Genetic and Molecular Basis of Heterogeneous NK Cell Responses against Acute Leukemia

 ,
,
Abstract
:
1. Introduction
2. Results
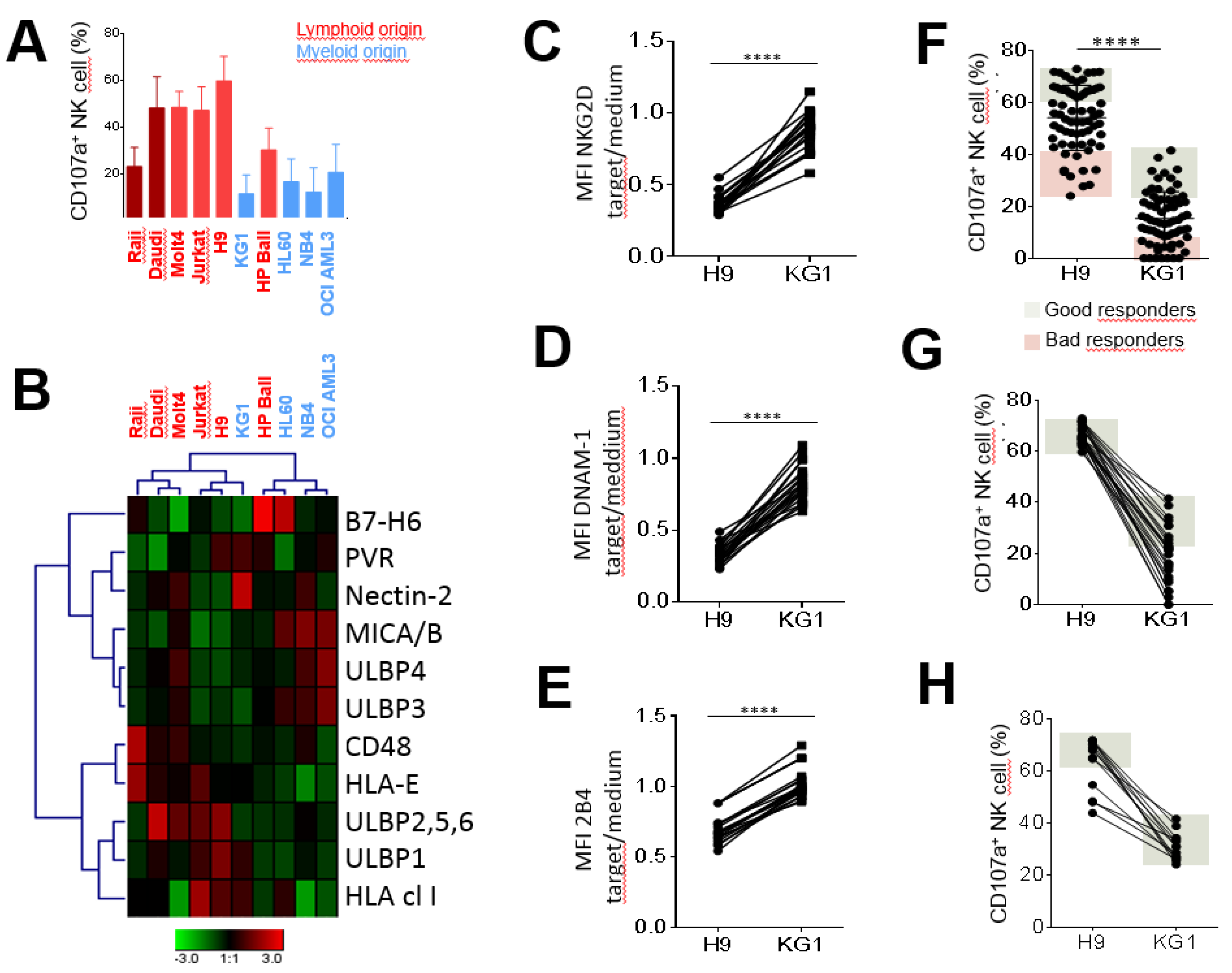
2.1. Lymphoid Cell Lines More Strongly Stimulated NK Cell Degranulation than Myeloid Cell Lines
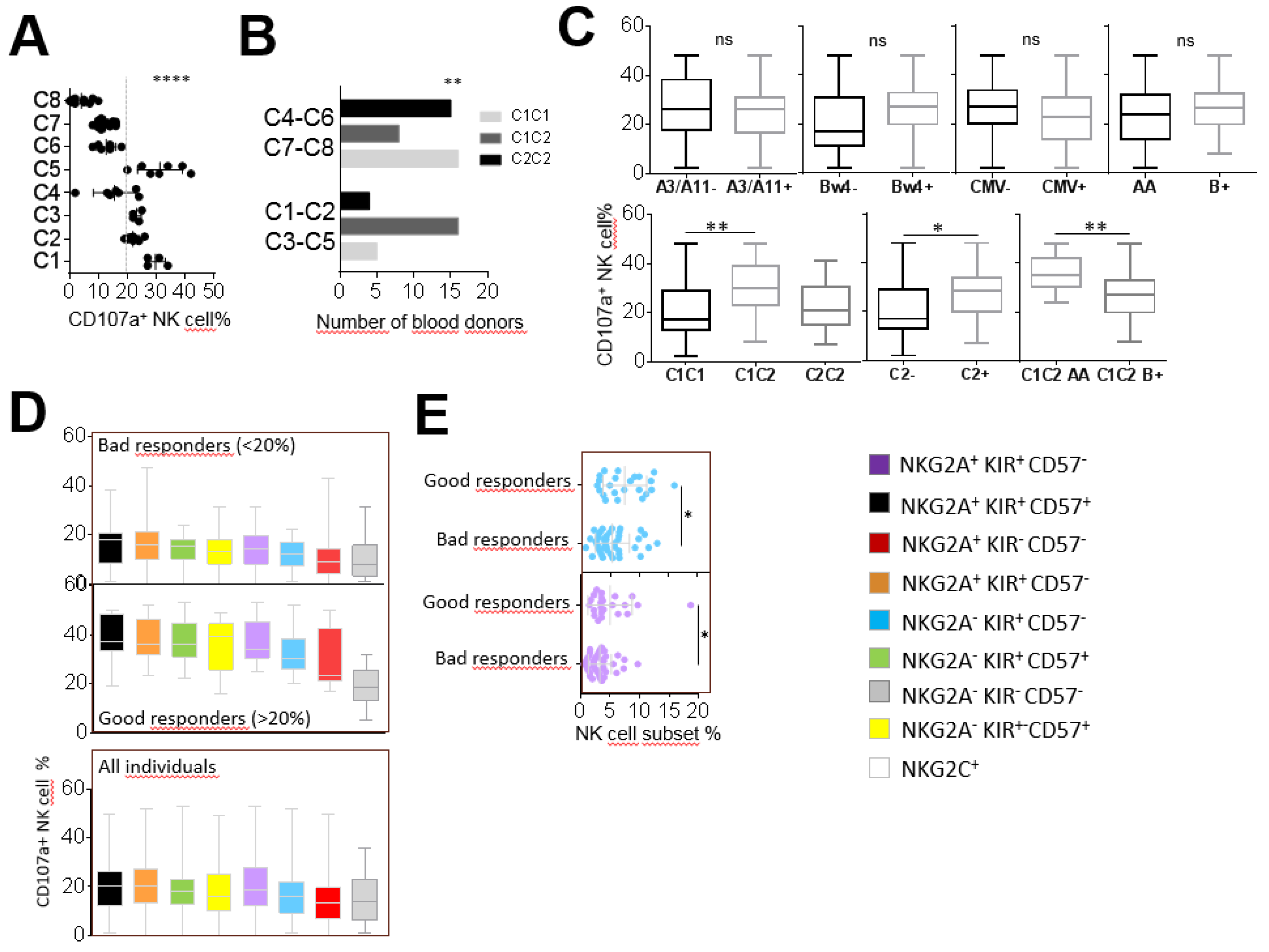
2.2. Inter-Individual Diversity of NK Cell Responses against Leukemia Target Cells
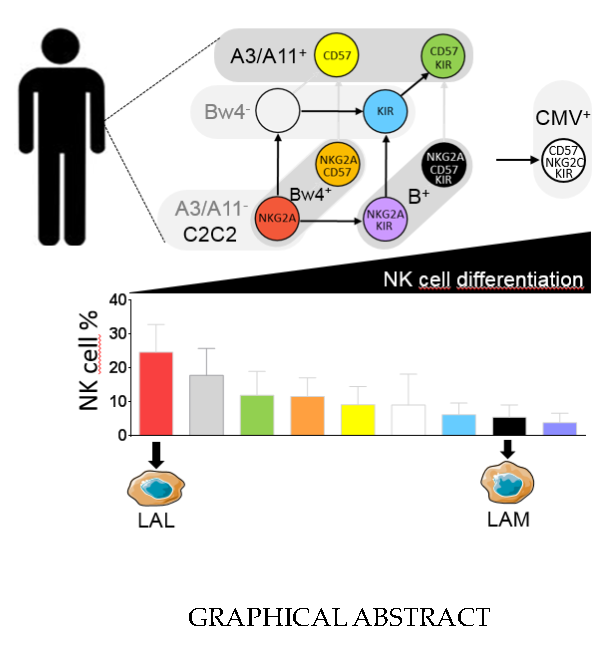
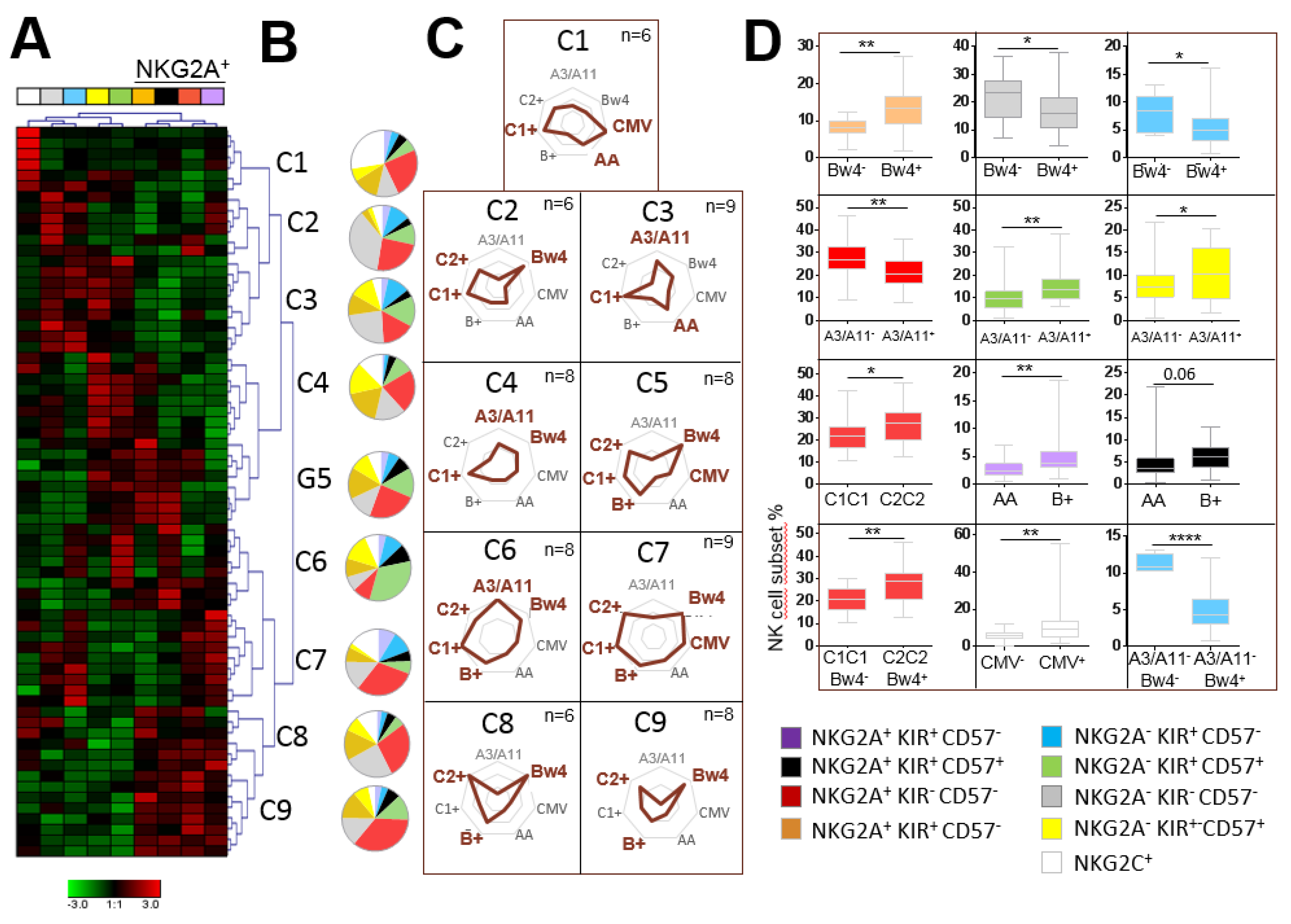
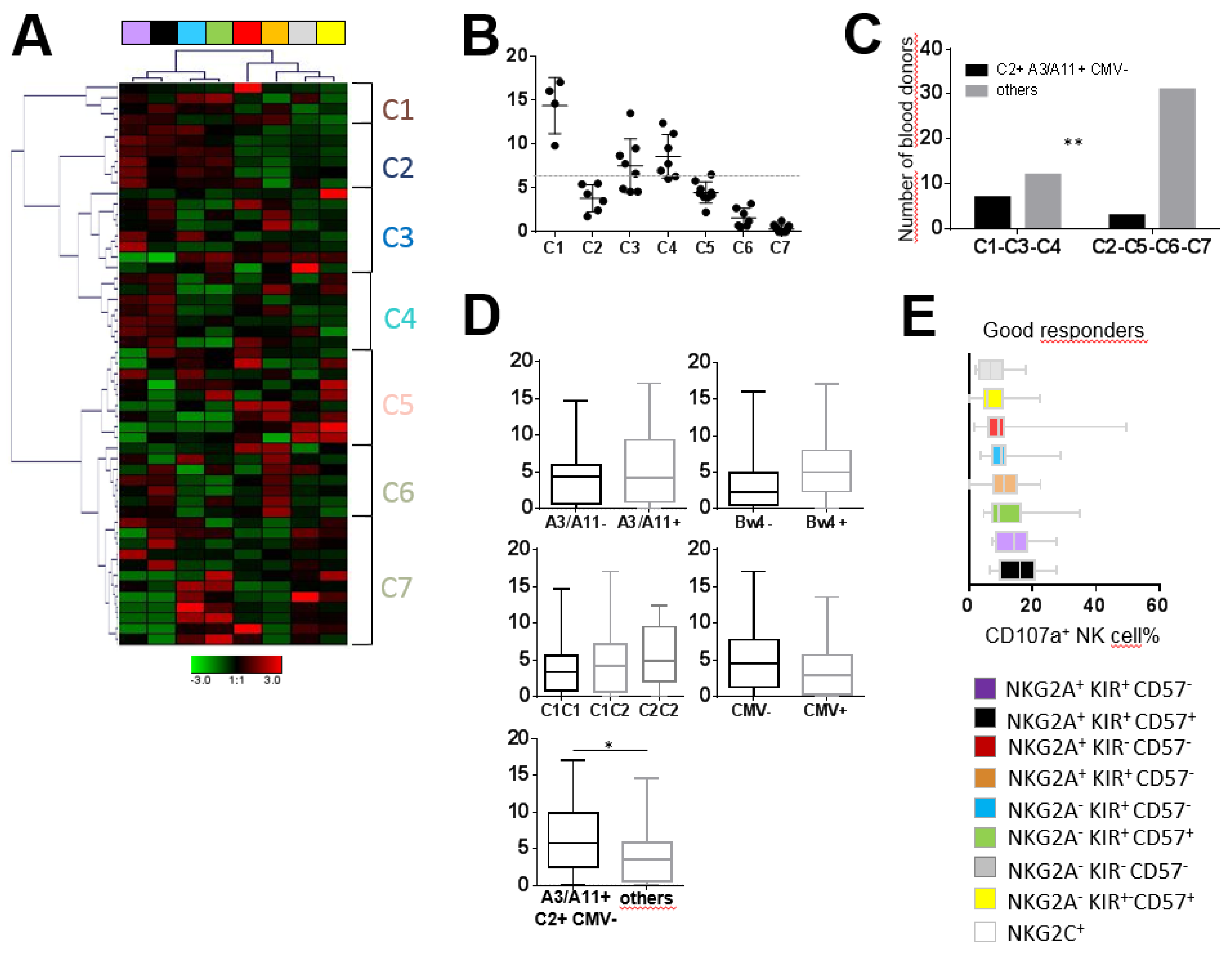
2.3. The Significant Impact of Immunogenetic HLA/KIR Polymorphisms and CMV Status on the Phenotypic Structuration of the NK Cell Repertoire
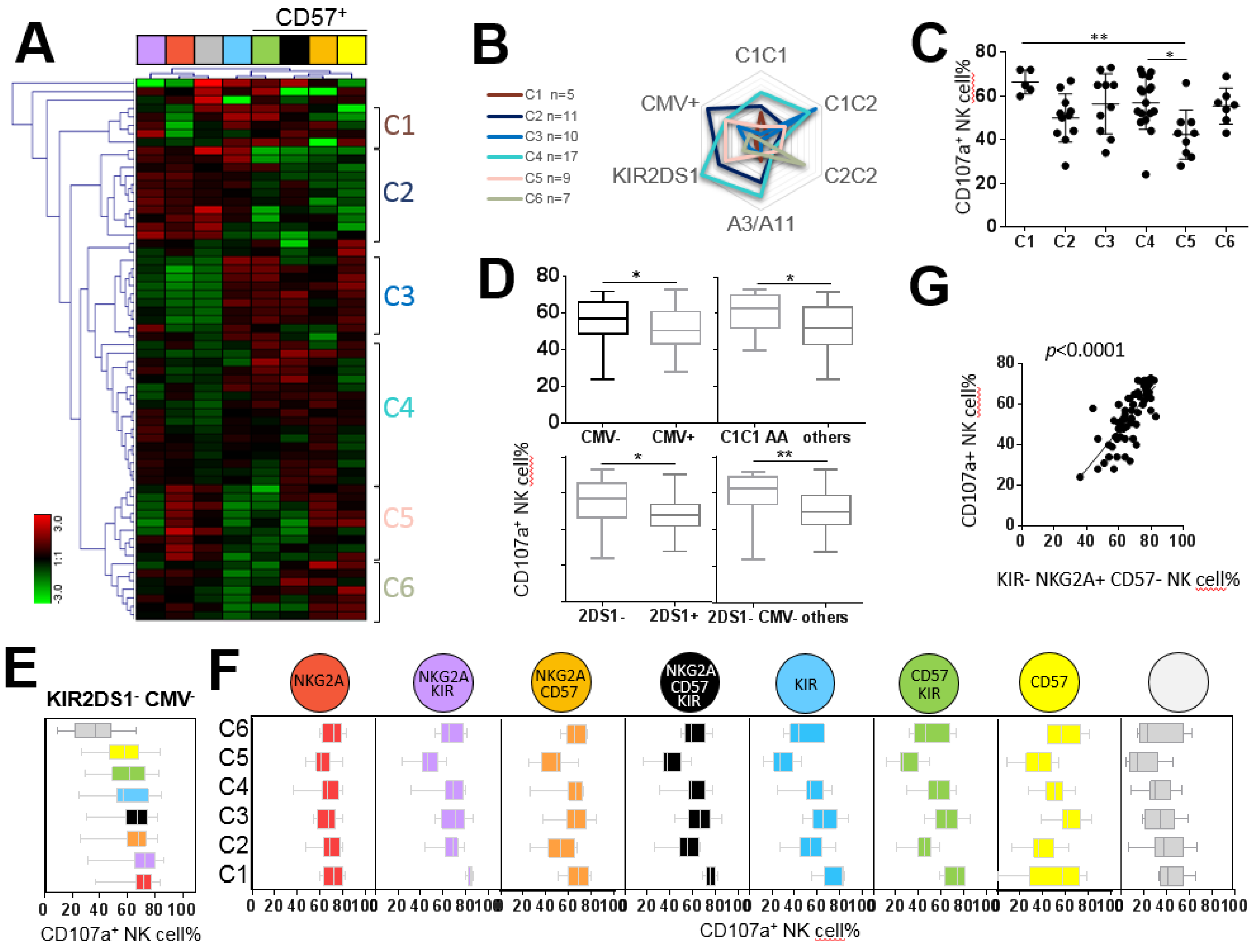
2.4. NKG2A+ NK Cell Subsets are the Most Efficient against ALL Target Cells
2.5. KIR+ and CD57+ NK Cell Subsets are the Most Efficient against AML Target Cells
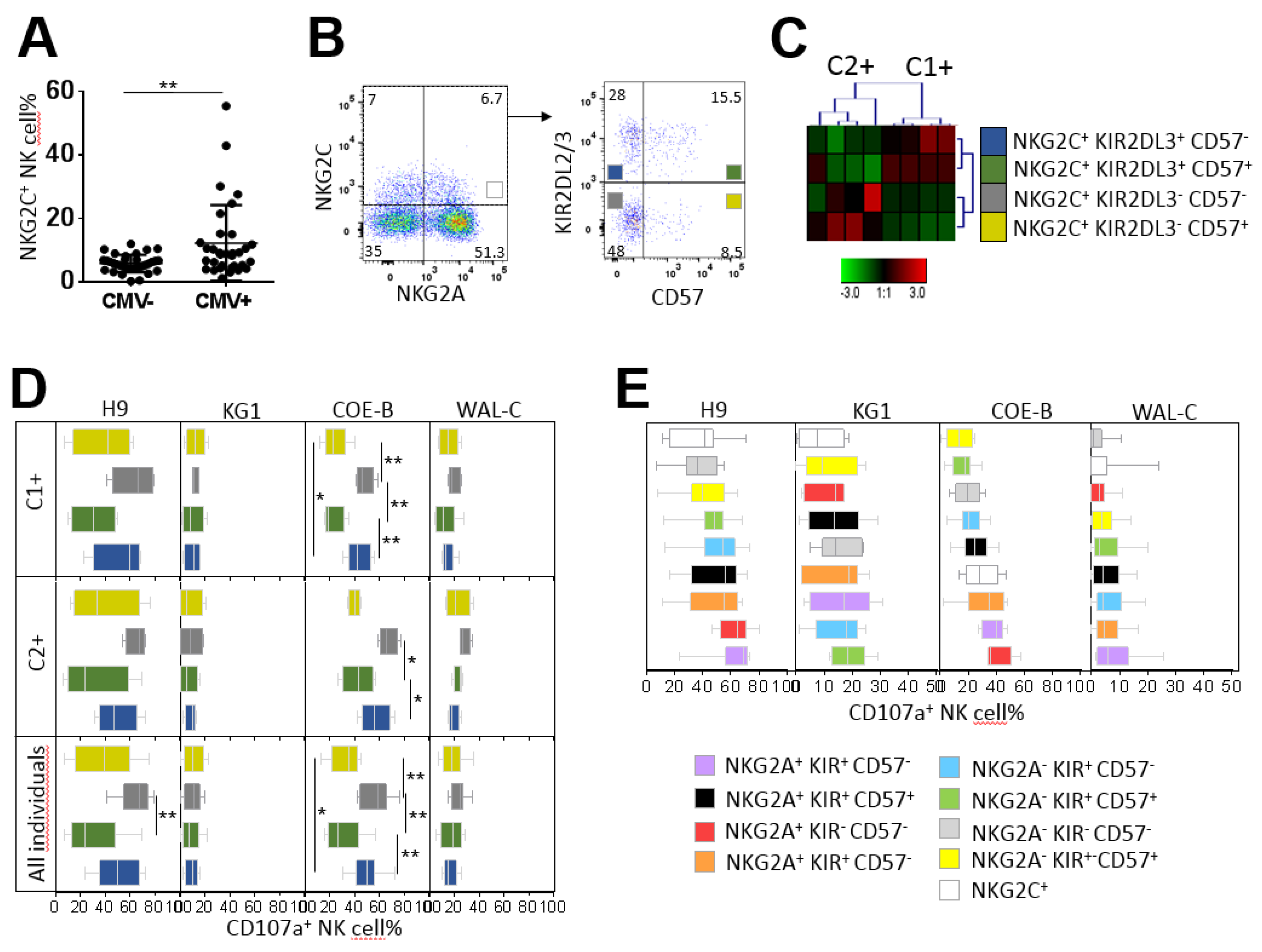
2.6. Hyporesponsiveness of CMV-Driven NKG2C+ NK Cell Subsets against Leukemia Target Cells
3. Discussion
4. Materials and Methods
4.1. Peripheral Blood Mononuclear Cells (PBMCs), Cell Lines and Primary Target Cells
4.2. HLA and KIR Genotyping
4.3. HCMV Serology
4.4. Phenotypic Analysis of NK Cells, Cell Lines and Primary Leukemic Cells by Flow Cytometry
4.5. CD107a Mobilization Assay Detected by Flow Cytometry
4.6. Hierarchical Clustering Analysis of NK Cell Phenotype and Degranulation
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
A.1. HLA and KIR Genotyping
A.2. NK Cell Phenotypic Analysis by Flow Cytometry
A.3. Phenotypic Analysis of Cell Lines and Primary Leukemic Cells by Flow Cytometry
References
- Iannello, A.; Thompson, T.W.; Ardolino, M.; Marcus, A.; Raulet, D.H. Immunosurveillance and immunotherapy of tumors by innate immune cells. Curr. Opin. Immunol. 2016, 38, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Waggoner, S.N.; Reighard, S.D.; Gyurova, I.E.; Cranert, S.A.; Mahl, S.E.; Karmele, E.P.; McNally, J.P.; Moran, M.T.; Brooks, T.R.; Yaqoob, F.; et al. Roles of natural killer cells in antiviral immunity. Curr. Opin. Virol. 2016, 16, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Kärre, K. MHC gene control of the natural killer system at the level of the target and the host. Semin. Cancer Biol. 1991, 2, 295–309. [Google Scholar] [PubMed]
