XIST-Promoter Demethylation as Tissue Biomarker for Testicular Germ Cell Tumors and Spermatogenesis Quality




 and
and 
Abstract
:1. Introduction
2. Results
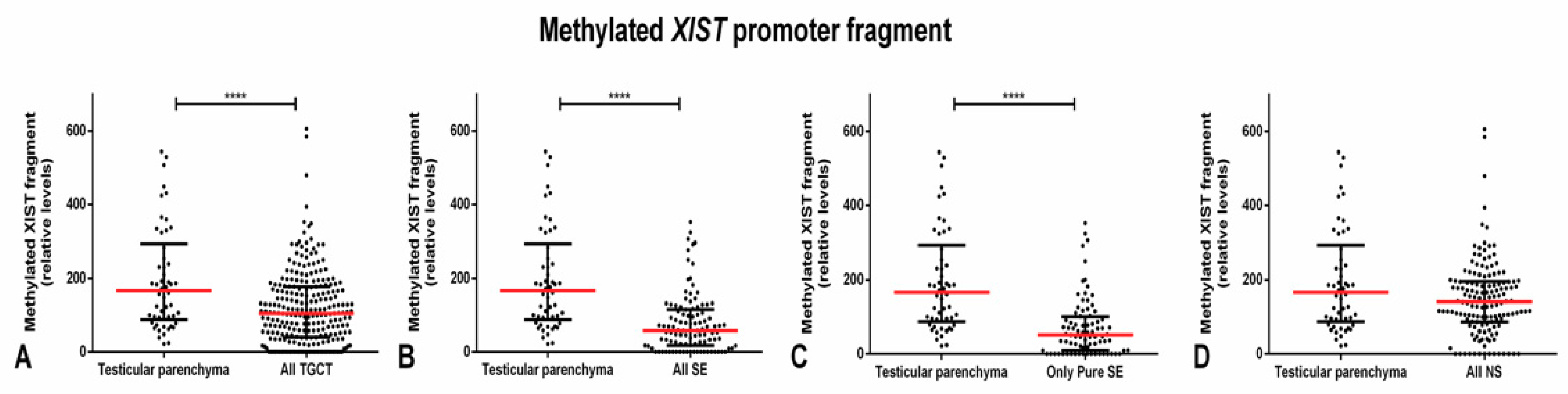
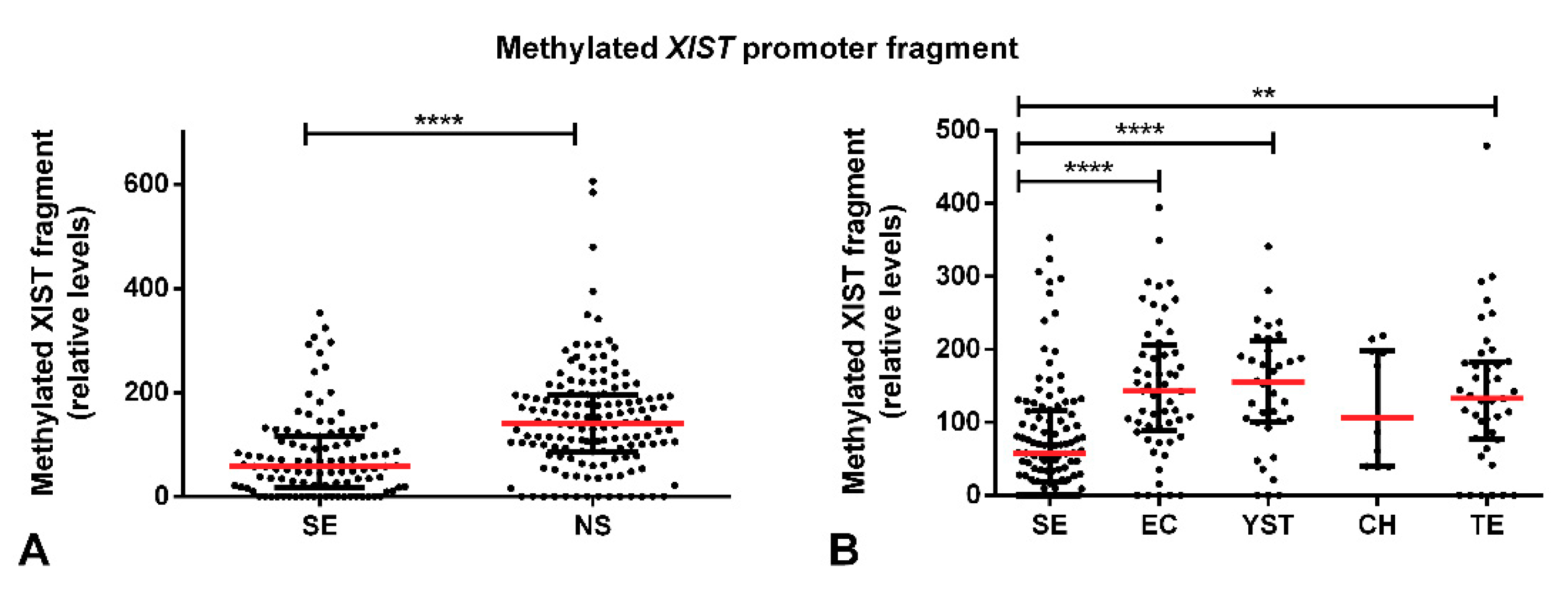
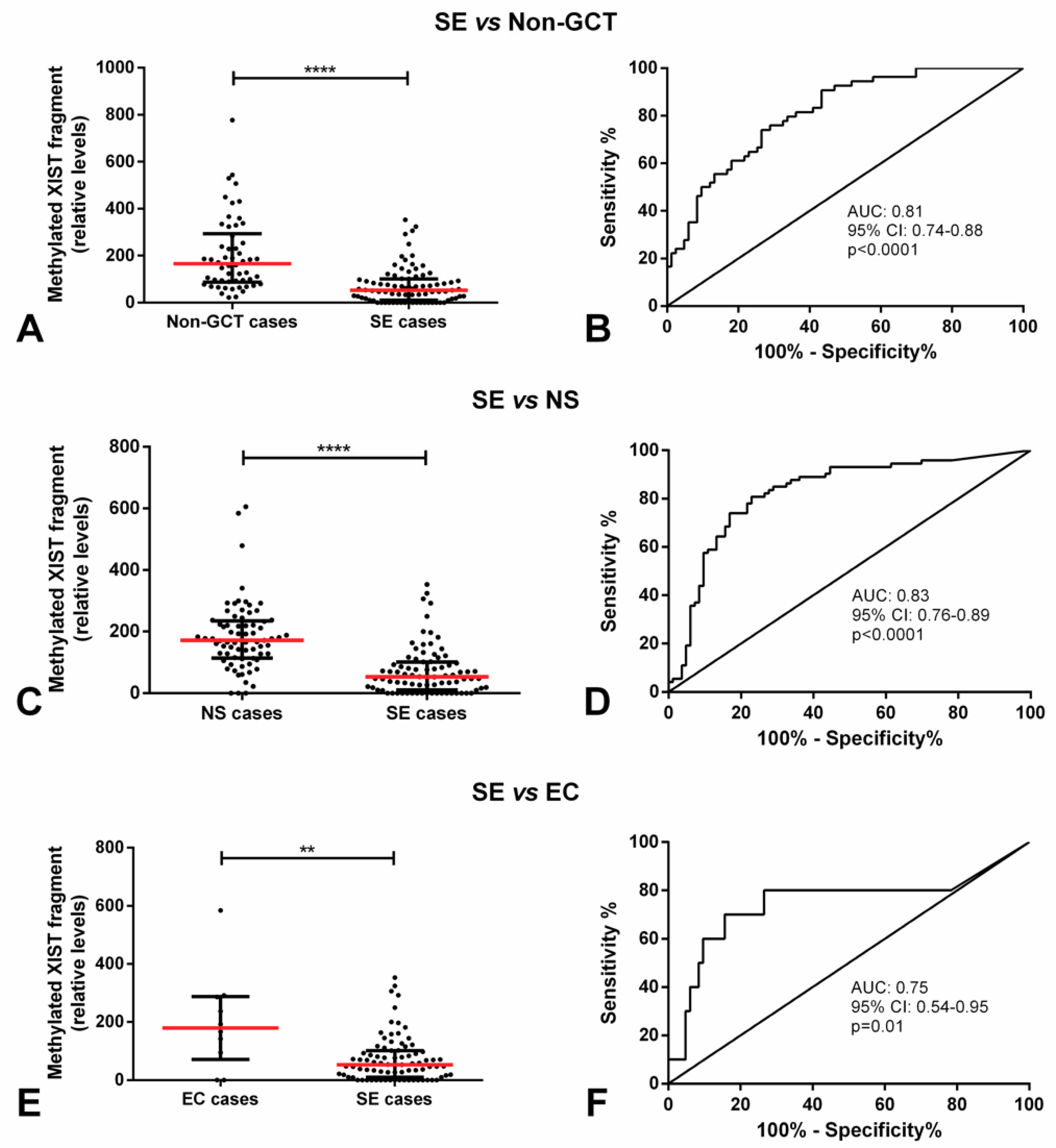
2.1. Methylated XIST Promoter in TGCTs
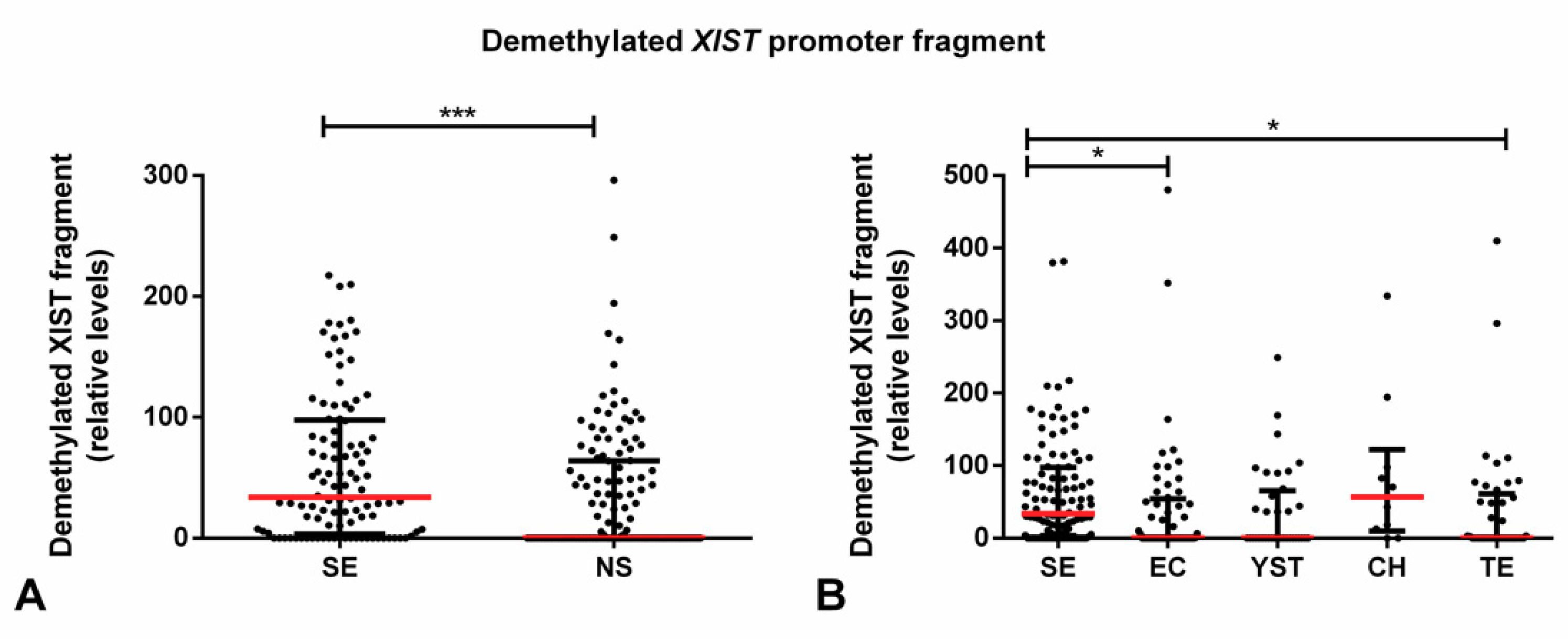
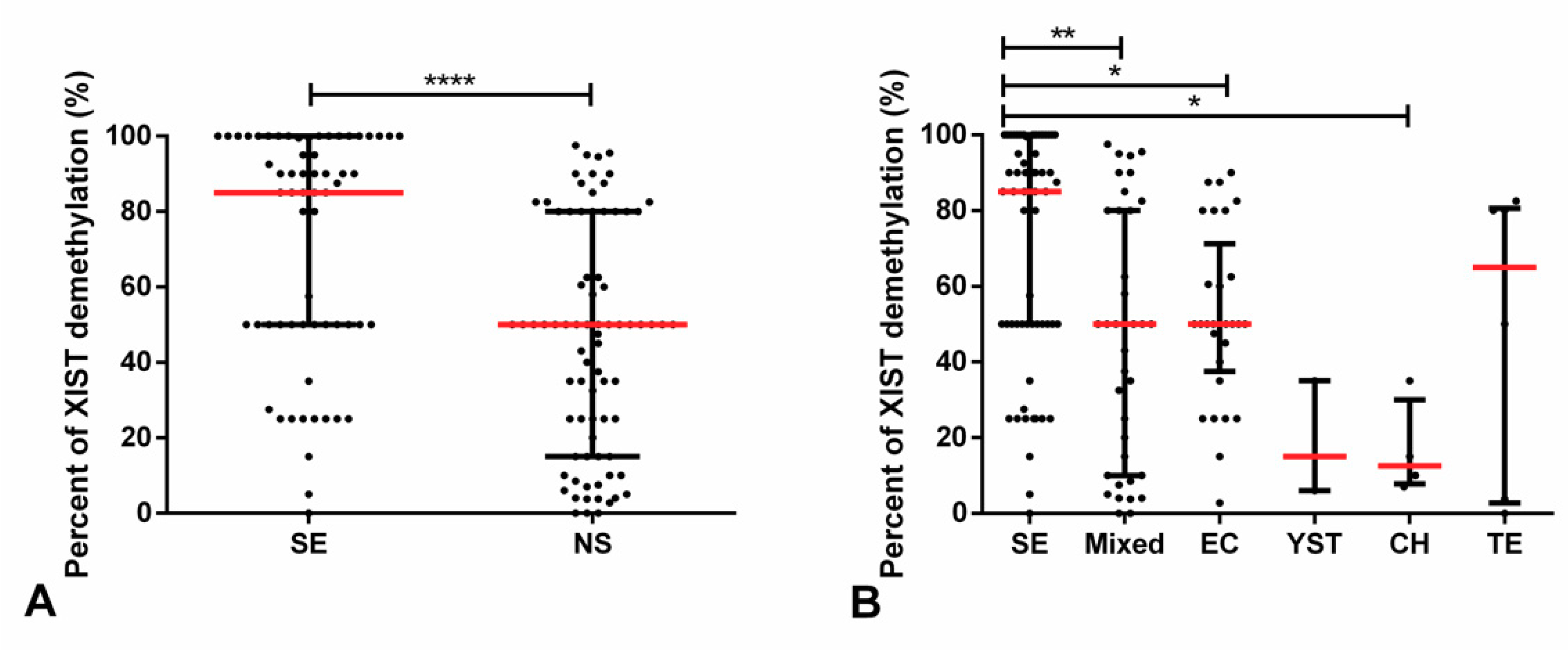
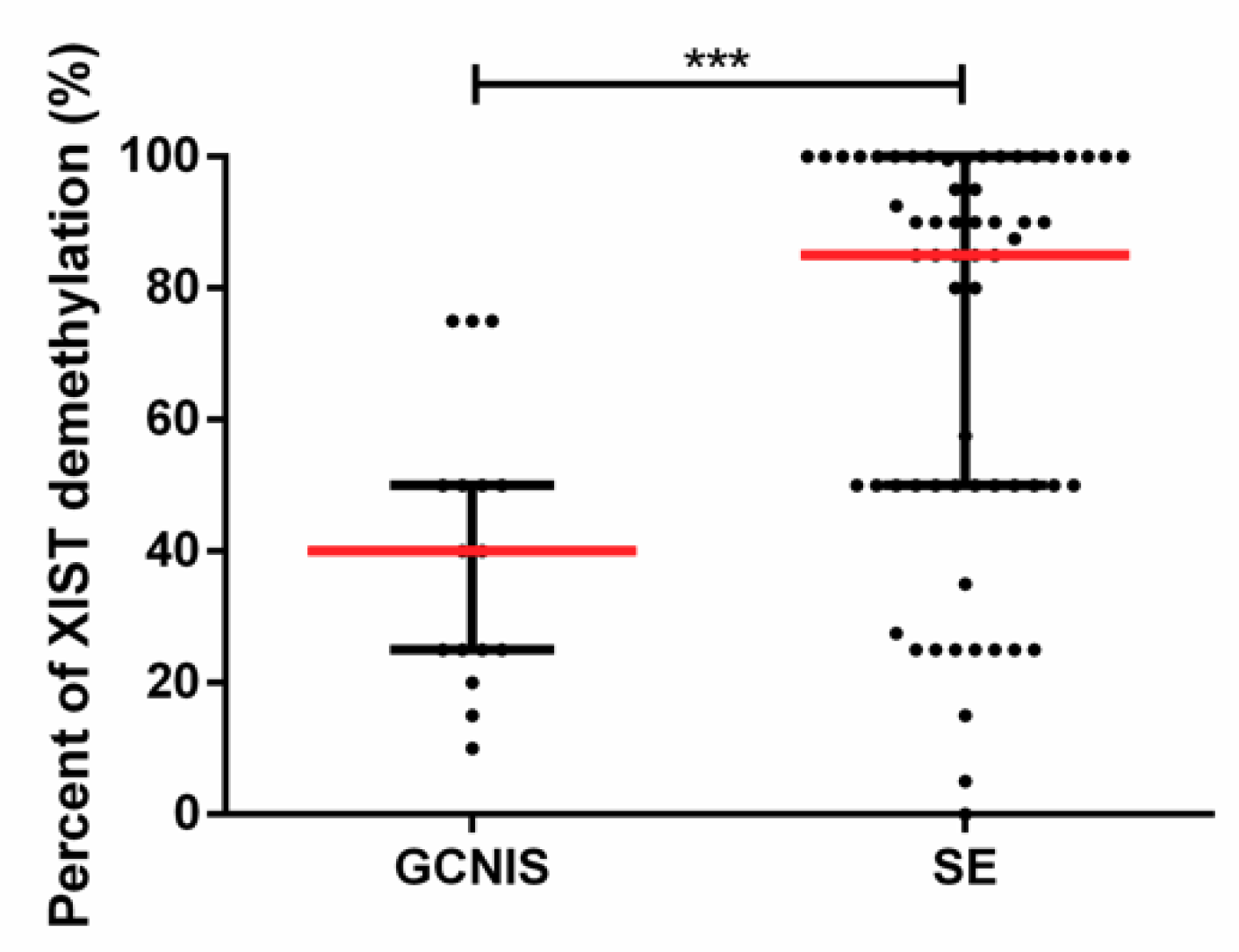
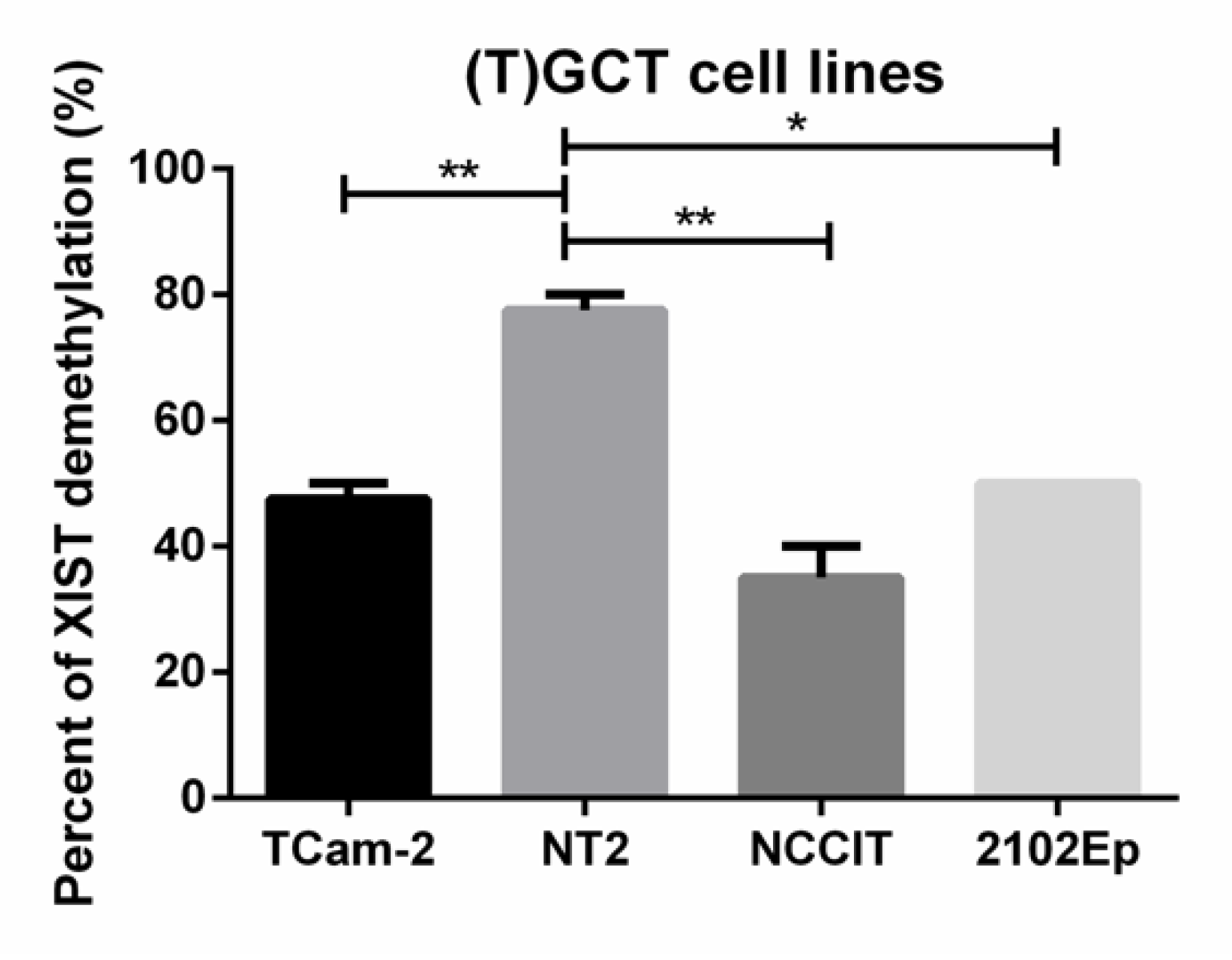
2.2. Demethylated XIST Promoter in TGCTs, GCNIS, and in (T)GCT Cell Lines
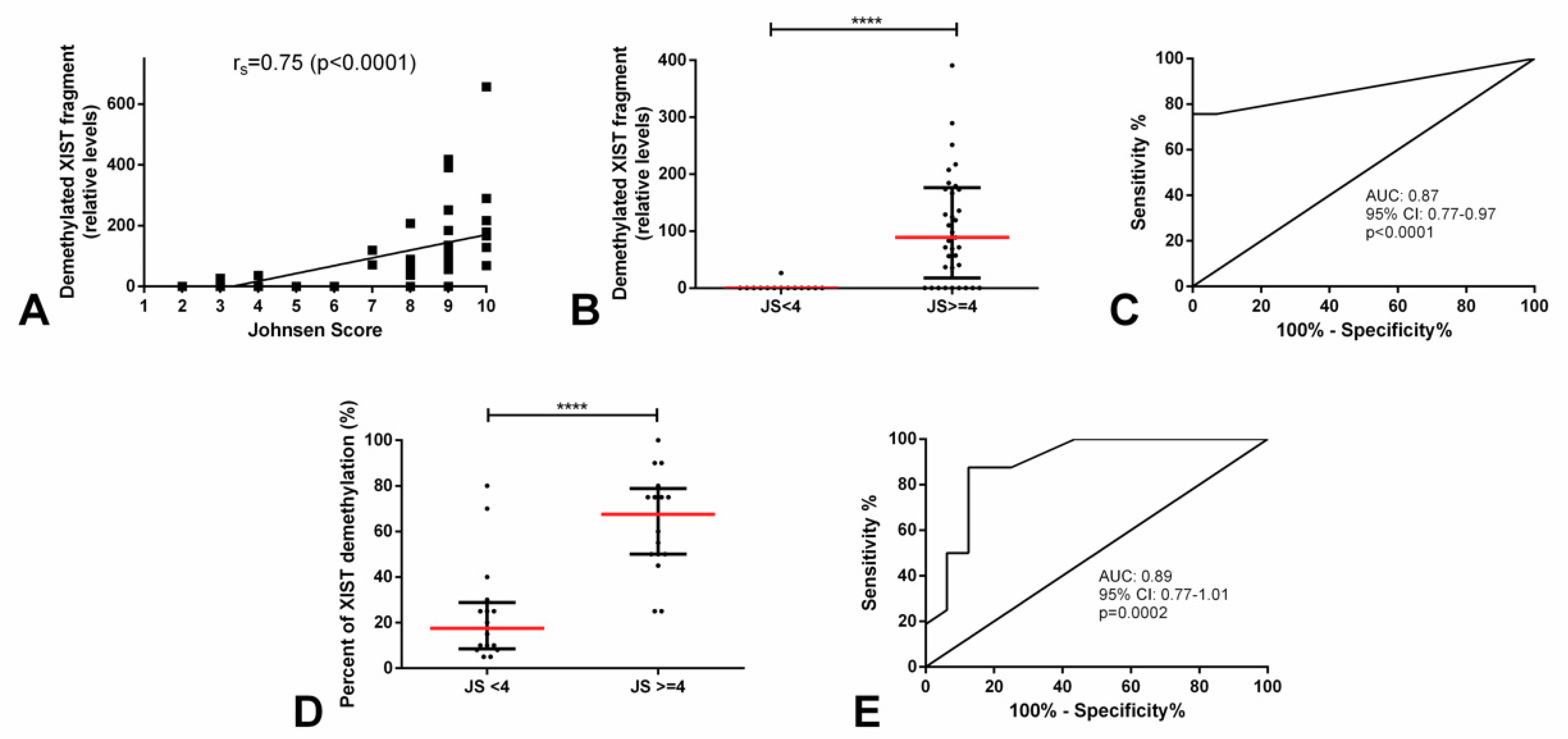
2.3. Demethylated XIST Promoter in Testicular Parenchyma
3. Discussion
4. Materials and Methods
4.1. Discovery Cohort
4.1.1. Testicular Germ Cell Tumor and Testicular Parenchyma Samples Selection
4.1.2. DNA Extraction and Bisulfite Treatment
4.1.3. Methylation Analyses
4.2. Validation Cohort
4.2.1. Testicular Germ Cell Tumor and Testicular Parenchyma Samples Selection
4.2.2. Cell Lines
4.2.3. DNA Extraction and Bisulfite Treatment
4.2.4. Methylation Analysis (High-Resolution Melting)
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lobo, J.; Gillis, A.J.M.; Jeronimo, C.; Henrique, R.; Looijenga, L.H.J. Human Germ Cell Tumors are Developmental Cancers: Impact of Epigenetics on Pathobiology and Clinic. Int. J. Mol. Sci. 2019, 20, 258. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.L.; Lobo, J.; Jeronimo, C.; Henrique, R. The epigenetics of testicular germ cell tumors: Looking for novel disease biomarkers. Epigenomics 2017, 9, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Rijlaarsdam, M.A.; Looijenga, L.H. An oncofetal and developmental perspective on testicular germ cell cancer. Semin. Cancer Biol. 2014, 29, 59–74. [Google Scholar] [CrossRef] [PubMed]
