Potential of Tyrosine Kinase Receptor TIE-1 as Novel Therapeutic Target in High-PI3K-Expressing Ovarian Cancer

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
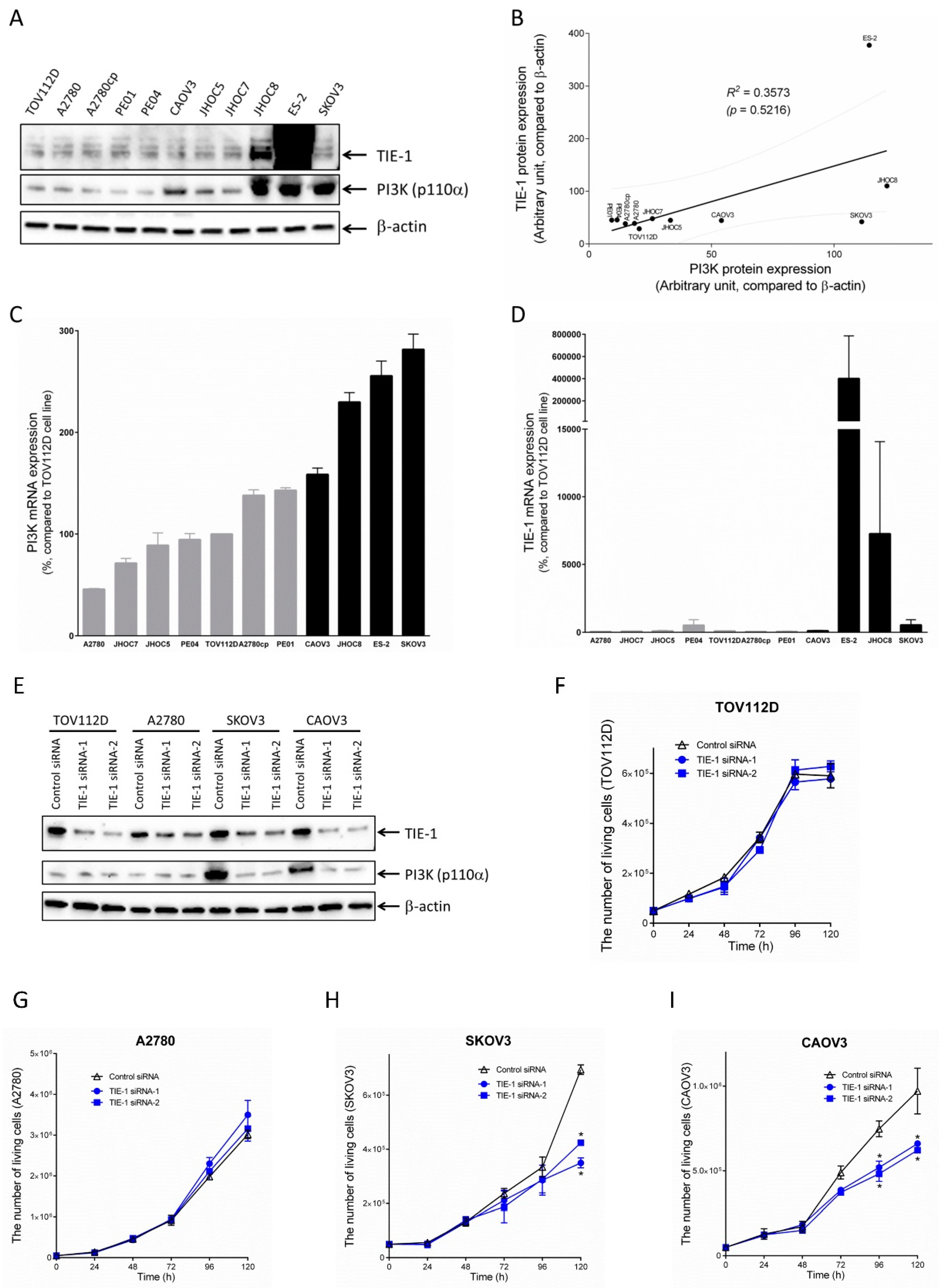
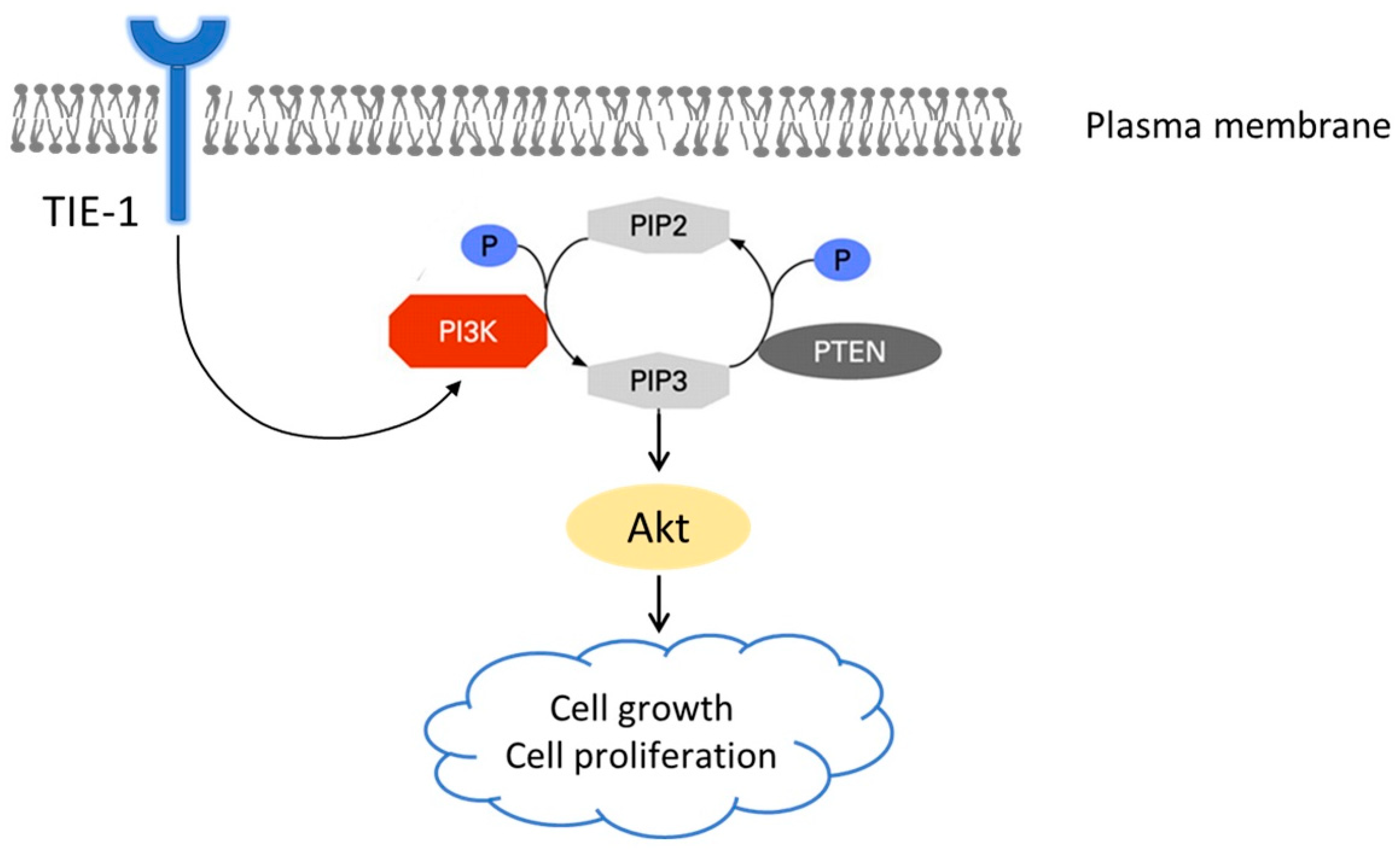
2.1. TIE-1 Might Signal through PI3K/Akt Pathway
2.2. TIE-1 Inhibition Decreases Cell Growth in High-PI3K-Expressing Cell Line.
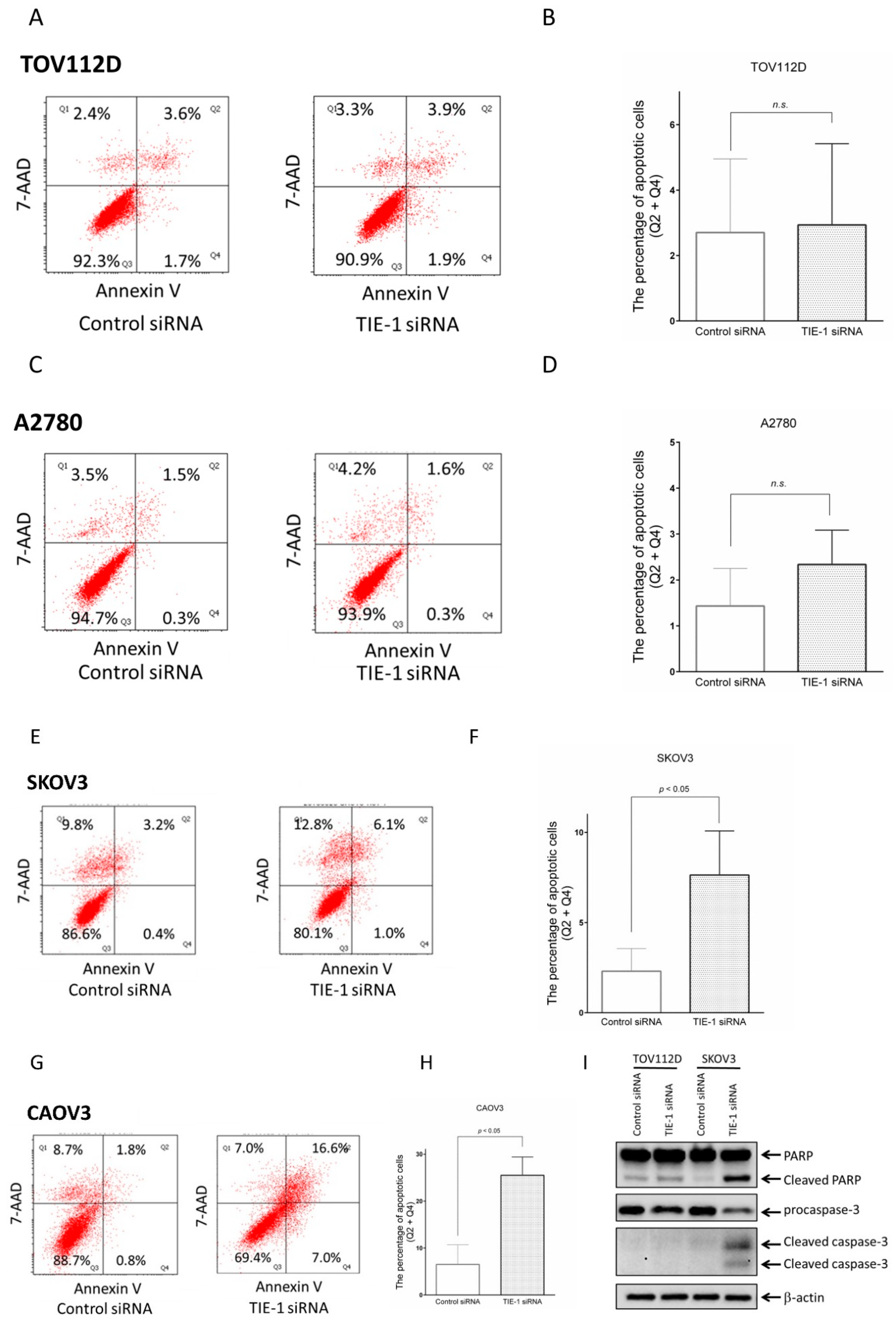
2.3. TIE-1 Inhibition Induces Apoptosis in High-PI3K-Expressing Cell Lines
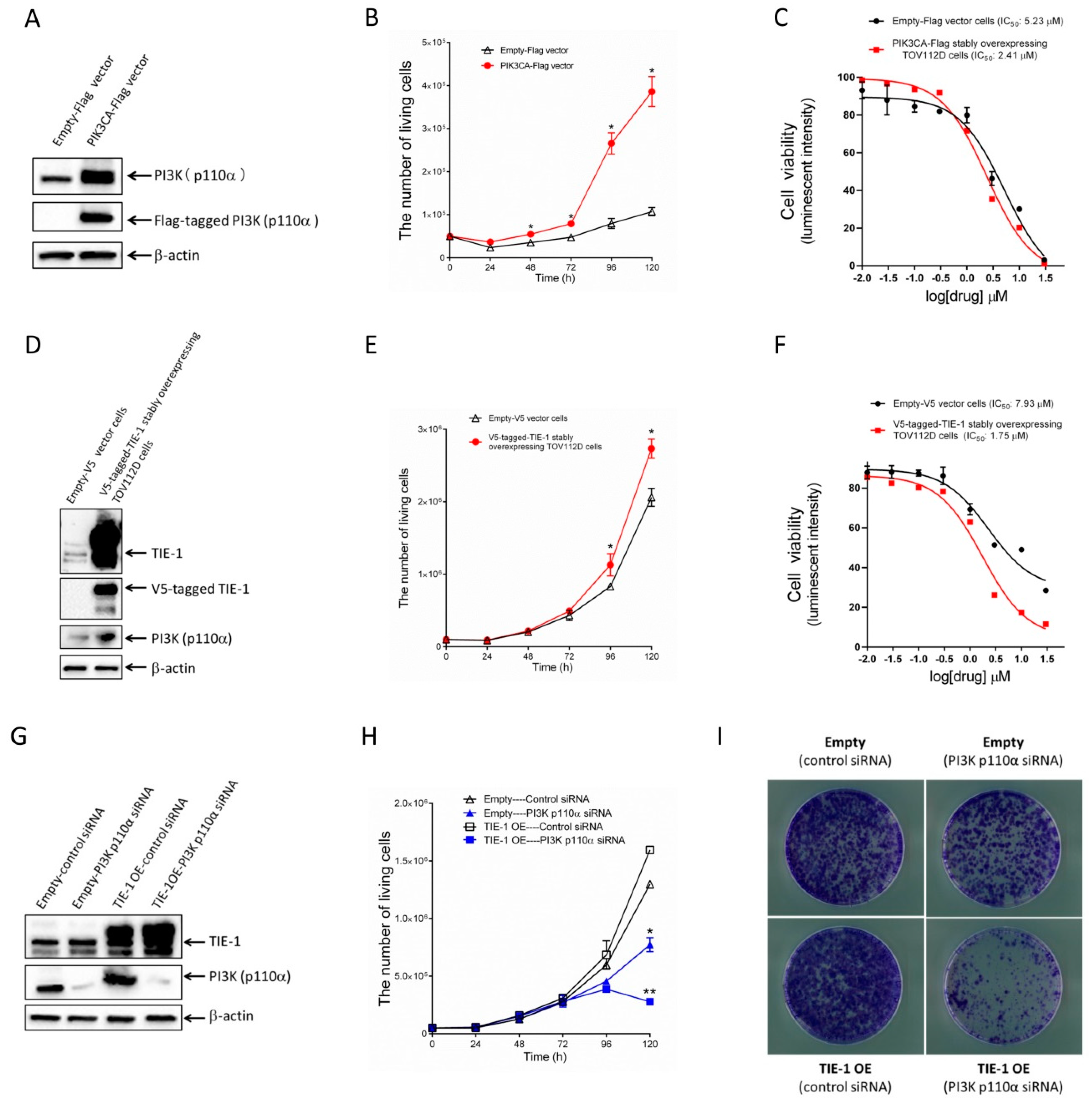
2.4. Increased TIE-1 Expression Induces TIE-1/PI3K-Mediated Cell Growth in Low-PI3K-Expressing Ovarian-Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Antibodies and Reagents
4.2. Cell Culture
4.3. Preparation of TIE-1–V5 Vector
4.4. siRNA transfections
4.5. Vector transfections
4.6. Immunoblotting
4.7. Cell-Viability Assay
4.8. Quantitative Real-Time PCR
4.9. Apoptosis Assay (Annexin V/7-AAD)
4.10. Clonogenic Assay
4.11. Trypan-Blue Exclusion Assay
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Kaye, S.B. New strategies in the treatment of ovarian cancer: Current clinical perspectives and future potential. Clinical. Cancer Res. 2013, 19, 961–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lengyel, E. Ovarian cancer development and metastasis. Am. J. Pathol. 2010, 177, 1053–1064. [Google Scholar] [CrossRef]
- Agarwal, R.; Kaye, S.B. Ovarian cancer: Strategies for overcoming resistance to chemotherapy. Nat. Rev. Cancer 2003, 3, 502–516. [Google Scholar] [CrossRef]