- Anfossi, N.; André, P.; Guia, S.; Falk, C.S.; Roetynck, S.; Stewart, C.A.; Breso, V.; Frassati, C.; Reviron, D.; Middleton, D.; et al. Human NK cell education by inhibitory receptors for MHC class I. Immunity 2006, 25, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Boudreau, J.E.; Hsu, K.C. Natural killer cell education in human health and disease. Curr. Opin. Immunol. 2018, 50, 102–111. [Google Scholar] [CrossRef]
- Braud, V.M.; Allan, D.S.J.; O’Callaghan, C.A.; Söderström, K.; D’Andrea, A.; Ogg, G.S.; Lazetic, S.; Young, N.T.; Bell, J.I.; Phillips, J.H.; et al. HLA-E binds to natural killer cell receptors CD94/NKG2A, B and C. Nature 1998, 391, 795–799. [Google Scholar] [CrossRef] [PubMed]
- Del Zotto, G.; Marcenaro, E.; Vacca, P.; Sivori, S.; Pende, D.; Della Chiesa, M.; Moretta, F.; Ingegnere, T.; Mingari, M.C.; Moretta, A.; et al. Markers and function of human NK cells in normal and pathological conditions: Markers and Function Of Human Nk Cells. Cytometry B Clin. Cytom. 2017, 92, 100–114. [Google Scholar] [CrossRef] [Green Version]
- Chiossone, L.; Vienne, M.; Kerdiles, Y.M.; Vivier, E. Natural killer cell immunotherapies against cancer: Checkpoint inhibitors and more. Semin. Immunol. 2017, 31, 55–63. [Google Scholar] [CrossRef]
- Gaggero, S.; Bruschi, M.; Petretto, A.; Parodi, M.; Del Zotto, G.; Lavarello, C.; Prato, C.; Santucci, L.; Barbuto, A.; Bottino, C.; et al. Nidogen-1 is a novel extracellular ligand for the NKp44 activating receptor. Oncoimmunology 2018, 7, e1470730. [Google Scholar] [CrossRef]
- Niehrs, A.; Garcia-Beltran, W.F.; Norman, P.J.; Watson, G.M.; Hölzemer, A.; Chapel, A.; Richert, L.; Pommerening-Röser, A.; Körner, C.; Ozawa, M.; et al. A subset of HLA-DP molecules serve as ligands for the natural cytotoxicity receptor NKp44. Nat. Immunol. 2019, 20, 1129–1137. [Google Scholar] [CrossRef]