- Oosterhuis, J.W.; Looijenga, L.H. Testicular germ-cell tumours in a broader perspective. Nat. Rev. Cancer 2005, 5, 210–222. [Google Scholar] [CrossRef]
- Oosterhuis, J.W.; Looijenga, L.H.J. Human germ cell tumours from a developmental perspective. Nat. Rev. Cancer 2019, 19, 522–537. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Albers, P.; Berney, D.M.; Feldman, D.R.; Daugaard, G.; Gilligan, T.; Looijenga, L.H.J. Testicular cancer. Nat. Rev. Dis. Primers 2018, 4, 29. [Google Scholar] [CrossRef] [PubMed]
- Moch, H.; Ulbright, T.; Humphrey, P.; Reuter, V. WHO Classification of Tumours of the Urinary System and Male Genital Organs, 4th ed; IARC: Lyon, France, 2016. [Google Scholar]
- Looijenga, L.H.; Stoop, H.; Biermann, K. Testicular cancer: Biology and biomarkers. Virchows Arch. 2014, 464, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Lyon, M. The gift of observation: An interview with Mary Lyon. Interview by Jane Gitschier. PLoS Genet. 2010, 6, e1000813. [Google Scholar] [CrossRef]
- Gendrel, A.V.; Heard, E. Noncoding RNAs and epigenetic mechanisms during X-chromosome inactivation. Annu. Rev. Cell Dev. Biol. 2014, 30, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Chaligne, R.; Heard, E. X-chromosome inactivation in development and cancer. FEBS Lett. 2014, 588, 2514–2522. [Google Scholar] [CrossRef] [PubMed]
