Cytokine-Induced Guanylate Binding Protein 1 (GBP1) Release from Human Ovarian Cancer Cells

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Proteomic Analysis of IL-27- or IFN-γ-Stimulated EOC Cells Shows an Overlapping Pattern of Secreted Proteins, among Which GBP1 is Highly Expressed
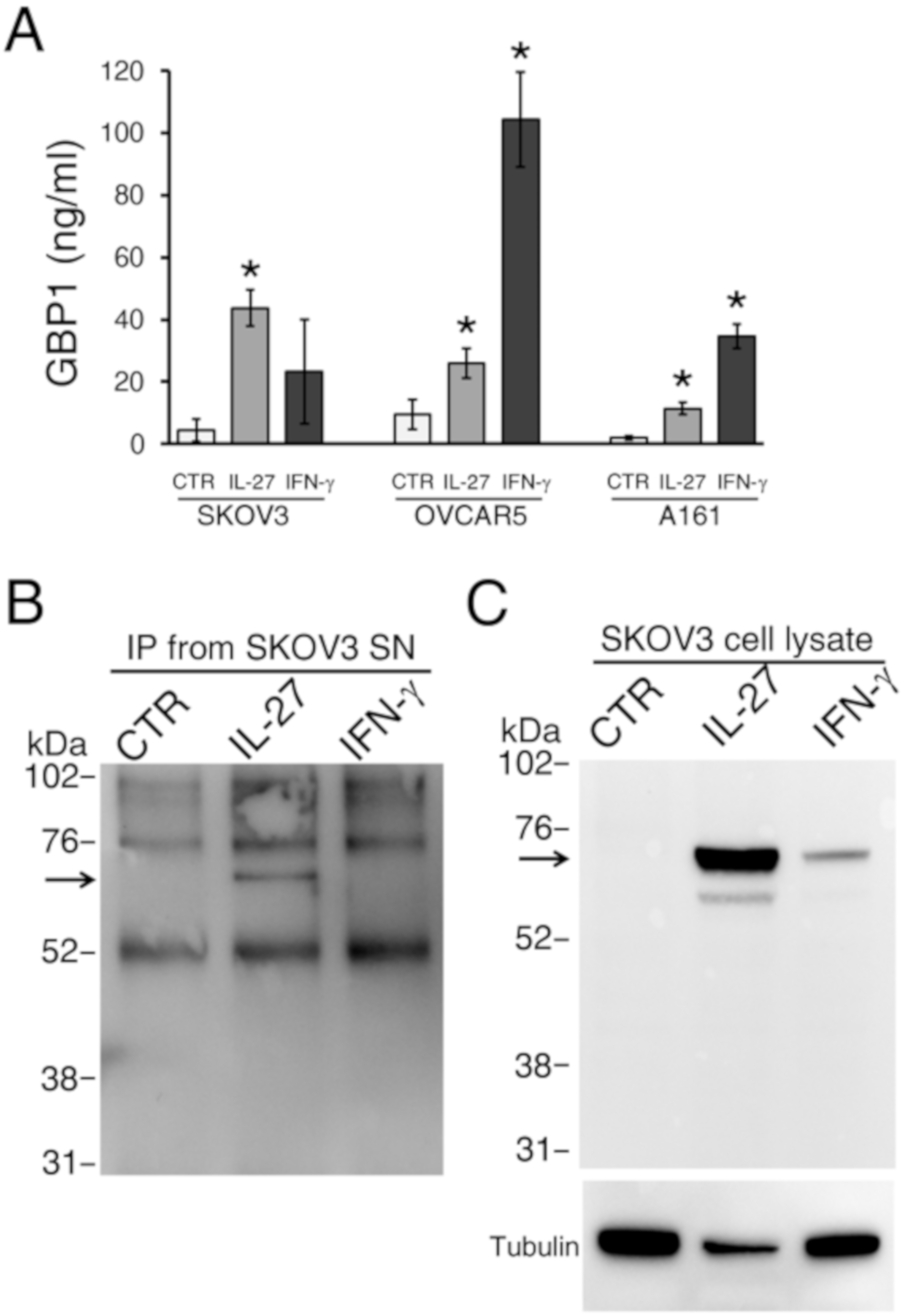
2.2. GBP1 is Released Predominantly As A Full-Length Molecule by Cytokine-Stimulated EOC Cells
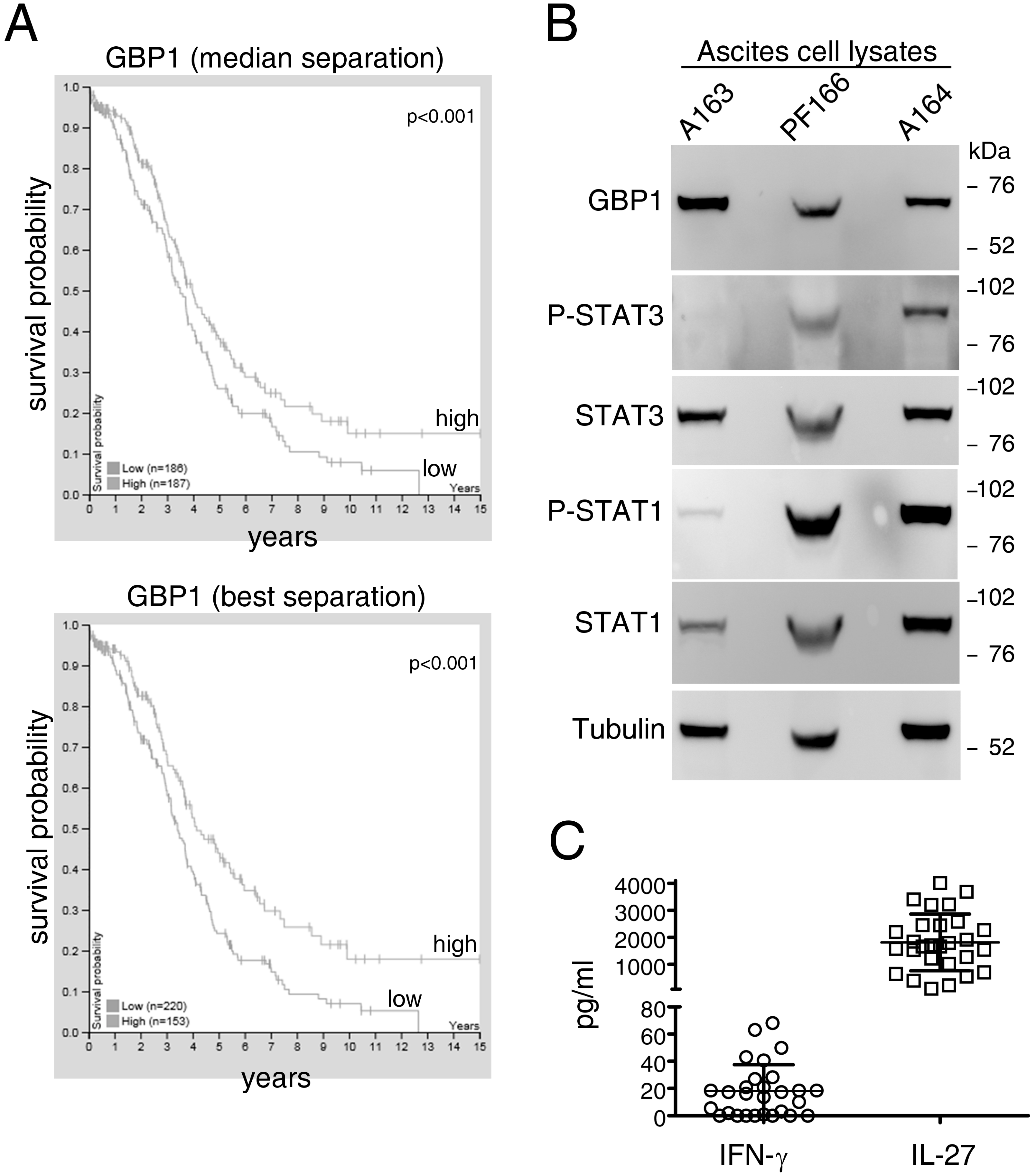
2.3. GBP1 is Expressed by EOC Tumors In Vivo and Accumulates in the Ascites
2.4. The Full-Length, Soluble Molecular Form of GBP1 Accumulates in the Liquid Phase of EOC Ascites
2.5. Effects of GBP1 Gene Transfer on EOC Cells In Vitro
3. Discussion
4. Materials and Methods
4.1. Ethics Approval and Consent to Participate
4.2. Cells and Cell Treatments
4.3. Sample Preparation and Mass Spectrometer Setup
4.4. Western Blot
4.5. GBP1 Immunoprecipitation and GTP-Agarose Pull-Down
4.6. Microvesicles Isolation from Ascites
4.7. ELISA and Multiplex Immunoassay
4.8. Cell Transfection and MTT Assay
4.9. Statistical Analysis and Online Data Retrieval
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Qu, X.; Tang, Y.; Hua, S. Immunological Approaches Towards Cancer and Inflammation: A Cross Talk. Front. Immunol. 2018, 9, 563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marelli, G.; Sica, A.; Vannucci, L.; Allavena, P. Inflammation as target in cancer therapy. Curr. Opin. Pharmacol. 2017, 35, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Castro, F.; Cardoso, A.P.; Gonçalves, R.M.; Serre, K.; Oliveira, M.J. Interferon-Gamma at the Crossroads of Tumor Immune Surveillance or Evasion. Front. Immunol. 2018, 9, 847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rožman, P.; Švajger, U. The tolerogenic role of IFN-γ. Cytokine Growth Factor Rev. 2018, 41, 40–53. [Google Scholar] [CrossRef]
- Spranger, S.; Spaapen, R.M.; Zha, Y.; Williams, J.; Meng, Y.; Ha, T.T.; Gajewski, T.F. Up-regulation of PD-L1, IDO, and T(regs) in the melanoma tumor microenvironment is driven by CD8(+) T cells. Sci. Transl. Med. 2013, 5, 200ra116. [Google Scholar] [CrossRef] [Green Version]
- Prendergast, G.C.; Smith, C.; Thomas, S.; Mandik-Nayak, L.; Laury-Kleintop, L.; Metz, R.; Muller, A.J. Indoleamine 2,3-dioxygenase pathways of pathogenic inflammation and immune escape in cancer. Cancer Immunol. Immunother. CII 2014, 63, 721–735. [Google Scholar] [CrossRef]
