
Liver Immune Microenvironment and Metastasis from Colorectal Cancer-Pathogenesis and Therapeutic Perspectives


Abstract
:Simple Summary
Abstract
1. Introduction on Colorectal Cancer (CRC) Liver Metastasis
1.1. Liver Tropism in Cancer Metastasis
1.2. Clinical Significance of CRC Liver Metastasis
1.3. Pathogenesis of CRC Liver Metastases
2. Liver Immune Microenvironment for CRC Liver Metastasis
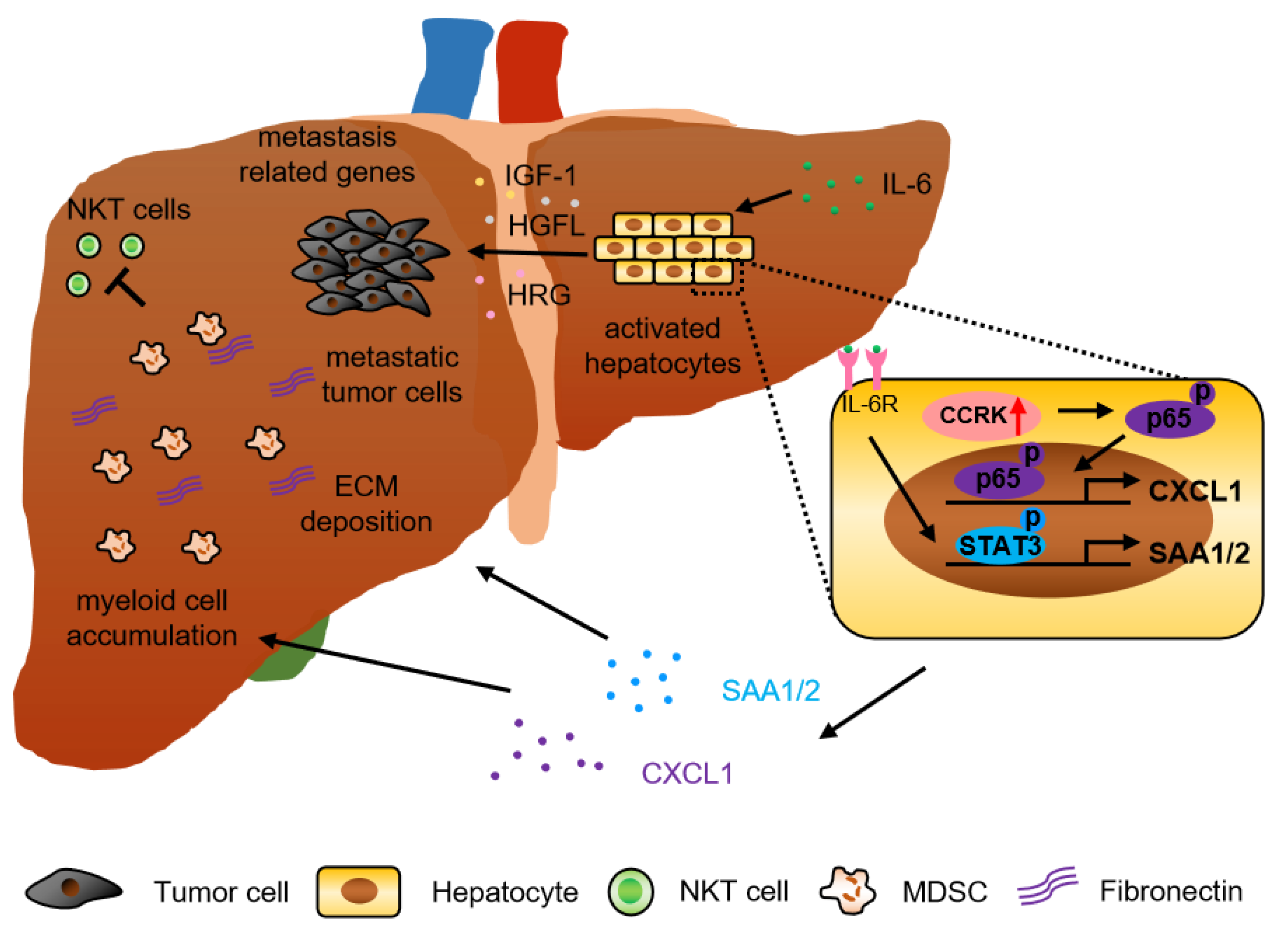
2.1. Hepatocyte-Derived Factors
2.1.1. Inflammatory Cytokines/Chemokines and Growth Factors
2.1.2. Cyclin-Dependent Kinases (CDKs)
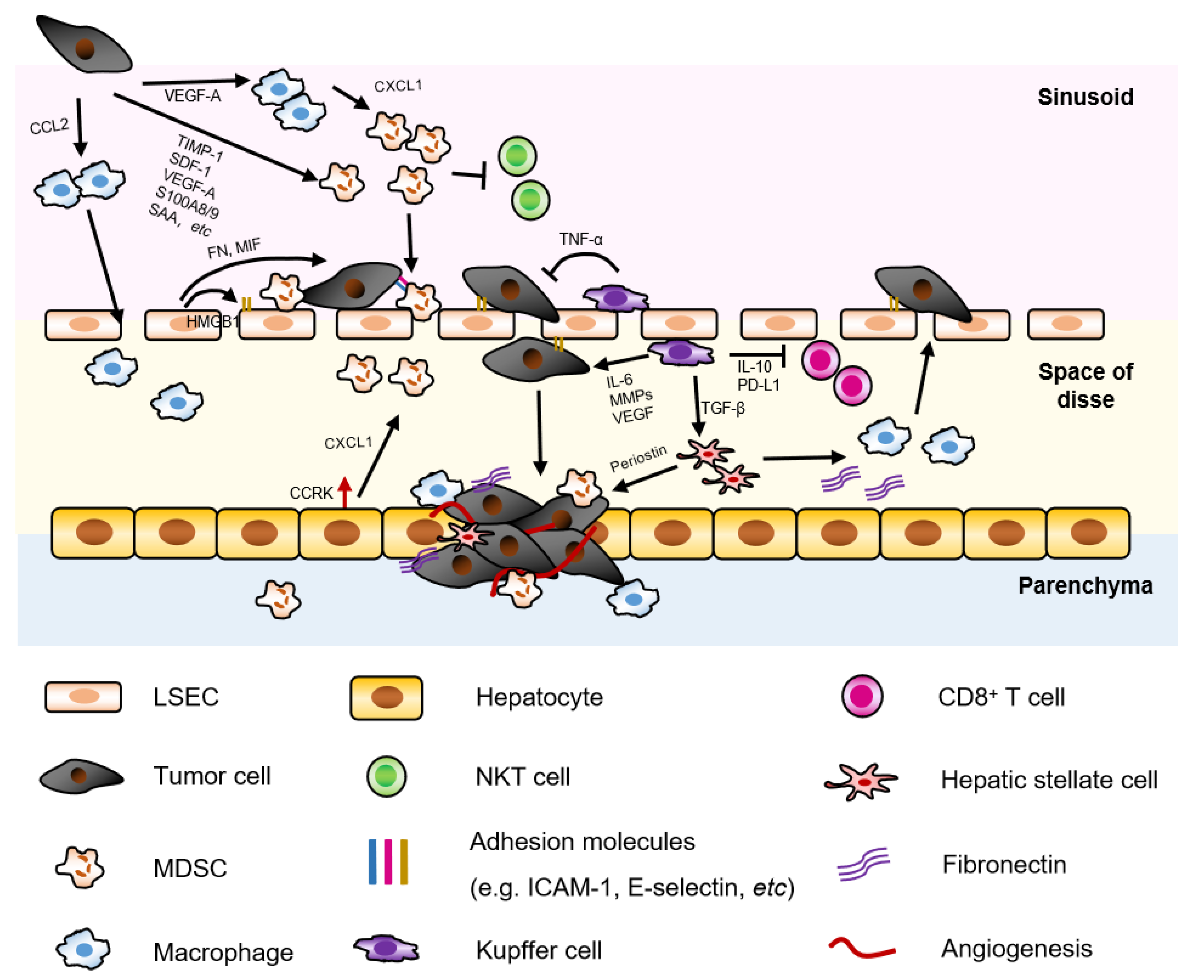
2.2. Liver Non-Parenchymal Cells
2.2.1. Hepatic Stellate Cell (HSC) and Cancer Associated Fibroblasts (CAF)
2.2.2. Liver Sinusoidal Endothelial Cell (LSEC)
2.2.3. Kupffer Cell
2.3. Liver-Infiltrating Immune Cells
2.3.1. Neutrophil and Myeloid-Derived Suppressor Cell (MDSC)
2.3.2. Monocyte, Tumor Associated Macrophage (TAM) or Metastasis-Associated Macrophage (MAM)
2.3.3. NK Cell
2.3.4. NKT Cell
2.3.5. Regulatory T Cells (Tregs) and Other Cells
2.4. Role of Extracellular Matrix (ECM)
2.5. Immune Checkpoint Molecules
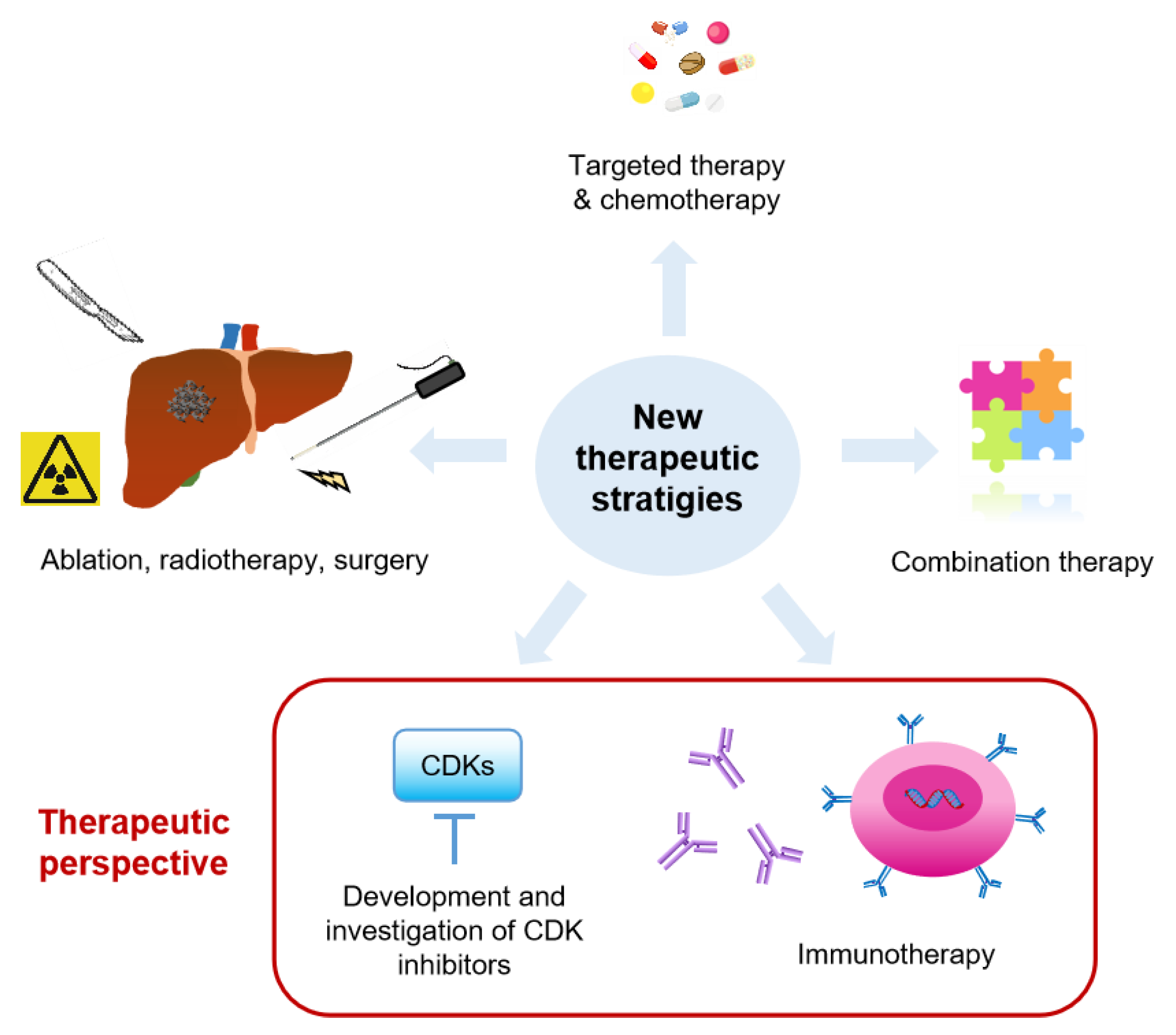
3. Therapeutics for CRC Liver Metastasis
3.1. Current Therapies for CRC Liver Metastasis
3.1.1. Surgical Resection
3.1.2. Systemic Therapy
3.1.3. Localized Therapy
3.2. Therapeutic Perspectives
3.2.1. Targeted Therapy Development, e.g., CDKs
3.2.2. Immunotherapy Development
3.3. Current Clinical Trials
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chaffer, C.L.; Weinberg, R.A. A Perspective on Cancer Cell Metastasis. Science 2011, 331, 1559–1564. [Google Scholar] [CrossRef] [PubMed]
- Budczies, J.; Von Winterfeld, M.; Klauschen, F.; Bockmayr, M.; Lennerz, J.K.; Denkert, C.; Wolf, T.; Warth, A.; Dietel, M.; Anagnostopoulos, I.; et al. The landscape of metastatic progression patterns across major human cancers. Oncotarget 2014, 6, 570–583. [Google Scholar] [CrossRef] [Green Version]
- Mielgo, A.; Schmid, M.C. Liver Tropism in Cancer: The Hepatic Metastatic Niche. Cold Spring Harb. Perspect. Med. 2019, 10, a037259. [Google Scholar] [CrossRef]
