Impact of ERCC1, XPF and DNA Polymerase β Expression on Platinum Response in Patient-Derived Ovarian Cancer Xenografts

 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Results
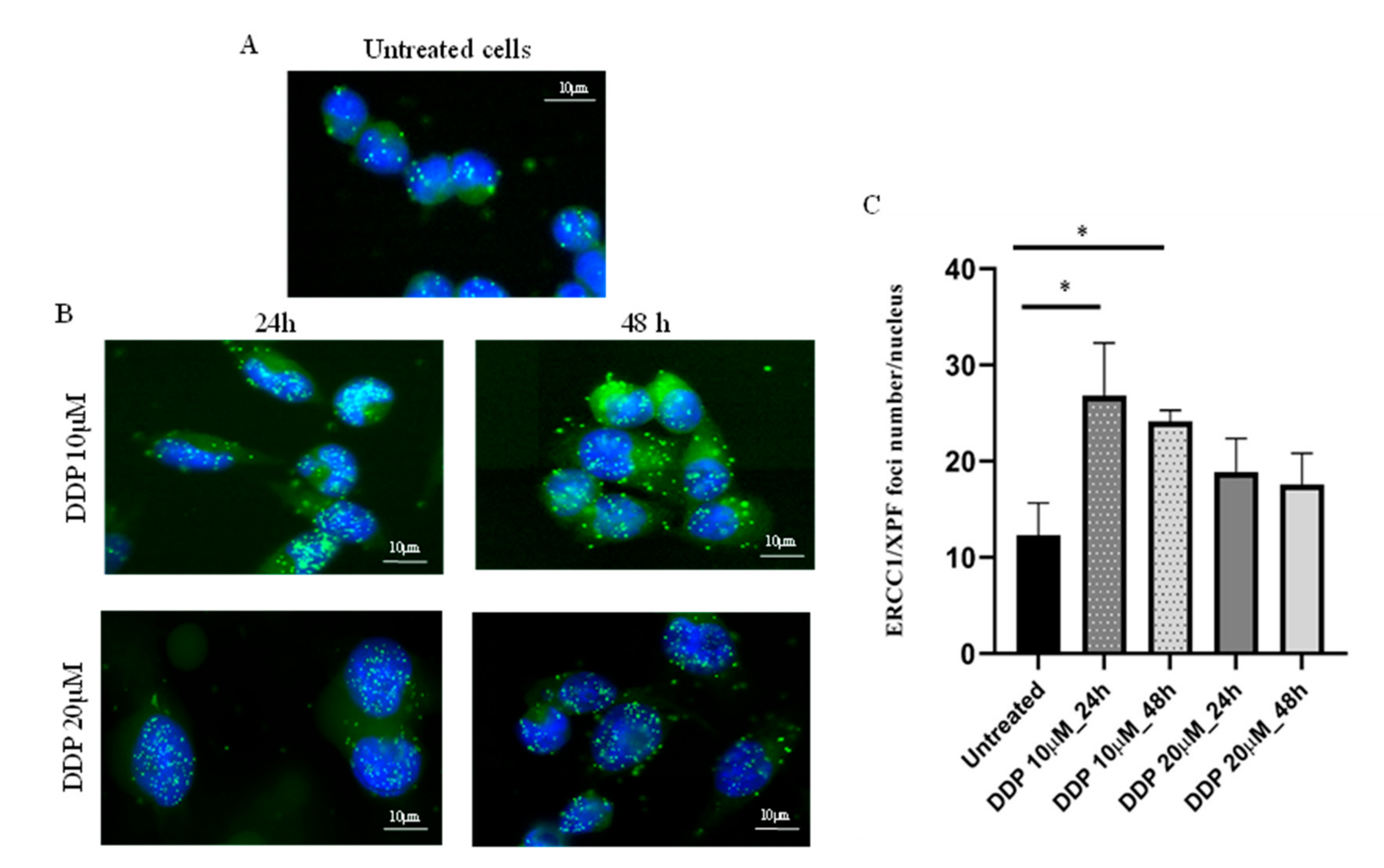
2.1. Relationship between ERCC1/XPF/DNA pol β mRNAs or Proteins and PLA-foci Expression in the Ovarian Xenobank.
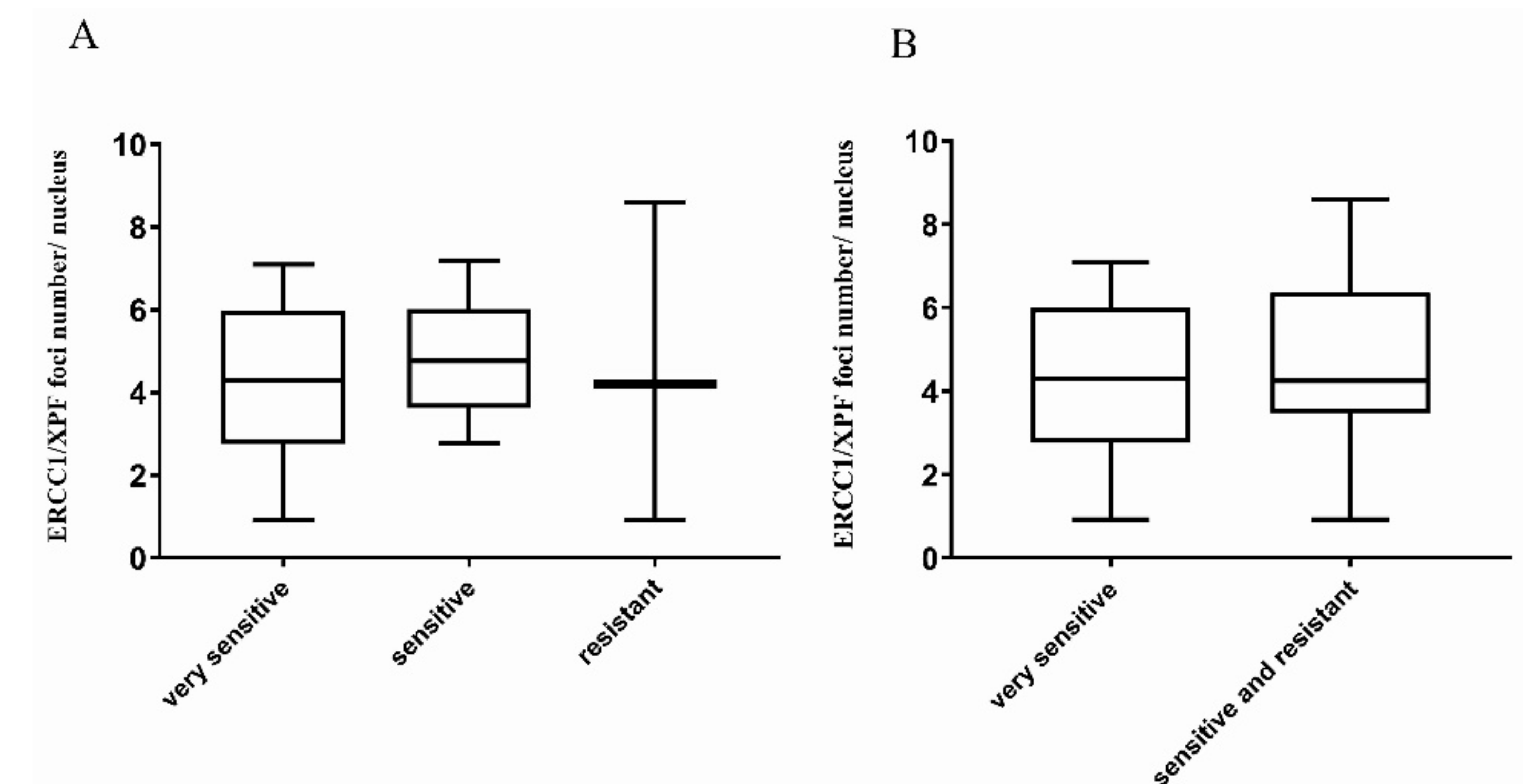
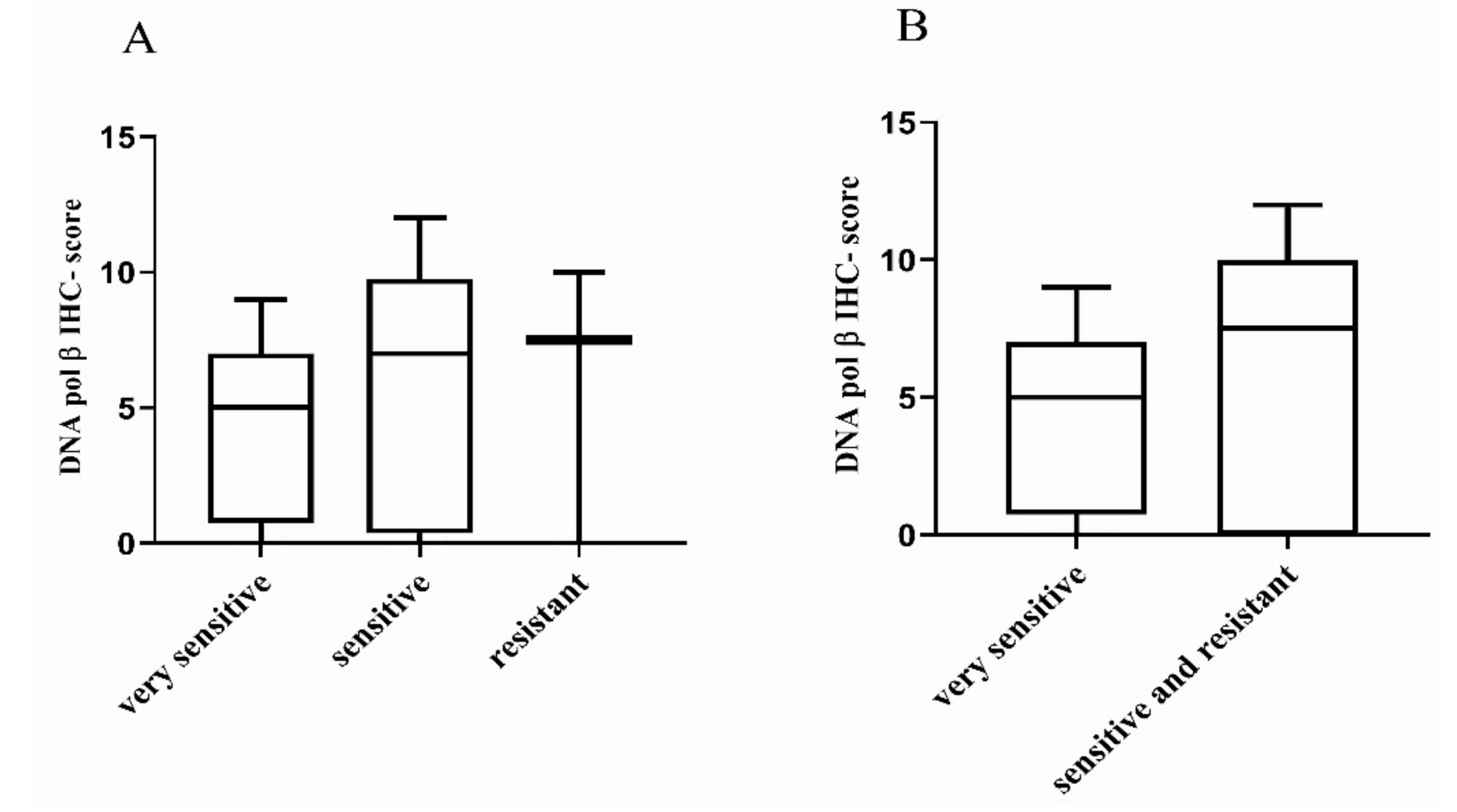
2.2. Correlation with Platinum Response
3. Discussion
4. Materials and Methods
4.1. Patient-Derived Ovarian Carcinoma Xenobank
4.2. DNA pol β and ERCC1 Immunohistochemistry (IHC) Expression in Ovarian OC-PDXs
4.3. Gene Expression
4.4. In Vitro Cell Culture and Drug Treatment
4.5. Proximity Ligation Assay (PLA) for the ERCC1/XPF Complex Detection
4.6. Statistical Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kelland, L. The resurgence of platinum-based cancer chemotherapy. Nat. Rev. Cancer 2007, 7, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Lheureux, S.; Braunstein, M.; Oza, A.M. Epithelial ovarian cancer: Evolution of management in the era of precision medicine. CA Cancer J. Clin. 2019, 69, 280–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slyskova, J.; Sabatella, M.; Ribeiro-Silva, C.; Stok, C.; Theil, A.F.; Vermeulen, W.; Lans, H. Base and nucleotide excision repair facilitate resolution of platinum drugs-induced transcription blockage. Nucleic Acids Res. 2018, 46, 9537–9549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, Y.; Lippard, S.J. Direct cellular responses to platinum-induced DNA damage. Chem. Rev. 2007, 107, 1387–1407. [Google Scholar] [CrossRef] [PubMed]
- Damia, G.; Imperatori, L.; Stefanini, M.; D’Incalci, M. Sensitivity of CHO mutant cell lines with specific defects in nucleotide excision repair to different anti-cancer agents. Int. J. Cancer 1996, 66, 779–783. [Google Scholar] [CrossRef]
- Kuraoka, I.; Kobertz, W.R.; Ariza, R.R.; Biggerstaff, M.; Essigmann, J.M.; Wood, R.D. Repair of an interstrand DNA cross-link initiated by ERCC1-XPF repair/recombination nuclease. J. Biol. Chem. 2000, 275, 26632–26636. [Google Scholar] [CrossRef] [Green Version]
- Macerelli, M.; Ganzinelli, M.; Gouedard, C.; Broggini, M.; Garassino, M.C.; Linardou, H.; Damia, G.; Wiesmuller, L. Can the response to a platinum-based therapy be predicted by the DNA repair status in non-small cell lung cancer? Cancer Treat. Rev. 2016, 48, 8–19. [Google Scholar] [CrossRef]
