MiR-155-5p and MiR-203a-3p Are Prognostic Factors in Soft Tissue Sarcoma

 ,
,  ,
, 

Abstract
:1. Introduction
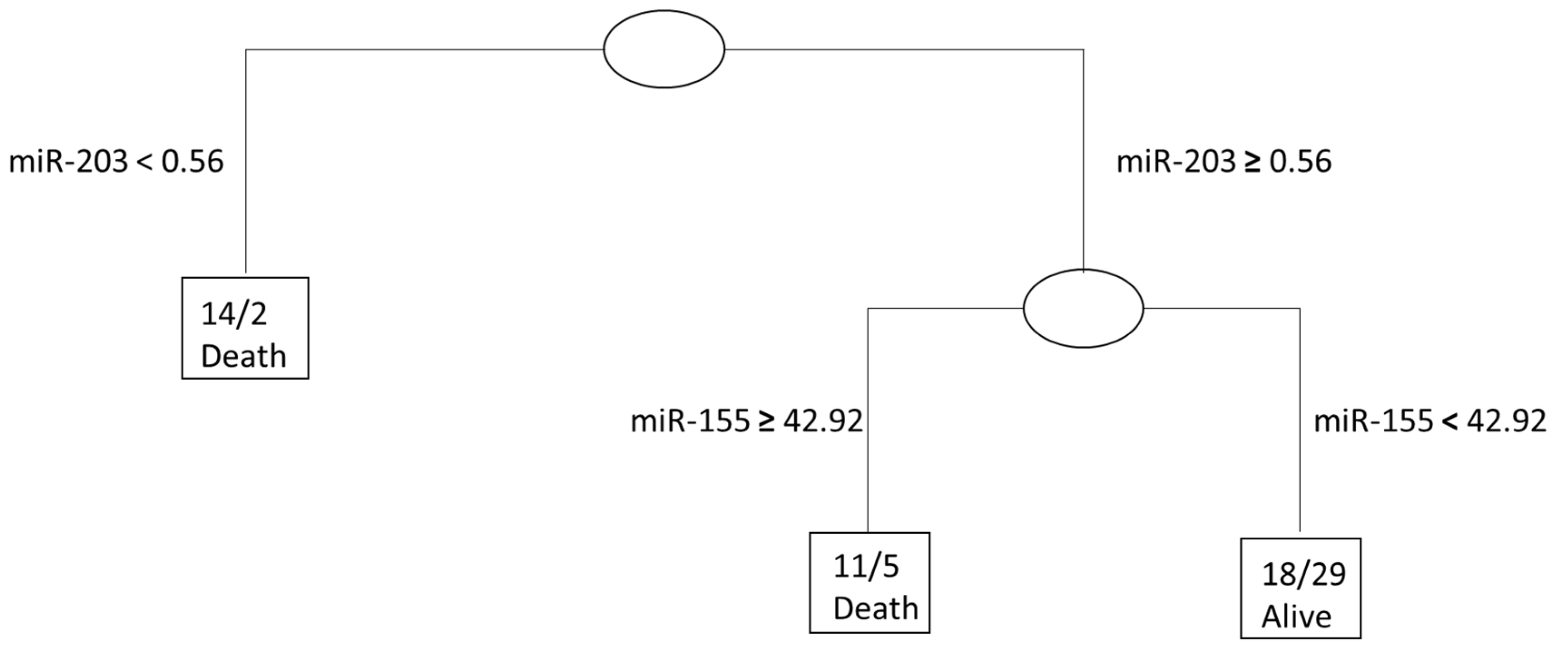
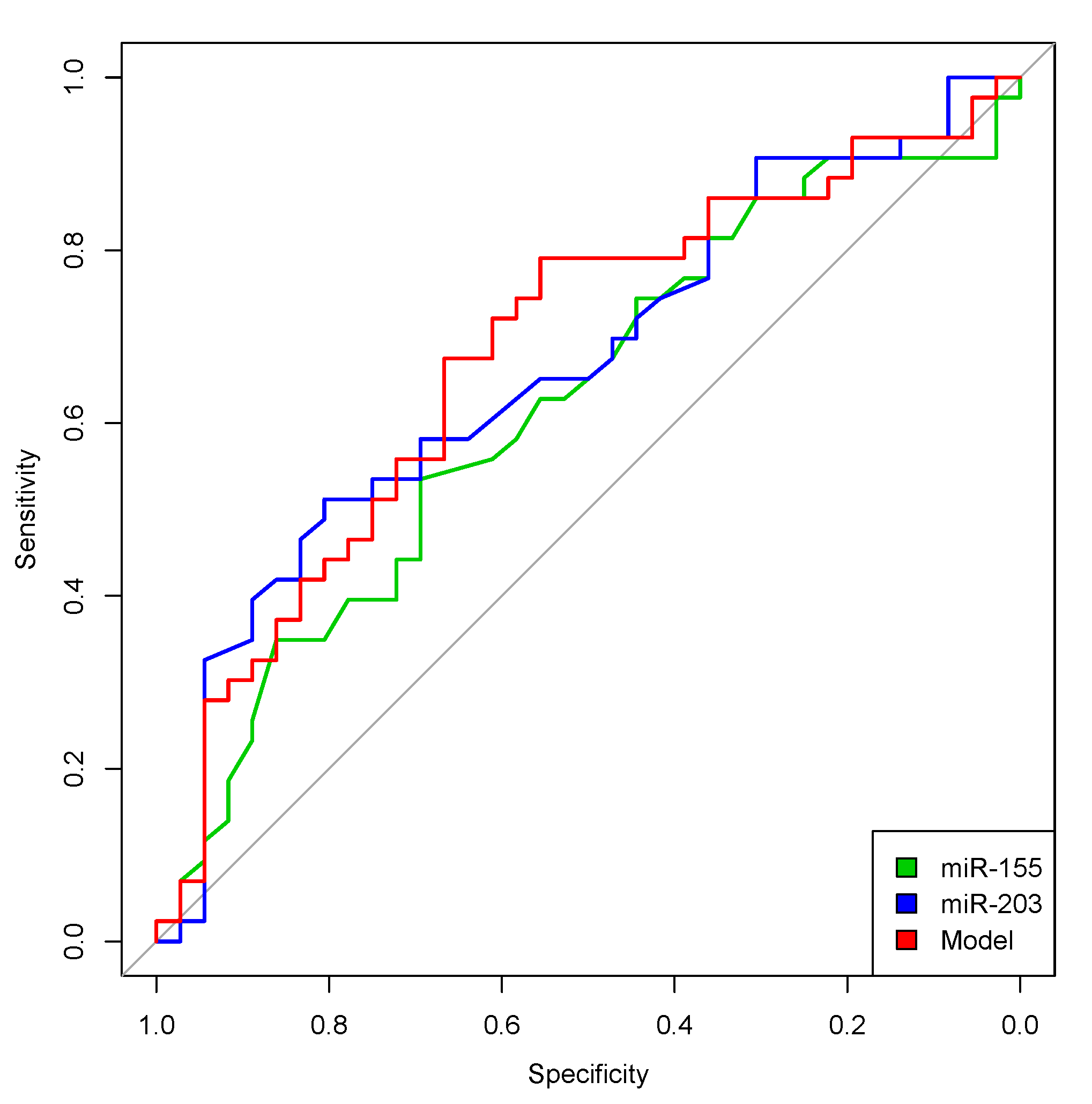
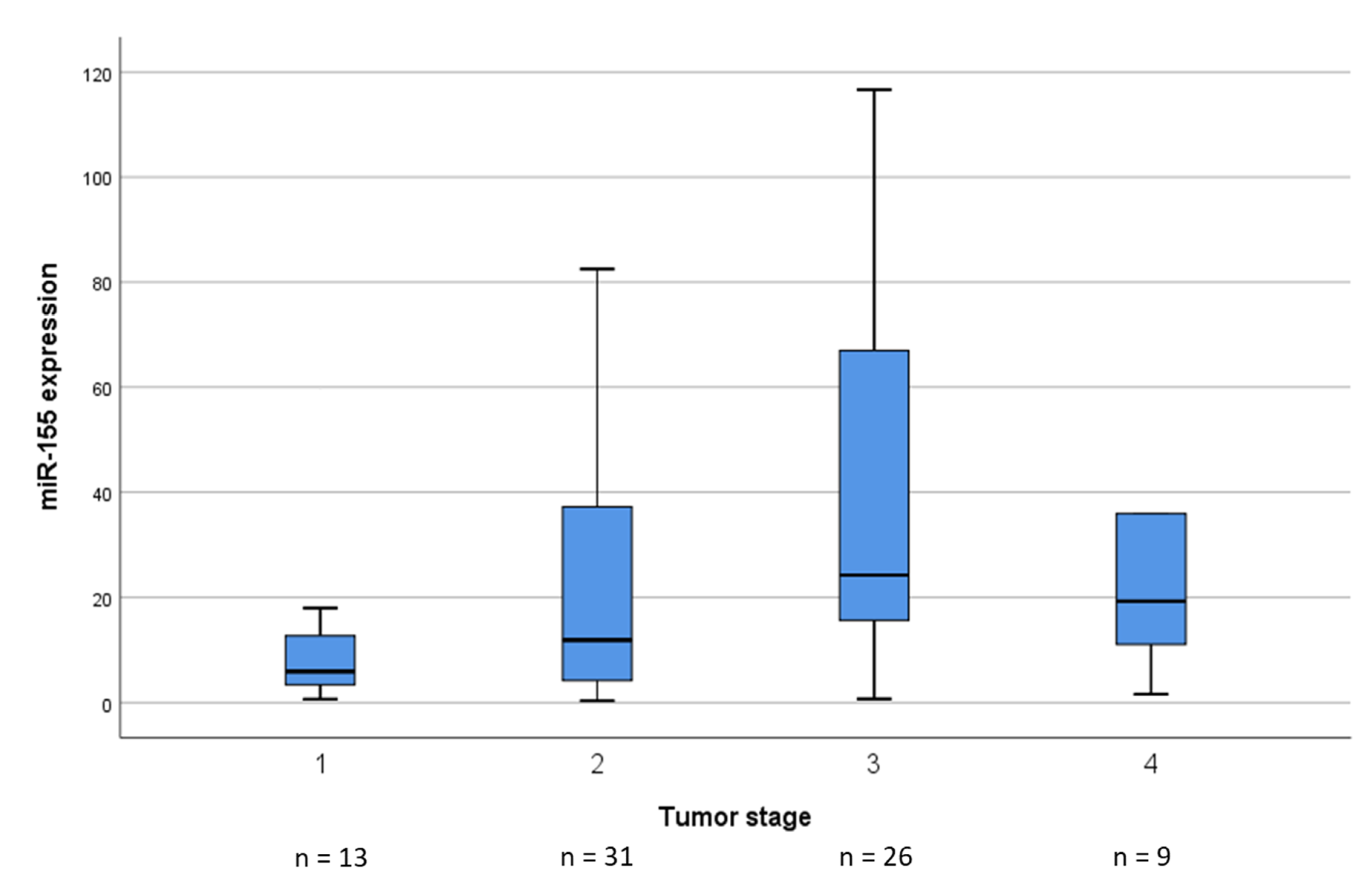
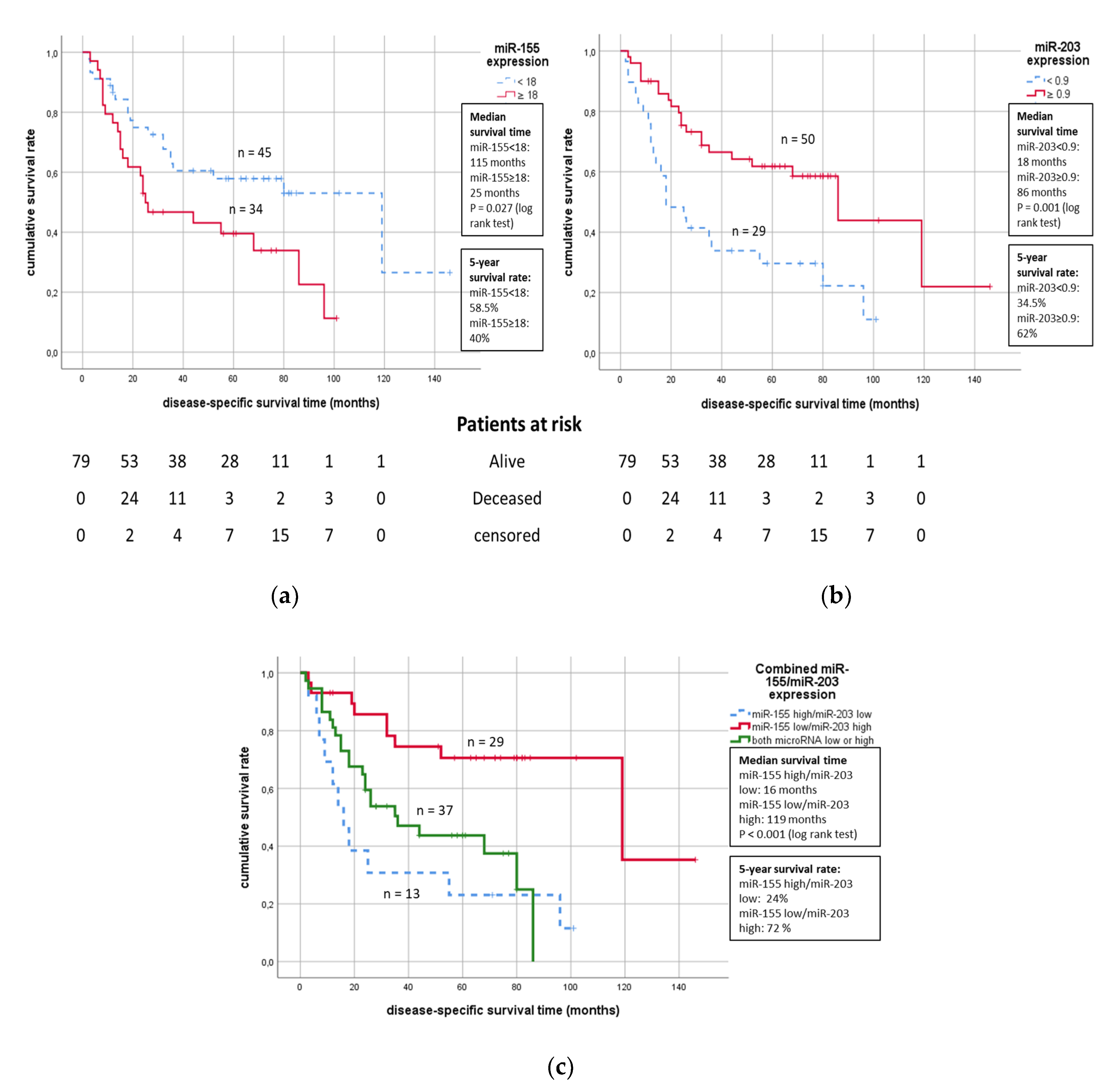
2. Results
2.1. miR-155 and miR-203 Expression in STS Specimens
2.2. Bivariate Correlation Analyses
2.3. Survival Analyses
3. Discussion
4. Materials and Methods
4.1. Patients and Specimens
4.2. RNA Isolation and cDNA Synthesis
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Taylor, B.S.; Barretina, J.; Maki, R.G.; Antonescu, C.R.; Singer, S.; Ladanyi, M. Advances in sarcoma genomics and new therapeutic targets. Nat. Rev. Cancer 2011, 11, 541–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducimetière, F.; Lurkin, A.; Ranchère-Vince, D.; Decouvelaere, A.-V.; Péoc’h, M.; Istier, L.; Chalabreysse, P.; Muller, C.; Alberti, L.; Bringuier, P.-P.; et al. Incidence of sarcoma histotypes and molecular subtypes in a prospective epidemiological study with central pathology review and molecular testing. PLoS ONE 2011, 6, e20294. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, A.; Sultan, I.; Huang, T.T.; Rodriguez-Galindo, C.; Shehadeh, A.; Meazza, C.; Ness, K.K.; Casanova, M.; Spunt, S.L. Soft tissue sarcoma across the age spectrum: a population-based study from the Surveillance Epidemiology and End Results database. Pediatr. Blood Cancer 2011, 57, 943–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, D.P. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Smolle, M.A.; Leithner, A.; Posch, F.; Szkandera, J.; Liegl-Atzwanger, B.; Pichler, M. MicroRNAs in Different Histologies of Soft Tissue Sarcoma: A Comprehensive Review. Int. J. Mol. Sci. 2017, 18, 1960. [Google Scholar] [CrossRef] [Green Version]
- Greither, T.; Grochola, L.F.; Udelnow, A.; Lautenschläger, C.; Würl, P.; Taubert, H. Elevated expression of microRNAs 155, 203, 210 and 222 in pancreatic tumors is associated with poorer survival. Int. J. Cancer 2010, 126, 73–80. [Google Scholar] [CrossRef]
- Greither, T.; Würl, P.; Grochola, L.; Bond, G.; Bache, M.; Kappler, M.; Lautenschläger, C.; Holzhausen, H.-J.; Wach, S.; Eckert, A.W.; et al. Expression of microRNA 210 associates with poor survival and age of tumor onset of soft-tissue sarcoma patients. Int. J. Cancer 2012, 130, 1230–1235. [Google Scholar] [CrossRef] [PubMed]