- Pflanz, S.; Timans, J.C.; Cheung, J.; Rosales, R.; Kanzler, H.; Gilbert, J.; Hibbert, L.; Churakova, T.; Travis, M.; Vaisberg, E.; et al. IL-27, a heterodimeric cytokine composed of EBI3 and p28 protein, induces proliferation of naive CD4+ T cells. Immunity 2002, 16, 779–790. [Google Scholar] [CrossRef] [Green Version]
- Trinchieri, G.; Pflanz, S.; Kastelein, R.A. The IL-12 family of heterodimeric cytokines: New players in the regulation of T cell responses. Immunity 2003, 19, 641–644. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, H.; Hunter, C.A. The immunobiology of interleukin-27. Annu. Rev. Immunol. 2015, 33, 417–443. [Google Scholar] [CrossRef]
- Carbotti, G.; Barisione, G.; Airoldi, I.; Mezzanzanica, D.; Bagnoli, M.; Ferrero, S.; Petretto, A.; Fabbi, M.; Ferrini, S. IL-27 induces the expression of IDO and PD-L1 in human cancer cells. Oncotarget 2015, 6, 43267–43280. [Google Scholar] [CrossRef] [Green Version]
- Carbotti, G.; Barisione, G.; Orengo, A.M.; Brizzolara, A.; Airoldi, I.; Bagnoli, M.; Pinciroli, P.; Mezzanzanica, D.; Centurioni, M.G.; Fabbi, M.; et al. The IL-18 antagonist IL-18-binding protein is produced in the human ovarian cancer microenvironment. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2013, 19, 4611–4620. [Google Scholar] [CrossRef] [Green Version]
- Petretto, A.; Carbotti, G.; Inglese, E.; Lavarello, C.; Pistillo, M.P.; Rigo, V.; Croce, M.; Longo, L.; Martini, S.; Vacca, P.; et al. Proteomic analysis uncovers common effects of IFN-γ and IL-27 on the HLA class I antigen presentation machinery in human cancer cells. Oncotarget 2016, 7, 72518–72536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbotti, G.; Nikpoor, A.R.; Vacca, P.; Gangemi, R.; Giordano, C.; Campelli, F.; Ferrini, S.; Fabbi, M. IL-27 mediates HLA class I up-regulation, which can be inhibited by the IL-6 pathway, in HLA-deficient Small Cell Lung Cancer cells. J. Exp. Clin. Cancer Res. CR 2017, 36, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocco, C.; Giuliani, N.; Di Carlo, E.; Ognio, E.; Storti, P.; Abeltino, M.; Sorrentino, C.; Ponzoni, M.; Ribatti, D.; Airoldi, I. Interleukin-27 acts as multifunctional antitumor agent in multiple myeloma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2010, 16, 4188–4197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kachroo, P.; Lee, M.-H.; Zhang, L.; Baratelli, F.; Lee, G.; Srivastava, M.K.; Wang, G.; Walser, T.C.; Krysan, K.; Sharma, S.; et al. IL-27 inhibits epithelial-mesenchymal transition and angiogenic factor production in a STAT1-dominant pathway in human non-small cell lung cancer. J. Exp. Clin. Cancer Res. CR 2013, 32, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabbi, M.; Carbotti, G.; Ferrini, S. Dual Roles of IL-27 in Cancer Biology and Immunotherapy. Mediators Inflamm. 2017, 2017, 3958069. [Google Scholar] [CrossRef] [Green Version]