- Ganzinelli, M.; Mariani, P.; Cattaneo, D.; Fossati, R.; Fruscio, R.; Corso, S.; Ricci, F.; Broggini, M.; Damia, G. Expression of DNA repair genes in ovarian cancer samples: Biological and clinical considerations. Eur. J. Cancer 2011, 47, 1086–1094. [Google Scholar] [CrossRef]
- Postel-Vinay, S.; Soria, J.C. ERCC1 as Predictor of Platinum Benefit in Non-Small-Cell Lung Cancer. J. Clin. Oncol. 2017, 35, 384–386. [Google Scholar] [CrossRef]
- Mesquita, K.A.; Alabdullah, M.; Griffin, M.; Toss, M.S.; Fatah, T.; Alblihy, A.; Moseley, P.; Chan, S.Y.T.; Rakha, E.A.; Madhusudan, S. ERCC1-XPF deficiency is a predictor of olaparib induced synthetic lethality and platinum sensitivity in epithelial ovarian cancers. Gynecol. Oncol. 2019, 153, 416–424. [Google Scholar] [CrossRef]
- Lee, S.M.; Falzon, M.; Blackhall, F.; Spicer, J.; Nicolson, M.; Chaudhuri, A.; Middleton, G.; Ahmed, S.; Hicks, J.; Crosse, B. Randomized Prospective Biomarker Trial of ERCC1 for Comparing Platinum and Nonplatinum Therapy in Advanced Non-Small-Cell Lung Cancer: ERCC1 Trial (ET). J. Clin. Oncol. 2017, 35, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Friboulet, L.; Olaussen, K.A.; Pignon, J.P.; Shepherd, F.A.; Tsao, M.S.; Graziano, S.; Kratzke, R.; Douillard, J.Y.; Seymour, L.; Pirker, R.; et al. ERCC1 isoform expression and DNA repair in non-small-cell lung cancer. N. Engl. J. Med. 2013, 368, 1101–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friboulet, L.; Postel-Vinay, S.; Sourisseau, T.; Adam, J.; Stoclin, A.; Ponsonnailles, F.; Dorvault, N.; Commo, F.; Saulnier, P.; Salome-Desmoulez, S.; et al. ERCC1 function in nuclear excision and interstrand crosslink repair pathways is mediated exclusively by the ERCC1-202 isoform. Cell Cycle 2013, 12, 3298–3306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, M.S.; Adam, J.; Dorvault, N.; Robin, A.; Friboulet, L.; Soria, J.C.; Olaussen, K.A. A novel antibody-based approach to detect the functional ERCC1-202 isoform. DNA Repair (Amst.) 2018, 64, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Guffanti, F.; Fratelli, M.; Ganzinelli, M.; Bolis, M.; Ricci, F.; Bizzaro, F.; Chila, R.; Sina, F.P.; Fruscio, R.; Lupia, M.; et al. Platinum sensitivity and DNA repair in a recently established panel of patient-derived ovarian carcinoma xenografts. Oncotarget 2018, 9, 24707–24717. [Google Scholar] [CrossRef] [Green Version]
