Jagged Ligands Enhance the Pro-Angiogenic Activity of Multiple Myeloma Cells

 , , , , , , ,
, , , , , , ,  ,
, 
 and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
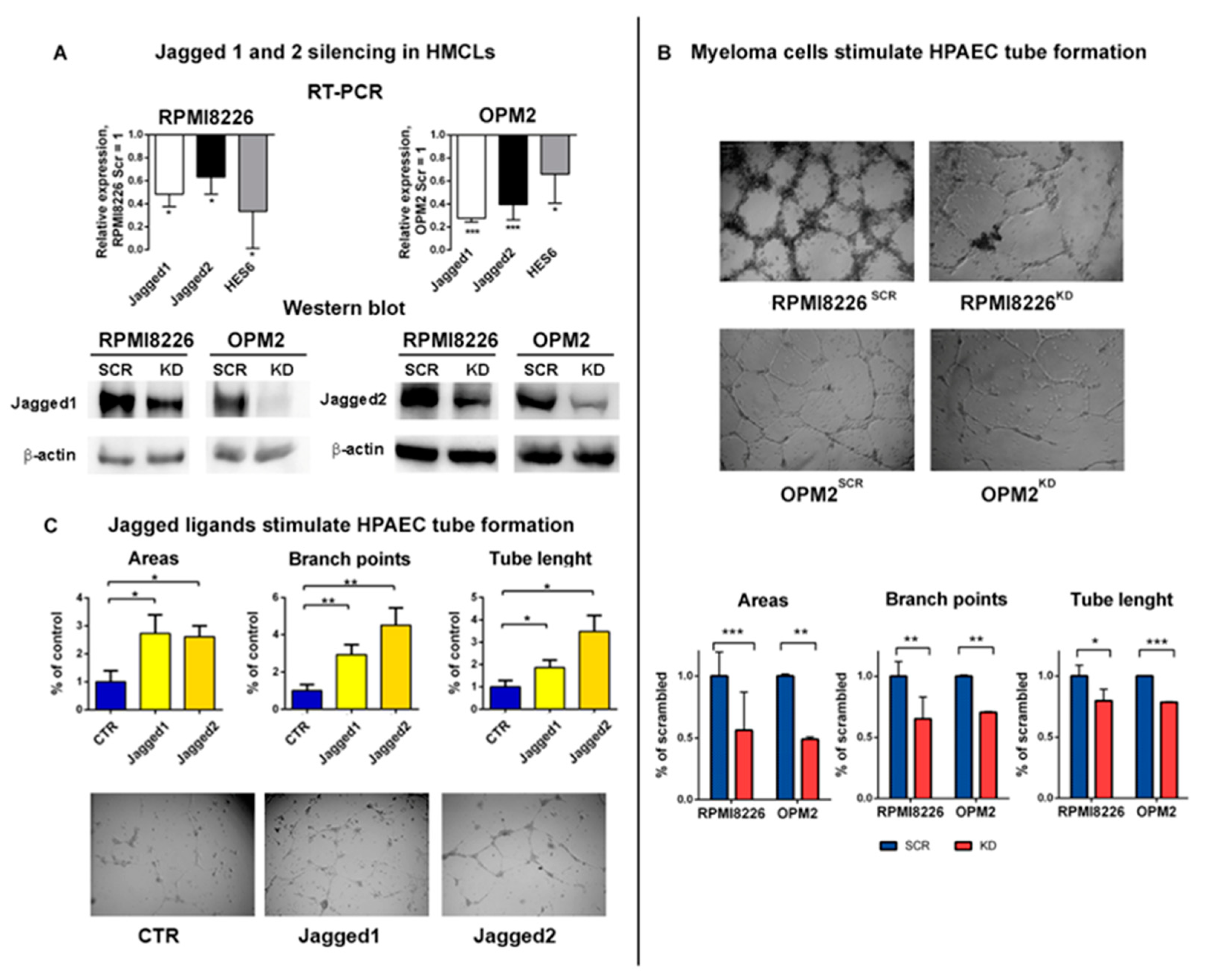
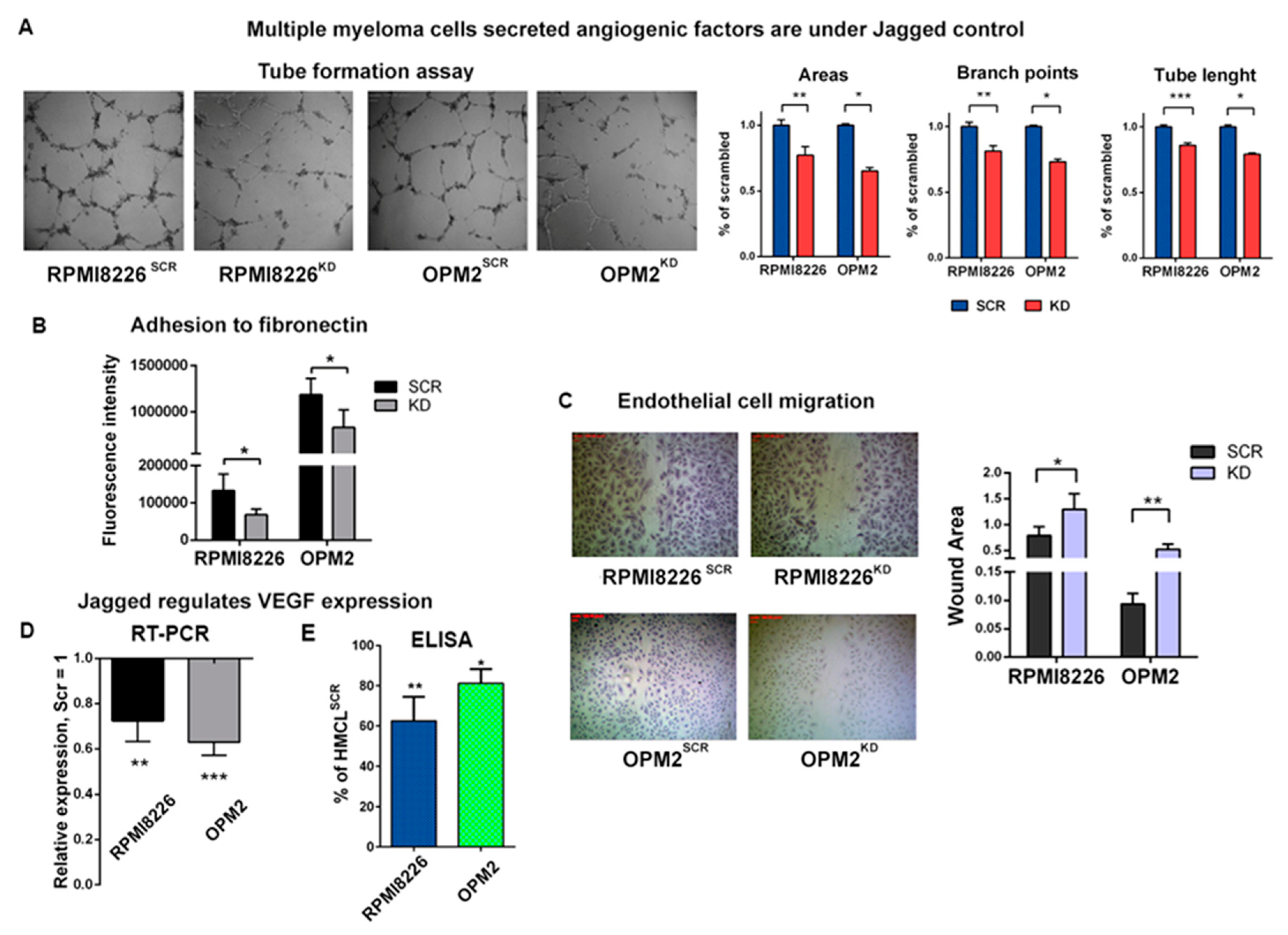
2.1. Myeloma Cell-Derived Jagged Ligands Regu Late Myeloma Cell Angiogenic Potential
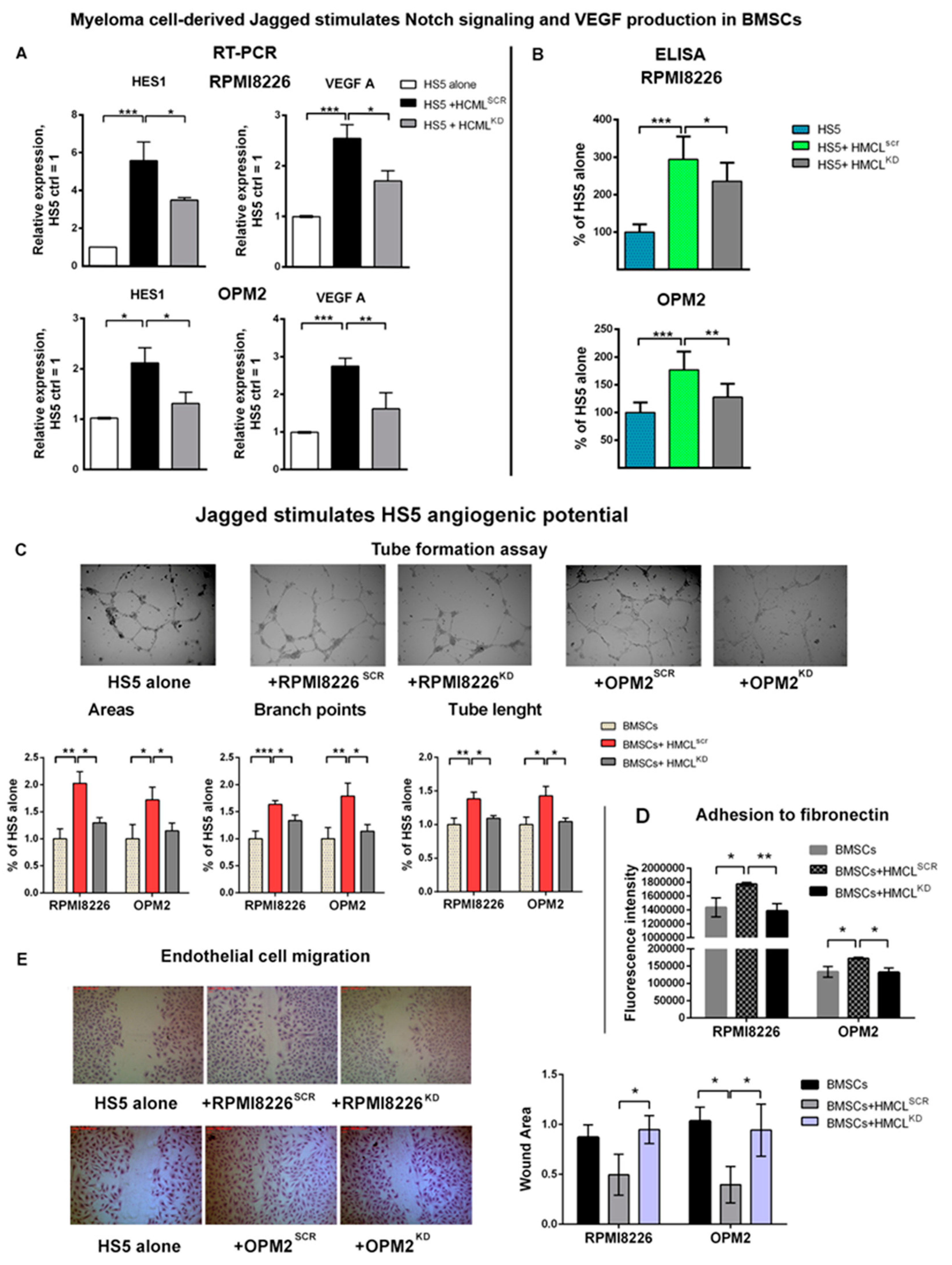
2.2. Myeloma Cell-Derived Jagged Ligands Increase the Angiogenic Potential of Stromal Cells
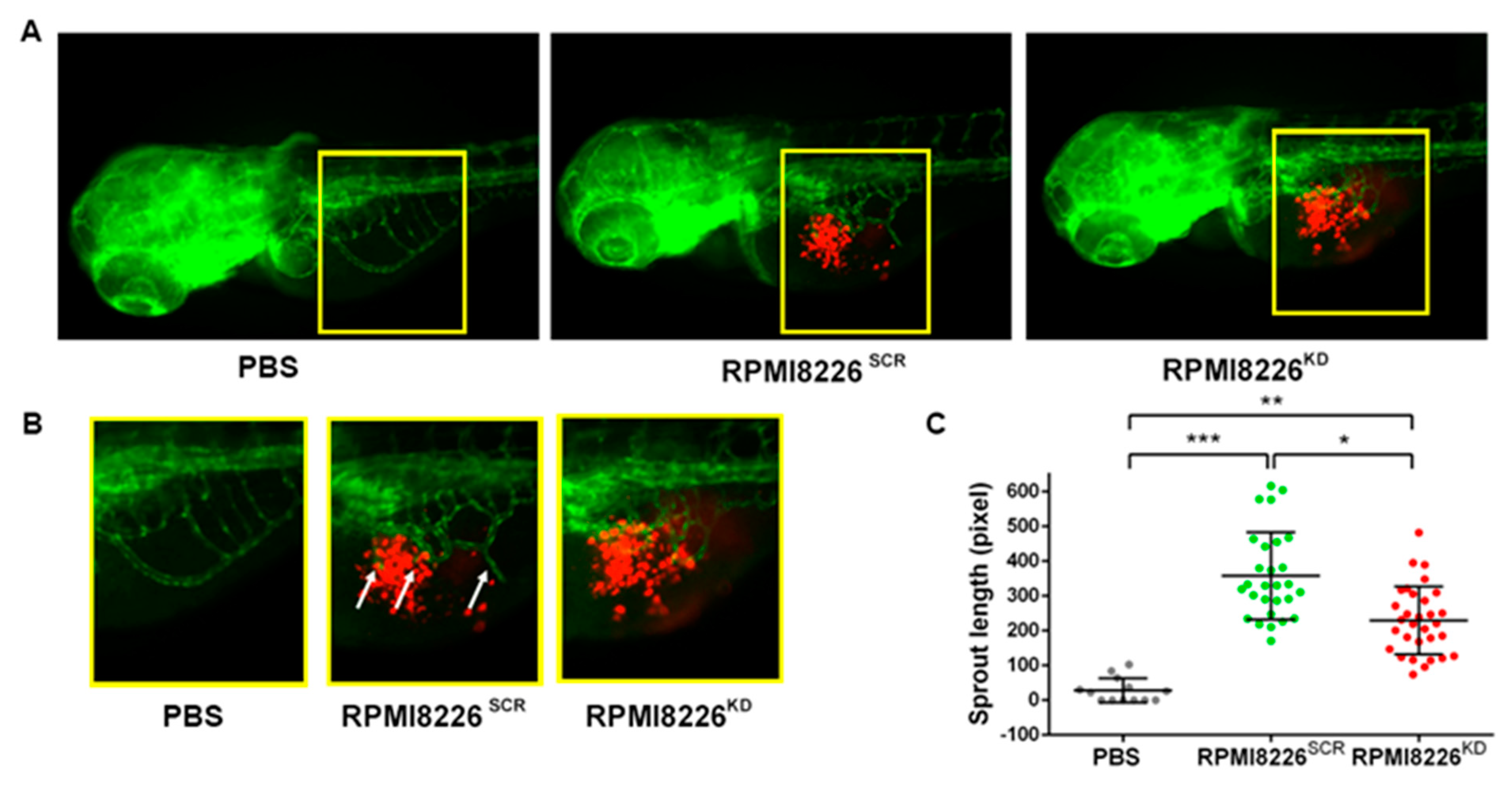
2.3. Jagged Ligands Promote MM-Associated Angiogenesis in a Zebrafish Model
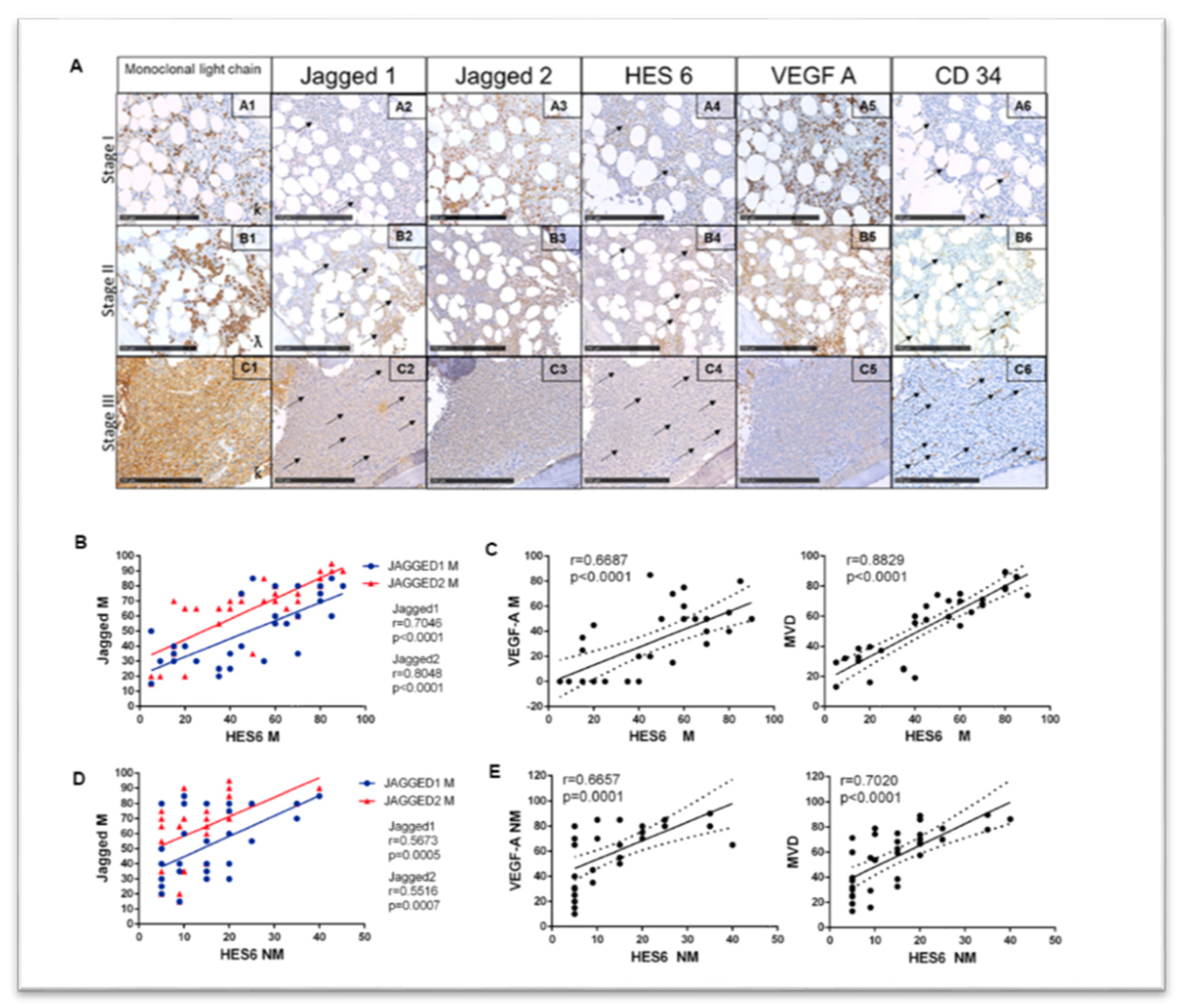
2.4. Identification of a Correlation between Jagged Expression in MM Cells and Angiogenesis in Patients’ Bone Marrow
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. HMCL Knockdown and Co-Culture Experiments
4.3. Tube Formation Assay
4.4. Adhesion Assay
4.5. Wound Healing Assay
4.6. RNA Extraction and qRT-PCR
4.7. ELISA for VEGF-A
4.8. Zebrafish Injection
4.9. Immunohistochemical Analysis of Human Bone Marrow Biopsies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ribatti, D.; Vacca, A. Role of Endothelial Cells and Fibroblasts in Multiple Myeloma Angiogenic Switch. Cancer Treat. Res. 2016, 169, 51–61. [Google Scholar]
- Hideshima, T.; Mitsiades, C.; Tonon, G.; Richardson, P.G.; Anderson, K.C. Understanding multiple myeloma pathogenesis in the bone marrow to identify new therapeutic targets. Nat. Rev. Cancer 2007, 7, 585–598. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Vacca, A. The role of microenvironment in tumor angiogenesis. Genes Nutr. 2008, 3, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ria, R.; Reale, A.; De Luisi, A.; Ferrucci, A.; Moschetta, M.; Vacca, A. Bone marrow angiogenesis and progression in multiple myeloma. Am. J. Blood Res. 2011, 1, 76–89. [Google Scholar] [PubMed]
- Vacca, A.; Ria, R.; Semeraro, F.; Merchionne, F.; Coluccia, M.; Boccarelli, A.; Scavelli, C.; Nico, B.; Gernone, A.; Battelli, F.; et al. Endothelial cells in the bone marrow of patients with multiple myeloma. Blood 2003, 102, 3340–3348. [Google Scholar] [CrossRef] [Green Version]