- Elton, T.S.; Selemon, H.; Elton, S.M.; Parinandi, N.L. Regulation of the MIR155 host gene in physiological and pathological processes. Gene 2013, 532, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Bill, K.; Liu, J.; Young, E.; Peng, T.; Bolshakov, S.; Hoffman, A.; Song, Y.; Demicco, E.G.; Terrada, D.L.; et al. MiR-155 is a liposarcoma oncogene that targets casein kinase-1α and enhances β-catenin signaling. Cancer Res. 2012, 72, 1751–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincenzi, B.; Iuliani, M.; Zoccoli, A.; Pantano, F.; Fioramonti, M.; de Lisi, D.; Frezza, A.M.; Rabitti, C.; Perrone, G.; Onetti Muda, A.; et al. Deregulation of dicer and mir-155 expression in liposarcoma. Oncotarget 2015, 6, 10586–10591. [Google Scholar] [CrossRef] [Green Version]
- Skalsky, R.L.; Samols, M.A.; Plaisance, K.B.; Boss, I.W.; Riva, A.; Lopez, M.C.; Baker, H.V.; Renne, R. Kaposi’s sarcoma-associated herpesvirus encodes an ortholog of miR-155. J. Virol. 2007, 81, 12836–12845. [Google Scholar] [CrossRef] [Green Version]
- Gottwein, E.; Mukherjee, N.; Sachse, C.; Frenzel, C.; Majoros, W.H.; Chi, J.-T.A.; Braich, R.; Manoharan, M.; Soutschek, J.; Ohler, U.; et al. A viral microRNA functions as an orthologue of cellular miR-155. Nature 2007, 450, 1096–1099. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Geng, X.; Zhang, J.; Zhao, H.; Liu, Y. miR-155 promotes the growth of osteosarcoma in a HBP1-dependent mechanism. Mol. Cell. Biochem. 2015, 403, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tang, B.; Han, H.; Mao, D.; Chen, J.; Zeng, Y.; Xiong, M. miR-155 Affects Osteosarcoma MG-63 Cell Autophagy Induced by Adriamycin Through Regulating PTEN-PI3K/AKT/mTOR Signaling Pathway. Cancer Biother. Radiopharm. 2018, 33, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Liao, Q.-S.; Tang, L. MiR-155 affects osteosarcoma cell proliferation and invasion through regulating NF-κB signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7633–7639. [Google Scholar] [CrossRef] [PubMed]
- Griffiths-Jones, S.; Grocock, R.J.; van Dongen, S.; Bateman, A.; Enright, A.J. miRBase: microRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef] [PubMed]
- Hezova, R.; Kovarikova, A.; Srovnal, J.; Zemanova, M.; Harustiak, T.; Ehrmann, J.; Hajduch, M.; Svoboda, M.; Sachlova, M.; Slaby, O. Diagnostic and prognostic potential of miR-21, miR-29c, miR-148 and miR-203 in adenocarcinoma and squamous cell carcinoma of esophagus. Diagn. Pathol. 2015, 10, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.; Yang, W.; Zhu, Y.; Yuan, Y. Prognostic role of microRNA-203 in various carcinomas: evidence from a meta-analysis involving 13 studies. Springerplus 2016, 5, 1538. [Google Scholar] [CrossRef] [Green Version]
- Diao, Y.; Guo, X.; Jiang, L.; Wang, G.; Zhang, C.; Wan, J.; Jin, Y.; Wu, Z. miR-203, a tumor suppressor frequently down-regulated by promoter hypermethylation in rhabdomyosarcoma. J. Biol. Chem. 2014, 289, 529–539. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Liu, G.; Wang, K. miR-203 Acts as a Tumor Suppressor Gene in Osteosarcoma by Regulating RAB22A. PLoS ONE 2015, 10, e0132225. [Google Scholar] [CrossRef]