- Looijenga, L.H.; Gillis, A.J.; van Gurp, R.J.; Verkerk, A.J.; Oosterhuis, J.W. X inactivation in human testicular tumors. XIST expression and androgen receptor methylation status. Am. J. Pathol. 1997, 151, 581–590. [Google Scholar] [PubMed]
- Kawakami, T.; Okamoto, K.; Ogawa, O.; Okada, Y. XIST unmethylated DNA fragments in male-derived plasma as a tumour marker for testicular cancer. Lancet 2004, 363, 40–42. [Google Scholar] [CrossRef]
- Ushida, H.; Kawakami, T.; Minami, K.; Chano, T.; Okabe, H.; Okada, Y.; Okamoto, K. Methylation profile of DNA repetitive elements in human testicular germ cell tumor. Mol. Carcinog. 2012, 51, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Looijenga, L.H.; Oosterhuis, J.W. Clinical value of the X chromosome in testicular germ-cell tumours. Lancet 2004, 363, 6–8. [Google Scholar] [CrossRef]
- Ostrowski, K.A.; Walsh, T.J. Infertility with Testicular Cancer. Urol. Clin. N. Am. 2015, 42, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Richler, C.; Soreq, H.; Wahrman, J. X inactivation in mammalian testis is correlated with inactive X-specific transcription. Nat. Genet. 1992, 2, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Salido, E.C.; Yen, P.H.; Mohandas, T.K.; Shapiro, L.J. Expression of the X-inactivation-associated gene XIST during spermatogenesis. Nat. Genet. 1992, 2, 196–199. [Google Scholar] [CrossRef]
- Yan, W.; McCarrey, J.R. Sex chromosome inactivation in the male. Epigenetics 2009, 4, 452–456. [Google Scholar] [CrossRef] [Green Version]
- Johnsen, S.G. Testicular biopsy score count—A method for registration of spermatogenesis in human testes: Normal values and results in 335 hypogonadal males. Hormones 1970, 1, 2–25. [Google Scholar] [CrossRef]