- Saltarella, I.; Frassanito, M.A.; Lamanuzzi, A.; Brevi, A.; Leone, P.; Desantis, V.; Di Marzo, L.; Bellone, M.; Derudas, D.; Ribatti, D.; et al. Homotypic and Heterotypic Activation of the Notch Pathway in Multiple Myeloma-Enhanced Angiogenesis: A Novel Therapeutic Target? Neoplasia 2019, 21, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Galletti, S.; Bulfamante, G.; Falleni, M.; Tosi, D.; Todoerti, K.; Lazzari, E.; Crews, L.A.; Jamieson, C.H.; Ravaioli, S.; et al. Multiple myeloma-derived Jagged ligands increases autocrine and paracrine interleukin-6 expression in bone marrow niche. Oncotarget 2016, 7, 56013–56029. [Google Scholar] [CrossRef] [Green Version]
- Platonova, N.; Lesma, E.; Basile, A.; Bignotto, M.; Garavelli, S.; Palano, M.T.; Moschini, A.; Neri, A.; Colombo, M.; Chiaramonte, R. Targeting Notch as a Therapeutic Approach for Human Malignancies. Curr. Pharm. Des. 2017, 23, 108–134. [Google Scholar]
- Kopan, R.; Ilagan, M.X. The canonical Notch signaling pathway: Unfolding the activation mechanism. Cell 2009, 137, 216–233. [Google Scholar] [CrossRef] [Green Version]
- Colombo, M.; Garavelli, S.; Mazzola, M.; Platonova, N.; Giannandrea, D.; Colella, R.; Apicella, L.; Lancellotti, M.; Lesma, E.; Ancona, S.; et al. Multiple myeloma exploits Jagged1 and Jagged2 to promote intrinsic and bone marrow-dependent drug resistance. Haematologica 2019, 105, e1925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nefedova, Y.; Cheng, P.; Alsina, M.; Dalton, W.S.; Gabrilovich, D.I. Involvement of Notch-1 signaling in bone marrow stroma-mediated de novo drug resistance of myeloma and other malignant lymphoid cell lines. Blood 2004, 103, 3503–3510. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Thummler, K.; Mirandola, L.; Garavelli, S.; Todoerti, K.; Apicella, L.; Lazzari, E.; Lancellotti, M.; Platonova, N.; Akbar, M.; et al. Notch signaling drives multiple myeloma induced osteoclastogenesis. Oncotarget 2014, 5, 