- Ciner, A.T.; Jones, K.; Muschel, R.J.; Brodt, P. The unique immune microenvironment of liver metastases: Challenges and opportunities. Semin. Cancer Biol. 2021, 71, 143–156. [Google Scholar] [CrossRef]
- de Ridder, J.; de Wilt, J.H.; Simmer, F.; Overbeek, L.; Lemmens, V.; Nagtegaal, I. Incidence and origin of histologically confirmed liver metastases: An explorative case-study of 23,154 patients. Oncotarget 2016, 7, 55368–55376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Gastroenterol. Rev. 2019, 14, 89–103. [Google Scholar] [CrossRef]
- Millikan, K.W.; Staren, E.D.; Doolas, A. Invasive Therapy of Metastatic colorectal cancer to the liver. Surg. Clin. N. Am. 1997, 77, 27–48. [Google Scholar] [CrossRef]
- Alberts, S.R.; Wagman, L.D. Chemotherapy for Colorectal Cancer Liver Metastases. Oncologist 2008, 13, 1063–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konopke, R.; Kersting, S.; Distler, M.; Dietrich, J.; Gastmeier, J.; Heller, A.; Kulisch, E.; Saeger, H.-D. Prognostic factors and evaluation of a clinical score for predicting survival after resection of colorectal liver metastases. Liver Int. 2009, 29, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Murray, T.; Ward, E.; Samuels, A.; Tiwari, R.C.; Ghafoor, A.; Feuer, E.J.; Thun, M.J. Cancer Statistics, 2005. CA Cancer J. Clin. 2005, 55, 10–30. [Google Scholar] [CrossRef] [PubMed]
- Galjart, B.; Nierop, P.M.H.; Van Der Stok, E.P.; Braak, R.R.J.C.V.D.; Höppener, D.J.; Daelemans, S.; Dirix, L.Y.; Verhoef, C.; Vermeulen, P.B.; Grünhagen, D.J. Angiogenic desmoplastic histopathological growth pattern as a prognostic marker of good outcome in patients with colorectal liver metastases. Angiogenesis 2019, 22, 355–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Vicien, G.; Mezheyeuski, A.; Banuls, M.; Ruiz-Roig, N.; Mollevi, D.G. The Tumor Microenvironment in Liver Metastases from Colorectal Carcinoma in the Context of the Histologic Growth Patterns. Int. J. Mol. Sci. 2021, 22, 1544. [Google Scholar] [CrossRef]