- Tang, W.H.; Zhou, S.J.; Song, S.D.; He, H.Y.; Wu, H.; Zhang, Z.; Yang, Y.Z.; Zhang, H.L.; Mao, J.M.; Liu, D.F.; et al. A clinical trial on the consistency of bilateral testicular tissue histopathology and Johnsen score: Single side or bilateral side biopsy? Oncotarget 2018, 9, 23848–23859. [Google Scholar] [CrossRef]
- Esteves, S.C.; Miyaoka, R.; Orosz, J.E.; Agarwal, A. An update on sperm retrieval techniques for azoospermic males. Clinics 2013, 68 (Suppl. 1), 99–110. [Google Scholar] [CrossRef]
- Killian, J.K.; Dorssers, L.C.; Trabert, B.; Gillis, A.J.; Cook, M.B.; Wang, Y.; Waterfall, J.J.; Stevenson, H.; Smith, W.I., Jr.; Noyes, N.; et al. Imprints and DPPA3 are bypassed during pluripotency- and differentiation-coupled methylation reprogramming in testicular germ cell tumors. Genome Res. 2016, 26, 1490–1504. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Shih, J.; Hollern, D.P.; Wang, L.; Bowlby, R.; Tickoo, S.K.; Thorsson, V.; Mungall, A.J.; Newton, Y.; Hegde, A.M.; et al. Integrated Molecular Characterization of Testicular Germ Cell Tumors. Cell Rep. 2018, 23, 3392–3406. [Google Scholar] [CrossRef] [PubMed]
- Lobo, J.; Costa, A.L.; Vilela-Salgueiro, B.; Rodrigues, A.; Guimaraes, R.; Cantante, M.; Lopes, P.; Antunes, L.; Jeronimo, C.; Henrique, R. Testicular germ cell tumors: Revisiting a series in light of the new WHO classification and AJCC staging systems, focusing on challenges for pathologists. Hum. Pathol. 2018, 82, 113–124. [Google Scholar] [CrossRef]
- Looijenga, L.H.; Gillis, A.J.; Verkerk, A.J.; van Putten, W.L.; Oosterhuis, J.W. Heterogeneous X inactivation in trophoblastic cells of human full-term female placentas. Am. J. Hum. Genet. 1999, 64, 1445–1452. [Google Scholar] [CrossRef] [PubMed]