- Mandai, M.; Hamanishi, J.; Abiko, K.; Matsumura, N.; Baba, T.; Konishi, I. Dual Faces of IFNγ in Cancer Progression: A Role of PD-L1 Induction in the Determination of Pro- and Antitumor Immunity. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2016, 22, 2329–2334. [Google Scholar] [CrossRef] [Green Version]
- Hibbert, L.; Pflanz, S.; De Waal Malefyt, R.; Kastelein, R.A. IL-27 and IFN-alpha signal via Stat1 and Stat3 and induce T-Bet and IL-12Rbeta2 in naive T cells. J. Interferon Cytokine Res. Off. J. Int. Soc. Interferon Cytokine Res. 2003, 23, 513–522. [Google Scholar] [CrossRef]
- Anderson, S.L.; Carton, J.M.; Lou, J.; Xing, L.; Rubin, B.Y. Interferon-induced guanylate binding protein-1 (GBP-1) mediates an antiviral effect against vesicular stomatitis virus and encephalomyocarditis virus. Virology 1999, 256, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Meunier, E.; Broz, P. Interferon-inducible GTPases in cell autonomous and innate immunity. Cell. Microbiol. 2016, 18, 168–180. [Google Scholar] [CrossRef] [Green Version]
- Guenzi, E.; Töpolt, K.; Lubeseder-Martellato, C.; Jörg, A.; Naschberger, E.; Benelli, R.; Albini, A.; Stürzl, M. The guanylate binding protein-1 GTPase controls the invasive and angiogenic capability of endothelial cells through inhibition of MMP-1 expression. EMBO J. 2003, 22, 3772–3782. [Google Scholar] [CrossRef] [PubMed]
- Naschberger, E.; Bauer, M.; Stürzl, M. Human guanylate binding protein-1 (hGBP-1) characterizes and establishes a non-angiogenic endothelial cell activation phenotype in inflammatory diseases. Adv. Enzyme Regul. 2005, 45, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Weinländer, K.; Naschberger, E.; Lehmann, M.H.; Tripal, P.; Paster, W.; Stockinger, H.; Hohenadl, C.; Stürzl, M. Guanylate binding protein-1 inhibits spreading and migration of endothelial cells through induction of integrin α 4 expression. FASEB J. 2008, 22, 4168–4178. [Google Scholar] [CrossRef] [PubMed]
- Naschberger, E.; Croner, R.S.; Merkel, S.; Dimmler, A.; Tripal, P.; Amann, K.U.; Kremmer, E.; Brueckl, W.M.; Papadopoulos, T.; Hohenadl, C.; et al. Angiostatic immune reaction in colorectal carcinoma: Impact on survival and perspectives for antiangiogenic therapy. Int. J. Cancer 2008, 123, 2120–2129. [Google Scholar] [CrossRef] [PubMed]