- Höppener, D.J.; Nierop, P.M.H.; Hof, J.; Sideras, K.; Zhou, G.; Visser, L.; Gouw, A.S.H.; De Jong, K.P.; Sprengers, D.; Kwekkeboom, J.; et al. Enrichment of the tumour immune microenvironment in patients with desmoplastic colorectal liver metastasis. Br. J. Cancer 2020, 123, 196–206. [Google Scholar] [CrossRef]
- Brunner, S.M.; Kesselring, R.; Rubner, C.; Martin, M.; Jeiter, T.; Boerner, T.; Ruemmele, P.; Schlitt, H.J.; Fichtner-Feigl, S. Prognosis according to histochemical analysis of liver metastases removed at liver resection. Br. J. Surg. 2014, 101, 1681–1691. [Google Scholar] [CrossRef] [PubMed]
- van Dam, P.-J.; Daelemans, S.; Ross, E.; Waumans, Y.; Van Laere, S.; Latacz, E.; Van Steen, R.; De Pooter, C.; Kockx, M.; Dirix, L.; et al. Histopathological growth patterns as a candidate biomarker for immunomodulatory therapy. Semin. Cancer Biol. 2018, 52, 86–93. [Google Scholar] [CrossRef]
- van Dam, P.-J.; van der Stok, E.P.; Teuwen, L.A.; Van den Eynden, G.G.; Illemann, M.; Frentzas, S.; Majeed, A.W.; Eefsen, R.L.; Coebergh van den Braak, R.R.J.; Lazaris, A.; et al. International consensus guidelines for scoring the histopathological growth patterns of liver metastasis. Br. J. Cancer. 2017, 117, 1427–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, K.; Rolff, H.C.; Eefsen, R.L.; Vainer, B. The morphological growth patterns of colorectal liver metastases are prognostic for overall survival. Mod. Pathol. 2014, 27, 1641–1648. [Google Scholar] [CrossRef] [Green Version]
- Stremitzer, S.; Vermeulen, P.; Graver, S.; Kockx, M.; Dirix, L.; Yang, D.; Zhang, W.; Stift, J.; Wrba, F.; Gruenberger, T.; et al. Immune phenotype and histopathological growth pattern in patients with colorectal liver metastases. Br. J. Cancer. 2020, 122, 1518–1524. [Google Scholar] [CrossRef]
- Rees, M.; Tekkis, P.P.; Welsh, F.K.; O’Rourke, T.; John, T.G. Evaluation of long-term survival after hepatic resection for metastatic colorectal cancer: A multifactorial model of 929 patients. Ann. Surg. 2008, 247, 125–135. [Google Scholar] [CrossRef]