- David, S.S.; O’Shea, V.L.; Kundu, S. Base-excision repair of oxidative DNA damage. Nature 2007, 447, 941–950. [Google Scholar] [CrossRef] [Green Version]
- Sobol, R.W.; Prasad, R.; Evenski, A.; Baker, A.; Yang, X.P.; Horton, J.K.; Wilson, S.H. The lyase activity of the DNA repair protein beta-polymerase protects from DNA-damage-induced cytotoxicity. Nature 2000, 405, 807–810. [Google Scholar] [CrossRef]
- Wang, M.; Li, E.; Lin, L.; Kumar, A.K.; Pan, F.; He, L.; Zhang, J.; Hu, Z.; Guo, Z. Enhanced Activity of Variant DNA Polymerase beta (D160G) Contributes to Cisplatin Therapy by Impeding the Efficiency of NER. Mol. Cancer Res. 2019, 17, 2077–2088. [Google Scholar] [CrossRef] [Green Version]
- Canitrot, Y.; Capp, J.P.; Puget, N.; Bieth, A.; Lopez, B.; Hoffmann, J.S.; Cazaux, C. DNA polymerase beta overexpression stimulates the Rad51-dependent homologous recombination in mammalian cells. Nucleic Acids Res. 2004, 32, 5104–5112. [Google Scholar] [CrossRef]
- Ray, S.; Breuer, G.; DeVeaux, M.; Zelterman, D.; Bindra, R.; Sweasy, J.B. DNA polymerase beta participates in DNA End-joining. Nucleic Acids Res. 2018, 46, 242–255. [Google Scholar] [CrossRef]
- Horton, J.K.; Srivastava, D.K.; Zmudzka, B.Z.; Wilson, S.H. Strategic down-regulation of DNA polymerase beta by antisense RNA sensitizes mammalian cells to specific DNA damaging agents. Nucleic Acids Res. 1995, 23, 3810–3815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemec, A.A.; Abriola, L.; Merkel, J.S.; de Stanchina, E.; DeVeaux, M.; Zelterman, D.; Glazer, P.M.; Sweasy, J.B. DNA Polymerase Beta Germline Variant Confers Cellular Response to Cisplatin Therapy. Mol. Cancer Res. 2017, 15, 269–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, T.; Pan, F.; Cao, Y.; Han, Y.; Zhao, J.; Sun, H.; Zhou, X.; Wu, X.; He, L.; Hu, Z.; et al. R152C DNA Pol beta mutation impairs base excision repair and induces cellular transformation. Oncotarget 2016, 7, 6902–6915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albertella, M.R.; Lau, A.; O’Connor, M.J. The overexpression of specialized DNA polymerases in cancer. DNA Repair (Amst.) 2005, 4, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Canitrot, Y.; Cazaux, C.; Frechet, M.; Bouayadi, K.; Lesca, C.; Salles, B.; Hoffmann, J.S. Overexpression of DNA polymerase beta in cell results in a mutator phenotype and a decreased sensitivity to anticancer drugs. Proc. Natl. Acad. Sci. USA 1998, 95, 12586–12590. [Google Scholar] [CrossRef] [Green Version]