- Willard, H.F. X chromosome inactivation, XIST, and pursuit of the X-inactivation center. Cell 1996, 86, 5–7. [Google Scholar] [CrossRef]
- Norris, D.P.; Patel, D.; Kay, G.F.; Penny, G.D.; Brockdorff, N.; Sheardown, S.A.; Rastan, S. Evidence that random and imprinted Xist expression is controlled by preemptive methylation. Cell 1994, 77, 41–51. [Google Scholar] [CrossRef]
- Kawakami, T.; Okamoto, K.; Sugihara, H.; Hattori, T.; Reeve, A.E.; Ogawa, O.; Okada, Y. The roles of supernumerical X chromosomes and XIST expression in testicular germ cell tumors. J. Urol. 2003, 169, 1546–1552. [Google Scholar] [CrossRef]
- McCarrey, J.R.; Watson, C.; Atencio, J.; Ostermeier, G.C.; Marahrens, Y.; Jaenisch, R.; Krawetz, S.A. X-chromosome inactivation during spermatogenesis is regulated by an Xist/Tsix-independent mechanism in the mouse. Genesis 2002, 34, 257–266. [Google Scholar] [CrossRef]
- Turner, J.M.; Mahadevaiah, S.K.; Elliott, D.J.; Garchon, H.J.; Pehrson, J.R.; Jaenisch, R.; Burgoyne, P.S. Meiotic sex chromosome inactivation in male mice with targeted disruptions of Xist. J. Cell Sci. 2002, 115, 4097–4105. [Google Scholar] [CrossRef]
- Amin, M.B.; Edge, S.; Greene, F.; Byrd, D.R.; Brookland, R.K.; Washington, M.K.; Gershenwald, J.E.; Compton, C.C.; Hess, K.R.; Sullivan, D.C.; et al. AJCC Cancer Staging Manual, 8th ed; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- International Germ Cell Cancer Collaborative Group. International Germ Cell Consensus Classification: A prognostic factor-based staging system for metastatic germ cell cancers. J. Clin. Oncol. 1997, 15, 594–603. [Google Scholar] [CrossRef] [PubMed]