- Nordlinger, B.; Guiguet, M.; Vaillant, J.C.; Balladur, P.; Boudjema, K.; Bachellier, P.; Jaeck, D. Surgical resection of colorectal carcinoma metastases to the liver. A prognostic scoring system to improve case selection, based on 1568 patients. Association Française de Chirurgie. Cancer 1996, 77, 1254–1262. [Google Scholar] [CrossRef]
- Hurwitz, H.; Fehrenbacher, L.; Novotny, W.; Cartwright, T.; Hainsworth, J.; Heim, W.; Berlin, J.; Baron, A.; Griffing, S.; Holmgren, E.; et al. Bevacizumab plus Irinotecan, Fluorouracil, and Leucovorin for Metastatic Colorectal Cancer. N. Engl. J. Med. 2004, 350, 2335–2342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiello, E.; Giuliani, F.; Gebbia, V.; Piano, A.; Agueli, R.; Colucci, G. Cetuximab: Clinical results in colorectal cancer. Ann. Oncol. 2007, 18, vi8–vi10. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, S.Y.; Baek, J.Y.; Cha, Y.J.; Ahn, J.B.; Kim, H.S.; Lee, K.W.; Kim, J.W.; Kim, T.Y.; Chang, W.J.; et al. A Phase II Study of Avelumab Monotherapy in Patients with Mismatch Repair-Deficient/Microsatellite Instability-High or POLE-Mutated Metastatic or Unresectable Colorectal Cancer. Cancer Res. Treat. 2020, 52, 1135–1144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, Z.; Yang, Z.; Wang, M.; Li, S.; Li, Y.; Zhang, R.; Xiong, Z.; Wei, Z.; Shen, J.; et al. Phase I Escalating-Dose Trial of CAR-T Therapy Targeting CEA + Metastatic Colorectal Cancers. Mol. Ther. 2017, 25, 1248–1258. [Google Scholar] [CrossRef]
- Lambert, A.W.; Pattabiraman, D.R.; Weinberg, R.A. Emerging Biological Principles of Metastasis. Cell 2017, 168, 670–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armaghany, T.; Wilson, J.D.; Chu, Q.; Mills, G. Genetic Alterations in Colorectal Cancer. Gastrointest. Cancer Res. 2012, 5, 19–27. [Google Scholar]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F.R. Cancer-related inflammation. Nat. Cell Biol. 2008, 454, 436–444. [Google Scholar] [CrossRef]
- Kitamura, T.; Qian, B.-Z.; Pollard, J.W. Immune cell promotion of metastasis. Nat. Rev. Immunol. 2015, 15, 73–86. [Google Scholar] [CrossRef]