- Barrow, A.D.; Edeling, M.A.; Trifonov, V.; Luo, J.; Goyal, P.; Bohl, B.; Bando, J.K.; Kim, A.H.; Walker, J.; Andahazy, M.; et al. Natural Killer Cells Control Tumor Growth by Sensing a Growth Factor. Cell 2018, 172, 534–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastaglio, S.; Wong, E.; Perera, T.; Ripley, J.; Blombery, P.; Smyth, M.J.; Koldej, R.; Ritchie, D. Natural killer receptor ligand expression on acute myeloid leukemia impacts survival and relapse after chemotherapy. Blood Adv. 2018, 2, 335–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pende, D.; Spaggiari, G.M.; Marcenaro, S.; Martini, S.; Rivera, P.; Capobianco, A.; Falco, M.; Lanino, E.; Pierri, I.; Zambello, R.; et al. Analysis of the receptor-ligand interactions in the natural killer-mediated lysis of freshly isolated myeloid or lymphoblastic leukemias: Evidence for the involvement of the Poliovirus receptor (CD155) and Nectin-2 (CD112). Blood 2005, 105, 2066–2073. [Google Scholar] [CrossRef] [PubMed]
- Willem, C.; Makanga, D.R.; Guillaume, T.; Maniangou, B.; Legrand, N.; Gagne, K.; Peterlin, P.; Garnier, A.; Béné, M.C.; Cesbron, A.; et al. Impact of KIR/HLA Incompatibilities on NK Cell Reconstitution and Clinical Outcome after T Cell–Replete Haploidentical Hematopoietic Stem Cell Transplantation with Posttransplant Cyclophosphamide. J. Immunol. 2019, 202, 2141–2152. [Google Scholar] [CrossRef] [Green Version]
- Parham, P. Co-evolution of lymphocyte receptors with MHC class I. Immunol. Rev. 2015, 267, 1–5. [Google Scholar] [CrossRef]
- Björkström, N.K.; Riese, P.; Heuts, F.; Andersson, S.; Fauriat, C.; Ivarsson, M.A.; Björklund, A.T.; Flodström-Tullberg, M.; Michaëlsson, J.; Rottenberg, M.E.; et al. Expression patterns of NKG2A, KIR, and CD57 define a process of CD56dim NK-cell differentiation uncoupled from NK-cell education. Blood 2010, 116, 3853–3864. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, C.M.; White, M.J.; Goodier, M.R.; Riley, E.M. Functional Significance of CD57 Expression on Human NK Cells and Relevance to Disease. Front. Immunol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Pfefferle, A.; Jacobs, B.; Netskar, H.; Ask, E.H.; Lorenz, S.; Clancy, T.; Goodridge, J.P.; Sohlberg, E.; Malmberg, K.-J. Intra-lineage Plasticity and Functional Reprogramming Maintain Natural Killer Cell Repertoire Diversity. Cell Rep. 2019, 29, 2284–2294.e4. [Google Scholar] [CrossRef] [Green Version]
- Cichocki, F.; Grzywacz, B.; Miller, J.S. Human NK Cell Development: One Road or Many? Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Béziat, V.; Liu, L.L.; Malmberg, J.-A.; Ivarsson, M.A.; Sohlberg, E.; Björklund, A.T.; Retière, C.; Sverremark-Ekström, E.; Traherne, J.; Ljungman, P.; et al. NK cell responses to cytomegalovirus infection lead to stable imprints in the human KIR repertoire and involve activating KIRs. Blood 2013, 121, 2678–2688. [Google Scholar] [CrossRef]
- Della Chiesa, M.; Muccio, L.; Moretta, A. CMV induces rapid NK cell maturation in HSCT recipients. Immunol. Lett. 2013, 155, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Foley, B.; Cooley, S.; Verneris, M.R.; Curtsinger, J.; Luo, X.; Waller, E.K.; Anasetti, C.; Weisdorf, D.; Miller, J.S. Human Cytomegalovirus (CMV)-Induced Memory-like NKG2C+ NK Cells Are Transplantable and Expand In Vivo in Response to Recipient CMV Antigen. J. Immunol. 2012, 189, 5082–5088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djaoud, Z.; David, G.; Bressollette, C.; Willem, C.; Rettman, P.; Gagne, K.; Legrand, N.; Mehlal, S.; Cesbron, A.; Imbert-Marcille, B.-M.; et al. Amplified NKG2C+ NK Cells in Cytomegalovirus (CMV) Infection Preferentially Express Killer Cell Ig-like Receptor 2DL: Functional Impact in Controlling CMV-Infected Dendritic Cells. J. Immunol. 