- Fankhauser, C.D.; Gerke, T.A.; Roth, L.; Sander, S.; Grossmann, N.C.; Kranzbuhler, B.; Eberli, D.; Sulser, T.; Beyer, J.; Hermanns, T. Pre-orchiectomy tumor marker levels should not be used for International Germ Cell Consensus Classification (IGCCCG) risk group assignment. J. Cancer Res. Clin. Oncol. 2019, 145, 781–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobo, J.; Costa, A.L.; Cantante, M.; Guimaraes, R.; Lopes, P.; Antunes, L.; Braga, I.; Oliveira, J.; Pelizzola, M.; Henrique, R.; et al. m(6)A RNA modification and its writer/reader VIRMA/YTHDF3 in testicular germ cell tumors: A role in seminoma phenotype maintenance. J. Transl. Med. 2019, 17, 79. [Google Scholar] [CrossRef] [PubMed]
- Vilela-Salgueiro, B.; Barros-Silva, D.; Lobo, J.; Costa, A.L.; Guimaraes, R.; Cantante, M.; Lopes, P.; Braga, I.; Oliveira, J.; Henrique, R.; et al. Germ cell tumour subtypes display differential expression of microRNA371a-3p. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2018, 373, 20170338. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.L.; Moreira-Barbosa, C.; Lobo, J.; Vilela-Salgueiro, B.; Cantante, M.; Guimaraes, R.; Lopes, P.; Braga, I.; Oliveira, J.; Antunes, L.; et al. DNA methylation profiling as a tool for testicular germ cell tumors subtyping. Epigenomics 2018, 10, 1511–1523. [Google Scholar] [CrossRef] [PubMed]