- Puri, M.C.; Partanen, J.; Rossant, J.; Bernstein, A. Interaction of the TEK and TIE receptor tyrosine kinases during cardiovascular development. Development 1999, 126, 4569–4580. [Google Scholar]
- Eklund, L.; Kangas, J.; Saharinen, P. Angiopoietin-Tie signalling in the cardiovascular and lymphatic systems. Clin. Sci. 2017, 131, 87–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.N.; Tozawa, Y.; Deutsch, U.; Wolburg-Buchholz, K.; Fujiwara, Y.; Gendron-Maguire, M.; Gridley, T.; Wolburg, H.; Risau, W.; Qin, Y. Distinct roles of the receptor tyrosine kinases Tie-1 and Tie-2 in blood vessel formation. Nature 1995, 376, 70–74. [Google Scholar] [CrossRef]
- Fagiani, E.; Christofori, G. Angiopoietins in angiogenesis. Cancer Lett. 2013, 328, 18–26. [Google Scholar] [CrossRef]
- Xu, M.; Xu, H.H.; Lin, Y.; Sun, X.; Wang, L.J.; Fang, Z.P.; Su, X.H.; Liang, X.J.; Hu, Y.; Liu, Z.M.; et al. LECT2, a ligand for Tie1, plays a crucial role in liver fibrogenesis. Cell 2019, 178, 1478–1492. [Google Scholar] [CrossRef]
- Yang, P.; Chen, N.; Jia, J.H.; Gao, X.J.; Li, S.H.; Cai, J.; Wang, Z. Tie-1: A potential target for anti-angiogenesis therapy. J. Huazhong Univ. Sci. Technol. [Med. Sci.] 2015, 35, 615–622. [Google Scholar] [CrossRef]
- Puri, M.C.; Rossant, J.; Alitalo, K.; Bernstein, A.; Partanen, J. The receptor tyrosine kinase TIE is required for integrity and survival of vascular endothelial cells. EMBO J. 1995, 14, 5884–5891. [Google Scholar] [CrossRef] [PubMed]