2013, 191, 2708–2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, Q.; Rückert, T.; Borst, E.M.; Dunst, J.; Haubner, A.; Durek, P.; Heinrich, F.; Gasparoni, G.; Babic, M.; Tomic, A.; et al. Peptide-specific recognition of human cytomegalovirus strains controls adaptive natural killer cells. Nat. Immunol. 2018, 19, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Fehniger, T.A.; Cooper, M.A. Harnessing NK Cell Memory for Cancer Immunotherapy. Trends Immunol. 2016, 37, 877–888. [Google Scholar] [CrossRef] [Green Version]
- Torelli, G.F.; Peragine, N.; Raponi, S.; Pagliara, D.; De Propris, M.S.; Vitale, A.; Bertaina, A.; Barberi, W.; Moretta, L.; Basso, G.; et al. Recognition of adult and pediatric acute lymphoblastic leukemia blasts by natural killer cells. Haematologica 2014, 99, 1248–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locatelli, F.; Moretta, F.; Brescia, L.; Merli, P. Natural killer cells in the treatment of high-risk acute leukaemia. Semin. Immunol. 2014, 26, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, C.; Vyas, P. The Graft-Versus-Leukemia Effect in AML. Front. Oncol. 2019, 9. [Google Scholar] [CrossRef]
- Paczulla, A.M.; Rothfelder, K.; Raffel, S.; Konantz, M.; Steinbacher, J.; Wang, H.; Tandler, C.; Mbarga, M.; Schaefer, T.; Falcone, M.; et al. Absence of NKG2D ligands defines leukaemia stem cells and mediates their immune evasion. Nature 2019, 572, 254–259. [Google Scholar] [CrossRef]
- David, G.; Djaoud, Z.; Willem, C.; Legrand, N.; Rettman, P.; Gagne, K.; Cesbron, A.; Retière, C. Large Spectrum of HLA-C Recognition by Killer Ig–like Receptor (KIR)2DL2 and KIR2DL3 and Restricted C1 Specificity of KIR2DS2: Dominant Impact of KIR2DL2/KIR2DS2 on KIR2D NK Cell Repertoire Formation. J. Immunol. 2013, 191, 4778–4788. [Google Scholar] [CrossRef] [Green Version]
- Hansasuta, P.; Dong, T.; Thananchai, H.; Weekes, M.; Willberg, C.; Aldemir, H.; Rowland-Jones, S.; Braud, V.M. Recognition of HLA-A3 and HLA-A11 by KIR3DL2 is peptide-specific. Eur. J. Immunol. 2004, 34, 1673–1679. [Google Scholar] [CrossRef] [PubMed]