- Lipnik, K.; Naschberger, E.; Gonin-Laurent, N.; Kodajova, P.; Petznek, H.; Rungaldier, S.; Astigiano, S.; Ferrini, S.; Stürzl, M.; Hohenadl, C. Interferon gamma-induced human guanylate binding protein 1 inhibits mammary tumor growth in mice. Mol. Med. Camb. Mass 2010, 16, 177–187. [Google Scholar] [PubMed]
- Britzen-Laurent, N.; Lipnik, K.; Ocker, M.; Naschberger, E.; Schellerer, V.S.; Croner, R.S.; Vieth, M.; Waldner, M.; Steinberg, P.; Hohenadl, C.; et al. GBP-1 acts as a tumor suppressor in colorectal cancer cells. Carcinogenesis 2013, 34, 153–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naschberger, E.; Liebl, A.; Schellerer, V.S.; Schütz, M.; Britzen-Laurent, N.; Kölbel, P.; Schaal, U.; Haep, L.; Regensburger, D.; Wittmann, T.; et al. Matricellular protein SPARCL1 regulates tumor microenvironment-dependent endothelial cell heterogeneity in colorectal carcinoma. J. Clin. Investig. 2016, 126, 4187–4204. [Google Scholar] [CrossRef] [Green Version]
- Britzen-Laurent, N.; Herrmann, C.; Naschberger, E.; Croner, R.S.; Stürzl, M. Pathophysiological role of guanylate-binding proteins in gastrointestinal diseases. World J. Gastroenterol. 2016, 22, 6434–6443. [Google Scholar] [CrossRef]
- Muzny, D.M.; Bainbridge, M.N.; Chang, K.; Dinh, H.H.; Drummond, J.A.; Fowler, G.; Kovar, C.L.; Lewis, L.R.; Morgan, M.B.; Newsham, I.F.; et al. Cancer Genome Atlas Network Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330–337. [Google Scholar]
- Li, L.; Ma, G.; Jing, C.; Liu, Z. Guanylate-binding protein 1 (GBP1) promotes lymph node metastasis in human esophageal squamous cell carcinoma. Discov. Med. 2015, 20, 369–378. [Google Scholar]
- Lan, Q.; Wang, A.; Cheng, Y.; Mukasa, A.; Ma, J.; Hong, L.; Yu, S.; Sun, L.; Huang, Q.; Purow, B.; et al. Guanylate binding protein-1 mediates EGFRvIII and promotes glioblastoma growth in vivo but not in vitro. Oncotarget 2016, 7, 9680–9691. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.; Foster, R.; Brakora, K.A.; Yusuf, R.Z.; Seiden, M.V. GBP1 overexpression is associated with a paclitaxel resistance phenotype. Cancer Chemother. Pharmacol. 2006, 57, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, M.; Amanuma, T.; Kuwahara, Y.; Shimura, T.; Suzuki, M.; Mori, S.; Kumamoto, H.; Saito, Y.; Ohkubo, Y.; Duan, Z.; et al. Guanine nucleotide-binding protein 1 is one of the key molecules contributing to cancer cell radioresistance. Cancer Sci. 2014, 105, 