- Taubert, H.; Meye, A.; Würl, P. Soft tissue sarcomas and p53 mutations. Mol Med. 1998, 4, 365–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bache, M.; Kappler, M.; Wichmann, H.; Rot, S.; Hahnel, A.; Greither, T.; Said, H.M.; Kotzsch, M.; Würl, P.; Taubert, H.; et al. Elevated tumor and serum levels of the hypoxia-associated protein osteopontin are associated with prognosis for soft tissue sarcoma patients. BMC Cancer 2010, 10, 132. [Google Scholar] [CrossRef] [Green Version]
- Kotzsch, M.; Magdolen, V.; Greither, T.; Kappler, M.; Bache, M.; Lautenschläger, C.; Füssel, S.; Eckert, A.W.; Luther, T.; Baretton, G.; et al. Combined mRNA expression levels of members of the urokinase plasminogen activator (uPA) system correlate with disease-associated survival of soft-tissue sarcoma patients. BMC Cancer 2011, 11, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rot, S.; Taubert, H.; Bache, M.; Greither, T.; Würl, P.; Eckert, A.W.; Schubert, J.; Vordermark, D.; Kappler, M. A novel splice variant of the stem cell marker LGR5/GPR49 is correlated with the risk of tumor-related death in soft-tissue sarcoma patients. BMC Cancer 2011, 11, 429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahnel, A.; Wichmann, H.; Greither, T.; Kappler, M.; Würl, P.; Kotzsch, M.; Taubert, H.; Vordermark, D.; Bache, M. Prognostic impact of mRNA levels of osteopontin splice variants in soft tissue sarcoma patients. BMC Cancer 2012, 12, 131. [Google Scholar] [CrossRef] [Green Version]
- Rot, S.; Taubert, H.; Bache, M.; Greither, T.; Würl, P.; Holzhausen, H.-J.; Eckert, A.W.; Vordermark, D.; Kappler, M. Low HIF-1α and low EGFR mRNA Expression Significantly Associate with Poor Survival in Soft Tissue Sarcoma Patients; the Proteins React Differently. Int. J. Mol. Sci. 2018, 19, 3842. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Ma, T.; Huang, C.; Hu, T.; Li, J. The pivotal role of microRNA-155 in the control of cancer. J. Cell. Physiol. 2014, 229, 545–550. [Google Scholar] [CrossRef]
- Jurkovicova, D.; Magyerkova, M.; Kulcsar, L.; Krivjanska, M.; Krivjansky, V.; Gibadulinova, A.; Oveckova, I.; Chovanec, M. miR-155 as a diagnostic and prognostic marker in hematological and solid malignancies. Neoplasma 2014, 61, 241–251. [Google Scholar] [CrossRef]
- Furuta, M.; Kozaki, K.-i.; Tanaka, S.; Arii, S.; Imoto, I.; Inazawa, J. miR-124 and miR-203 are epigenetically silenced tumor-suppressive microRNAs in hepatocellular carcinoma. Carcinogenesis 2010, 31, 766–776. [Google Scholar] [CrossRef] [Green Version]