- Morvan, M.; David, G.; Sébille, V.; Perrin, A.; Gagne, K.; Willem, C.; Kerdudou, N.; Denis, L.; Clémenceau, B.; Folléa, G.; et al. Autologous and allogeneic HLA KIR ligand environments and activating KIR control KIR NK-cell functions. Eur. J. Immunol. 2008, 38, 3474–3486. [Google Scholar] [CrossRef]
- Fauriat, C.; Andersson, S.; Björklund, A.T.; Carlsten, M.; Schaffer, M.; Björkström, N.K.; Baumann, B.C.; Michaëlsson, J.; Ljunggren, H.-G.; Malmberg, K.-J. Estimation of the Size of the Alloreactive NK Cell Repertoire: Studies in Individuals Homozygous for the Group A KIR Haplotype. J. Immunol. 2008, 181, 6010–6019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Heller, G.; Chewning, J.; Kim, S.; Yokoyama, W.M.; Hsu, K.C. Hierarchy of the Human Natural Killer Cell Response Is Determined by Class and Quantity of Inhibitory Receptors for Self-HLA-B and HLA-C Ligands. J. Immunol. 2007, 179, 5977–5989. [Google Scholar] [CrossRef]
- Hilton, H.G.; Guethlein, L.A.; Goyos, A.; Nemat-Gorgani, N.; Bushnell, D.A.; Norman, P.J.; Parham, P. Polymorphic HLA-C Receptors Balance the Functional Characteristics of KIR Haplotypes. J. Immunol. 2015, 195, 3160–3170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morvan, M.; Willem, C.; Gagne, K.; Kerdudou, N.; David, G.; Sébille, V.; Folléa, G.; Bignon, J.-D.; Retière, C. Phenotypic and Functional Analyses of KIR3DL1+ and KIR3DS1+ NK Cell Subsets Demonstrate Differential Regulation by Bw4 Molecules and Induced KIR3DS1 Expression on Stimulated NK Cells. J. Immunol. 2009, 182, 6727–6735. [Google Scholar] [CrossRef] [Green Version]
- Goodridge, J.P.; Önfelt, B.; Malmberg, K.-J. Newtonian cell interactions shape natural killer cell education. Immunol. Rev. 2015, 267, 197–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatton, O.; Strauss-Albee, D.M.; Zhao, N.Q.; Haggadone, M.D.; Pelpola, J.S.; Krams, S.M.; Martinez, O.M.; Blish, C.A. NKG2A-Expressing Natural Killer Cells Dominate the Response to Autologous Lymphoblastoid Cells Infected with Epstein–Barr Virus. Front. Immunol. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Forslund, E.; Sohlberg, E.; Enqvist, M.; Olofsson, P.E.; Malmberg, K.-J.; Önfelt, B. Microchip-Based Single-Cell Imaging Reveals That CD56dim CD57− KIR− NKG2A+ NK Cells Have More Dynamic Migration Associated with Increased Target Cell Conjugation and Probability of Killing Compared to CD56dim CD57− KIR− NKG2A− NK Cells. J. Immunol. 2015, 195, 3374–3381. [Google Scholar] [CrossRef] [Green Version]
- Russo, A.; Oliveira, G.; Berglund, S.; Greco, R.; Gambacorta, V.; Cieri, N.; Toffalori, C.; Zito, L.; Lorentino, F.; Piemontese, S.; et al. NK cell recovery after haploidentical HSCT with posttransplant cyclophosphamide: Dynamics and clinical implications. Blood 2018, 131, 247–262. [Google Scholar] [CrossRef] [Green Version]
- Mancusi, A.; Ruggeri, L.; Velardi, A. Haploidentical hematopoietic transplantation for the cure of leukemia: From its biology to clinical translation. Blood 2016, 128, 2616–2623. [Google Scholar] [CrossRef] [Green Version]
- Locatelli, F.; Pende, D.; Mingari, M.C.; Bertaina, A.; Falco, M.; Moretta, A.; Moretta, L. Cellular and molecular basis of haploidentical hematopoietic stem cell transplantation in the successful treatment of high-risk leukemias: Role of alloreactive NK cells. Front. Immunol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, S.; Dhedin, N.; Vernant, J.-P.; Kuentz, M.; Jijakli, A.A.; Rouas-Freiss, N.; Carosella, E.D.; Boudifa, A.; Debré, P.; Vieillard, V. NK-cell reconstitution after haploidentical hematopoietic stem-cell transplantations: Immaturity of NK cells and inhibitory effect of NKG2A override GvL effect. Blood 2005, 105, 4135–4142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodgins, J.J.; Khan, S.T.; Park, M.M.; Auer, R.C.; Ardolino, M. Killers 2.0: NK Cell Therapies at the Forefront of Cancer Control. Available online: http://www.jci.org/articles/view/129338/pdf (accessed on 7 May 2020).
