Components of the Lectin Pathway of Complement in Haematologic Malignancies



Abstract
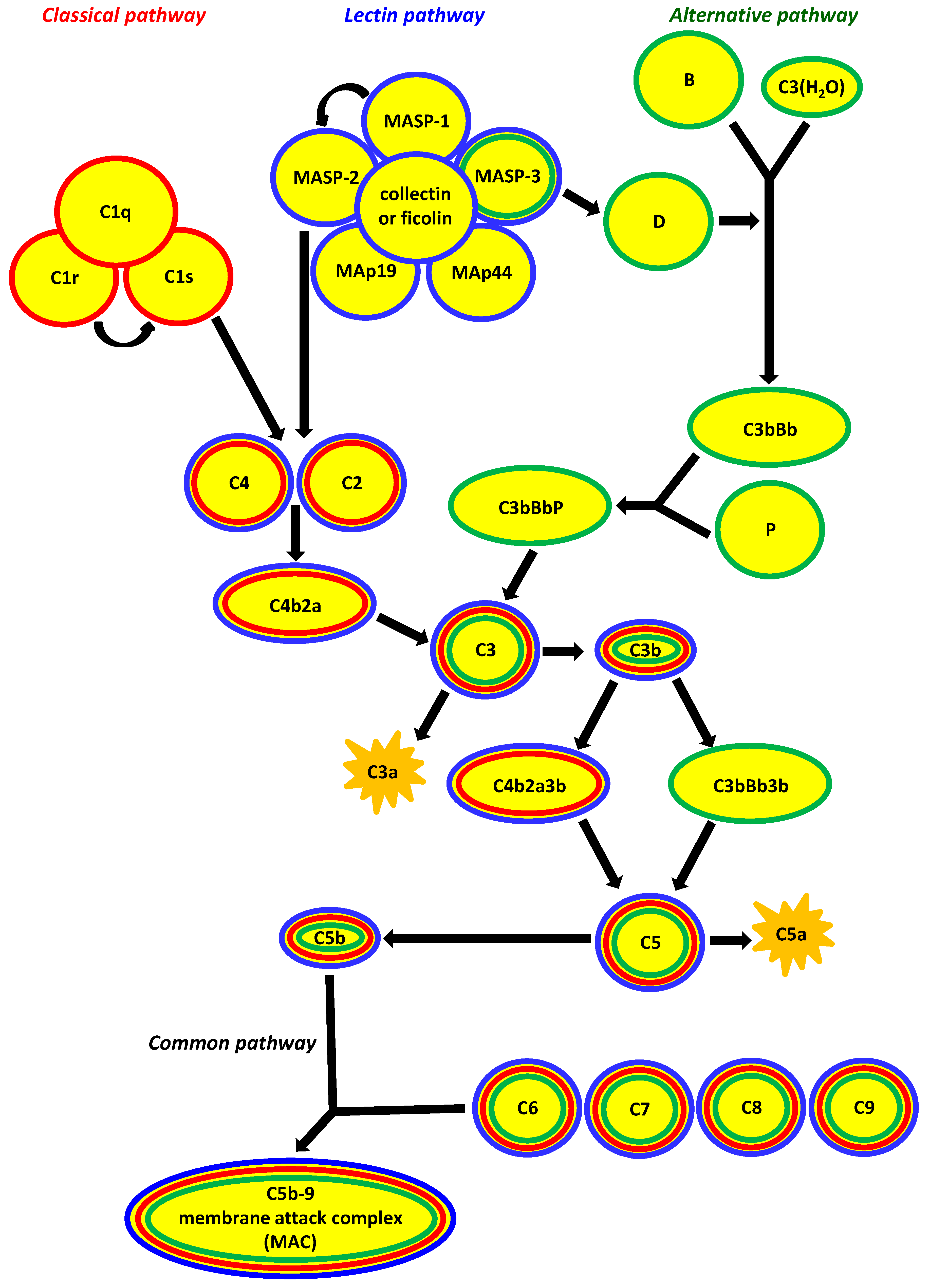
:1. Factors Specific for Activation of the Complement System via the Lectin Pathway
2. Associations of the Complement System with Cancer
3. Haematologic Malignancies
4. Lectin Pathway-Related Pattern-Recognising Molecules
4.1. Collectins
4.2. Ficolins
5. MBL-Associated Serine Proteases (MASP) and Their Related Proteins
5.1. MASP-1, MASP-3 and MAp44
5.2. MASP-2 and MAp19
6. Associations of Lectin Pathway Components with Haematologic Malignancies
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thiel, S. Complement activating soluble pattern recognition molecules with collagen-like regions, mannan binding lectin, ficolins and associated proteins. Mol. Immunol. 2007, 44, 3875–3888. [Google Scholar] [CrossRef] [PubMed]
- Thiel, S.; Gadjeva, M. Humoral pattern recognition molecules: Mannan-binding lectin and ficolins. In Target Pattern Recognition in Innate Immunity; Kishore, U., Ed.; Springer: New York, NY, USA, 2009; pp. 58–73. [Google Scholar]
- Cedzynski, M.; Swierzko, A.S.; Kilpatrick, D.C. Factors of the lectin pathway of complement activation and their clinical associations in neonates. J. Biomed. Biotechnol. 2012, 2012, 364246. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, M.; Endo, Y.; Fujita, T. Structural and functional overview of the lectin complement pathway: Its molecular basis and physiological implication. Arch. Immunol. Ther. Exp. 2013, 61, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Cedzyński, M.; Kilpatrick, D.C.; Świerzko, A.S. Mannose-binding lectin. In The Complement Factsbook, 2nd ed.; Barnum, S., Schein, T., Eds.; Academic Press, Elsevier: London, UK, 2018; pp. 33–43. [Google Scholar]
- Hansen, S.W.; Ohtani, K.; Roy, N.; Wakamiya, N. The collectins CL-L1, CL-K1 and CL-P1, and their roles in complement and innate immunity. Immunobiology 2016, 221, 1058–1067. [Google Scholar] [CrossRef] [Green Version]
- Hansen, S.W.K.; Aagaard, J.B.; Bjerrum, K.B.; Hejbol, E.K.; Nielsen, O.; Schroder, H.D.; Skjoedt, K.; Sorensen, A.L.; Graversen, J.H.; Henriksen, M.L. CL-L1 and CL-K1 exhibit widespread tissue distribution with high and co-localized expression in secretory epithelia and mucosa. Front. Immunol. 2018, 9, 1757. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, K.; Wakamiya, N. The collectins. In The Complement Factsbook, 2nd ed.; Barnum, S., Schein, T., Eds.; Academic Press, Elsevier: London, UK, 2018; pp. 57–65. [Google Scholar]
- Keshi, H.; Sakamoto, T.; Kawai, T.; Ohtani, K.; Katoh, T.; Jang, S.J.; Motomura, W.; Yoshizaki, T.; Fukuda, M.; Koyama, S.; et al. Identification and characterization of a novel human collectin CL-K1. Mol. Immunol. 2006, 50, 1001–1013. [Google Scholar] [CrossRef] [Green Version]
- Hansen, S.; Selman, L.; Palaniyar, N.; Ziegler, K.; Brandt, J.; Kliem, A.; Jonasson, M.; Skjoedt, M.O.; Nielsen, O.; Hartshorn, K.; et al. Collectin 11 (CL-11, CL-K1) is a MASP-1/3-associated plasma collectin with microbial-binding activity. J. Immunol. 2010, 185, 6096–6104. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, M. Ficolins. In The Complement Factsbook, 2nd ed.; Barnum, S., Schein, T., Eds.; Academic Press, Elsevier: London, UK, 2018; pp. 45–56. [Google Scholar]
- Garlatti, V.; Belloy, N.; Martin, L.; Lacroix, M.; Matsushita, M.; Endo, Y.; Fujita, T.; Fontecilla-Camps, J.C.; Arlaud, G.J.; Thielens, N.M.; et al. Structural insights into the innate immune recognition specificities of L- and H-ficolins. EMBO J. 2007, 26, 623–633. [Google Scholar] [CrossRef] [Green Version]
- Degn, S.E.; Jensen, L.; Hansen, A.G.; Duman, D.; Tekin, M.; Jensenius, J.C.; Thiel, S. Mannan-binding lectin-associated serine protease (MASP)-1 is crucial for lectin pathway activation in human serum, whereas neither MASP-1 nor MASP-3 is required for alternative pathway function. J. Immunol. 2012, 189, 3957–3969. [Google Scholar] [CrossRef] [Green Version]
- Yongqing, T.; Drentin, N.; Duncan, R.C.; Wijeyewickrema, L.C.; Pike, R.N. Mannose-binding lectin serine proteases and associated proteins of the lectin pathway of complement: Two genes, five proteins and many functions? Biochim. Biophys. Acta 2012, 1824, 253–262. [Google Scholar] [CrossRef]
- Pihl, R.; Jensenius, J.C.; Thiel, S. MASP-1. In The Complement Factsbook, 2nd ed.; Barnum, S., Schein, T., Eds.; Academic Press, Elsevier: London, UK, 2018; pp. 69–78. [Google Scholar]
