Mitochondrial Respiration Correlates with Prognostic Markers in Chronic Lymphocytic Leukemia and Is Normalized by Ibrutinib Treatment

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
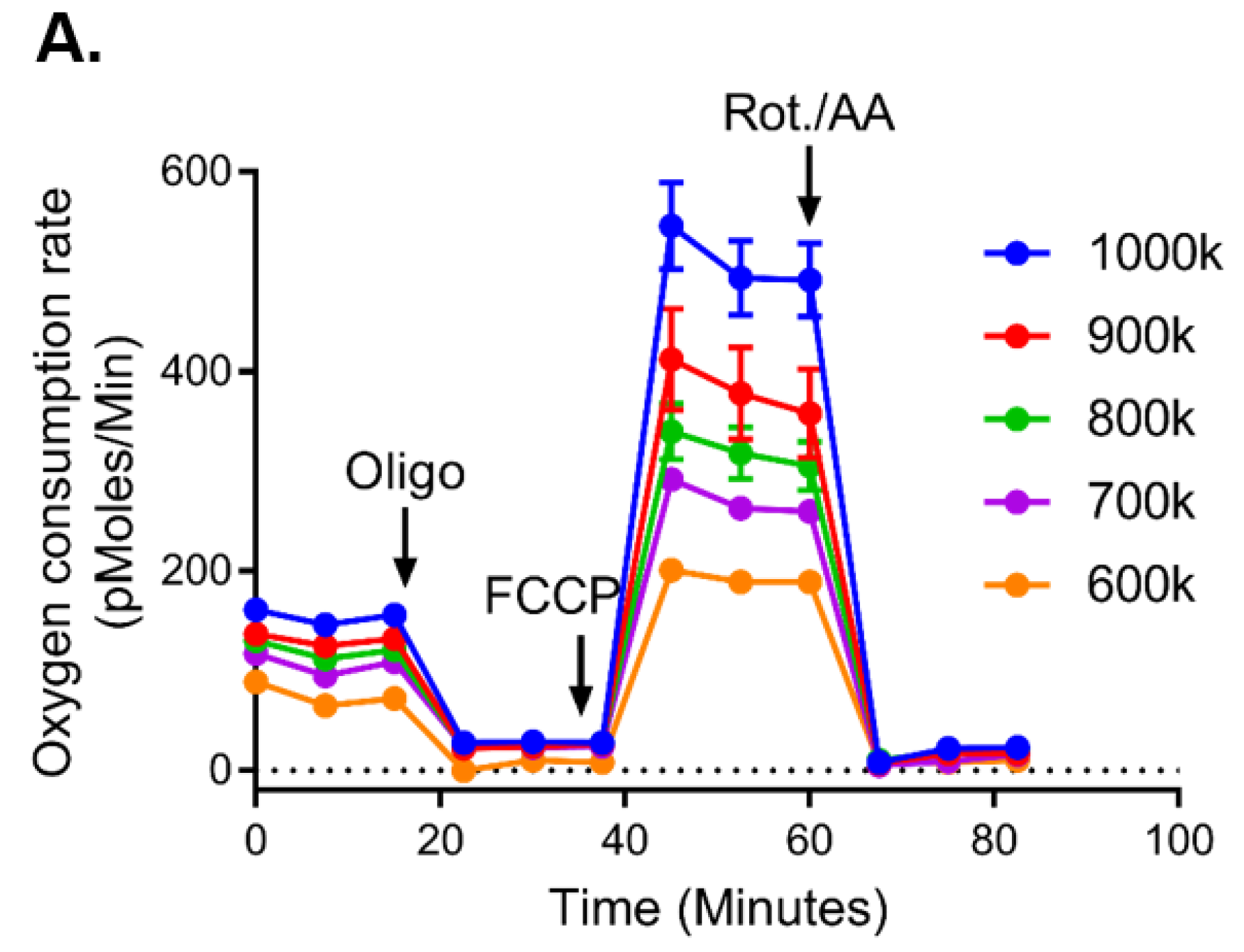
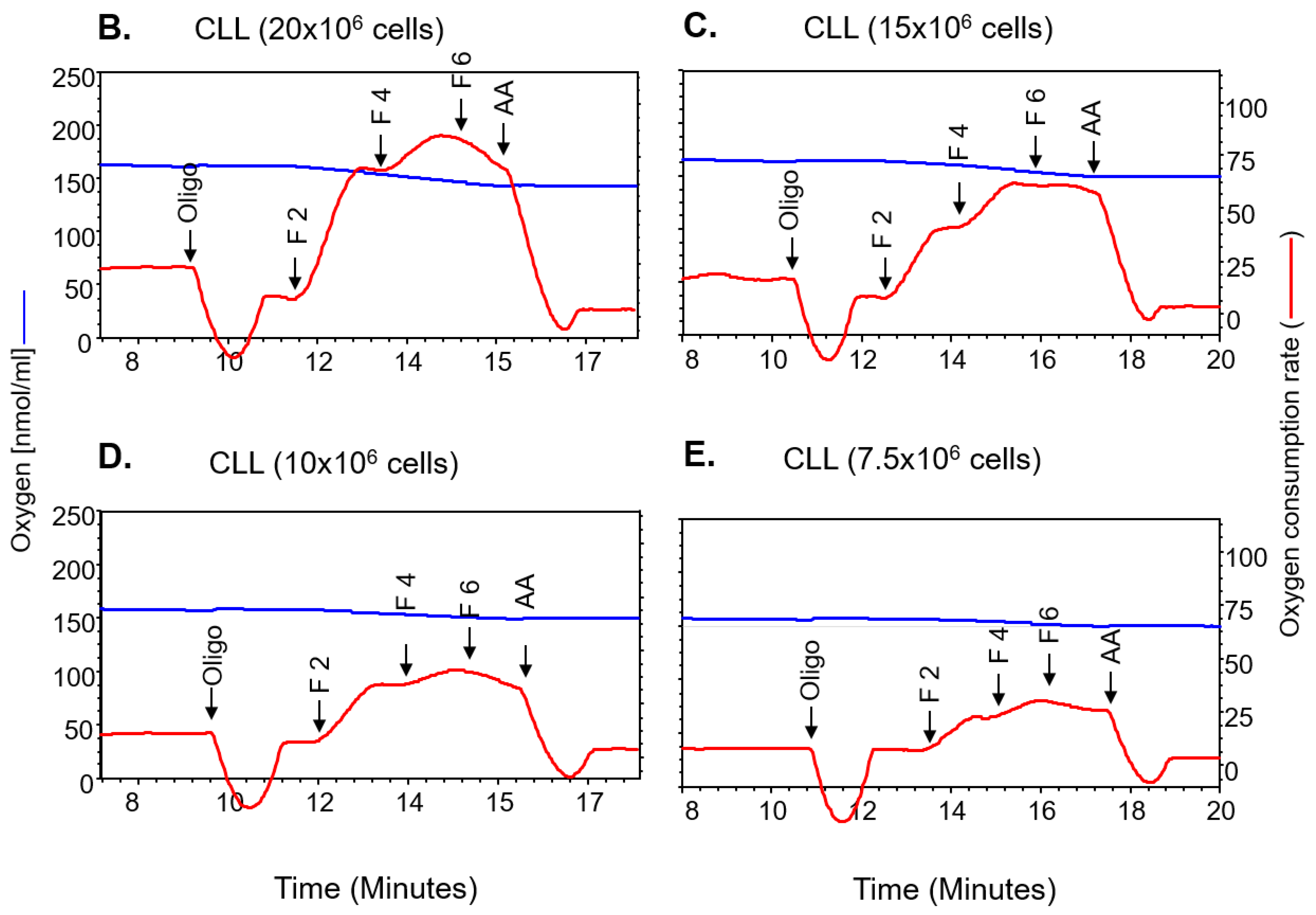
2.1. Mitochondrial Bioenergetics Measured Across Two Platforms Are Equivalent
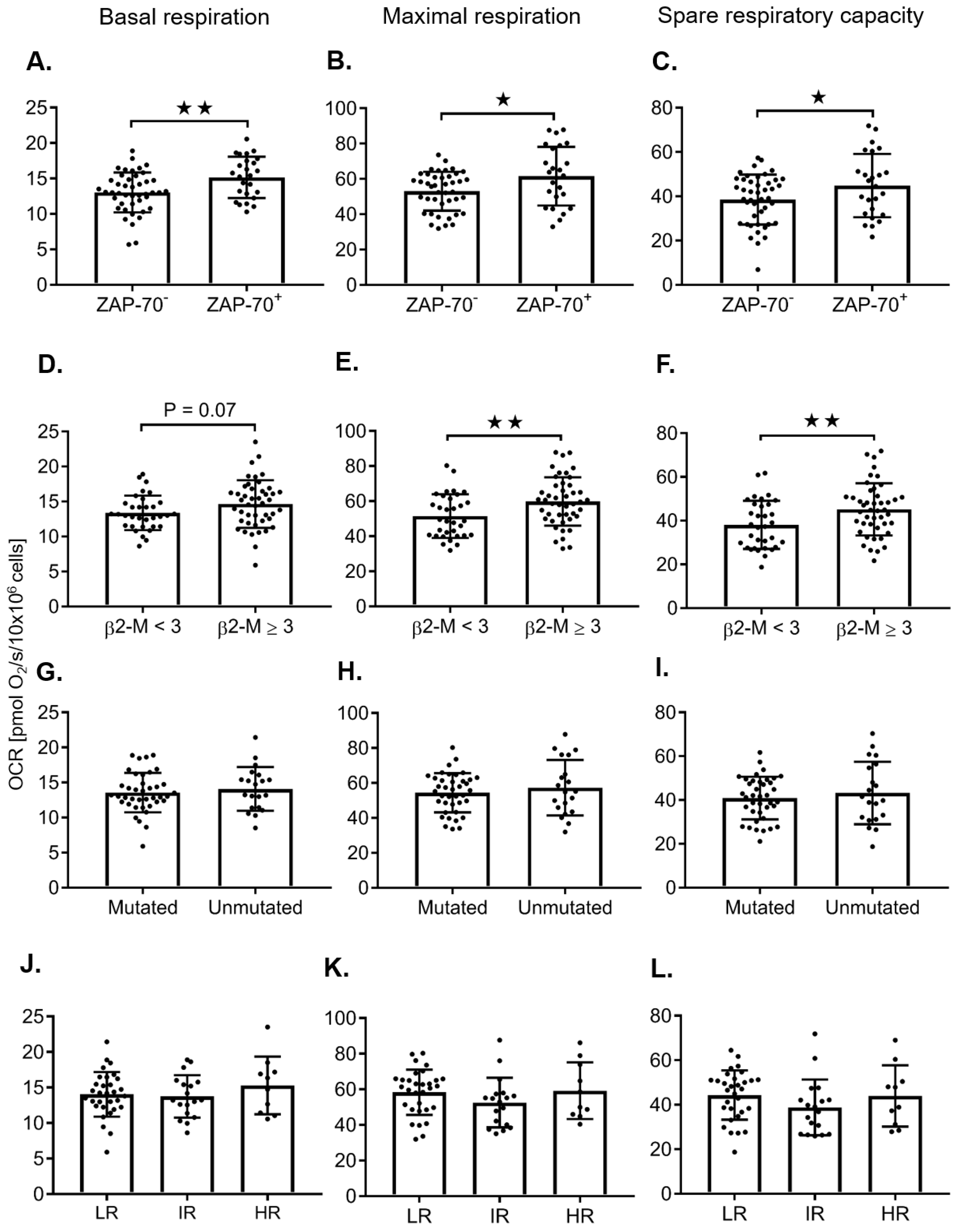
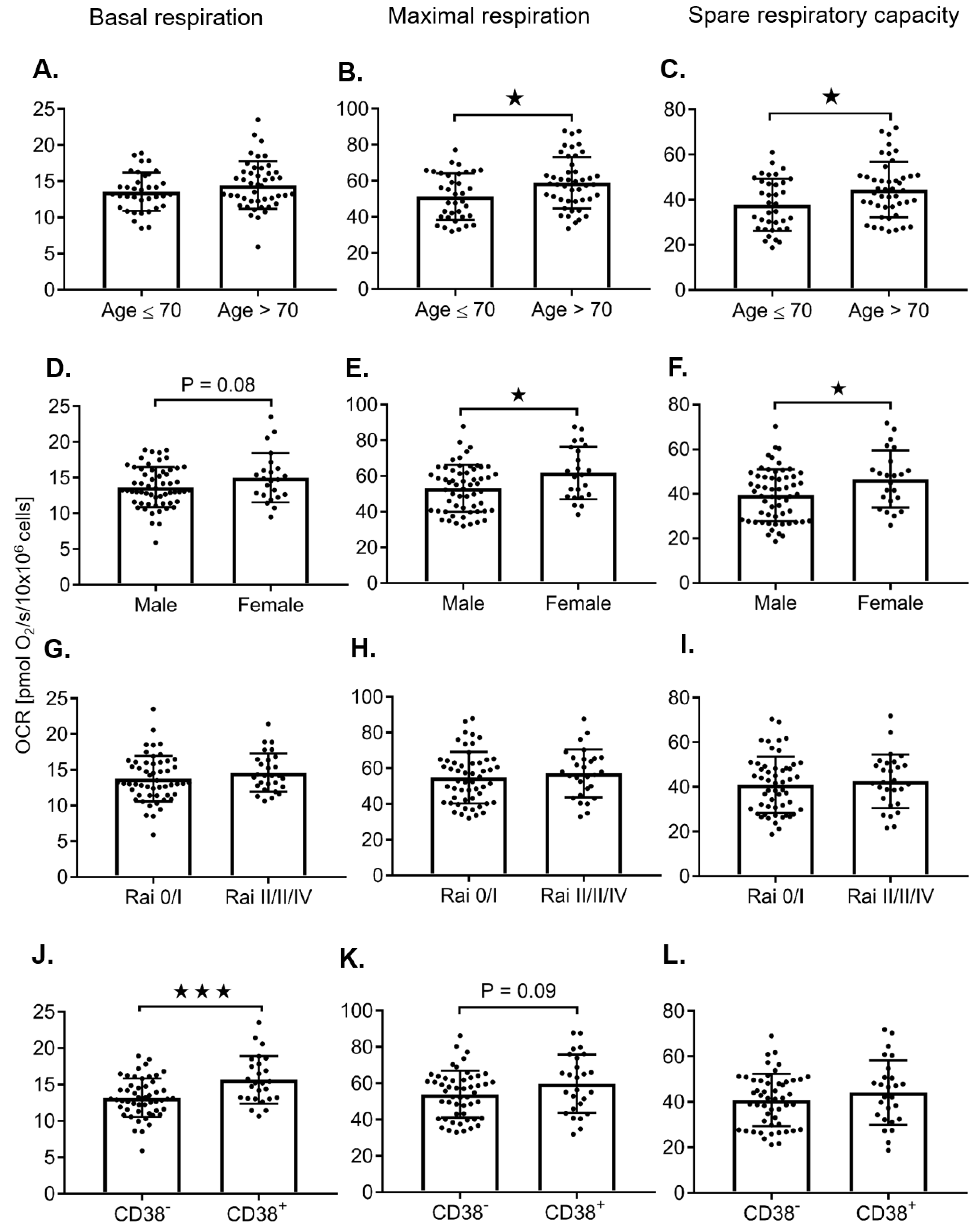
2.2. Adverse Prognostic Markers Predict Increased Maximal Respiration Rates
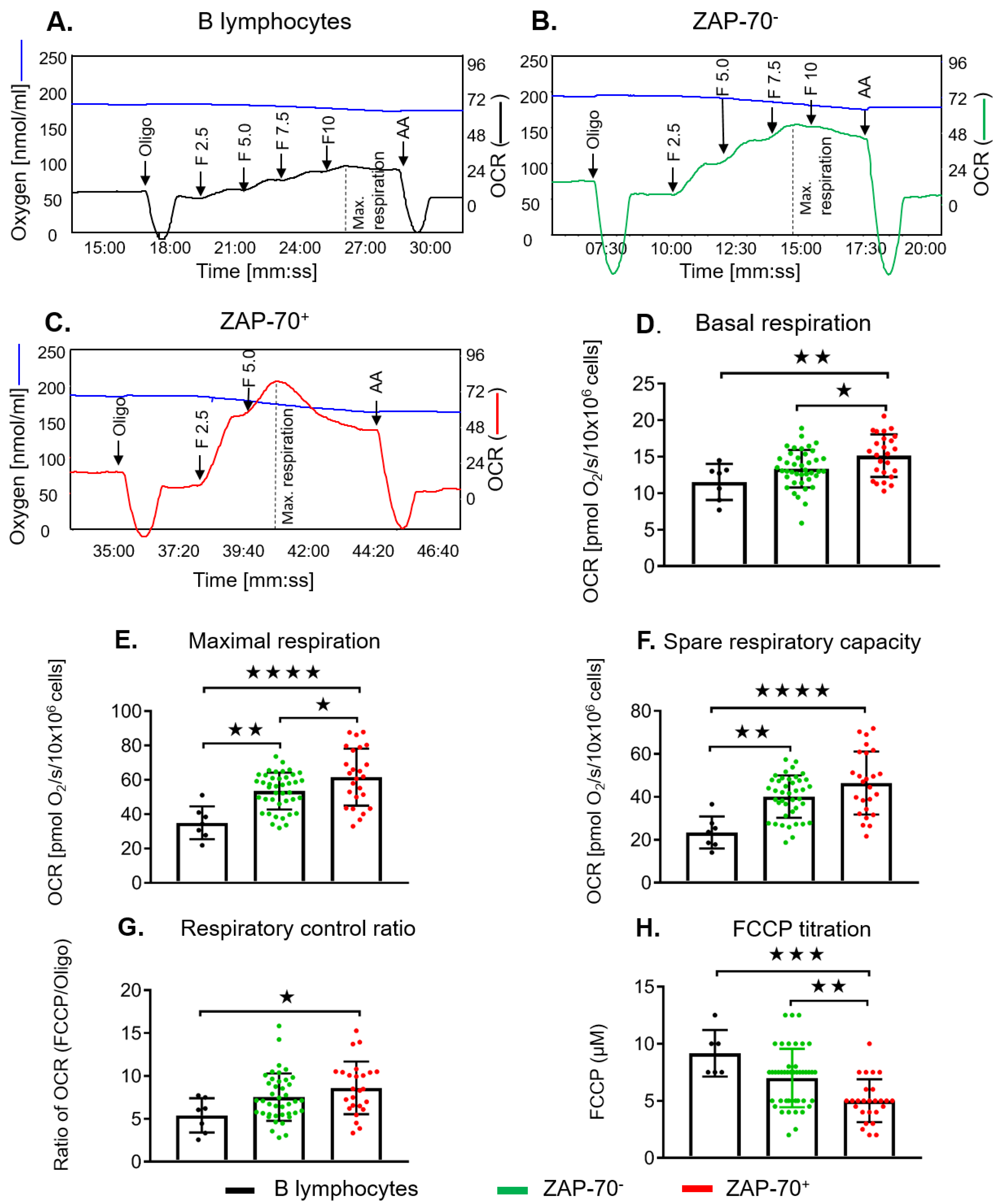
2.3. ZAP-70+ CLL Cells Have Increased Maximal Respiration
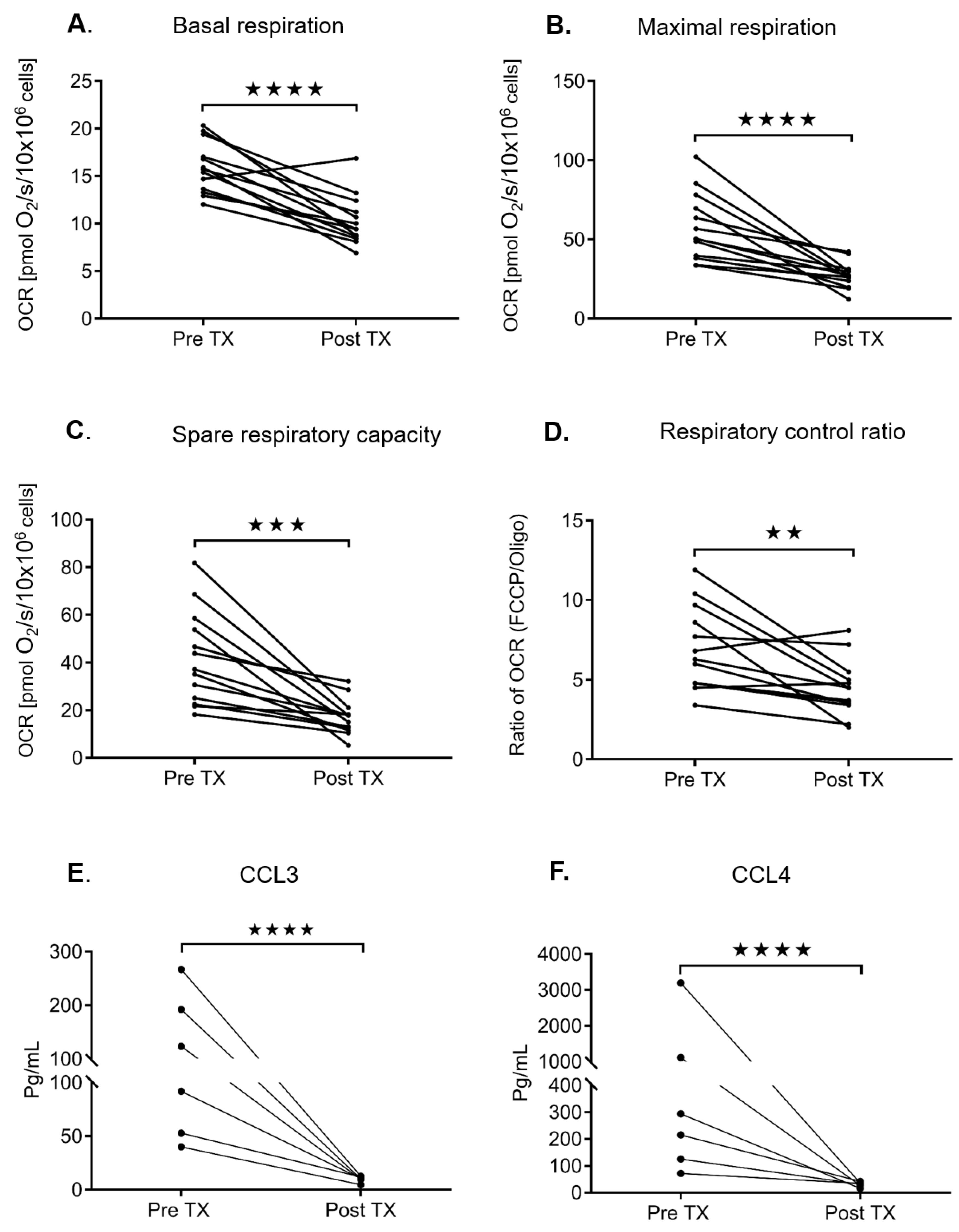
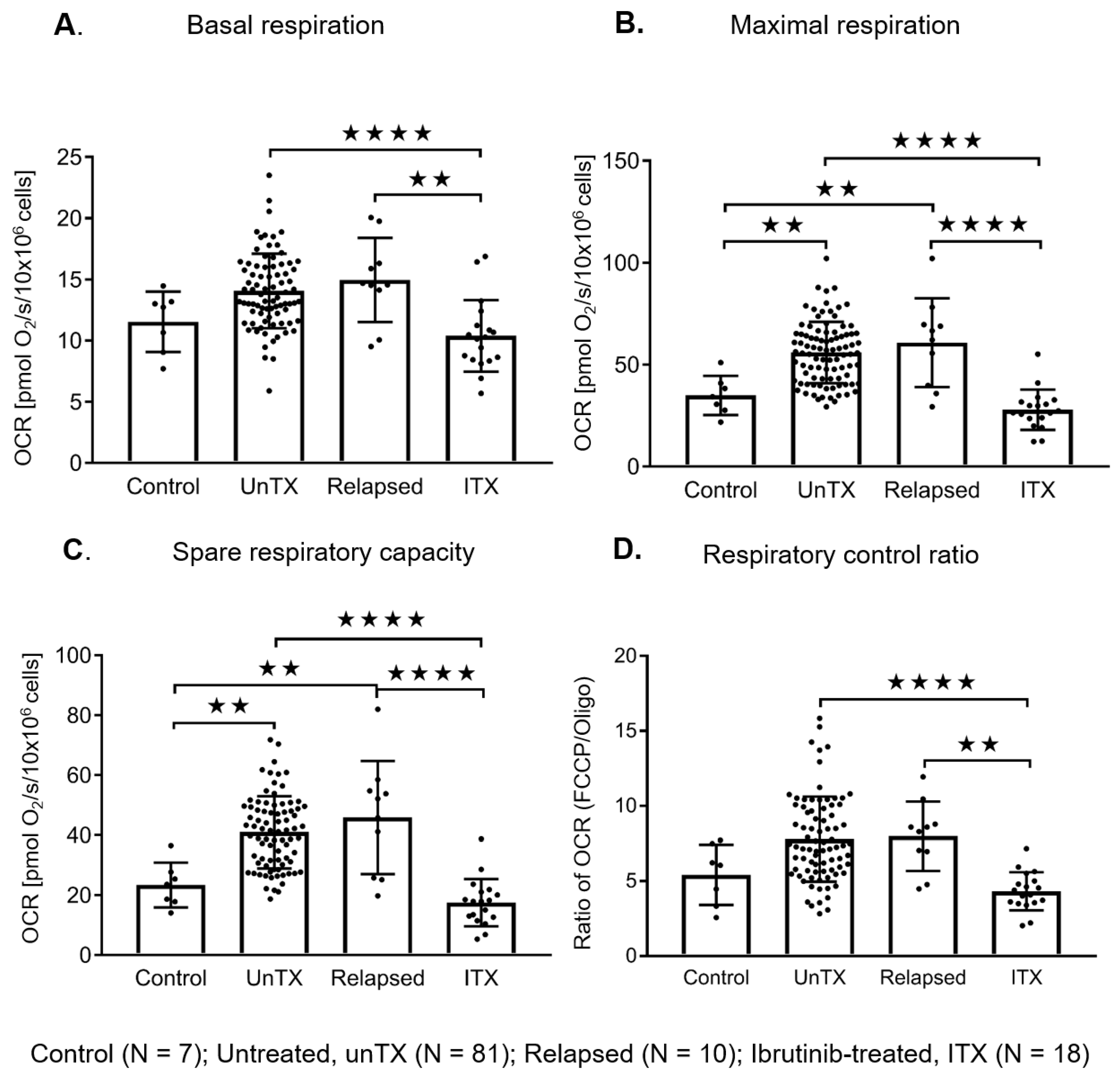
2.4. Ibrutinib Treatment Normalizes the Mitochondrial Respiration Profile and Down Regulates the BCR Signaling in CLL Cells
3. Discussion
4. Materials and Methods
4.1. Primary Samples
4.2. Measurement of Mitochondrial Respiration in Isolated CLL Cells and B Lymphocytes
4.3. Cell Viability Ex Vivo in CLL Cells from Untreated and Ibrutinib-Treated Patients
4.4. Quantification of CCL3/CCL4 Levels in Plasma from Ibrutinib-Treated CLL Patients
4.5. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Antimycin A |
| ATP | Adenosine triphosphate |
| BCR | B cell receptor |
| BTK | Burton’s tyrosine kinase |
| CD38 | Cluster of differentiation 38 |
| CCL3 | Chemokine (C-C motif) ligand 3 |
| CCL4 | Chemokine (C-C motif) ligand 4 |
| CLL | Chronic lymphocytic leukemia |
| β2-M | β2-microglobulin |
| ECAR | Extracellular acidification rate |
| ETC | Electron transport chain |
| FCCP | Carbonyl cyanide-p-trifluoro-methoxyphenylhydrazone |
| FISH | Fluorescence in situ hybridization |
| IgVH | Immunoglobulin variable heavy chain |
| MBL | Monoclonal B cell lymphocytosis |
| OCR | Oxygen consumption rates |
| Oligo | Oligomycin |
| PBMC | Peripheral blood mononuclear cells |
| PI3K | Phosphoinositide 3-kinase |
| ROS | Reactive oxygen species |
| Rot. | Rotenone |
| WBC | White blood cell count |
| ZAP-70 | Zeta-chain-associated protein of 70 kD |
References
- Hallek, M. Chronic lymphocytic leukemia: 2015 Update on diagnosis, risk stratification, and treatment. Am. J. Hematol. 2015, 90, 446–460. [Google Scholar] [CrossRef]