10393–10406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houde, C.; Li, Y.; Song, L.; Barton, K.; Zhang, Q.; Godwin, J.; Nand, S.; Toor, A.; Alkan, S.; Smadja, N.V.; et al. Overexpression of the NOTCH ligand JAG2 in malignant plasma cells from multiple myeloma patients and cell lines. Blood 2004, 104, 3697–3704. [Google Scholar] [CrossRef] [Green Version]
- Skrtic, A.; Korac, P.; Kristo, D.R.; Ajdukovic Stojisavljevic, R.; Ivankovic, D.; Dominis, M. Immunohistochemical analysis of NOTCH1 and JAGGED1 expression in multiple myeloma and monoclonal gammopathy of undetermined significance. Hum. Pathol. 2010, 41, 1702–1710. [Google Scholar] [CrossRef] [PubMed]
- Pedrosa, A.R.; Trindade, A.; Fernandes, A.C.; Carvalho, C.; Gigante, J.; Tavares, A.T.; Dieguez-Hurtado, R.; Yagita, H.; Adams, R.H.; Duarte, A. Endothelial Jagged1 antagonizes Dll4 regulation of endothelial branching and promotes vascular maturation downstream of Dll4/Notch1. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1134–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kangsamaksin, T.; Tattersall, I.W.; Kitajewski, J. Notch functions in developmental and tumour angiogenesis by diverse mechanisms. Biochem. Soc. Trans. 2014, 42, 1563–1568. [Google Scholar] [CrossRef]
- Bridges, E.; Oon, C.E.; Harris, A. Notch regulation of tumor angiogenesis. Future Oncol. (Lond. Engl.) 2011, 7, 569–588. [Google Scholar] [CrossRef]
- Ghiabi, P.; Jiang, J.; Pasquier, J.; Maleki, M.; Abu-Kaoud, N.; Rafii, S.; Rafii, A. Endothelial cells provide a notch-dependent pro-tumoral niche for enhancing breast cancer survival, stemness and pro-metastatic properties. PLoS ONE 2014, 9, e112424. [Google Scholar] [CrossRef]
- Phng, L.K.; Gerhardt, H. Angiogenesis: A team effort coordinated by notch. Dev. Cell 2009, 16, 196–208. [Google Scholar] [CrossRef]
- Rajkumar, S.V.; Leong, T.; Roche, P.C.; Fonseca, R.; Dispenzieri, A.; Lacy, M.Q.; Lust, J.A.; Witzig, T.E.; Kyle, R.A.; Gertz, M.A.; et al. Prognostic value of bone marrow angiogenesis in multiple myeloma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2000, 6, 3111–3116. [Google Scholar]