- Sonkoly, E.; Lovén, J.; Xu, N.; Meisgen, F.; Wei, T.; Brodin, P.; Jaks, V.; Kasper, M.; Shimokawa, T.; Harada, M.; et al. MicroRNA-203 functions as a tumor suppressor in basal cell carcinoma. Oncogenesis 2012, 1, e3. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.-R.; Guo, G.-L.; Zhang, W.; Huang, G.-L.; Hu, X.-Q.; Zhu, J.; Huang, Q.-D.; You, J.; Zhang, X.-H. Clinical significance of miR-155 expression in breast cancer and effects of miR-155 ASO on cell viability and apoptosis. Oncol. Rep. 2012, 27, 1149–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.-D.; Zhang, W.; Liang, H.-J.; Ji, W.-Y. Overexpression of miR -155 promotes proliferation and invasion of human laryngeal squamous cell carcinoma via targeting SOCS1 and STAT3. PLoS ONE. 2013, 8, e56395. [Google Scholar] [CrossRef] [PubMed]
- Osaka, E.; Kelly, A.D.; Spentzos, D.; Choy, E.; Yang, X.; Shen, J.K.; Yang, P.; Mankin, H.J.; Hornicek, F.J.; Duan, Z. MicroRNA-155 expression is independently predictive of outcome in chordoma. Oncotarget 2015, 6, 9125–9139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Dong, J.; He, X.; Shen, L.; Long, C.; Liu, F.; Liu, X.; Lin, T.; He, D.; Wei, G. MiR-155-5p exerts tumor-suppressing functions in Wilms tumor by targeting IGF2 via the PI3K signaling pathway. Biomed. Pharmacother. 2020, 125, 109880. [Google Scholar] [CrossRef]
- Fasanaro, P.; D’Alessandra, Y.; Di Stefano, V.; Melchionna, R.; Romani, S.; Pompilio, G.; Capogrossi, M.C.; Martelli, F. MicroRNA-210 modulates endothelial cell response to hypoxia and inhibits the receptor tyrosine kinase ligand Ephrin-A3. J. Biol. Chem. 2008, 283, 15878–15883. [Google Scholar] [CrossRef] [Green Version]
- Kong, W.; He, L.; Richards, E.J.; Challa, S.; Xu, C.-X.; Permuth-Wey, J.; Lancaster, J.M.; Coppola, D.; Sellers, T.A.; Djeu, J.Y.; et al. Upregulation of miRNA-155 promotes tumour angiogenesis by targeting VHL and is associated with poor prognosis and triple-negative breast cancer. Oncogene 2014, 33, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Neilsen, P.M.; Noll, J.E.; Mattiske, S.; Bracken, C.P.; Gregory, P.A.; Schulz, R.B.; Lim, S.P.; Kumar, R.; Suetani, R.J.; Goodall, G.J.; et al. Mutant p53 drives invasion in breast tumors through up-regulation of miR-155. Oncogene 2013, 32, 2992–3000. [Google Scholar] [CrossRef] [Green Version]
- Kunz, C.; Pebler, S.; Otte, J.; von der Ahe, D. Differential regulation of plasminogen activator and inhibitor gene transcription by the tumor suppressor p53. Nucleic Acids Res. 1995, 23, 3710–3717. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Guo, D.; Wang, Q.; Liu, P.; Xiao, Y.; Wu, P.; Wang, Y.; Chen, B.; Liu, Z.; Liu, Q. Lgr5-mediated p53 Repression through PDCD5 leads to doxorubicin resistance in Hepatocellular Carcinoma. Theranostics 2019, 9, 2967–2983. [Google Scholar] [CrossRef]