- Oppezzo, P.; Dighiero, G. Role of the B-cell receptor and the microenvironment in chronic lymphocytic leukemia. Blood Cancer J. 2013, 3, e149. [Google Scholar] [CrossRef] [Green Version]
- Shanafelt, T.D.; Geyer, S.M.; Kay, N.E. Prognosis at diagnosis: Integrating molecular biologic insights into clinical practice for patients with CLL. Blood 2004, 103, 1202–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Bockstaele, F.; Verhasselt, B.; Philippe, J. Prognostic markers in chronic lymphocytic leukemia: A comprehensive review. Blood Rev. 2009, 23, 25–47. [Google Scholar] [CrossRef] [PubMed]
- Orchard, J.A.; Ibbotson, R.E.; Davis, Z.; Wiestner, A.; Rosenwald, A.; Thomas, P.W.; Hamblin, T.J.; Staudt, L.M.; Oscier, D.G. ZAP-70 expression and prognosis in chronic lymphocytic leukaemia. Lancet 2004, 363, 105–111. [Google Scholar] [CrossRef]
- Herishanu, Y.; Perez-Galan, P.; Liu, D.; Biancotto, A.; Pittaluga, S.; Vire, B.; Gibellini, F.; Njuguna, N.; Lee, E.; Stennett, L.; et al. The lymph node microenvironment promotes B-cell receptor signaling, NF-kappaB activation, and tumor proliferation in chronic lymphocytic leukemia. Blood 2011, 117, 563–574. [Google Scholar] [CrossRef]
- Burger, J.A.; Quiroga, M.P.; Hartmann, E.; Burkle, A.; Wierda, W.G.; Keating, M.J.; Rosenwald, A. High-level expression of the T-cell chemokines CCL3 and CCL4 by chronic lymphocytic leukemia B cells in nurselike cell cocultures and after BCR stimulation. Blood 2009, 113, 3050–3058. [Google Scholar] [CrossRef]
- Sivina, M.; Hartmann, E.; Kipps, T.J.; Rassenti, L.; Krupnik, D.; Lerner, S.; LaPushin, R.; Xiao, L.; Huang, X.; Werner, L.; et al. CCL3 (MIP-1α) plasma levels and the risk for disease progression in chronic lymphocytic leukemia. Blood 2011, 117, 1662–1669. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Sivina, M.; Hoellenriegel, J.; Oki, Y.; Hagemeister, F.B.; Fayad, L.; Romaguera, J.E.; Fowler, N.; Fanale, M.A.; Kwak, L.W.; et al. CCL3 and CCL4 are biomarkers for B cell receptor pathway activation and prognostic serum markers in diffuse large B cell lymphoma. Br. J. Haematol. 2015, 171, 726–735. [Google Scholar] [CrossRef]