- Vande Broek, I.; Vanderkerken, K.; Van Camp, B.; Van Riet, I. Extravasation and homing mechanisms in multiple myeloma. Clin. Exp. Metastasis 2008, 25, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, A.; Ria, R.; Di Pietro, G.; Cirulli, T.; Surico, G.; Pennisi, A.; Morabito, F.; Ribatti, D.; Vacca, A. Bone marrow endothelial cells in multiple myeloma secrete CXC-chemokines that mediate interactions with plasma cells. Br. J. Haematol. 2005, 129, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Moschetta, M.; Vacca, A. Microenvironment and multiple myeloma spread. Thromb. Res. 2014, 133, S102–S106. [Google Scholar] [CrossRef]
- Kawano, Y.; Moschetta, M.; Manier, S.; Glavey, S.; Gorgun, G.T.; Roccaro, A.M.; Anderson, K.C.; Ghobrial, I.M. Targeting the bone marrow microenvironment in multiple myeloma. Immunol. Rev. 2015, 263, 160–172. [Google Scholar] [CrossRef]
- Hori, K.; Sen, A.; Artavanis-Tsakonas, S. Notch signaling at a glance. J. Cell Sci. 2013, 126, 2135–2140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muguruma, Y.; Yahata, T.; Warita, T.; Hozumi, K.; Nakamura, Y.; Suzuki, R.; Ito, M.; Ando, K. Jagged1-induced Notch activation contributes to the acquisition of bortezomib resistance in myeloma cells. Blood Cancer J. 2017, 7, e650. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Hu, J.; De Bruyne, E.; Menu, E.; Schots, R.; Vanderkerken, K.; Van Valckenborgh, E. Dll1/Notch activation contributes to bortezomib resistance by upregulating CYP1A1 in multiple myeloma. Biochem. Biophys. Res. Commun. 2012, 428, 518–524. [Google Scholar] [CrossRef]
- Nefedova, Y.; Sullivan, D.M.; Bolick, S.C.; Dalton, W.S.; Gabrilovich, D.I. Inhibition of Notch signaling induces apoptosis of myeloma cells and enhances sensitivity to chemotherapy. Blood 2008, 111, 2220–2229. [Google Scholar] [CrossRef] [Green Version]