- Dobo, J.; Major, B.; Kekesi, K.A.; Szabo, I.; Megyeri, M.; Hajela, K.; Juhasz, G.; Zavodszky, P.; Gal, P. Cleavage of kininogen and subsequent bradykinin release by the complement component: Mannose-binding lectin-associated serine protease (MASP)-1. PLoS ONE 2011, 6, e20036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pihl, R.; Jensenius, J.C.; Thiel, S. MASP-3. In The Complement Factsbook, 2nd ed.; Barnum, S., Schein, T., Eds.; Academic Press, Elsevier: London, UK, 2018; pp. 89–97. [Google Scholar]
- Dobo, J.; Szakacs, D.; Oroszlan, G.; Kortvely, E.; Kiss, B.; Boros, E.; Szasz, R.; Zavodszky, P.; Gal, P.; Pal, G. MASP-3 is the exclusive pro-factor D activator in resting blood: The lectin and the alternative complement pathways are fundamentally linked. Sci. Rep. 2016, 6, 31877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garred, P.; Genster, N.; Pilely, K.; Bayarri-Olmos, R.; Rosbjerg, A.; Ma, Y.J.; Skjoedt, M.O. A journey through the lectin pathway of complement-MBL and beyond. Immunol. Rev. 2016, 274, 74–97. [Google Scholar] [CrossRef] [PubMed]
- Pihl, R.; Jensenius, J.C.; Thiel, S. MASP-2. In The Complement Factsbook, 2nd ed.; Barnum, S., Schein, T., Eds.; Academic Press, Elsevier: London, UK, 2018; pp. 79–87. [Google Scholar]
- Markiewski, M.M.; DeAngelis, R.A.; Benencia, F.; Ricklin-Lichtsteiner, S.K.; Koutoulaki, A.; Gerard, C.; Coukos, G.; Lambris, J.D. Modulation of the antitumor immune response by complement. Nat. Immunol. 2008, 11, 1225–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markiewski, M.; Lambris, J.D. Is complement good or bad for cancer patients? A new perspective on an old dilemma. Trends Immunol. 2009, 30, 286–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swierzko, A.S.; Kilpatrick, D.C.; Cedzynski, M. Mannan-binding lectin in malignancy. Mol. Immunol. 2012, 55, 16–21. [Google Scholar] [CrossRef]
- Pio, R.; Corrales, L.; Lambris, J.D. The role of complement in tumor growth. Adv. Exp. Med. Biol. 2014, 772, 229–262. [Google Scholar]
- Martin, M.; Blom, A.M. Complement in removal of the dead—Balancing inflammation. Immunol. Rev. 2016, 274, 218–232. [Google Scholar] [CrossRef]
- Kochanek, D.M.; Ghouse, S.M.; Karbowniczek, M.M.; Markiewski, M.M. Complementing cancer metastasis. Front. Immunol. 2018, 9, 1629. [Google Scholar] [CrossRef]
- Rus, H.G.; Niculescu, F.; Shin, M.L. Sublytic complement attack induces cell cycle in oligodendrocytes. J. Immunol. 1996, 156, 4892–4900. [Google Scholar]
- Rus, H.G.; Niculescu, F.I.; Shin, M.L. Role of the C5b-9 complement complex in cell cycle and apoptosis. Immunol. Rev. 2001, 180, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Fosbrink, M.; Niculescu, F.; Rus, H. The role of C5b-9 terminal complement complex in activation of the cell cycle and transcription. Immunol. Res. 2005, 31, 37–46. [Google Scholar] [CrossRef]
- Rousseau, S.; Dolado, I.; Beardmore, V.; Shpiro, N.; Marquez, R.; Nebreda, A.R.; Arthur, J.S.; Case, L.M.; Tessier-Lavigne, M.; Gaestel, M.; et al. CXCL12 and C5a trigger cell migration via a PAK1/2-p38alpha MAPK-MAPKAP-K2-HSP27 pathway. Cell Signal. 2006, 18, 1897–1905. [Google Scholar] [CrossRef] [PubMed]
- Rutkowski, M.J.; Sughrue, M.E.; Kane, A.J.; Ahn, B.J.; Fang, S.; Parsa, A.T. The complement cascade as a mediator of tissue growth and regeneration. Inflamm. Res. 2010, 59, 897–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bamberg, C.E.; Mackay, C.R.; Lee, H.; Zahra, D.; Jackson, J.; Lim, Y.S.; Whitfeld, P.L.; Craig, S.; Corsini, E.; Lu, B.; et al. The C5a receptor (C5aR) C5L2 is a modulator of C5aR-mediated signal transduction. J. Biol. Chem. 2010, 285, 7633–7644. [Google Scholar] [CrossRef] [Green Version]
- Mamidi, S.; Hone, S.; Kirschfink, M. The complement system in cancer: Ambivalence between tumour destruction and promotion. Immunobiology 2012, 222, 45–54. [Google Scholar] [CrossRef]
- Liu, X.-Y.; Wang, X.-Y.; Li, R.-Y.; Jia, S.-C.; Sun, P.; Zhao, M.; Fang, C. Recent progress in the understanding of complement activation and its role in tumor growth and anti-tumor therapy. Biomed. Pharmacother. 2017, 91, 446–456. [Google Scholar] [CrossRef]
- Tang, Z.; Lu, B.; Hatch, E.; Sacks, S.H.; Sheerin, N.S. C3a mediates epithelial-to-mesenchymal transition in proteinuric nephropathy. J. Am. Soc. Nephrol. 2009, 20, 593–603. [Google Scholar] [CrossRef] [Green Version]
- Speidl, W.S.; Kastl, S.P.; Hutter, R.; Katsaros, K.M.; Kaun, C.; Bauriedel, G.; Maurer, G.; Huber, K.; Badimon, J.J.; Wojta, J. The complement component C5a is present in human coronary lesions in vivo and induces the expression of MMP-1 and MMP-9 in human macrophages in vitro. FASEB J. 2011, 25, 35–44. [Google Scholar] [CrossRef]
- Corrales, L.; Ajona, D.; Rafail, S.; Lasatre, J.J.; Riezu-Boj, J.I.; Lambris, J.D.; Rouzaut, A.; Pajares, M.J.; Montuenga, L.M.; Pio, R. Anaphylatoxin C5a creates a favorable microenvironment for lung cancer progression. J. Biol. Chem. 2012, 189, 4674–4683. [Google Scholar] [CrossRef]