- Chen, L.S.; Bose, P.; Cruz, N.D.; Jiang, Y.; Wu, Q.; Thompson, P.A.; Feng, S.; Kroll, M.H.; Qiao, W.; Huang, X.; et al. A pilot study of lower doses of ibrutinib in patients with chronic lymphocytic leukemia. Blood 2018, 132, 2249–2259. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenabeele, P.; Galluzzi, L.; Vanden Berghe, T.; Kroemer, G. Molecular mechanisms of necroptosis: An ordered cellular explosion. Nat. Rev. Mol. Cell Biol. 2010, 11, 700–714. [Google Scholar] [CrossRef] [PubMed]
- Ward, P.S.; Thompson, C.B. Metabolic reprogramming: A cancer hallmark even warburg did not anticipate. Cancer Cell 2012, 21, 297–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeBerardinis, R.J.; Mancuso, A.; Daikhin, E.; Nissim, I.; Yudkoff, M.; Wehrli, S.; Thompson, C.B. Beyond aerobic glycolysis: Transformed cells can engage in glutamine metabolism that exceeds the requirement for protein and nucleotide synthesis. Proc. Natl. Acad. Sci. USA 2007, 104, 19345–19350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyanaraman, B.; Cheng, G.; Hardy, M.; Ouari, O.; Lopez, M.; Joseph, J.; Zielonka, J.; Dwinell, M.B. A review of the basics of mitochondrial bioenergetics, metabolism, and related signaling pathways in cancer cells: Therapeutic targeting of tumor mitochondria with lipophilic cationic compounds. Redox. Biol. 2018, 14, 316–327. [Google Scholar] [CrossRef]
- Carew, J.S.; Nawrocki, S.T.; Xu, R.H.; Dunner, K.; McConkey, D.J.; Wierda, W.G.; Keating, M.J.; Huang, P. Increased mitochondrial biogenesis in primary leukemia cells: The role of endogenous nitric oxide and impact on sensitivity to fludarabine. Leukemia 2004, 18, 1934–1940. [Google Scholar] [CrossRef] [PubMed]
- Jitschin, R.; Hofmann, A.D.; Bruns, H.; Giessl, A.; Bricks, J.; Berger, J.; Saul, D.; Eckart, M.J.; Mackensen, A.; Mougiakakos, D. Mitochondrial metabolism contributes to oxidative stress and reveals therapeutic targets in chronic lymphocytic leukemia. Blood 2014, 123, 2663–2672. [Google Scholar] [CrossRef] [Green Version]
- Byrd, J.C.; Furman, R.R.; Coutre, S.E.; Flinn, I.W.; Burger, J.A.; Blum, K.A.; Grant, B.; Sharman, J.P.; Coleman, M.; Wierda, W.G.; et al. Targeting BTK with ibrutinib in relapsed chronic lymphocytic leukemia. N. Engl. J. Med. 2013, 369, 32–42. [Google Scholar] [CrossRef]