- Ruggeri, L.; Parisi, S.; Urbani, E.; Curti, A. Alloreactive Natural Killer Cells for the Treatment of Acute Myeloid Leukemia: From Stem Cell Transplantation to Adoptive Immunotherapy. Front. Immunol. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, E.; Marin, D.; Banerjee, P.; Macapinlac, H.A.; Thompson, P.; Basar, R.; Kerbauy, L.N.; Overman, B.; Thall, P.; Kaplan, M.; et al. Use of CAR-Transduced Natural Killer Cells in CD19-Positive Lymphoid Tumors. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Manser, A.R.; Scherenschlich, N.; Thöns, C.; Hengel, H.; Timm, J.; Uhrberg, M. KIR Polymorphism Modulates the Size of the Adaptive NK Cell Pool in Human Cytomegalovirus–Infected Individuals. J. Immunol. 2019, 203, 2301–2309. [Google Scholar] [CrossRef]
- Liu, L.L.; Béziat, V.; Oei, V.Y.S.; Pfefferle, A.; Schaffer, M.; Lehmann, S.; Hellström-Lindberg, E.; Söderhäll, S.; Heyman, M.; Grandér, D.; et al. Ex Vivo Expanded Adaptive NK Cells Effectively Kill Primary Acute Lymphoblastic Leukemia Cells. Cancer Immunol. Res. 2017, 5, 654–665. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Verges, S.; Milush, J.M.; Schwartz, B.S.; Pando, M.J.; Jarjoura, J.; York, V.A.; Houchins, J.P.; Miller, S.; Kang, S.-M.; Norris, P.J.; et al. Expansion of a unique CD57+NKG2Chi natural killer cell subset during acute human cytomegalovirus infection. Proc. Natl. Acad. Sci. USA 2011, 108, 14725–14732. [Google Scholar] [CrossRef] [Green Version]
- Merino, A.; Zhang, B.; Dougherty, P.; Luo, X.; Wang, J.; Blazar, B.R.; Miller, J.S.; Cichocki, F. Chronic stimulation drives human NK cell dysfunction and epigenetic reprograming. J. Clin. Invest. 2019, 129, 3770–3785. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.Y.; Gaidulis, L.; Miller, M.M.; Goto, R.M.; Rodriguez, R.; Forman, S.J.; Senitzer, D. Development of a multiplex PCR-SSP method for Killer-cell immunoglobulin-like receptor genotyping. Tissue Antigens 2004, 64, 462–468. [Google Scholar] [CrossRef]
- Alter, G.; Malenfant, J.M.; Altfeld, M. CD107a as a functional marker for the identification of natural killer cell activity. J. Immunol. Methods 2004, 294, 15–22. [Google Scholar] [CrossRef]
- Penack, O.; Gentilini, C.; Fischer, L.; Asemissen, A.M.; Scheibenbogen, C.; Thiel, E.; Uharek, L. CD56dimCD16neg cells are responsible for natural cytotoxicity against tumor targets. Leukemia 2005, 19, 835–840. [Google Scholar] [CrossRef] [Green Version]
- Sturn, A.; Quackenbush, J.; Trajanoski, Z. Genesis: Cluster analysis of microarray data. Bioinformatics 2002, 18, 207–208. [Google Scholar] [CrossRef] [Green Version]
- Hsu, K.C.; Chida, S.; Geraghty, D.E.; Dupont, B. The killer cell immunoglobulin-like receptor (KIR) genomic region: Gene-order, haplotypes and allelic polymorphism. Immunol. Rev. 2002, 190, 40–52. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NK Receptors | CD Number | Ligands |
|---|---|---|
| Activating | ||
| DNAM-1 | CD226 | Nectin-2 (CD112), PVR(CD155) |
| 2B4 | CD244 | CD48 |
| NKp30 | CD337 | B7-H6, BAT3 a, PfEMP1 a |
| NKp44 | CD336 | Nidogen-1, PCNA a, HS a, HLA-DP |
| NKp46 | CD335 | CFP a, viral HA, HN a or envelope proteins, PfEMP1 a |
| NKG2D | CD314 | MIC-A a, MIC-B a, ULBP1-6 |
| NKG2C | CD159a | HLA-E |
| Inhibitory | ||
| NKG2A | CD159a | HLA-E |
| KIR2DL2/3 | CD158b | HLA-C1, HLA-C2 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makanga, D.R.; Da Rin de Lorenzo, F.; David, G.; Willem, C.; Dubreuil, L.; Legrand, N.; Guillaume, T.; Peterlin, P.; Lebourgeois, A.; Béné, M.C.; et al. Genetic and Molecular Basis of Heterogeneous NK Cell Responses against Acute Leukemia. Cancers 2020, 12, 1927. https://doi.org/10.3390/cancers12071927
Makanga DR, Da Rin de Lorenzo F, David G, Willem C, Dubreuil L, Legrand N, Guillaume T, Peterlin P, Lebourgeois A, Béné MC, et al. Genetic and Molecular Basis of Heterogeneous NK Cell Responses against Acute Leukemia. Cancers. 2020; 12(7):1927. https://doi.org/10.3390/cancers12071927
Chicago/Turabian StyleMakanga, Dhon Roméo, Francesca Da Rin de Lorenzo, Gaëlle David, Catherine Willem, Léa Dubreuil, Nolwenn Legrand, Thierry Guillaume, Pierre Peterlin, Amandine Lebourgeois, Marie Christine Béné, and et al. 2020. "Genetic and Molecular Basis of Heterogeneous NK Cell Responses against Acute Leukemia" Cancers 12, no. 7: 1927. https://doi.org/10.3390/cancers12071927
APA StyleMakanga, D. R., Da Rin de Lorenzo, F., David, G., Willem, C., Dubreuil, L., Legrand, N., Guillaume, T., Peterlin, P., Lebourgeois, A., Béné, M. C., Garnier, A., Chevallier, P., Gendzekhadze, K., Cesbron, A., Gagne, K., Clemenceau, B., & Retière, C. (2020). Genetic and Molecular Basis of Heterogeneous NK Cell Responses against Acute Leukemia. Cancers, 12(7), 1927. https://doi.org/10.3390/cancers12071927