- Mirandola, L.; Apicella, L.; Colombo, M.; Yu, Y.; Berta, D.G.; Platonova, N.; Lazzari, E.; Lancellotti, M.; Bulfamante, G.; Cobos, E.; et al. Anti-Notch treatment prevents multiple myeloma cells localization to the bone marrow via the chemokine system CXCR4/SDF-1. Leukemia 2013, 27, 1558–1566. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Calle, J.; Anderson, J.; Cregor, M.D.; Hiasa, M.; Chirgwin, J.M.; Carlesso, N.; Yoneda, T.; Mohammad, K.S.; Plotkin, L.I.; Roodman, G.D.; et al. Bidirectional Notch Signaling and Osteocyte-Derived Factors in the Bone Marrow Microenvironment Promote Tumor Cell Proliferation and Bone Destruction in Multiple Myeloma. Cancer Res. 2016, 76, 1089–1100. [Google Scholar] [CrossRef] [Green Version]
- Ghoshal, P.; Nganga, A.J.; Moran-Giuati, J.; Szafranek, A.; Johnson, T.R.; Bigelow, A.J.; Houde, C.M.; Avet-Loiseau, H.; Smiraglia, D.J.; Ersing, N.; et al. Loss of the SMRT/NCoR2 corepressor correlates with JAG2 overexpression in multiple myeloma. Cancer Res. 2009, 69, 4380–4387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, T.; Adachi, Y.; Ohtsuki, Y. Skeletrophin, a novel ubiquitin ligase to the intracellular region of Jagged-2, is aberrantly expressed in multiple myeloma. Am. J. Pathol. 2005, 166, 1817–1826. [Google Scholar] [CrossRef] [Green Version]
- Vacca, A.; Ria, R.; Reale, A.; Ribatti, D. Angiogenesis in multiple myeloma. Chem. Immunol. Allergy 2014, 99, 180–196. [Google Scholar] [PubMed]
- Lawson, N.D.; Weinstein, B.M. In vivo imaging of embryonic vascular development using transgenic zebrafish. Dev. Biol. 2002, 248, 307–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitale, G.; Gaudenzi, G.; Dicitore, A.; Cotelli, F.; Ferone, D.; Persani, L. Zebrafish as an innovative model for neuroendocrine tumors. Endocr. Related. Cancer 2014, 21, R67–R83. [Google Scholar] [CrossRef]
- Chen, J.Y.; Li, C.F.; Chu, P.Y.; Lai, Y.S.; Chen, C.H.; Jiang, S.S.; Hou, M.F.; Hung, W.C. Lysine demethylase 2A promotes stemness and angiogenesis of breast cancer by upregulating Jagged1. Oncotarget 2016, 7, 27689–27710. [Google Scholar] [CrossRef] [Green Version]
- Pietras, A.; von Stedingk, K.; Lindgren, D.; Pahlman, S.; Axelson, H. JAG2 induction in hypoxic tumor cells alters Notch signaling and enhances endothelial cell tube formation. Mol. Cancer Res. 2011, 9, 626–636. [Google Scholar] [CrossRef] [Green Version]