- Nitta, H.; Wada, Y.; Kawano, Y.; Murakami, Y.; Irie, A.; Taniguchi, K.; Kikuchi, K.; Yamada, G.; Suzuki, K.; Honda, J.; et al. Enhancement of human cancer cell motility and invasiveness by anaphylatoxin C5a via aberrantly expressed C5a receptor (CD88). Clin. Cancer Res. 2013, 19, 2004–2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayegh, E.T.; Bloch, O.; Parsa, A.T. Complement anaphylatoxins as immune regulators in cancer. Cancer Med. 2014, 4, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, M.Z.; Reca, R.; Wysoczynski, M.; Kucia, M.; Baran, J.T.; Allendorf, D.J.; Ratajczak, J.; Ross, G.D. Transplantation studies in C3-deficient animals reveal a novel role of the third complement component (C3) in engrafment of bone marrow cells. Leukemia 2004, 18, 1482–1490. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.M.; Wu, W.; Wysoczynski, M.; Liu, R.; Zuba-Surma, E.K.; Kucia, M.; Ratajczak, J.; Ratajczak, M.Z. Impaired mobilization in of hematopoietic stem/progenitor cells in C5-deficient mice supports the pivotal involvement of innate immunity in this process and reveals novel promobilization effects of granulocytes. Leukemia 2009, 23, 2052–2062. [Google Scholar] [CrossRef] [PubMed]
- Borkowska, S.; Suszynska, M.; Mierzejewska, K.; Ismail, A.; Budkowska, M.; Salata, D.; Dolegowska, B.; Kucia, M.; Ratajczak, J.; Ratajczak, M.Z. Novel evidence that crosstalk between the complement, coagulation and fibrinolysis proteolytic cascades is involved in mobilization of hematopoietic stem/progenitor cells (HSPCs). Leukemia 2014, 28, 2148–2154. [Google Scholar] [CrossRef] [Green Version]
- Borkowska, S.; Suszynska, M.; Wysoczynski, M.; Ratajczak, M.Z. Mobilization studies in C3-deficient mice unravel the involvement of a novel crosstalk between the coagulation and complement cascades in mobilization of hematopoietic stem/progenitor cells (HSPCs). Leukemia 2013, 27, 1928–1930. [Google Scholar] [CrossRef] [Green Version]
- Bryder, D.; Rossi, D.J.; Weissman, I.L. Hematopoietic stem cells. The paradigmatic tissue-specific stem cell. Am. J. Pathol. 2006, 169, 338–346. [Google Scholar] [CrossRef] [Green Version]
- Rossi, D.J.; Bryder, D.; Weissman, I.L. Hematopoietic stem cells aging: Mechanism and consequence. Exp. Gerontol. 2007, 42, 385–390. [Google Scholar] [CrossRef] [Green Version]
- Wahlestedt, M.; Pronk, C.J.; Bryder, D. Concise review: Hematopoietic stem cell aging and the prospects of rejuvenation. Stem Cells Transl. Med. 2015, 4, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Gallipoli, P.; Huntly, B.J.P. Novel epigenetic therapies in hematological malignancies. Current status and beyond. Semin. Cancer Biol. 2018, 51, 198–210. [Google Scholar] [CrossRef] [Green Version]
- Safdar, A.; Armstrong, D. Infections in patients with hematologic neoplasms and hematopoietic stem cell transplantation: Neutropenia, humoral, and splenic defects. Clin. Infect. Dis. 2011, 53, 798–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Świerzko, A.S.; Michalski, M.; Sokołowska, A.; Nowicki, M.; Eppa, Ł.; Szala-Poździej, A.; Mitrus, I.; Szmigielska-Kapłon, A.; Sobczyk-Kruszelnicka, M.; Michalak, K.; et al. The role of complement activating collectins and associated serine proteases in patients with hematological malignancies, receiving high-dose chemotherapy, and autologous hematopoietic stem cell transplantations (auto-HSCT). Front. Immunol. 2018, 9, 2153. [Google Scholar] [CrossRef] [PubMed]
- Tomaiuolo, R.; Ruocco, A.; Salapete, C.; Carru, C.; Baggio, G.; Franceschi, C.; Zinellu, A.; Vaupel, J.; Bellia, C.; Lo Sasso, B.; et al. Activity of mannose-binding lectin (MBL) in centenarians. Aging Cell 2012, 11, 394–400. [Google Scholar] [CrossRef] [Green Version]
- Nauta, A.J.; Raashou-Jensen, N.; Roos, A.; Daha, M.R.; Madsen, H.O.; Borrias-Essers, M.C.; Ryder, L.P.; Koch, C.; Garred, P. Mannose-binding lectin engagement with late apoptotic and necrotic cells. Eur. J. Immunol. 2003, 33, 2853–2863. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T.; Taira, S.; Kodama, N.; Matsushita, M.; Fujita, T. Mannose-binding protein recognizes glioma cells: In vitro analysis of complement activation on glioma cells via the lectin pathway. Jpn. J. Cancer Res. 1995, 86, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Zanetti, K.A.; Haznadar, M.; Welsh, J.A.; Robles, A.I.; Ryan, B.M.; McClary, A.C.; Bowman, E.D.; Goodman, J.E.; Bernig, T.; Chanock, S.J.; et al. 3′UTR and functional secretor haplotypes in mannose-binding lectin 2 are associated with increased colon cancer risk in African Americans. Cancer Res. 2012, 72, 1467–1477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henriksen, M.L.; Madsen, K.L.; Skjoedt, K.; Hansen, S. Calcium-sensitive immunoaffinity chromatography: Gentle and highly specific retrieval of a scarce plasma antigen, collectin-LK (CL-LK). J. Immunol. Methods 2014, 413, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.J.; Skjoedt, M.O.; Garred, P. Collectin-11/MASP complex formation triggers activation of the lectin complement pathway—The fifth lectin pathway initiation complex. J. Innate Immun. 2013, 5, 242–250. [Google Scholar] [CrossRef]