- Vaisman, A.; Chaney, S.G. The efficiency and fidelity of translesion synthesis past cisplatin and oxaliplatin GpG adducts by human DNA polymerase beta. J. Biol. Chem. 2000, 275, 13017–13025. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Hoffman, R.M. Human ovarian carcinoma metastatic models constructed in nude mice by orthotopic transplantation of histologically-intact patient specimens. Anticancer Res. 1993, 13, 283–286. [Google Scholar]
- Maru, Y.; Hippo, Y. Current Status of Patient-Derived Ovarian Cancer Models. Cells 2019, 8, 505. [Google Scholar] [CrossRef] [Green Version]
- Ricci, F.; Broggini, M.; Damia, G. Revisiting ovarian cancer preclinical models: Implications for a better management of the disease. Cancer Treat. Rev. 2013, 39, 561–568. [Google Scholar] [CrossRef]
- Palmer, A.C.; Plana, D.; Gao, H.; Korn, J.M.; Yang, G.; Green, J.; Zhang, X.; Velazquez, R.; McLaughlin, M.E.; Ruddy, D.A.; et al. A proof of concept for biomarker-guided targeted therapy against ovarian cancer based on patient-derived tumor xenografts. Cancer Res. 2020. [Google Scholar] [CrossRef]
- Ricci, F.; Bizzaro, F.; Cesca, M.; Guffanti, F.; Ganzinelli, M.; Decio, A.; Ghilardi, C.; Perego, P.; Fruscio, R.; Buda, A.; et al. Patient-derived ovarian tumor xenografts recapitulate human clinicopathology and genetic alterations. Cancer Res. 2014, 74, 6980–6990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran Chau, V.; Liu, W.; Gerbe de Thore, M.; Meziani, L.; Mondini, M.; O’Connor, M.J.; Deutsch, E.; Clemenson, C. Differential therapeutic effects of PARP and ATR inhibition combined with radiotherapy in the treatment of subcutaneous versus orthotopic lung tumour models. Br. J. Cancer 2020. [Google Scholar] [CrossRef] [PubMed]
- Kuo, T.H.; Kubota, T.; Watanabe, M.; Furukawa, T.; Kase, S.; Tanino, H.; Saikawa, Y.; Ishibiki, K.; Kitajima, M.; Hoffman, R.M. Site-specific chemosensitivity of human small-cell lung carcinoma growing orthotopically compared to subcutaneously in SCID mice: The importance of orthotopic models to obtain relevant drug evaluation data. Anticancer Res. 1993, 13, 627–630. [Google Scholar] [PubMed]