- Soares, R.; Balogh, G.; Guo, S.; Gartner, F.; Russo, J.; Schmitt, F. Evidence for the notch signaling pathway on the role of estrogen in angiogenesis. Mol. Endocrinol. (Baltim. Md.) 2004, 18, 2333–2343. [Google Scholar] [CrossRef]
- Zeng, Q.; Li, S.; Chepeha, D.B.; Giordano, T.J.; Li, J.; Zhang, H.; Polverini, P.J.; Nor, J.; Kitajewski, J.; Wang, C.Y. Crosstalk between tumor and endothelial cells promotes tumor angiogenesis by MAPK activation of Notch signaling. Cancer Cell 2005, 8, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Boareto, M.; Jolly, M.K.; Ben-Jacob, E.; Onuchic, J.N. Jagged mediates differences in normal and tumor angiogenesis by affecting tip-stalk fate decision. Proc. Natl. Acad. Sci. USA 2015, 112, E3836–E3844. [Google Scholar] [CrossRef] [Green Version]
- Masiero, M.; Li, D.; Whiteman, P.; Bentley, C.; Greig, J.; Hassanali, T.; Watts, S.; Stribbling, S.; Yates, J.; Bealing, E.; et al. Development of Therapeutic Anti-JAGGED1 Antibodies for Cancer Therapy. Mol. Cancer Ther. 2019, 18, 2030–2042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platonova, N.; Parravicini, C.; Sensi, C.; Paoli, A.; Colombo, M.; Neri, A.; Eberini, I.; Chiaramonte, R. Identification of small molecules uncoupling the Notch:Jagged interaction through an integrated high-throughput screening. PLoS ONE 2017, 12, e0182640. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PATIENT N. | INFILTRATION DEGREE | LIGHT CHAIN | JAGGED1 M | JAGGED1 NM | JAGGED2 M | JAGGED2 NM | HES6 M | HES6 NM | VEGF-A M | VEGF-A NM | MVD |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | I | K | 50 | 5 | 20 | 15 | 5 | 5 | 0 | 30 | 13.0 |
| 2 | I | L | 30 | 5 | 35 | 40 | 15 | 5 | 0 | 40 | 30.3 |
| 3 | I | L | 30 | 9 | 20 | 25 | 9 | 5 | 0 | 15 | 32.0 |
| 4 | I | L | 40 | 9 | 20 | 20 | 20 | 9 | 0 | 35 | 16.0 |
| 5 | I | L | 15 | 9 | 15 | 20 | 5 | 9 | 0 | 35 | 29.3 |
| 6 | I | L | 20 | 5 | 65 | 35 | 35 | 5 | 0 | 31 | 24.7 |
| 7 | I | K | 25 | 5 | 70 | 40 | 40 | 5 | 0 | 10 | 19.0 |