- Henriksen, M.L.; Brandt, J.; Iyer, S.S.; Thielens, N.M.; Hansen, S. Characterization of the interaction between collectin 11 (CL-11, CL-K1) and nucleic acids. Mol. Immunol. 2013, 56, 757–767. [Google Scholar] [CrossRef]
- Farrar, C.A.; Tran, D.; Li, K.; Wu, W.; Peng, Q.; Schwaeble, W.; Zhou, W.; Sacks, S.H. Collectin-11 detects stress-induced L-fucose pattern to trigger renal epithelial injury. J. Clin. Investig. 2016, 126, 1911–1925. [Google Scholar] [CrossRef] [Green Version]
- Nauser, C.L.; Howard, M.C.; Fanelli, G.; Farrar, C.A.; Sacks, S. Collectin-11 (CL-11) Is a major sentinel at epithelial surfaces and key pattern recognition molecule in complement-mediated ischaemic injury. Front. Immunol. 2018, 9, 2023. [Google Scholar] [CrossRef] [PubMed]
- Bayarri-Olmos, R.; Hansen, S.; Henriksen, M.L.; Storm, L.; Thiel, S.; Garred, P.; Munthe-Fog, L. Genetic variation of COLEC10 and COLEC11 and association with serum levels of collectin liver 1 (CL-L1) and collectin kidney 1 (CL-K1). PLoS ONE 2015, 10, e0114883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selman, L.; Henriksen, M.L.; Brandt, J.; Palarasah, Y.; Waters, A.; Beales, P.L.; Holmskov, U.; Jorgensen, T.J.; Nielsen, C.; Skjodt, K.; et al. An enzyme-linked immunosorbent assay (ELISA) for quantification of human collectin 11 (CL-11, CL-K1). J. Immunol. Methods 2012, 375, 182–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rooryck, C.; Diaz-Font, A.; Osborn, D.P.; Chabchoub, E.; Hernandez-Hernandez, V.; Shamseldin, H.; Kenny, J.; Waters, A.; Jenkins, D.; Kaissi, A.A.; et al. Mutations in lectin complement pathway genes COLEC11 and MASP1 cause 3MC syndrome. Nat. Genet. 2011, 43, 197–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatraman Girija, U.; Furze, C.M.; Gingras, A.R.; Yoshizaki, T.; Ohtani, K.; Marshall, J.E.; Wallis, A.K.; Schwaeble, W.J.; El-Mezgueldi, M.; Mitchell, D.A.; et al. Molecular basis of sugar recognition by collectin-K1 and the effects of mutations associated with 3MC syndrome. BMC Biol. 2015, 13, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munye, M.M.; Diaz-Font, A.; Ocaka, L.; Henriksen, M.L.; Lees, M.; Brady, A.; Jenkins, D.; Morton, J.; Hansen, S.W.; Bacchelli, C.; et al. COLEC10 is mutated in 3MC patients and regulates early craniofacial development. PLoS Genet. 2017, 13, e1006679. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, C.R.; Jensen, L.; Dagnaes-Hansen, F.; Holm, I.E.; Endo, Y.; Fujita, T.; Thiel, S.; Jensenius, J.C.; Degn, S.E. Mitochondria and the lectin pathway of complement. J. Biol. Chem. 2013, 288, 8016–8027. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.J.; Doni, A.; Romani, L.; Jurgensen, H.J.; Behrendt, N.; Mantovani, A.; Garred, P. Ficolin-1-PTX3 complex formation promotes clearance of altered self-cells and modulates IL-8 production. J. Immunol. 2013, 191, 1324–1333. [Google Scholar] [CrossRef] [Green Version]
- Hummelshoj, T.; Munthe-Fog, L.; Madsen, H.O.; Garred, P. Functional SNPs in the human ficolin (FCN) genes reveal distinct geographical patterns. Mol. Immunol. 2008, 45, 2508–2520. [Google Scholar] [CrossRef]
- Ammitzboll, C.G.; Kjaer, T.R.; Steffensen, R.; Stengaard-Pedersen, K.; Nielsen, H.J.; Thiel, S.; Bogsted, M.; Jensenius, J.C. Non-synonymous polymorphisms in the FCN1 gene determine ligand-binding ability and serum levels of M-ficolin. PLoS ONE 2012, 11, e50585. [Google Scholar] [CrossRef]
- Munthe-Fog, L.; Hummelshoj, T.; Honore, C.; Moller, M.E.; Skjoedt, M.O.; Palsgaard, I.; Borregaard, N.; Madsen, H.O.; Garred, P. Variation in FCN1 affects biosynthesis of ficolin-1 and is associated with outcome of systemic inflammation. Genes Immun. 2012, 13, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Kuraya, M.; Ming, Z.; Liu, X.; Matsushita, M.; Fujita, T. Specific binding of L-ficolin and H-ficolin to apoptotic cells leads to complement activation. Immunobiology 2005, 209, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.L.; Honore, C.; Hummelshoj, T.; Hansen, B.E.; Madsen, H.O.; Garred, P. Ficolin-2 recognizes DNA and participates in the clearance of dying host cells. Mol. Immunol. 2007, 44, 856–865. [Google Scholar] [CrossRef] [PubMed]