1351–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreoli, M.; Persico, M.; Kumar, A.; Orteca, N.; Kumar, V.; Pepe, A.; Mahalingam, S.; Alegria, A.E.; Petrella, L.; Sevciunaite, L.; et al. Identification of the first inhibitor of the GBP1:PIM1 interaction. Implications for the development of a new class of anticancer agents against paclitaxel resistant cancer cells. J. Med. Chem. 2014, 57, 7916–7932. [Google Scholar] [CrossRef] [Green Version]
- Domcke, S.; Sinha, R.; Levine, D.A.; Sander, C.; Schultz, N. Evaluating cell lines as tumour models by comparison of genomic profiles. Nat. Commun. 2013, 4, 2126. [Google Scholar] [CrossRef]
- Naschberger, E.; Geißdörfer, W.; Bogdan, C.; Tripal, P.; Kremmer, E.; Stürzl, M.; Britzen-Laurent, N. Processing and secretion of guanylate binding protein-1 depend on inflammatory caspase activity. J. Cell. Mol. Med. 2017, 21, 1954–1966. [Google Scholar] [CrossRef]
- Guenzi, E.; Töpolt, K.; Cornali, E.; Lubeseder-Martellato, C.; Jörg, A.; Matzen, K.; Zietz, C.; Kremmer, E.; Nappi, F.; Schwemmle, M.; et al. The helical domain of GBP-1 mediates the inhibition of endothelial cell proliferation by inflammatory cytokines. EMBO J. 2001, 20, 5568–5577. [Google Scholar] [CrossRef] [Green Version]
- Uhlen, M.; Zhang, C.; Lee, S.; Sjöstedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A pathology atlas of the human cancer transcriptome. Science 2017, 357, eaan2507. [Google Scholar] [CrossRef] [Green Version]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Kim, J.; Gee, H.Y.; Lee, M.G. Unconventional protein secretion—New insights into the pathogenesis and therapeutic targets of human diseases. J. Cell Sci. 2018, 131, jcs213686. [Google Scholar] [CrossRef] [Green Version]
- Greening, D.W.; Ji, H.; Chen, M.; Robinson, B.W.S.; Dick, I.M.; Creaney, J.; Simpson, R.J. Secreted primary human malignant mesothelioma exosome signature reflects oncogenic cargo. Sci. Rep. 2016, 6, 32643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britzen-Laurent, N.; Bauer, M.; Berton, V.; Fischer, N.; Syguda, A.; Reipschläger, S.; Naschberger, E.; Herrmann, C.; Stürzl, M. Intracellular trafficking of guanylate-binding proteins is regulated by heterodimerization in a hierarchical manner. PLoS ONE 2010, 5, e14246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, W.; Shan, B.; Zhang, Y.; Ren, Y.; Liang, S.; Zhao, J.; Zhao, Z.; Wang, G.; Zhao, X.; Peng, D.; et al. Association between DNA damage repair gene somatic mutations and immune-related gene expression in ovarian cancer. Cancer Med. 