- Li, L.; Han, M.-x.; Li, S.; Xu, Y.; Wang, L. Hypoxia regulates the proliferation and osteogenic differentiation of human periodontal ligament cells under cyclic tensile stress via mitogen-activated protein kinase pathways. J. Periodontol. 2014, 85, 498–508. [Google Scholar] [CrossRef]
- Yang, X.; Li, X.; Lin, Q.; Xu, Q. Up-regulation of microRNA-203 inhibits myocardial fibrosis and oxidative stress in mice with diabetic cardiomyopathy through the inhibition of PI3K/Akt signaling pathway via PIK3CA. Gene 2019, 715, 143995. [Google Scholar] [CrossRef] [PubMed]
- Kapodistrias, N.; Mavridis, K.; Batistatou, A.; Gogou, P.; Karavasilis, V.; Sainis, I.; Briasoulis, E.; Scorilas, A. Assessing the clinical value of microRNAs in formalin-fixed paraffin-embedded liposarcoma tissues: Overexpressed miR-155 is an indicator of poor prognosis. Oncotarget 2017, 8, 6896–6913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donnem, T.; Eklo, K.; Berg, T.; Sorbye, S.W.; Lonvik, K.; Al-Saad, S.; Al-Shibli, K.; Andersen, S.; Stenvold, H.; Bremnes, R.M.; et al. Prognostic impact of MiR-155 in non-small cell lung cancer evaluated by in situ hybridization. J. Transl. Med. 2011, 9, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibuya, H.; Iinuma, H.; Shimada, R.; Horiuchi, A.; Watanabe, T. Clinicopathological and prognostic value of microRNA-21 and microRNA-155 in colorectal cancer. Oncology 2010, 79, 313–320. [Google Scholar] [CrossRef]
- Han, Z.-B.; Chen, H.-Y.; Fan, J.-W.; Wu, J.-Y.; Tang, H.-M.; Peng, Z.-H. Up-regulation of microRNA-155 promotes cancer cell invasion and predicts poor survival of hepatocellular carcinoma following liver transplantation. J. Cancer Res. Clin. Oncol. 2012, 138, 153–161. [Google Scholar] [CrossRef]
- Chen, J.; Wang, B.-C.; Tang, J.-H. Clinical significance of microRNA-155 expression in human breast cancer. J. Surg. Oncol. 2012, 106, 260–266. [Google Scholar] [CrossRef]
- Sun, J.; Shi, H.; Lai, N.; Liao, K.; Zhang, S.; Lu, X. Overexpression of microRNA-155 predicts poor prognosis in glioma patients. Med. Oncol. 2014, 31, 911. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Y.; Guo, Q.; Diao, Y.; Liu, H.; Song, G.; Wang, W.; Zhang, Z.; Yin, H.; Li, L. Prognostic role of microRNA-155 in patients with leukemia: A meta-analysis. Clin. Chim. Acta 2018, 483, 6–13. [Google Scholar] [CrossRef]
- Ikenaga, N.; Ohuchida, K.; Mizumoto, K.; Yu, J.; Kayashima, T.; Sakai, H.; Fujita, H.; Nakata, K.; Tanaka, M. MicroRNA-203 expression as a new prognostic marker of pancreatic adenocarcinoma. Ann. Surg. Oncol. 2010, 17, 3120–3128. [Google Scholar] [CrossRef]