- Arbogast, S.; Behnke, S.; Opitz, I.; Stahel, R.A.; Seifert, B.; Weder, W.; Moch, H.; Soltermann, A. Automated ERCC1 immunohistochemistry in non-small cell lung cancer: Comparison of anti-ERCC1 antibodies 8F1, D-10, and FL-297. Appl. Immunohistochem. Mol. Morphol. 2011, 19, 99–105. [Google Scholar] [CrossRef]
- Olaussen, K.A.; Soria, J.C. Validation of ERCC1-XPF immunodetection—Letter. Cancer Res. 2010, 70, 3851–3852. [Google Scholar] [CrossRef] [Green Version]
- Konstantinopoulos, P.A.; Ceccaldi, R.; Shapiro, G.I.; D’Andrea, A.D. Homologous Recombination Deficiency: Exploiting the Fundamental Vulnerability of Ovarian Cancer. Cancer Discov. 2015, 5, 1137–1154. [Google Scholar] [CrossRef] [Green Version]
- Ceccaldi, R.; O’Connor, K.W.; Mouw, K.W.; Li, A.Y.; Matulonis, U.A.; D’Andrea, A.D.; Konstantinopoulos, P.A. A unique subset of epithelial ovarian cancers with platinum sensitivity and PARP inhibitor resistance. Cancer Res. 2015, 75, 628–634. [Google Scholar] [CrossRef] [Green Version]
- Mouw, K.W.; D’Andrea, A.D.; Konstantinopoulos, P.A. Nucleotide excision repair (NER) alterations as evolving biomarkers and therapeutic targets in epithelial cancers. Oncoscience 2015, 2, 942–943. [Google Scholar] [CrossRef]
- Tavecchio, M.; Simone, M.; Erba, E.; Chiolo, I.; Liberi, G.; Foiani, M.; D’Incalci, M.; Damia, G. Role of homologous recombination in trabectedin-induced DNA damage. Eur. J. Cancer 2008, 44, 609–618. [Google Scholar] [CrossRef]
- Caiola, E.; Salles, D.; Frapolli, R.; Lupi, M.; Rotella, G.; Ronchi, A.; Garassino, M.C.; Mattschas, N.; Colavecchio, S.; Broggini, M.; et al. Base excision repair-mediated resistance to cisplatin in KRAS(G12C) mutant NSCLC cells. Oncotarget 2015, 6, 30072–30087. [Google Scholar] [CrossRef] [Green Version]
- Dabholkar, M.; Bostick-Bruton, F.; Weber, C.; Bohr, V.A.; Egwuagu, C.; Reed, E. ERCC1 and ERCC2 expression in malignant tissues from ovarian cancer patients. J. Natl. Cancer Inst. 1992, 84, 1512–1517. [Google Scholar] [CrossRef] [PubMed]
- Steffensen, K.D.; Smoter, M.; Waldstrom, M.; Grala, B.; Bodnar, L.; Stec, R.; Szczylik, C.; Jakobsen, A. Resistance to first line platinum paclitaxel chemotherapy in serous epithelial ovarian cancer: The prediction value of ERCC1 and Tau expression. Int. J. Oncol. 2014, 44, 1736–1744. [Google Scholar] [CrossRef] [PubMed]