- Joyce, J.A.; Pollard, J.W. Microenvironmental regulation of metastasis. Nat. Rev. Cancer 2008, 9, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Vanaclocha, F.; Crende, O.; de Durango, C.G.; Herreros-Pomares, A.; López-Doménech, S.; González, Á.; Ruiz-Casares, E.; Vilboux, T.; Caruso, R.; Durán, H.; et al. Liver prometastatic reaction: Stimulating factors and responsive cancer phenotypes. Semin. Cancer Biol. 2021, 71, 122–133. [Google Scholar] [CrossRef]
- Costa-Silva, B.; Aiello, N.M.; Ocean, A.J.; Singh, S.; Zhang, H.; Thakur, B.K.; Becker, A.; Hoshino, A.; Mark, M.T.; Molina, H.; et al. Pancreatic cancer exosomes initiate pre-metastatic niche formation in the liver. Nat. Cell Biol. 2015, 17, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Psaila, B.; Lyden, D. The metastatic niche: Adapting the foreign soil. Nat. Rev. Cancer 2009, 9, 285–293. [Google Scholar] [CrossRef]
- Thomas, H. IL-6 drives niche formation in pancreatic cancer liver metastasis. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 263. [Google Scholar] [CrossRef]
- Mohr, A.M.; Gould, J.J.; Kubik, J.L.; Talmon, G.A.; Casey, C.A.; Thomas, P.; Tuma, D.J.; McVicker, B.L. Enhanced colorectal cancer metastases in the alcohol-injured liver. Clin. Exp. Metastasis 2017, 34, 171–184. [Google Scholar] [CrossRef]
- Wang, N.; Rayes, R.F.; Elahi, S.M.; Lu, Y.; Hancock, M.A.; Massie, B.; Rowe, G.E.; Aomari, H.; Hossain, S.; Durocher, Y.; et al. The IGF-Trap: Novel Inhibitor of Carcinoma Growth and Metastasis. Mol. Cancer Ther. 2015, 14, 982–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagh, P.K.; Peace, B.E.; Waltz, S.E. Met-Related Receptor Tyrosine Kinase Ron in Tumor Growth and Metastasis. Adv. Cancer Res. 2008, 100, 1–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshioka, T.; Nishikawa, Y.; Ito, R.; Kawamata, M.; Doi, Y.; Yamamoto, Y.; Yoshida, M.; Omori, Y.; Kotanagi, H.; Masuko, T.; et al. Significance of integrin alphavbeta5 and erbB3 in enhanced cell migration and liver metastasis of colon carcinomas stimulated by hepatocyte-derived heregulin. Cancer Sci. 2010, 101, 2011–2018. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Vanaclocha, F. The Liver Prometastatic Reaction of Cancer Patients: Implications for Microenvironment-Dependent Colon Cancer Gene Regulation. Cancer Microenviron. 2011, 4, 163–180. [Google Scholar] [CrossRef] [Green Version]