- Wang, C.; Wang, X.; Liang, H.; Wang, T.; Yan, X.; Cao, M.; Wang, N.; Zhang, S.; Zen, K.; Zhang, C.; et al. miR-203 inhibits cell proliferation and migration of lung cancer cells by targeting PKCα. PLoS ONE 2013, 8, e73985. [Google Scholar] [CrossRef]
- Madhavan, D.; Peng, C.; Wallwiener, M.; Zucknick, M.; Nees, J.; Schott, S.; Rudolph, A.; Riethdorf, S.; Trumpp, A.; Pantel, K.; et al. Circulating miRNAs with prognostic value in metastatic breast cancer and for early detection of metastasis. Carcinogenesis 2016, 37, 461–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schetter, A.J.; Leung, S.Y.; Sohn, J.J.; Zanetti, K.A.; Bowman, E.D.; Yanaihara, N.; Yuen, S.T.; Chan, T.L.; Kwong, D.L.W.; Au, G.K.H.; et al. MicroRNA expression profiles associated with prognosis and therapeutic outcome in colon adenocarcinoma. JAMA 2008, 299, 425–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bovell, L.C.; Shanmugam, C.; Putcha, B.-D.K.; Katkoori, V.R.; Zhang, B.; Bae, S.; Singh, K.P.; Grizzle, W.E.; Manne, U. The prognostic value of microRNAs varies with patient race/ethnicity and stage of colorectal cancer. Clin. Cancer Res. 2013, 19, 3955–3965. [Google Scholar] [CrossRef] [Green Version]
- Costa, F.F.; Bischof, J.M.; Vanin, E.F.; Lulla, R.R.; Wang, M.; Sredni, S.T.; Rajaram, V.; Bonaldo, M.d.F.; Wang, D.; Goldman, S.; et al. Identification of microRNAs as potential prognostic markers in ependymoma. PLoS ONE 2011, 6, e25114. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Deng, Y.; Yang, G.; Xie, W. MicroRNA-203 down-regulation is associated with unfavorable prognosis in human glioma. J. Surg. Oncol. 2013, 108, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-Y.; Han, Z.-B.; Fan, J.-W.; Xia, J.; Wu, J.-Y.; Qiu, G.-Q.; Tang, H.-M.; Peng, Z.-H. miR-203 expression predicts outcome after liver transplantation for hepatocellular carcinoma in cirrhotic liver. Med. Oncol. 2012, 29, 1859–1865. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, X.; Wang, J.; Wen, Y.; Zhang, L.; Wang, D.; Chen, H.; Chen, Q.; Xiang, W. Upregulation of microRNA-203 is associated with advanced tumor progression and poor prognosis in epithelial ovarian cancer. Med. Oncol. 2013, 30, 681. [Google Scholar] [CrossRef]
- Bian, K.; Fan, J.; Zhang, X.; Yang, X.-W.; Zhu, H.-Y.; Wang, L.; Sun, J.-Y.; Meng, Y.-L.; Cui, P.-C.; Cheng, S.-Y.; et al. MicroRNA-203 leads to G1 phase cell cycle arrest in laryngeal carcinoma cells by directly targeting survivin. FEBS Lett. 2012, 586, 804–809. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Liang, H.; Zhou, Y.; Wang, C.; Zhang, S.; Pan, Y.; Wang, Y.; Yan, X.; Zhang, J.; Zhang, C.-Y.; et al. miR-203 suppresses the proliferation and migration and promotes the apoptosis of lung cancer cells by targeting SRC. PLoS ONE 2014, 9, e105570. [Google Scholar] [CrossRef]