- Hummelshoj, T.; Munthe-Fog, L.; Madsen, H.O.; Fujita, T.; Matsushita, M.; Garred, P. Polymorphisms in the FCN2 gene determine serum variation and function of Ficolin-2. Hum. Mol. Genet. 2005, 14, 1651–1658. [Google Scholar] [CrossRef]
- Herpers, B.L.; Immink, M.M.; de Jong, B.A.; van Velzen-Blad, H.; de Jongh, B.M.; van Hannen, E.J. Coding and non-coding polymorphisms in the lectin pathway activator L-ficolin gene in 188 Dutch blood bank donors. Mol. Immunol. 2006, 43, 851–855. [Google Scholar] [CrossRef]
- Cedzynski, M.; Nuytinck, L.; Atkinson, A.P.; Swierzko, A.S.; Zeman, K.; Szemraj, J.; Szala, A.; Turner, M.L.; Kilpatrick, D.C. Extremes of L-ficolin concentration in children with recurrent infections are associated with single nucleotide polymorphisms in the FCN2 gene. Clin. Exp. Immunol. 2007, 150, 99–104. [Google Scholar] [CrossRef]
- Kilpatrick, D.C.; Swierzko, A.S.; Matsushita, M.; Domzalska-Popadiuk, I.; Borkowska-Klos, M.; Szczapa, J.; Cedzynski, M. The relationship between FCN2 genotypes and serum ficolin-2 (L-ficolin) protein concentrations from a large cohort of neonates. Hum. Immunol. 2013, 74, 864–871. [Google Scholar] [CrossRef]
- Troldborg, A.; Hansen, A.; Hansen, S.W.K.; Jensenius, J.C.; Stengaard-Pedersen, K.; Thiel, S. Lectin complement pathway proteins in healthy individuals. Clin. Exp. Immunol. 2017, 188, 138–147. [Google Scholar] [CrossRef] [Green Version]
- Honore, C.; Hummelshoj, T.; Hansen, B.E.; Madsen, H.O.; Eggleton, P.; Garred, P. The innate immune component ficolin 3 (Hakata antigen) mediates the clearance of late apoptotic cells. Arthritis Rheum. 2007, 56, 1598–1607. [Google Scholar] [CrossRef]
- Michalski, M.; Świerzko, A.S.; Sawicki, S.; Kałużyński, A.; Łukasiewicz, J.; Maciejewska, A.; Wydra, D.; Cedzyński, M. Interactions of ficolin-3 with ovarian cancer cells. Immunobiology 2019, 224, 316–324. [Google Scholar] [CrossRef]
- Jarlhelt, I.; Pilely, K.; Clausen, J.B.; Skjoedt, M.O.; Bayarri-Olmos, R.; Garred, P. Circulating ficolin-2 and ficolin-3 form heterocomplexes. J. Immunol. 2020, 204, 1919–1928. [Google Scholar] [CrossRef]
- Munthe-Fog, L.; Hummelshoj, T.; Honore, C.; Madsen, H.O.; Permin, H.; Garred, P. Immunodeficiency associated with FCN3 mutation and ficolin-3 deficiency. N. Engl. J. Med. 2009, 360, 2637–2644. [Google Scholar] [CrossRef] [PubMed]
- Michalski, M.; Świerzko, A.S.; Pągowska-Klimek, I.; Niemir, Z.I.; Mazerant, K.; Domżalska-Popadiuk, I.; Moll, M.; Cedzyński, M. Primary Ficolin-3 deficiency—Is it associated with increased susceptibility to infections? Immunobiology 2015, 220, 711–713. [Google Scholar] [CrossRef] [PubMed]
- Kjaer, T.R.; Le, L.T.M.; Pedersen, J.S.; Sander, B.; Golas, M.M.; Jensenius, J.C.; Andersen, G.R.; Thiel, S. Structural insights into the initiating complex of the lectin pathway of complement activation. Structure 2015, 23, 342–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jani, P.K.; Kajdacsi, E.; Megyeri, M.; Dobo, J.; Doleschall, Z.; Futosi, K.; Timar, C.I.; Mocsai, A.; Mako, V.; Gal, P.; et al. MASP-1 induces a unique cytokine pattern in endothelial cells: A novel link between complement system and neutrophil granulocytes. PLoS ONE 2014, 9, e87104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwaner, E.; Nemeth, Z.; Jani, P.K.; Kajdacsi, E.; Debreczeni, M.L.; Doleschall, Z.; Dobo, J.; Gal, P.; Rigo, J.; Andras, K.; et al. Transcriptome analysis of inflammation-related gene expression in endothelial cells activated by complement MASP-1. Sci. Rep. 2017, 7, 10462. [Google Scholar] [CrossRef] [Green Version]
- Debreczeni, M.L.; Nemeth, Z.; Kajdacsi, E.; Schwaner, E.; Mako, V.; Masszi, A.; Doleschall, Z.; Rigo, J.; Walter, F.R.; Deli, M.A.; et al. MASP-1 increases endothelial permeability. Front. Immunol. 2019, 10, 991. [Google Scholar] [CrossRef]
- Holers, M.V.; Borodovsky, A.; Scheinman, R.I.; Ho, N.; Ramos Ramirez, J.; Dobo, J.; Gal, P.; Lindenberger, J.; Hansen, A.G.; Desai, D.; et al. Key components of the complement lectin pathway are not only required for the development of inflammatory arthritis but also regulate the transcription of factor D. Front. Immunol. 2020, 11, 201. [Google Scholar] [CrossRef] [Green Version]
- Pihl, R.; Jensen, L.; Hansen, A.G.; Thogersen, I.B.; Andres, S.; Dagnes-Hansen, F.; Oexle, K.; Enghild, J.J.; Thiel, S. Analysis of factor D isoforms in Malpuech-Michels-Mingarelli-Carnevale patients highlights the role of MASP-3 as a maturase in the alternative pathway of complement. J. Immunol. 2017, 199, 2158–2170. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, M.; Machida, T.; Ishida, Y.; Ogata, Y.; Omori, T.; Takasumi, M.; Endo, Y.; Suzuki, T.; Sekimata, M.; Homma, Y.; et al. Cutting edge: Role of MASP-3 in the physiological activation of factor D of the alternative complement pathway. J. Immunol. 2019, 203, 1411–1416. [Google Scholar] [CrossRef]