| 8 | I | K | 35 | 9 | 65 | 80 | 40 | 9 | 0 | 45 | 55.7 |
| 9 | I | L | 30 | 9 | 65 | 60 | 25 | 5 | 0 | 20 | 37.3 |
| 10 | I | L | 25 | 9 | 55 | 30 | 35 | 5 | 0 | 25 | 25.3 |
| 11 | II | L | 25 | 15 | 65 | 40 | 40 | 5 | 20 | 70 | 60.0 |
| 12 | II | K | 30 | 15 | 70 | 45 | 55 | 15 | 15 | 50 | 59.7 |
| 13 | II | L | 35 | 20 | 70 | 70 | 15 | 15 | 35 | 65 | 38.7 |
| 14 | II | K | 40 | 35 | 65 | 75 | 20 | 5 | 45 | 80 | 39.7 |
| 15 | II | K | 40 | 25 | 70 | 50 | 45 | 20 | 20 | 75 | 57.7 |
| 16 | II | K | 40 | 25 | 40 | 55 | 15 | 15 | 25 | 55 | 32.7 |
| 17 | II | K | 85 | 15 | 35 | 80 | 50 | 10 | 50 | 70 | 74.3 |
| 18 | II | K | 60 | 40 | 70 | 75 | 60 | 10 | 60 | 70 | 53.7 |
| 19 | II | K | 80 | 30 | 75 | 55 | 70 | 5 | 30 | 65 | 71.3 |
| 20 | II | L | 55 | 25 | 65 | 80 | 65 | 15 | 50 | 65 | 62.7 |
| 21 | II | K | 60 | 50 | 70 | 85 | 70 | 20 | 40 | 70 | 67.3 |
| 22 | III | K | 30 | 25 | 85 | 85 | 55 | 20 | 70 | 80 | 70.3 |
| 23 | III | K | 80 | 55 | 75 | 90 | 60 | 15 | 75 | 85 | 75.0 |
| 24 | III | L | 65 | 25 | 60 | 80 | 70 | 15 | 50 | 85 | 68.3 |
| 25 | III | L | 80 | 30 | 90 | 85 | 80 | 10 | 55 | 85 | 79.0 |
| 26 | III | K | 70 | 30 | 95 | 90 | 85 | 20 | 80 | 80 | 86.0 |
| 27 | III | K | 75 | 25 | 75 | 80 | 45 | 20 | 85 | 70 | 66.7 |
| 28 | III | K | 55 | 30 | 80 | 80 | 60 | 25 | 50 | 85 | 70.0 |
| 29 | III | K | 80 | 50 | 80 | 90 | 80 | 35 | 40 | 90 | 78.0 |
| 30 | III | K | 80 | 30 | 80 | 95 | N.A. | 25 | 45 | 80 | 79.0 |
| 31 | III | K | 75 | 25 | 85 | 95 | 80 | 20 | 40 | 80 | 89.0 |
| 32 | III | L | 85 | 55 | 90 | 90 | 85 | 40 | 80 | 65 | 86.3 |
| 33 | III | K | 70 | 45 | 80 | 85 | 80 | 35 | 40 | 80 | 89.7 |
| 34 | III | K | 80 | 30 | 90 | 85 | 90 | 20 | 50 | 70 | 74.0 |
| hGAPDH | 5′-ACAGTCAGCCG ATC TTC TT-3′ | 5′-AATGGAGGGGTCATTGATGG-3′ |
|---|---|---|
| h18S | 5′-GTAACCCGTTGAACCCCATT-3′ | 5′-CCATCCAATCGGTAGTAGCG-3′ |
| hJagged1 | 5′-GCAACACCTTCAACCTCAAG-3′ | 5′-GTTGAACGGTGTCATTACTGG-3′ |
| hJagged2 | 5′-TCATCCCCTTCCAGTTCG-3′ | 5′-TGGTATCGTTGTCCCAGTC-3′ |
| hHES1 | 5′-AGGCGGACATTCTGGAAATG-3′ | 5′-CGGTACTTCCCCAGCACACTT-3′ |