- Hoellenriegel, J.; Meadows, S.A.; Sivina, M.; Wierda, W.G.; Kantarjian, H.; Keating, M.J.; Giese, N.; O’Brien, S.; Yu, A.; Miller, L.L.; et al. The phosphoinositide 3’-kinase delta inhibitor, CAL-101, inhibits B-cell receptor signaling and chemokine networks in chronic lymphocytic leukemia. Blood 2011, 118, 3603–3612. [Google Scholar] [CrossRef] [Green Version]
- Macias-Perez, I.M.; Flinn, I.W. GS-1101: A delta-specific PI3K inhibitor in chronic lymphocytic leukemia. Curr. Hematol. Malig. Rep. 2013, 8, 22–27. [Google Scholar] [CrossRef]
- Woyach, J.A.; Smucker, K.; Smith, L.L.; Lozanski, A.; Zhong, Y.; Ruppert, A.S.; Lucas, D.; Williams, K.; Zhao, W.; Rassenti, L.; et al. Prolonged lymphocytosis during ibrutinib therapy is associated with distinct molecular characteristics and does not indicate a suboptimal response to therapy. Blood 2014, 123, 1810–1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vangapandu, H.V.; Havranek, O.; Ayres, M.L.; Kaipparettu, B.A.; Balakrishnan, K.; Wierda, W.G.; Keating, M.J.; Davis, R.E.; Stellrecht, C.M.; Gandhi, V. B-cell Receptor Signaling Regulates Metabolism in Chronic Lymphocytic Leukemia. Mol. Cancer Res. 2017, 15, 1692–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vangapandu, H.V.; Gandhi, V. Extracellular Flux Assays to Determine Oxidative Phosphorylation and Glycolysis in Chronic Lymphocytic Leukemia Cells. Methods Mol. Biol. 2019, 1881, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Mowery, Y.M.; Lanasa, M.C. Clinical aspects of monoclonal B-cell lymphocytosis. Cancer Control 2012, 19, 8–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventura-Clapier, R.; Moulin, M.; Piquereau, J.; Lemaire, C.; Mericskay, M.; Veksler, V.; Garnier, A. Mitochondria: A central target for sex differences in pathologies. Clin. Sci. 2017, 131, 803–822. [Google Scholar] [CrossRef] [PubMed]