- Steffensen, K.D.; Waldstrom, M.; Jakobsen, A. The relationship of platinum resistance and ERCC1 protein expression in epithelial ovarian cancer. Int. J. Gynecol. Cancer 2009, 19, 820–825. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Dou, X.; Yang, H.; Jia, L.; Qin, K.; Gao, X.; Yang, B.; Zhang, W.; Qin, C.; Zhang, F.; et al. Association of expression of p53, livin, ERCC1, BRCA1 and PARP1 in epithelial ovarian cancer tissue with drug resistance and prognosis. Pathol. Res. Pract. 2019, 152794. [Google Scholar] [CrossRef]
- Rubatt, J.M.; Darcy, K.M.; Tian, C.; Muggia, F.; Dhir, R.; Armstrong, D.K.; Bookman, M.A.; Niedernhofer, L.J.; Deloia, J.; Birrer, M.; et al. Pre-treatment tumor expression of ERCC1 in women with advanced stage epithelial ovarian cancer is not predictive of clinical outcomes: A Gynecologic Oncology Group study. Gynecol. Oncol. 2012, 125, 421–426. [Google Scholar] [CrossRef] [Green Version]
- Graeser, M.; McCarthy, A.; Lord, C.J.; Savage, K.; Hills, M.; Salter, J.; Orr, N.; Parton, M.; Smith, I.E.; Reis-Filho, J.S.; et al. A marker of homologous recombination predicts pathologic complete response to neoadjuvant chemotherapy in primary breast cancer. Clin. Cancer Res. 2010, 16, 6159–6168. [Google Scholar] [CrossRef] [Green Version]
- Naipal, K.A.; Verkaik, N.S.; Ameziane, N.; van Deurzen, C.H.; Ter Brugge, P.; Meijers, M.; Sieuwerts, A.M.; Martens, J.W.; O’Connor, M.J.; Vrieling, H.; et al. Functional ex vivo assay to select homologous recombination-deficient breast tumors for PARP inhibitor treatment. Clin. Cancer Res. 2014, 20, 4816–4826. [Google Scholar] [CrossRef] [Green Version]
- Meijer, T.G.; Verkaik, N.S.; Sieuwerts, A.M.; van Riet, J.; Naipal, K.A.T.; van Deurzen, C.H.M.; den Bakker, M.A.; Sleddens, H.; Dubbink, H.J.; den Toom, T.D.; et al. Functional Ex Vivo Assay Reveals Homologous Recombination Deficiency in Breast Cancer Beyond BRCA Gene Defects. Clin. Cancer Res. 2018, 24, 6277–6287. [Google Scholar] [CrossRef] [Green Version]
- Scurry, J.; van Zyl, B.; Gulliver, D.; Otton, G.; Jaaback, K.; Lombard, J.; Vilain, R.E.; Bowden, N.A. Nucleotide excision repair protein ERCC1 and tumour-infiltrating lymphocytes are potential biomarkers of neoadjuvant platinum resistance in high grade serous ovarian cancer. Gynecol. Oncol. 2018, 151, 306–310. [Google Scholar] [CrossRef]