2020, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumacher, T.N.; Schreiber, R.D. Neoantigens in cancer immunotherapy. Science 2015, 348, 69–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naschberger, E.; Lubeseder-Martellato, C.; Meyer, N.; Gessner, R.; Kremmer, E.; Gessner, A.; Stürzl, M. Human guanylate binding protein-1 is a secreted GTPase present in increased concentrations in the cerebrospinal fluid of patients with bacterial meningitis. Am. J. Pathol. 2006, 169, 1088–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.Y.; Gaggero, A.; Rubartelli, A.; Rosso, O.; Miotti, S.; Mezzanzanica, D.; Canevari, S.; Ferrini, S. Expression of interleukin-18 in human ovarian carcinoma and normal ovarian epithelium: Evidence for defective processing in tumor cells. Int. J. Cancer 2002, 98, 873–878. [Google Scholar] [CrossRef] [Green Version]
- Orengo, A.M.; Fabbi, M.; Miglietta, L.; Andreani, C.; Bruzzone, M.; Puppo, A.; Cristoforoni, P.; Centurioni, M.G.; Gualco, M.; Salvi, S.; et al. Interleukin (IL)-18, a biomarker of human ovarian carcinoma, is predominantly released as biologically inactive precursor. Int. J. Cancer 2011, 129, 1116–1125. [Google Scholar] [CrossRef]
- Feng, Q.; Li, P.; Salamanca, C.; Huntsman, D.; Leung, P.C.K.; Auersperg, N. Caspase-1alpha is down-regulated in human ovarian cancer cells and the overexpression of caspase-1alpha induces apoptosis. Cancer Res. 2005, 65, 8591–8596. [Google Scholar] [CrossRef] [Green Version]
- Jackson, E.K.; Cheng, D.; Jackson, T.C.; Verrier, J.D.; Gillespie, D.G. Extracellular guanosine regulates extracellular adenosine levels. Am. J. Physiol. Cell Physiol. 2013, 304, C406–C421. [Google Scholar] [CrossRef] [Green Version]
- Dubyak, G.R. Dueling nucleosides: Cross-regulation of extracellular adenosine by guanosine. Focus on “Extracellular guanosine regulates extracellular adenosine levels”. Am. J. Physiol. Cell Physiol. 2013, 304, C403–C405. [Google Scholar]
- Unterer, B.; Wiesmann, V.; Gunasekaran, M.; Sticht, H.; Tenkerian, C.; Behrens, J.; Leone, M.; Engel, F.B.; Britzen-Laurent, N.; Naschberger, E.; et al. IFN-γ-response mediator GBP-1 represses human cell proliferation by inhibiting the Hippo signaling transcription factor TEAD. Biochem. J. 2018, 475, 2955–2967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wadi, S.; Tipton, A.R.; Trendel, J.A.; Khuder, S.A.; Vestal, D.J. hGBP-1 Expression Predicts Shorter Progression-Free Survival in Ovarian Cancers, While Contributing to Paclitaxel Resistance. J. Cancer Ther. 2016, 7, 994–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wang, S.