- Koczula, K.M.; Ludwig, C.; Hayden, R.; Cronin, L.; Pratt, G.; Parry, H.; Tennant, D.; Drayson, M.; Bunce, C.M.; Khanim, F.L.; et al. Metabolic plasticity in CLL: Adaptation to the hypoxic niche. Leukemia 2016, 30, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Gdynia, G.; Robak, T.; Kopitz, J.; Heller, A.; Grekova, S.; Duglova, K.; Laukemper, G.; Heinzel-Gutenbrunner, M.; Gutenbrunner, C.; Roth, W.; et al. Distinct Activities of Glycolytic Enzymes Identify Chronic Lymphocytic Leukemia Patients with a more Aggressive Course and Resistance to Chemo-Immunotherapy. EBioMedicine 2018, 32, 125–133. [Google Scholar] [CrossRef]
- Chowdhury, S.K.; Zherebitskaya, E.; Smith, D.R.; Akude, E.; Chattopadhyay, S.; Jolivalt, C.G.; Calcutt, N.A.; Fernyhough, P. Mitochondrial respiratory chain dysfunction in dorsal root ganglia of streptozotocin-induced diabetic rats and its correction by insulin treatment. Diabetes 2010, 59, 1082–1091. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Widhopf, G.; Huynh, L.; Rassenti, L.; Rai, K.R.; Weiss, A.; Kipps, T.J. Expression of ZAP-70 is associated with increased B-cell receptor signaling in chronic lymphocytic leukemia. Blood 2002, 100, 4609–4614. [Google Scholar] [CrossRef]
- Seftel, M.D.; Demers, A.A.; Banerji, V.; Gibson, S.B.; Morales, C.; Musto, G.; Pitz, M.W.; Johnston, J.B. High incidence of chronic lymphocytic leukemia (CLL) diagnosed by immunophenotyping: A population-based Canadian cohort. Leuk. Res. 2009, 33, 1463–1468. [Google Scholar] [CrossRef] [PubMed]
- Catovsky, D.; Wade, R.; Else, M. The clinical significance of patients’ sex in chronic lymphocytic leukemia. Haematologica 2014, 99, 1088–1094. [Google Scholar] [CrossRef] [PubMed]
- Uminski, K.; Brown, K.; Bucher, O.; Hibbert, I.; Dhaliwal, D.H.; Johnston, J.B.; Geirnaert, M.; Dawe, D.E.; Banerji, V. Descriptive analysis of dosing and outcomes for patients with ibrutinib-treated relapsed or refractory chronic lymphocytic leukemia in a Canadian centre. Curr. Oncol. 2019, 26, e610–e617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy Chowdhury, S.; Banerji, V. Targeting Mitochondrial Bioenergetics as a Therapeutic Strategy for Chronic Lymphocytic Leukemia. Oxid. Med. Cell Longev. 2018, 2018. [Google Scholar] [CrossRef]
- Chowdhury, S.K.; Drahota, Z.; Floryk, D.; Calda, P.; Houstek, J. Activities of mitochondrial oxidative phosphorylation enzymes in cultured amniocytes. Clin. Chim. Acta 2000, 298, 157–173. [Google Scholar] [CrossRef]