- Damia, G.; Broggini, M. Platinum Resistance in Ovarian Cancer: Role of DNA Repair. Cancers 2019, 11. [Google Scholar] [CrossRef] [Green Version]
- Marusyk, A.; Janiszewska, M.; Polyak, K. Intratumor Heterogeneity: The Rosetta Stone of Therapy Resistance. Cancer Cell 2020, 37, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Winterhoff, B.; Talukdar, S.; Chang, Z.; Wang, J.; Starr, T.K. Single-cell sequencing in ovarian cancer: A new frontier in precision medicine. Curr. Opin. Obstet. Gynecol. 2019, 31, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Sawant, A.; Floyd, A.M.; Dangeti, M.; Lei, W.; Sobol, R.W.; Patrick, S.M. Differential role of base excision repair proteins in mediating cisplatin cytotoxicity. DNA Repair (Amst.) 2017, 51, 46–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Parsons, J.; Nicolay, N.H.; Caporali, S.; Harrington, C.F.; Singh, R.; Finch, D.; D’Atri, S.; Farmer, P.B.; Johnston, P.G.; et al. Cells deficient in the base excision repair protein, DNA polymerase beta, are hypersensitive to oxaliplatin chemotherapy. Oncogene 2010, 29, 463–468. [Google Scholar] [CrossRef]
- Bergoglio, V.; Canitrot, Y.; Hogarth, L.; Minto, L.; Howell, S.B.; Cazaux, C.; Hoffmann, J.S. Enhanced expression and activity of DNA polymerase beta in human ovarian tumor cells: Impact on sensitivity towards antitumor agents. Oncogene 2001, 20, 6181–6187. [Google Scholar] [CrossRef] [Green Version]
- Pilla, D.; Bosisio, F.M.; Marotta, R.; Faggi, S.; Forlani, P.; Falavigna, M.; Biunno, I.; Martella, E.; De Blasio, P.; Borghesi, S.; et al. Tissue microarray design and construction for scientific, industrial and diagnostic use. J. Pathol. Inform. 2012, 3, 42. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| OC-PDXs | Descriptive Statistics | IHC-Score | PLA | Normalized Gene Expression Levels | |||
|---|---|---|---|---|---|---|---|
| ERCC1 | DNA pol β | ERCC1/XPFFoci Number | ERCC1 | XPF | DNA pol β | ||
| All PDXs (n = 52) | Mean (SD) | 1.4(0.9) | 5.5 (4.2) | 5.0 (2.0) | 1.1 (0.8) | 0.0025 (0.0016) | 0.0448 (0.0336) |
| Min–Max | 0.0–3.5 | 0.0–12.0 | 0.9–8.8 | 0.2–3.1 | 0.0005–0.009 | 0.004–0.1619 | |
| Missing | 3 | 1 | 3 | 16 | 16 | 16 | |
| High-Grade PDXs (n = 41) | Mean (SD) | 1.2 (1.0) | 5.3 (4.0) | 5.1 (2.2) | 1.2 (0.8) | 0.0027 (0.0018) | 0.0498 (0.0374) |