- Mortensen, S.A.; Skov, L.L.; Kjaer-Sorensen, K.; Hansen, A.G.; Hansen, S.; Dagnaes-Hansen, F.; Jensenius, J.C.; Oxvig, C.; Thiel, S.; Degn, S.E. Endogenous natural complement inhibitor regulates cardiac development. J. Immunol. 2017, 198, 3118–3126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, G.; Madsen, H.O.; Garred, P. A novel mannose-binding lectin-associated serine protease 1/3 gene variant. Scand. J. Immunol. 2007, 65, 430–434. [Google Scholar] [CrossRef] [PubMed]
- Sirmaci, A.; Walsh, T.; Akay, H.; Spiliopoulos, M.; Bayezit Sakalar, Y.; Hasanefendioglu-Bayrak, A.; Duman, D.; Farooq, A.; King, M.-C.; Tekin, M. MASP1 mutations in patients with facial, umbilical, coccygeal, and auditory findings of Carnevale, Malpuech, OSA, and Michels syndromes. Am. J. Hum. Genet. 2010, 87, 679–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atik, T.; Koparir, A.; Bademci, G.; Foster, J., 2nd; Altunoglu, U.; Mutlu, G.Y.; Bowdin, S.; Elcioglu, N.; Tayfun, G.A.; Atik, S.S.; et al. Novel MASP1 mutations are associated with an expanded phenotype in 3MC1 syndrome. Orphanet J. Rare Dis. 2015, 10, 128. [Google Scholar] [CrossRef] [Green Version]
- Graul-Neumann, L.M.; Mensah, M.A.; Klopocki, E.; Uebe, S.; Ekici, A.B.; Thiel, C.T.; Reis, A.; Zweier, C. Biallelic intragenic deletion in MASP1 in an adult female with 3MC syndrome. Eur. J. Med. Genet. 2018, 61, 363–368. [Google Scholar] [CrossRef]
- Basdemirci, M.; Sen, A.; Ceylaner, S. Novel mutation in MASP1 gene in a new family with 3MC syndrome. Clin. Dysmorphol. 2019, 28, 91–93. [Google Scholar] [CrossRef]
- Haerynck, F.; Van Steen, K.; Cattaert, T.; Loeys, B.; Van Daele, S.; Schelstraete, P.; Claes, K.; Van Thielen, M.; De Canck, I.; Mahachie John, J.M.; et al. Polymorphisms in the lectin pathway genes as a possible cause of early chronic Pseudomonas aeruginosa colonization in cystic fibrosis patients. Hum. Immunol. 2012, 73, 1175–1183. [Google Scholar] [CrossRef]
- Sokolowska, A.; Szala, A.; Swierzko, A.S.; Kozinska, M.; Niemiec, T.; Blachnio, M.; Augustynowicz-Kopec, E.; Dziadek, J.; Cedzynski, M. Mannan-binding lectin-associated serine protease-2 (MASP-2) deficiency in two patients with pulmonary tuberculosis and one healthy control. Cell. Mol. Immunol. 2015, 12, 119–121. [Google Scholar] [CrossRef] [Green Version]
- Tulio, S.; Faucz, F.R.; Werneck, R.I.; Olandoski, M.; Alexandre, R.B.; Boldt, A.B.; Pedroso, M.L.; de Messias-Reason, I.J. MASP2 gene polymorphism is associated with susceptibility to hepatitis C virus infection. Hum. Immunol. 2011, 72, 912–915. [Google Scholar] [CrossRef]
- Degn, S.E.; Thiel, S.; Nielsen, O.; Hansen, A.G.; Steffensen, R.; Jensenius, J.C. MAp19, the alternative splice product of the MASP2 gene. J. Immunol. Methods 2011, 373, 89–101. [Google Scholar] [CrossRef]
- Sokołowska, A.; Świerzko, A.S.; Gajek, G.; Gołos, A.; Michalski, M.; Nowicki, M.; Szala-Poździej, A.; Wolska-Washer, A.; Brzezińska, O.; Wierzbowska, A.; et al. Associations of ficolins and mannose-binding lectin with acute myeloid leukaemia in adults. Sci. Rep. 2020, 10, 10561. [Google Scholar] [CrossRef] [PubMed]
- Adamiak, M.; Abdel-Latif, A.; Ratajczak, M.Z. Mannan binding lectin triggers mobilization of hematopoietic cells. Oncotarget 2017, 43, 73368–73369. [Google Scholar] [CrossRef] [PubMed]
- Adamiak, M.; Abdelbaset-Ismail, A.; Suszynska, M.; Abdel-Latif, A.; Ratajczak, J.; Ratajczak, M.Z. Novel evidence that the mannan-binding lectin pathway of complement activation plays a pivotal role in triggering mobilization of hematopoietic stem/progenitor cells by activation of both the complement and coagulation cascades. Leukemia 2017, 31, 262–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratajczak, M.Z.; Adamiak, M.; Kucia, M.; Tse, W.; Ratajczak, J.; Wiktor-Jędrzejczak, W. The emerging link between the complement cascade and purinergic signaling in stress hematopoiesis. Front. Immunol. 2018, 9, 1295. [Google Scholar] [CrossRef] [Green Version]
- Khayr, W.; Haddad, R.Y.; Noor, S.A. Infections in hematological malignancies. Dis. Mon. 2012, 58, 239–249. [Google Scholar] [CrossRef]
- Ruhnke, M.; Arnold, R.; Gastmeier, P. Infection control issues in patients with haematological malignancies in the era of multidrug-resistant bacteria. Lancet Oncol. 2014, 15, e606–e609. [Google Scholar] [CrossRef]
- Wójtowicz, A.; Bochud, P.-E. Risk stratification and immunogenetic risk for infections following stem cell transplantation. Virulence 2016, 7, 917–929. [Google Scholar] [CrossRef] [Green Version]