- Verstovsek, S.; Kantarjian, H.; Manshouri, T.; O’Brien, S.; Faderl, S.; Talpaz, M.; Cortes, J.; Albitar, M. Prognostic significance of Tie-1 protein expression in patients with early chronic phase chronic myeloid leukemia. Cancer 2002, 94, 1517–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verstovsek, S.; Estey, E.; Manshouri, T.; Keating, M.; Kantarjian, H.; Giles, F.J.; Albitar, M. High expression of the receptor tyrosine kinase Tie-1 in acute myeloid leukemia and myelodysplastic syndrome. Leuk. Lymphom. 2001, 42, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Rees, K.A.; Singh, H.; Brindle, N.P. The receptor tyrosine kinase Tie1 is expressed and activated in epithelial tumour cell lines. Int. J. Oncol. 2007, 31, 893–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeisberg, E.M.; Potenta, S.; Xie, L.; Zeisberg, M.; Kalluri, R. Discovery of endothelial to mesenchymal transition as a source for carcinoma-associated fibroblasts. Cancer Res. 2007, 67, 10123–10128. [Google Scholar] [CrossRef] [Green Version]
- Ito, Y.; Yoshida, H.; Uruno, T.; Nakano, K.; Takamura, Y.; Miya, A.; Kobayashi, K.; Yokozawa, T.; Matsuzuka, F.; Kuma, K.; et al. Tie-1 tyrosine kinase expression in human thyroid neoplasms. Histopathology 2004, 44, 318–322. [Google Scholar] [CrossRef]
- Salven, P.; Joensuu, H.; Heikkila, P.; Matikainen, M.T.; Wasenius, V.M.; Alanko, A.; Alitalo, K. Endothelial Tie growth factor receptor provides antigenic marker for assessment of breast cancer angiogenesis. Br. J. Cancer 1996, 74, 69–72. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, M.; Toyoshima, M.; Zhang, X.; Hasegawa-Minato, J.; Shigeta, S.; Usui, T.; Kemp, C.J.; Grandori, C.; Kitatani, K.; Yaegashi, N. Tyrosine kinase receptor TIE-1 mediates platinum resistance by promoting nucleotide excision repair in ovarian cancer. Sci. Rep. 2018, 8, 13207. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, Y.; Whitman, M.; Cantley, L.C.; Erikson, R.L. Evidence that the Rous sarcoma virus transforming gene product phosphorylates phosphatidylinositol and diacylglycerol. Proc. Natl. Acad. Sci. USA 1984, 81, 2117–2121. [Google Scholar] [CrossRef] [Green Version]
- Vivanco, I.; Sawyers, C.L. The phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat. Rev. Cancer 2002, 2, 489–501. [Google Scholar] [CrossRef]
- Engelman, J.A.; Luo, J.; Cantley, L.C. The evolution of phosphatidylinositol 3-kinases as regulators of growth and metabolism. Nat. Rev. Genet. 2006, 7, 606–619. [Google Scholar] [CrossRef]
- Fruman, D.A.; Meyers, R.E.; Cantley, L.C. Phosphoinositide kinases. Annu. Rev. Biochem. 1998, 67, 481–507. [Google Scholar] [CrossRef] [PubMed]
- Vanhaesebroeck, B.; Guillermet-Guibert, J.; Graupera, M.; Bilanges, B. The emerging mechanisms of isoform-specific PI3K signalling. Nat. Rev. Mol. Cell Biol. 2010, 11, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Jean, S.; Kiger, A.A. Classes of phosphoinositide 3-kinases at a glance. J. Cell Sci. 2014, 127 Pt 5, 923–928. [Google Scholar] [CrossRef] [Green Version]
- Samuels, Y.; Waldman, T. Oncogenic mutations of PIK3CA in human cancers. Curr. Top. Microbiol. Immunol. 2010, 347, 21–41. [Google Scholar] [PubMed]
- Karakas, B.; Bachman, K.E.; Park, B.H. Mutation of the PIK3CA oncogene in human cancers. Br. J. Cancer 2006, 94, 455–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantley, L.C. The phosphoinositide 3-kinase pathway. Science 2002, 296, 1655–1657. [Google Scholar] [CrossRef]