- Hill, B.G.; Dranka, B.P.; Zou, L.; Chatham, J.C.; Darley-Usmar, V.M. Importance of the bioenergetic reserve capacity in response to cardiomyocyte stress induced by 4-hydroxynonenal. Biochem. J. 2009, 424, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Kost, S.E.; Bouchard, E.D.; LaBossiere, E.; Ye, X.; Queau, M.L.; Liang, W.S.; Banerji, V.; Gibson, S.B.; Katyal, S.; Johnston, J.B. Cross-resistance and synergy with bendamustine in chronic lymphocytic leukemia. Leuk. Res. 2016, 50, 63–71. [Google Scholar] [CrossRef]
- Gehrke, I.; Bouchard, E.D.; Beiggi, S.; Poeppl, A.G.; Johnston, J.B.; Gibson, S.B.; Banerji, V. On-target effect of FK866, a nicotinamide phosphoribosyl transferase inhibitor, by apoptosis-mediated death in chronic lymphocytic leukemia cells. Clin. Cancer Res. 2014, 20, 4861–4872. [Google Scholar] [CrossRef] [Green Version]
- Hipp, S.; Tai, Y.T.; Blanset, D.; Deegen, P.; Wahl, J.; Thomas, O.; Rattel, B.; Adam, P.J.; Anderson, K.C.; Friedrich, M. A novel BCMA/CD3 bispecific T-cell engager for the treatment of multiple myeloma induces selective lysis in vitro and in vivo. Leukemia 2017, 31, 2278. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dependent | Basal Respiration | Maximal Respiration | Spare Respiratory Capacity | Respiratory Control Ratio | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variable | PE | SE | P | N | PE | SE | P | N | PE | SE | P | N | PE | SE | P | N |
| Age > 70 vs. ≤ 70 | - | - | - | 7.67 | 3.04 | 0.014 | 81 | 6.76 | 2.67 | 0.013 | 81 | 1.83 | 0.60 | 0.0032 | 81 | |
| WBC > 70 vs. ≤ 70 | - | - | - | −6.54 | 3.19 | 0.044 | 81 | −5.91 | 2.80 | 0.0382 | 81 | - | - | - | ||
| ZAP-70 ≥ 20 vs < 20 | 1.78 | 0.69 | 0.012 | 66 | 8.10 | 3.36 | 0.019 | 66 | 6.32 | 3.02 | 0.0403 | 66 | - | - | - | |
| CD38 ≥ 20 vs. < 20 | 2.43 | 0.74 | 0.0015 | 71 | - | - | - | - | - | - | - | - | - | |||
| β2-M ≥ 3 vs. < 3 | - | - | - | 8.32 | 3.03 | 0.0076 | 78 | 7.05 | 2.66 | 0.0097 | 78 | 1.26 | 0.62 | 0.0482 | 78 | |
| Male vs. Female | - | - | - | −8.57 | 3.35 | 0.012 | 81 | −7.26 | 2.95 | 0.016 | 81 | - | - | - | ||
| Age > 70 vs. ≤ 70 | - | - | - | - | - | - | - | - | - | 2.05 | 0.75 | 0.0081 | 58 | |||