- Peterslund, N.A.; Koch, C.; Jensenius, J.C.; Thiel, S. Association between deficiency of mannose-binding lectin and severe infections after chemotherapy. Lancet 2001, 358, 637–638. [Google Scholar] [CrossRef]
- Horiuchi, T.; Gondo, H.; Miyagawa, H.; Otsuka, J.; Inaba, S.; Nagafuji, K.; Takase, K.; Tsukamoto, H.; Koyama, T.; Mitoma, H.; et al. Association of MBL gene polymorphisms with major bacterial infection in patients treated with high-dose chemotherapy and autologous PBSCT. Genes Immun. 2005, 6, 162–166. [Google Scholar] [CrossRef] [Green Version]
- Moreto, A.; Farinas-Alvarez, C.; Puente, M.; Ocejo-Vinyals, J.G.; Sanchez-Velasco, P.; Horcajada, J.P.; Batlle, A.; Montes, C.; Santos, F.; Code, E.; et al. Mannose-binding lectin gene variants and infections in patients receiving autologous stem cell transplantation. BMC Immunol. 2014, 15, 17. [Google Scholar] [CrossRef] [Green Version]
- Molle, I.; Peterslund, N.A.; Thiel, S.; Steffensen, R. MBL2 polymorphism and risk of severe infections in multiple myeloma patients receiving high-dose melphalan and autologous stem cell transplantation. Bone Marrow Transplant. 2006, 38, 555–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molle, I.; Steffensen, R.; Thiel, S.; Peterslund, N.A. Chemotherapy-related infections in patients with multiple myeloma: Associations with mannan-binding lectin genotypes. Eur. J. Hematol. 2006, 77, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Eleutherakis-Papaiakovou, E.; Dimopoulos, M.A.; Kastritis, E.; Christoulas, D.; Roussou, M.; Migkou, M.; Gavriatopoulou, M.; Fotiou, D.; Panagiotidis, I.; Ziogas, D.C.; et al. Low circulating mannan-binding lectin levels correlate with increased frequency and severity of febrile episodes in myeloma patients who undergo ASCT and do not receive antibiotic prophylaxis. Bone Marrow Transplant. 2017, 52, 1537–1542. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, D.C.; McLintock, L.A.; Allan, E.K.; Kopland, M.; Fujita, T.; Jordanides, N.E.; Koch, C.; Matsushita, M.; Shiraki, H.; Stewart, K.; et al. No strong relationship between mannan binding lectin or plasma ficolins and chemotherapy-related infections. Clin. Exp. Immunol. 2003, 134, 279–284. [Google Scholar] [CrossRef]
- Bergmann, O.J.; Christiansen, M.; Laursen, I.; Bang, P.; Hansen, N.E.; Ellegaard, J.; Koch, C.; Andersen, V. Low levels of mannose-binding lectin do not affect occurrence of severe infections or duration of fever in acute myeloid leukaemia during remission induction therapy. Eur. J. Haematol. 2003, 70, 91–97. [Google Scholar] [CrossRef]
- Wong, M.; Ohrmalm, L.; Brollden, K.; Aust, C.; Hibberd, M.; Tolfvenstam, T. Mannose-binding lectin 2 polymorphisms do not influence frequency and type of infection in adults with chemotherapy induced neutropaenia. PLoS ONE 2012, 7, e30819. [Google Scholar] [CrossRef]
- Osthoff, M.; Rovo, A.; Stern, M.; Danner, D.; Gratwohl, A.; Tichelli, A.; Trendelenburg, M. Mannose-binding lectin levels and major infections in a cohort of very long-term survivors after allogeneic stem cell transplantation. Haematologica 2010, 95, 1389–1396. [Google Scholar] [CrossRef] [Green Version]
- Mullighan, C.G.; Heatley, S.L.; Danner, S.; Dean, M.M.; Doherty, K.; Hahn, U.; Bradstock, K.F.; Minchinton, R.; Schwarer, A.P.; Szer, J.; et al. Mannose-binding lectin status is associated with risk of major infection following myeloablative sibiling allogeneic hemopoietic stem cell transplantation. Blood 2008, 112, 2120–2128. [Google Scholar] [CrossRef] [Green Version]
- Radnay, Z.B.; Udvardy, M.; Papp, M.; Harsfalvi, J.; Rejto, L.; Pal, I.; Illes, A.; Kiss, A. Evaluation of mannose-binding lectin is a useful approach to predict the risk of infectious complications following autologous hematopoietic stem cell transplantation. Transplant. Proc. 2016, 48, 3397–3405. [Google Scholar] [CrossRef]
- Schlapbach, L.J.; Thiel, S.; Aebi, C.; Hirt, A.; Leibundgut, K.; Jensenius, J.C.; Ammann, R.A. M-ficolin in children with cancer. Immunobiology 2011, 216, 633–638. [Google Scholar] [CrossRef]
- Świerzko, A.S.; Michalski, M.; Sokołowska, A.; Nowicki, M.; Szala-Poździej, A.; Eppa, Ł.; Mitrus, I.; Szmigielska-Kapłon, A.; Sobczyk-Kruszelnicka, M.; Michalak, K.; et al. Associations of ficolins with haematological malignancies in patients receiving high-dose chemotherapy and autologous haematopoietic stem cell transplantations (auto-HSCT). Front. Immunol. 2020, 10, 3097. [Google Scholar] [CrossRef] [PubMed]