- Fruman, D.A.; Chiu, H.; Hopkins, B.D.; Bagrodia, S.; Cantley, L.C.; Abraham, R.T. The PI3K Pathway in Human Disease. Cell 2017, 170, 605–635. [Google Scholar] [CrossRef] [Green Version]
- Vanhaesebroeck, B.; Ali, K.; Bilancio, A.; Geering, B.; Foukas, L.C. Signalling by PI3K isoforms: Insights from gene-targeted mice. Trends Biochem. Sci. 2005, 30, 194–204. [Google Scholar] [CrossRef]
- Regad, T. Targeting RTK Signaling Pathways in Cancer. Cancers 2015, 7, 1758–1784. [Google Scholar] [CrossRef]
- Korhonen, E.A.; Lampinen, A.; Giri, H.; Anisimov, A.; Kim, M.; Allen, B.; Fang, S.; D’Amico, G.; Sipila, T.J.; Lohela, M.; et al. Tie1 controls angiopoietin function in vascular remodeling and inflammation. J. Clin. Investig. 2016, 126, 3495–3510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savant, S.; La Porta, S.; Budnik, A.; Busch, K.; Hu, J.; Tisch, N.; Korn, C.; Valls, A.F.; Benest, A.V.; Terhardt, D.; et al. The orphan receptor Tie1 controls angiogenesis and vascular remodeling by differentially regulating Tie2 in Tip and stalk cells. Cell Rep. 2015, 12, 1761–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kontos, C.D.; Cha, E.H.; York, J.D.; Peters, K.G. The endothelial receptor tyrosine kinase Tie1 activates phosphatidylinositol 3-kinase and Akt to inhibit apoptosis. Mol. Cell. Biol. 2002, 22, 1704–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domcke, S.; Sinha, R.; Levine, D.A.; Sander, C.; Schultz, N. Evaluating cell lines as tumour models by comparison of genomic profiles. Nat. Commun. 2013, 4, 2126. [Google Scholar] [CrossRef] [PubMed]