- Siu, M.K.; Abou-Kheir, W.; Yin, J.J.; Chang, Y.-S.; Barrett, B.; Suau, F.; Casey, O.; Chen, W.-Y.; Fang, L.; Hynes, P.; et al. Loss of EGFR signaling regulated miR-203 promotes prostate cancer bone metastasis and tyrosine kinase inhibitors resistance. Oncotarget 2014, 5, 3770–3784. [Google Scholar] [CrossRef] [Green Version]
- Teicher, B.A. Searching for molecular targets in sarcoma. Biochem. Pharmacol. 2012, 84, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bonneau, E.; Neveu, B.; Kostantin, E.; Tsongalis, G.J.; de Guire, V. How close are miRNAs from clinical practice? A perspective on the diagnostic and therapeutic market. EJIFCC 2019, 30, 114–127. [Google Scholar] [PubMed]
- Kappler, M.; Köhler, T.; Kampf, C.; Diestelkötter, P.; Würl, P.; Schmitz, M.; Bartel, F.; Lautenschläger, C.; Rieber, E.P.; Schmidt, H.; et al. Increased survivin transcript levels: an independent negative predictor of survival in soft tissue sarcoma patients. Int. J. Cancer 2001, 95, 360–363. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | rs | p | n | Reference | |
|---|---|---|---|---|---|
| miR-155 | p53 mutations | 0.260 | 0.040 | 63 | [21] |
| OPN protein | 0.361 | 0.002 | 72 | [22] | |
| uPA protein | 0.604 | <0.001 | 79 | [23] | |
| uPAR protein | 0.440 | <0.001 | 79 | [23] | |
| PAI-1 protein | 0.347 | 0.002 | 79 | [23] | |
| EFNA3 mRNA | −0.378 | 0.001 | 75 | n.p. | |
| LGR5 mRNA | −0.389 | 0.001 | 69 | [24] | |
| miR-210 | 0.349 | 0.002 | 78 | [7] | |
| miR-203 | miR-210 | 0.265 | 0.019 | 78 | [7] |
| OPN-a mRNA | −0.267 | 0.021 | 74 | [25] | |
| OPN-b mRNA | −0.324 | 0.005 | 74 | [25] | |
| OPN-c mRNA | −0.260 | 0.025 | 74 | [25] | |
| pAKT473 protein | −0.434 | <0.001 | 73 | [26] |
| Parameters | Univariate Cox Regression Analysis | Multivariate Cox Regression Analysis | |||||
|---|---|---|---|---|---|---|---|
| n | p | RR | n | p | RR | ||
| miR-155 low | 45 | Reference | 45 | Reference | Tumor-specific survival | ||
| miR-155 high | 34 | 0.031 | 1.96 (1.06–3.61) | 34 | 0.766 | 1.13 (0.52–2.46) | |
| miR-203 low | 29 | 0.002 | 2.59 (1.41–4.76) | 29 | 0.077 | 1.99 (0.93–4.25) | |
| miR-203 high | 50 | Reference | 50 | Reference | |||
| miR-155 low + miR-203 high | 29 | Reference | 29 | Reference | |||
| miR-155 high + miR-203 low | 13 | 0.001 | 4.76 (1.89–12.05) | 13 | 0.21 | 4.05 (0.46–35.7) | |
| miR-155 low | 45 | Reference | 45 | Reference | Relapse-free survival | ||
| miR-155 high | 34 | 0.985 | 1.01 (0.5–2.01) | 34 | 0.512 | 1.32 (0.34–1.73) | |