- Ameye, L.; Paesmans, M.; Thiel, S.; Jensenius, J.C.; Aoun, M. M-ficolin levels are associated with the occurrence of severe infections in patients with haematological cancer undergoing chemotherapy. Clin. Exp. Immunol. 2012, 167, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Pana, Z.D.; Samarah, F.; Papi, R.; Antachopoulus, C.; Papageorgiou, T.; Farmaki, E.; Hatzipantelis, E.; Tragiannidis, A.; Vavatsi-Christaki, N.; Kyriakidis, D.; et al. Mannose-binding lectin and ficolin-2 polymorphisms are associated with increased risk for bacterial infections in children with B acute lymphoblastic leukemia. Pediatr. Blood Cancer 2014, 61, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Islak Mutcali, S.; Saltoglu, N.; Balkan, I.I.; Ozaras, R.; Yemisen, M.; Mete, B.; Ozturk, R.; Ongoren, S.; Baslar, Z.; Aydin, Y.; et al. Early changes of mannose-binding lectin, H-ficolin, and procalcitonin in patients with febrile neutropenia: A prospective observational study. Turk. J. Hematol. 2016, 33, 304–310. [Google Scholar] [CrossRef]
- Schlapbach, L.J.; Aebi, C.; Hansen, A.G.; Hirt, A.; Jensenius, J.C.; Ammann, R. H-ficolin serum concentration and susceptibility to fever and neutropenia in paediatric cancer patients. Clin. Exp. Immunol. 2009, 157, 83–89. [Google Scholar] [CrossRef]
- Fisch, U.; Zehnder, A.; Hirt, A.; Niggli, F.; Simon, A.; Ozsahin, H.; Schlapbach, L.; Ammann, R. Mannan-binding lectin (MBL) and MBL-associated serine protease-2 in children with cancer. Swiss Med. Wkly. 2011, 141, w13191. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Bassig, B.A.; Xu, J.; Zheng, T.; Zhang, Y.; Berndt, S.I.; Holford, T.R.; Hosgood, H.D., 3rd; Leaderer, B.; Yeager, M.; et al. Polymorphisms in pattern-recognition genes in the innate immunity system and risk of non-Hodgkin lymphoma. Environ. Mol. Mutagenesis 2013, 54, 72–77. [Google Scholar] [CrossRef]
- Charbonneau, B.; Maurer, M.J.; Fredericksen, Z.S.; Zent, C.S.; Link, B.K.; Novak, A.J.; Ansell, S.M.; Weiner, G.J.; Wang, A.H.; Witzig, T.E.; et al. Germline variation in complement genes and event-free survival in follicular and diffuse large B-cell lymphoma. Am. J. Hematol. 2012, 87, 880–885. [Google Scholar] [CrossRef] [Green Version]
- Zehnder, A.; Fisch, U.; Hirt, A.; Niggli, F.K.; Simon, A.; Ozsahin, H.; Schlapbach, L.J.; Ammann, R.A. Prognosis in pediatric hematologic malignancies is associated with serum concentration of mannose-binding lectin-associated serine protease-2 (MASP-2). Pediatr. Blood Cancer 2009, 53, 53–57. [Google Scholar] [CrossRef]
- Swierzko, A.S.; Cedzynski, M.; Domzalska-Popadiuk, I.; MacDonald, S.L.; Borkowska-Klos, M.; Atkinson, A.P.M.; Szala, A.; Jopek, A.; Jensenius, J.C.; Kawakami, M.; et al. Mannan-binding lectin-associated serine protease-2 (MASP-2) in a large cohort of neonates and its clinical associations. Mol. Immunol. 2009, 46, 1696–1701. [Google Scholar] [CrossRef]


{kind=link}
| Family | Protein | Ligands/Substrates | Gene and Its Chromosomal Location | Primary Sites of Expression | References |
|---|---|---|---|---|---|
| Collectins | MBL | D-Man D-GlcNAc L-Fuc | MBL2 10q11.2-q21 | hepatocytes | [1,2,5] |
| CL-10 (CL-L1) | D-Man L-Fuc D-Fuc D-Gal D-GlcNAc | COLEC10 8q24.12 | hepatocytes | [6,7,8] | |
| CL-11 (CL-K1) | D-Man L-Fuc D-ManNAc | COLEC11 2p25.3 | Kidney adrenal gland hepatocytes | [8,9,10] | |
| Ficolins | Ficolin-1 (M-ficolin) | D-GlcNAc D-ManNAc D-GalNAc Sialic acid | FCN1 9q34.3 | bone marrow, monocytes, neutrophils | [1,2,11] |
| Ficolin-2 (L-ficolin) | D-GlcNAc D-GalNAc D-ManNAc D-Gal N-acetylated cysteine acetylocholine | FCN2 9q34.3 | hepatocytes | [4,11,12] | |
| Ficolin-3 (H-ficolin) | D-GlcNAc D-GalNAc D-Gal D-Fuc L-Fuc | FCN3 1p36.11 | hepatocytes, alveolar type II pneumocytes, ciliated bronchial cells | [1,2,11] | |
| MASP | MASP-1 | MASP-2 C2 Fibrinogen factor XIII TAFI PAR-4 kininogen | MASP1/3 3q27-q28 | hepatocytes. | [13,14,15,16] |
| MASP-3 | Pro-D IGFPB-5 | MASP1/3 3q27-q28 | hepatocytes, cervix | [14,17,18] | |
| MASP-2 | C4 C2 prothrombin kininogen | MASP2 1p36.3–p36.2 | hepatocytes | [16,19,20] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cedzyński, M.; Świerzko, A.S. Components of the Lectin Pathway of Complement in Haematologic Malignancies. Cancers 2020, 12, 1792. https://doi.org/10.3390/cancers12071792
Cedzyński M, Świerzko AS. Components of the Lectin Pathway of Complement in Haematologic Malignancies. Cancers. 2020; 12(7):1792. https://doi.org/10.3390/cancers12071792
Chicago/Turabian StyleCedzyński, Maciej, and Anna S. Świerzko. 2020. "Components of the Lectin Pathway of Complement in Haematologic Malignancies" Cancers 12, no. 7: 1792. https://doi.org/10.3390/cancers12071792
APA StyleCedzyński, M., & Świerzko, A. S. (2020). Components of the Lectin Pathway of Complement in Haematologic Malignancies. Cancers, 12(7), 1792. https://doi.org/10.3390/cancers12071792