- Gasparri, M.L.; Bardhi, E.; Ruscito, I.; Papadia, A.; Farooqi, A.A.; Marchetti, C.; Bogani, G.; Ceccacci, I.; Mueller, M.D.; Benedetti Panici, P. PI3K/AKT/mTOR pathway in ovarian cancer treatment: Are we on the right track? Geburtshilfe und Frauenheilkd. 2017, 77, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zeng, J.; Shen, K. PI3K/AKT/mTOR signaling pathway as a therapeutic target for ovarian cancer. Arch. Gynecol. Obstet. 2014, 290, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Li, H.; Li, W.; Gui, T.; Yang, J.; Cao, D.; Shen, K. The PI3K/AKT/mTOR pathway is a potential predictor of distinct invasive and migratory capacities in human ovarian cancer cell lines. Oncotarget 2015, 6, 25520–25532. [Google Scholar] [CrossRef]
- Wu, P.; Liu, T.; Hu, Y. PI3K inhibitors for cancer therapy: What has been achieved so far? Curr. Med. Chem. 2009, 16, 916–930. [Google Scholar] [CrossRef]
- Deng, J.; Bai, X.; Feng, X.; Ni, J.; Beretov, J.; Graham, P.; Li, Y. Inhibition of PI3K/Akt/mTOR signaling pathway alleviates ovarian cancer chemoresistance through reversing epithelial–mesenchymal transition and decreasing cancer stem cell marker expression. BMC Cancer 2019, 19, 618. [Google Scholar] [CrossRef]
- Cheaib, B.; Auguste, A.; Leary, A. The PI3K/Akt/mTOR pathway in ovarian cancer: Therapeutic opportunities and challenges. Chin. J. Cancer 2015, 34, 4–16. [Google Scholar] [CrossRef]
- Yang, J.; Nie, J.; Ma, X.; Wei, Y.; Peng, Y.; Wei, X. Targeting PI3K in cancer: Mechanisms and advances in clinical trials. Mol. Cancer 2019, 18, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Name | Histological Type |