- Eads, C.A.; Lord, R.V.; Wickramasinghe, K.; Long, T.I.; Kurumboor, S.K.; Bernstein, L.; Peters, J.H.; DeMeester, S.R.; DeMeester, T.R.; Skinner, K.A.; et al. Epigenetic patterns in the progression of esophageal adenocarcinoma. Cancer Res. 2001, 61, 3410–3418. [Google Scholar] [PubMed]
- Gillis, A.J.; Stoop, H.; Biermann, K.; van Gurp, R.J.; Swartzman, E.; Cribbes, S.; Ferlinz, A.; Shannon, M.; Oosterhuis, J.W.; Looijenga, L.H. Expression and interdependencies of pluripotency factors LIN28, OCT3/4, NANOG and SOX2 in human testicular germ cells and tumours of the testis. Int. J. Androl. 2011, 34, e160-174. [Google Scholar] [CrossRef]
- Hussmann, D.; Hansen, L.L. Methylation-Sensitive High Resolution Melting (MS-HRM). Methods Mol. Biol. 2018, 1708, 551–571. [Google Scholar] [CrossRef]
- Wojdacz, T.K.; Dobrovic, A. Methylation-sensitive high resolution melting (MS-HRM): A new approach for sensitive and high-throughput assessment of methylation. Nucleic Acids Res. 2007, 35, e41. [Google Scholar] [CrossRef]
- Youden, W.J. Index for rating diagnostic tests. Cancer 1950, 3, 32–35. [Google Scholar] [CrossRef]
- Schisterman, E.F.; Perkins, N.J.; Liu, A.; Bondell, H. Optimal cut-point and its corresponding Youden Index to discriminate individuals using pooled blood samples. Epidemiology 2005, 16, 73–81. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Context. | AUC (95% CI) | Sensitivity (%) | Specificity (%) | PPV (%) | NPV (%) | Accuracy (%) |
|---|---|---|---|---|---|---|
| SE vs. non-GCT cases | 0.81 (0.74–0.88) | 73.5 | 74.1 | 81.3 | 64.5 | 73.7 |
| SE vs. NS cases | 0.83 (0.76–0.89) | 77.1 | 80.8 | 82.1 | 75.6 | 78.8 |
| SE vs. EC cases | 0.75 (0.54–0.95) | 84.3 | 70.0 | 95.9 | 35.0 | 82.8 |