- Malumbres, M. Cyclin-dependent kinases. Genome Biol. 2014, 15, 122. [Google Scholar] [CrossRef] [Green Version]
- Raycheva, J.; Karanikolova, T.; Krusteva, R.; Taushanova-Hadjieva, M. Efficacy of CDK4/6 Inhibitor in Treatment of Metastatic Breast Cancer and Colon Cancer. Int. J. Clin. Oncol. Cancer Res. 2020, 5, 1–4. [Google Scholar] [CrossRef]
- Liang, J.; Chen, M.; Hughes, D.; Chumanevich, A.A.; Altilia, S.; Kaza, V.; Lim, C.U.; Kiaris, H.; Mythreye, K.; Pena, M.M.; et al. CDK8 Selectively Promotes the Growth of Colon Cancer Metastases in the Liver by Regulating Gene Expression of TIMP3 and Matrix Metalloproteinases. Cancer Res. 2018, 78, 6594–6606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, S.; Baber, J.; Fujii, T.; Coito, A.J. Matrix metalloproteinases in liver injury, repair and fibrosis. Matrix Biol. 2015, 44–46, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Zhou, J.; Xiong, Z.; Sun, H.; Yang, W.; Mok, M.T.S.; Wang, J.; Li, J.; Liu, M.; Tang, W.; et al. Cell cycle-related kinase reprograms the liver immune microenvironment to promote cancer metastasis. Cell. Mol. Immunol. 2020, 18, 1005–1015. [Google Scholar] [CrossRef]
- Tsuchida, T.; Friedman, S.L. Mechanisms of hepatic stellate cell activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhou, J.; Liu, X.; Feng, Y.; Yang, W.; Wu, F.; Cheung, O.K.-W.; Sun, H.; Zeng, X.; Tang, W.; et al. Targeting monocyte-intrinsic enhancer reprogramming improves immunotherapy efficacy in hepatocellular carcinoma. Gut 2020, 69, 365–379. [Google Scholar] [CrossRef]
- Bao, S.; Ouyang, G.; Bai, X.; Huang, Z.; Ma, C.; Liu, M.; Shao, R.; Anderson, R.M.; Rich, J.N.; Wang, X.-F. Periostin potently promotes metastatic growth of colon cancer by augmenting cell survival via the Akt/PKB pathway. Cancer Cell 2004, 5, 329–339. [Google Scholar] [CrossRef]
- Eveno, C.; Hainaud, P.; Rampanou, A.; Bonnin, P.; Bakhouche, S.; Dupuy, E.; Contreres, J.O.; Pocard, M. Proof of prometastatic niche induction by hepatic stellate cells. J. Surg. Res. 2015, 194, 496–504. [Google Scholar] [CrossRef]