-S.; Zhou, L.; Yu, L.; Zhang, L.-M. A network-pathway based module identification for predicting the prognosis of ovarian cancer patients. J. Ovarian Res. 2016, 9, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulak, N.A.; Pichler, G.; Paron, I.; Nagaraj, N.; Mann, M. Minimal, encapsulated proteomic-sample processing applied to copy-number estimation in eukaryotic cells. Nat. Methods 2014, 11, 319–324. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell. Proteomics MCP 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [Green Version]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Lubeseder-Martellato, C.; Guenzi, E.; Jörg, A.; Töpolt, K.; Naschberger, E.; Kremmer, E.; Zietz, C.; Tschachler, E.; Hutzler, P.; Schwemmle, M.; et al. Guanylate-binding protein-1 expression is selectively induced by inflammatory cytokines and is an activation marker of endothelial cells during inflammatory diseases. Am. J. Pathol. 2002, 161, 1749–1759. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | SKOV3 Cell Lysate | SKOV3 Conditioned Media | ||
|---|---|---|---|---|
| IFN-γ | IL-27 | IFN-γ | IL-27 | |
| GBP1 | 8.50 a | 8.47 | 6.43 | 3.20 |
| GBP2 | 5.95 | 6.18 | NaN b | NaN |
| GBP4 | 1.59 | 2.53 | NaN | NaN |
| GBP5 | 5.73 | 6.78 | NaN | NaN |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carbotti, G.; Petretto, A.; Naschberger, E.; Stürzl, M.; Martini, S.; Mingari, M.C.; Filaci, G.; Ferrini, S.; Fabbi, M. Cytokine-Induced Guanylate Binding Protein 1 (GBP1) Release from Human Ovarian Cancer Cells. Cancers 2020, 12, 488. https://doi.org/10.3390/cancers12020488
Carbotti G, Petretto A, Naschberger E, Stürzl M, Martini S, Mingari MC, Filaci G, Ferrini S, Fabbi M. Cytokine-Induced Guanylate Binding Protein 1 (GBP1) Release from Human Ovarian Cancer Cells. Cancers. 2020; 12(2):488. https://doi.org/10.3390/cancers12020488
Chicago/Turabian StyleCarbotti, Grazia, Andrea Petretto, Elisabeth Naschberger, Michael Stürzl, Stefania Martini, Maria Cristina Mingari, Gilberto Filaci, Silvano Ferrini, and Marina Fabbi. 2020. "Cytokine-Induced Guanylate Binding Protein 1 (GBP1) Release from Human Ovarian Cancer Cells" Cancers 12, no. 2: 488. https://doi.org/10.3390/cancers12020488
APA StyleCarbotti, G., Petretto, A., Naschberger, E., Stürzl, M., Martini, S., Mingari, M. C., Filaci, G., Ferrini, S., & Fabbi, M. (2020). Cytokine-Induced Guanylate Binding Protein 1 (GBP1) Release from Human Ovarian Cancer Cells. Cancers, 12(2), 488. https://doi.org/10.3390/cancers12020488