| Context. | AUC (95% CI) | Sensitivity (%) | Specificity (%) | PPV (%) | NPV (%) | Accuracy (%) |
|---|---|---|---|---|---|---|
| JS ≥ 4 vs. JS < 4 | 0.87 (0.77-0.97) | 75.7 | 100 | 100 | 60.9 | 82.4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lobo, J.; Nunes, S.P.; Gillis, A.J.M.; Barros-Silva, D.; Miranda-Gonçalves, V.; Berg, A.v.d.; Cantante, M.; Guimarães, R.; Henrique, R.; Jerónimo, C.; et al. XIST-Promoter Demethylation as Tissue Biomarker for Testicular Germ Cell Tumors and Spermatogenesis Quality. Cancers 2019, 11, 1385. https://doi.org/10.3390/cancers11091385
Lobo J, Nunes SP, Gillis AJM, Barros-Silva D, Miranda-Gonçalves V, Berg Avd, Cantante M, Guimarães R, Henrique R, Jerónimo C, et al. XIST-Promoter Demethylation as Tissue Biomarker for Testicular Germ Cell Tumors and Spermatogenesis Quality. Cancers. 2019; 11(9):1385. https://doi.org/10.3390/cancers11091385
Chicago/Turabian StyleLobo, João, Sandra P. Nunes, Ad J. M. Gillis, Daniela Barros-Silva, Vera Miranda-Gonçalves, Annette van den Berg, Mariana Cantante, Rita Guimarães, Rui Henrique, Carmen Jerónimo, and et al. 2019. "XIST-Promoter Demethylation as Tissue Biomarker for Testicular Germ Cell Tumors and Spermatogenesis Quality" Cancers 11, no. 9: 1385. https://doi.org/10.3390/cancers11091385
APA StyleLobo, J., Nunes, S. P., Gillis, A. J. M., Barros-Silva, D., Miranda-Gonçalves, V., Berg, A. v. d., Cantante, M., Guimarães, R., Henrique, R., Jerónimo, C., & Looijenga, L. H. J. (2019). XIST-Promoter Demethylation as Tissue Biomarker for Testicular Germ Cell Tumors and Spermatogenesis Quality. Cancers, 11(9), 1385. https://doi.org/10.3390/cancers11091385