- Takatsuna, M.; Morohashi, S.; Yoshizawa, T.; Hirai, H.; Haga, T.; Ota, R.; Wu, Y.; Morohashi, H.; Hakamada, K.; Terai, S.; et al. Myofibroblasts of the muscle layer stimulate the malignant potential of colorectal cancer. Oncol. Rep. 2016, 36, 1251–1257. [Google Scholar] [CrossRef] [Green Version]
- Solano-Iturri, J.D.; Beitia, M.; Errarte, P.; Calvete-Candenas, J.; Etxezarraga, M.C.; Loizate, A.; Echevarria, E.; Badiola, I.; Larrinaga, G. Altered expression of fibroblast activation protein-alpha (FAP) in colorectal adenoma-carcinoma sequence and in lymph node and liver metastases. Aging 2020, 12, 10337–10358. [Google Scholar] [CrossRef]
- Zhang, R.; Qi, F.; Shao, S.; Li, G.; Feng, Y. Human colorectal cancer-derived carcinoma associated fibroblasts promote CD44-mediated adhesion of colorectal cancer cells to endothelial cells by secretion of HGF. Cancer Cell Int. 2019, 19, 192. [Google Scholar] [CrossRef]
- Gonzalez-Zubeldia, I.; Dotor, J.; Redrado, M.; Bleau, A.M.; Manrique, I.; de Aberasturi, A.L.; Villalba, M.; Calvo, A. Co-migration of colon cancer cells and CAFs induced by TGFbeta (1) enhances liver metastasis. Cell Tissue Res. 2015, 359, 829–839. [Google Scholar] [CrossRef]
- Hu, J.L.; Wang, W.; Lan, X.L.; Zeng, Z.C.; Liang, Y.S.; Yan, Y.R.; Song, F.Y.; Wang, F.F.; Zhu, X.H.; Liao, W.J.; et al. CAFs secreted exosomes promote metastasis and chemotherapy resistance by enhancing cell stemness and epithelial-mesenchymal transition in colorectal cancer. Mol. Cancer 2019, 18, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, H.X.; Gong, W.Z.; Zhou, K.; Xiao, Z.G.; Hou, F.T.; Huang, T.; Zhang, L.; Dong, H.Y.; Zhang, W.L.; Liu, Y.; et al. CXCR4/TGF-beta1 mediated hepatic stellate cells differentiation into carcinoma-associated fibroblasts and promoted liver metastasis of colon cancer. Cancer Biol. Ther. 2020, 21, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Wahid, A.; Cydzik, M.; Fischer, N.W.; Prodeus, A.; Shively, J.E.; Martel, A.; Alminawi, S.; Ghorab, Z.; Berinstein, N.L.; Gariepy, J. Serum-derived carcinoembryonic antigen (CEA) activates fibroblasts to induce a local re-modeling of the extracellular matrix that favors the engraftment of CEA-expressing tumor cells. Int. J. Cancer 2018, 143, 1963–1977. [Google Scholar] [CrossRef]