| hHES6 | 5′-CGTGAGGATGAGGACGG-3′ | 5′-AGGCTCTCGTTGATCCG-3′ |
| hVEGF-A | 5′-GGGCAGAATCATCACGAAGT-3′ | 5′-TGGTGATGTTGGACTCCTCA-3′ |
| hHPRT | 5′-TTTATGTCCCCTGTTGACTGGT-3′ | 5′-GTAGCCCTCTGTGTGCTCAA-3′ |
| Antigen | Clone | Source | Dilution/Time | Unmasking |
|---|---|---|---|---|
| Kappa light chain | - | Agilent | 1:10; 1 h | FLEX TRS Low pH |
| Lambda light chain | - | Agilent | 1:10; 1 h | FLEX TRS Low pH |
| Jagged 1 | AF1277 Goat | R&D syst. | 1:100; 1 h | EDTA |
| Jagged 2 | 4F10 Mouse | Santa Cruz | 1:200; o.n. | Citrate |
| HES6 | Polyclonal Rabbit | Abcam | 1:300; o.n. | Citrate |
| VEGF-A | A-20 Polyclonal Rabbit | Santa Cruz | 1:800; 1 h | Citrate |
| CD34 | QBEnd 10 Monoclonal Mouse | Agilent | Ready-to-Use | FLEX TRS Low pH |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palano, M.T.; Giannandrea, D.; Platonova, N.; Gaudenzi, G.; Falleni, M.; Tosi, D.; Lesma, E.; Citro, V.; Colombo, M.; Saltarella, I.; et al. Jagged Ligands Enhance the Pro-Angiogenic Activity of Multiple Myeloma Cells. Cancers 2020, 12, 2600. https://doi.org/10.3390/cancers12092600
Palano MT, Giannandrea D, Platonova N, Gaudenzi G, Falleni M, Tosi D, Lesma E, Citro V, Colombo M, Saltarella I, et al. Jagged Ligands Enhance the Pro-Angiogenic Activity of Multiple Myeloma Cells. Cancers. 2020; 12(9):2600. https://doi.org/10.3390/cancers12092600
Chicago/Turabian StylePalano, Maria Teresa, Domenica Giannandrea, Natalia Platonova, Germano Gaudenzi, Monica Falleni, Delfina Tosi, Elena Lesma, Valentina Citro, Michela Colombo, Ilaria Saltarella, and et al. 2020. "Jagged Ligands Enhance the Pro-Angiogenic Activity of Multiple Myeloma Cells" Cancers 12, no. 9: 2600. https://doi.org/10.3390/cancers12092600
APA StylePalano, M. T., Giannandrea, D., Platonova, N., Gaudenzi, G., Falleni, M., Tosi, D., Lesma, E., Citro, V., Colombo, M., Saltarella, I., Ria, R., Amodio, N., Taiana, E., Neri, A., Vitale, G., & Chiaramonte, R. (2020). Jagged Ligands Enhance the Pro-Angiogenic Activity of Multiple Myeloma Cells. Cancers, 12(9), 2600. https://doi.org/10.3390/cancers12092600