|---|---|
| TOV112D | Endometrioid |
| A2780 | Non-serous |
| A2780cp | Non-serous |
| PE01 | High grade serous carcinoma |
| PE04 | High grade serous carcinoma |
| CAOV3 | High grade serous carcinoma |
| JHOC5 | Clear cell carcinoma |
| JHOC7 | Clear cell carcinoma |
| JHOC8 | Clear cell carcinoma |
| ES-2 | Clear cell carcinoma |
| SKOV3 | Non-serous |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Ishibashi, M.; Kitatani, K.; Shigeta, S.; Tokunaga, H.; Toyoshima, M.; Shimada, M.; Yaegashi, N. Potential of Tyrosine Kinase Receptor TIE-1 as Novel Therapeutic Target in High-PI3K-Expressing Ovarian Cancer. Cancers 2020, 12, 1705. https://doi.org/10.3390/cancers12061705
Zhang X, Ishibashi M, Kitatani K, Shigeta S, Tokunaga H, Toyoshima M, Shimada M, Yaegashi N. Potential of Tyrosine Kinase Receptor TIE-1 as Novel Therapeutic Target in High-PI3K-Expressing Ovarian Cancer. Cancers. 2020; 12(6):1705. https://doi.org/10.3390/cancers12061705
Chicago/Turabian StyleZhang, Xuewei, Masumi Ishibashi, Kazuyuki Kitatani, Shogo Shigeta, Hideki Tokunaga, Masafumi Toyoshima, Muneaki Shimada, and Nobuo Yaegashi. 2020. "Potential of Tyrosine Kinase Receptor TIE-1 as Novel Therapeutic Target in High-PI3K-Expressing Ovarian Cancer" Cancers 12, no. 6: 1705. https://doi.org/10.3390/cancers12061705
APA StyleZhang, X., Ishibashi, M., Kitatani, K., Shigeta, S., Tokunaga, H., Toyoshima, M., Shimada, M., & Yaegashi, N. (2020). Potential of Tyrosine Kinase Receptor TIE-1 as Novel Therapeutic Target in High-PI3K-Expressing Ovarian Cancer. Cancers, 12(6), 1705. https://doi.org/10.3390/cancers12061705