| CD38 ≥ 20 vs.< 20 | 2.00 | 0.80 | 0.016 | 50 | - | - | - | - | - | - | - | - | - | |||
| β2-M ≥ 3 vs. < 3 | - | - | - | 7.75 | 3.35 | 0.025 | 55 | 6.78 | 2.95 | 0.0255 | 19 | - | - | - | ||
| ZAP-70 ≥ 20 vs. < 20 | 2.97 | 1.10 | 0.015 | 19 | 16.30 | 6.09 | 0.016 | 19 | 13.33 | 5.57 | 0.0286 | 19 | - | - | - | |
| CD38 ≥ 20 vs. < 20 | 4.05 | 1.50 | 0.014 | 21 | - | - | - | - | - | - | - | - | - | |||
| FISH 1 (0 = base) | −0.78 | 1.79 | 0.67 | 16 | - | - | - | - | - | - | - | - | - | |||
| FISH 2 (0 = base) | 5.53 | 2.50 | 0.046 | 16 | - | - | - | - | - | - | - | - | - | |||
| FISH high risk vs. 0.1 | 5.81 | 2.35 | 0.027 | 16 | - | - | - | - | - | - | - | - | - | |||
| Dependent | Basal Respiration | Maximal Respiration | Spare Respiratory Capacity | Respiratory Control Ratio | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variable | PE | SE | P | PE | SE | P | PE | SE | P | PE | SE | P |
| Age > 70 | - | - | - | - | - | - | - | - | - | 1.83 | 0.60 | 0.003 |
| WBC > 70 | - | - | - | −8.57 | 3.11 | 0.007 | −7.60 | 2.73 | 0.007 | - | - | - |
| CD38 ≥ 20% | 2.43 | 0.74 | 0.0015 | - | - | - | - | - | - | - | - | - |
| β2-M ≥ 3 | - | - | - | 10.31 | 3.00 | 0.0010 | 8.82 | 2.62 | 0.001 | - | - | - |
| N | 71 | - | - | 78 | - | - | 78 | - | - | 81 | - | - |
| Age > 70 | - | - | - | - | - | - | - | - | - | 2.05 | 0.75 | 0.008 |
| WBC > 70 | - | - | - | - | - | - | -6.66 | 3.03 | 0.032 | - | - | - |
| CD38 ≥ 20% | 2.00 | 0.80 | 0.016 | - | - | - | - | - | - | - | - | - |
| β2-M ≥ 3 | - | - | - | 7.75 | 3.35 | 0.025 | 8.51 | 2.95 | 0.006 | - | - | - |
| N | 50 | - | - | 55 | - | - | 55 | - | - | 58 | - | - |
| ZAP-7 0 ≥ 20% | 2.97 | 1.10 | 0.015 | 16.30 | 6.09 | 0.016 | 13.33 | 5.57 | 0.029 | - | - | - |
| N | 19 | - | - | 19 | - | - | 19 | - | - | - | - | - |
| Variable | Untreated | Relapsed | Ibrutinib | p-Value | |||
|---|---|---|---|---|---|---|---|
| N | 78 | 10 | 13 | 3 Groups | Relapsed vs. Untreated | Treated vs. Untreated | |
| Age > 70 (%) | 55 | 40 | 54 | 0.69 | 0.50 | 0.64 | |
| WBC > 70 (%) | 36 | 50 | 62 | 0.18 | 0.49 | 0.094 | |
| Male (%) | 72 | 40 | 54 | 0.11 | 0.068 | 0.045 | |
| ZAP-70 ≥ 20 (%) | 38 | 44 | 42 | 1 | 1 | 0.80 | |