| Min–Max | 0.0–3.5 | 0.0–12.0 | 0.9–8.8 | 0.2–3.1 | 0.0009–0.0090 | 0.0040–0.1619 | |
| Missing | 1 | 1 | 2 | 2 | 2 | 2 | |
| Method | Molecular Target | IHC-Score | PLA | Normalized Gene Expression Levels | |||
|---|---|---|---|---|---|---|---|
| ERCC1 | DNA pol β | ERCC1/XPF Foci Number | ERCC1 | XPF | DNA pol β | ||
| IHC-Score | ERCC1 | 1 | |||||
| 39 | |||||||
| DNA pol β | 0.03 | 1 | |||||
| 0.85 | |||||||
| 38 | 40 | ||||||
| PLA | ERCC1/XPF Foci Number | 0.26 | 0.62 | 1 | |||
| 0.13 | <0.0001 | ||||||
| 36 | 37 | 38 | |||||
| Normalized Gene Expression Levels by RT-PCR | ERCC1 | 0.02 | −0.07 | 0.15 | 1 | ||
| 0.94 | 0.72 | 0.47 | |||||
| 25 | 25 | 25 | 26 | ||||
| XPF | 0.69 | −0.05 | 0.19 | 0.04 | 1 | ||
| 0.0001 | 0.83 | 0.36 | 0.83 | ||||
| 25 | 25 | 25 | 26 | 26 | |||
| DNA pol β | 0.11 | 0.17 | 0.12 | 0.24 | 0.36 | 1 | |
| 0.62 | 0.42 | 0.58 | 0.24 | 0.07 | |||
| 25 | 25 | 25 | 26 | 26 | 26 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guffanti, F.; Alvisi, M.F.; Caiola, E.; Ricci, F.; De Maglie, M.; Soldati, S.; Ganzinelli, M.; Decio, A.; Giavazzi, R.; Rulli, E.; et al. Impact of ERCC1, XPF and DNA Polymerase β Expression on Platinum Response in Patient-Derived Ovarian Cancer Xenografts. Cancers 2020, 12, 2398. https://doi.org/10.3390/cancers12092398
Guffanti F, Alvisi MF, Caiola E, Ricci F, De Maglie M, Soldati S, Ganzinelli M, Decio A, Giavazzi R, Rulli E, et al. Impact of ERCC1, XPF and DNA Polymerase β Expression on Platinum Response in Patient-Derived Ovarian Cancer Xenografts. Cancers. 2020; 12(9):2398. https://doi.org/10.3390/cancers12092398
Chicago/Turabian StyleGuffanti, Federica, Maria Francesca Alvisi, Elisa Caiola, Francesca Ricci, Marcella De Maglie, Sabina Soldati, Monica Ganzinelli, Alessandra Decio, Raffaella Giavazzi, Eliana Rulli, and et al. 2020. "Impact of ERCC1, XPF and DNA Polymerase β Expression on Platinum Response in Patient-Derived Ovarian Cancer Xenografts" Cancers 12, no. 9: 2398. https://doi.org/10.3390/cancers12092398
APA StyleGuffanti, F., Alvisi, M. F., Caiola, E., Ricci, F., De Maglie, M., Soldati, S., Ganzinelli, M., Decio, A., Giavazzi, R., Rulli, E., & Damia, G. (2020). Impact of ERCC1, XPF and DNA Polymerase β Expression on Platinum Response in Patient-Derived Ovarian Cancer Xenografts. Cancers, 12(9), 2398. https://doi.org/10.3390/cancers12092398