| miR-203 low | 29 | 0.189 | 1.61 (0.79–3.26) | 29 | 0.405 | 1.60 (0.21–1.90) | |
| miR-203 high | 50 | Reference | 50 | Reference | |||
| miR-155 low + miR-203 high | 29 | 0.455 | 1.5 (0.23–1.93) | 29 | Reference | ||
| miR-155 high + miR-203 low | 13 | Reference | 13 | 0.247 | 3.04 (0.46–19.99) | ||
| Parameters | miR-155 Low (<18.0) | miR-155 High (≥18.0) | Chi2 Test (p-Value) | miR-203 Low (<0.9) | miR-203 High (≥0.9) | Chi2 Test (p-Value) | |
|---|---|---|---|---|---|---|---|
| age | <60 years | 23 | 20 | n.s. | 15 | 28 | n.s. |
| >60 years | 22 | 14 | 14 | 22 | |||
| gender | female | 24 | 12 | n.s. | 16 | 20 | n.s. |
| male | 21 | 22 | 13 | 30 | |||
| patients status | alive | 25 | 11 | 0.04 | 7 | 29 | 0.004 |
| deceased | 20 | 23 | 22 | 21 | |||
| tumor stage a | I | 12 | 1 | 0.011 | 3 | 10 | n.s. |
| II | 19 | 12 | 10 | 21 | |||
| III | 11 | 15 | 11 | 15 | |||
| IV | 3 | 6 | 5 | 4 | |||
| resection | radical (R0) | 32 | 21 | n.s. | 17 | 36 | n.s. |
| not radical (R1) | 13 | 13 | 12 | 14 | |||
| tumor localization | extremities | 29 | 23 | n.s. | 18 | 34 | n.s. |
| trunk wall | 3 | 2 | 2 | 3 | |||
| head/neck | 2 | 0 | 1 | 1 | |||
| abdomen/peritoneum | 9 | 9 | 6 | 12 | |||
| multiple locations | 2 | 0 | 2 | 0 | |||
| histological subtypes | LS | 12 | 9 | 0.011 | 10 | 11 | n.s. |
| FS + NOS | 12 | 8 | 4 | 16 | |||
| NS | 4 | 3 | 3 | 4 | |||
| RMS + LMS | 11 | 10 | 8 | 13 | |||
| other | 6 | 4 | 4 | 6 | |||
| tumor size | T1 | 6 | 5 | n.s. | 3 | 8 | n.s. |
| T2 | 39 | 29 | 26 | 42 | |||
| number of relapses | 0 | 28 | 20 | n.s. | 15 | 33 | n.s. |
| 1 | 8 | 5 | 6 | 7 | |||
| >2 | 9 | 9 | 8 | 10 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Greither, T.; Koser, F.; Holzhausen, H.-J.; Güttler, A.; Würl, P.; Kappler, M.; Wach, S.; Taubert, H. MiR-155-5p and MiR-203a-3p Are Prognostic Factors in Soft Tissue Sarcoma. Cancers 2020, 12, 2254. https://doi.org/10.3390/cancers12082254
Greither T, Koser F, Holzhausen H-J, Güttler A, Würl P, Kappler M, Wach S, Taubert H. MiR-155-5p and MiR-203a-3p Are Prognostic Factors in Soft Tissue Sarcoma. Cancers. 2020; 12(8):2254. https://doi.org/10.3390/cancers12082254
Chicago/Turabian StyleGreither, Thomas, Franziska Koser, Hans-Jürgen Holzhausen, Antje Güttler, Peter Würl, Matthias Kappler, Sven Wach, and Helge Taubert. 2020. "MiR-155-5p and MiR-203a-3p Are Prognostic Factors in Soft Tissue Sarcoma" Cancers 12, no. 8: 2254. https://doi.org/10.3390/cancers12082254
APA StyleGreither, T., Koser, F., Holzhausen, H. -J., Güttler, A., Würl, P., Kappler, M., Wach, S., & Taubert, H. (2020). MiR-155-5p and MiR-203a-3p Are Prognostic Factors in Soft Tissue Sarcoma. Cancers, 12(8), 2254. https://doi.org/10.3390/cancers12082254