| n | 63 | 9 | 12 | - | - | - | |
| CD38 ≥ 20 (%) | 31 | 38 | 67 | 0.070 | 1 | 0.065 | |
| n | 68 | 8 | 12 | - | - | - | |
| β2-M ≥ 3.5 (%) | 37 | 80 | 50 | 0.031 | 0.0015 | 0.031 | |
| β2-M ≥ 3.0 (%) | 58 | 90 | 67 | 0.12 | 0.081 | 0.13 | |
| n | 76 | 10 | 12 | - | - | - | |
| Rai II–IV (%) | 33 | 100 | 62 | <0.001 | 0.0000 | 0.0003 | |
| n | 78 | 10 | 13 | - | - | - | |
| FISH (%) | 0 | 53 | 56 | 15 | 0.6 | 1 | 0.2 |
| 1 | 30 | 22 | 46 | - | - | - | |
| 2 | 18 | 22 | 38 | - | - | - | |
| n | 57 | 9 | 13 | - | - | - | |
| Um IgVH (%) | 33 | 50 | 73 | 0.048 | 0.71 | 0.038 | |
| n | 58 | 9 | 11 | - | - | - | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roy Chowdhury, S.; Bouchard, E.D.J.; Saleh, R.; Nugent, Z.; Peltier, C.; Mejia, E.; Hou, S.; McFall, C.; Squires, M.; Hewitt, D.; et al. Mitochondrial Respiration Correlates with Prognostic Markers in Chronic Lymphocytic Leukemia and Is Normalized by Ibrutinib Treatment. Cancers 2020, 12, 650. https://doi.org/10.3390/cancers12030650
Roy Chowdhury S, Bouchard EDJ, Saleh R, Nugent Z, Peltier C, Mejia E, Hou S, McFall C, Squires M, Hewitt D, et al. Mitochondrial Respiration Correlates with Prognostic Markers in Chronic Lymphocytic Leukemia and Is Normalized by Ibrutinib Treatment. Cancers. 2020; 12(3):650. https://doi.org/10.3390/cancers12030650
Chicago/Turabian StyleRoy Chowdhury, Subir, Eric D. J. Bouchard, Ryan Saleh, Zoann Nugent, Cheryl Peltier, Edgard Mejia, Sen Hou, Carly McFall, Mandy Squires, Donna Hewitt, and et al. 2020. "Mitochondrial Respiration Correlates with Prognostic Markers in Chronic Lymphocytic Leukemia and Is Normalized by Ibrutinib Treatment" Cancers 12, no. 3: 650. https://doi.org/10.3390/cancers12030650
APA StyleRoy Chowdhury, S., Bouchard, E. D. J., Saleh, R., Nugent, Z., Peltier, C., Mejia, E., Hou, S., McFall, C., Squires, M., Hewitt, D., Davidson, L., Shen, G. X., Johnston, J. B., Doucette, C., Hatch, G. M., Fernyhough, P., Marshall, A., Gibson, S. B., Dawe, D. E., & Banerji, V. (2020). Mitochondrial Respiration Correlates with Prognostic Markers in Chronic Lymphocytic Leukemia and Is Normalized by Ibrutinib Treatment. Cancers, 12(3), 650. https://doi.org/10.3390/cancers12030650