- Ouahoud, S.; Voorneveld, P.W.; van der Burg, L.R.A.; de Jonge-Muller, E.S.M.; Schoonderwoerd, M.J.A.; Paauwe, M.; de Vos, T.; de Wit, S.; van Pelt, G.W.; Mesker, W.E.; et al. Bidirectional tumor/stroma crosstalk promotes metastasis in mesenchymal colorectal cancer. Oncogene 2020, 39, 2453–2466. [Google Scholar] [CrossRef]
- Kobayashi, H.; Gieniec, K.A.; Wright, J.A.; Wang, T.; Asai, N.; Mizutani, Y.; Lida, T.; Ando, R.; Suzuki, N.; Lannagan, T.R.; et al. The Balance of Stromal BMP Signaling Mediated by GREM1 and ISLR Drives Colorectal Carcinogenesis. Gastroenterology 2021, 160, 1224–1239.e30. [Google Scholar] [CrossRef] [PubMed]
- Shetty, S.; Lalor, P.F.; Adams, D.H. Liver sinusoidal endothelial cells—gatekeepers of hepatic immunity. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 555–567. [Google Scholar] [CrossRef]
- Heymann, F.; Tacke, F. Immunology in the liver—from homeostasis to disease. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 88–110. [Google Scholar] [CrossRef]
- Benedicto, A.; Marquez, J.; Herrero, A.; Olaso, E.; Kolaczkowska, E.; Arteta, B. Decreased expression of the beta2 integrin on tumor cells is associated with a reduction in liver metastasis of colorectal cancer in mice. BMC Cancer 2017, 17, 827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatib, A.-M.; Fallavollita, L.; Wancewicz, E.V.; Monia, B.P.; Brodt, P. Inhibition of hepatic endothelial E-selectin expression by C-raf antisense oligonucleotides blocks colorectal carcinoma liver metastasis. Cancer Res. 2002, 62, 5393–5398. [Google Scholar]
- Khatib, A.-M.; Auguste, P.; Fallavollita, L.; Wang, N.; Samani, A.; Kontogiannea, M.; Meterissian, S.; Brodt, P. Characterization of the Host Proinflammatory Response to Tumor Cells during the Initial Stages of Liver Metastasis. Am. J. Pathol. 2005, 167, 749–759. [Google Scholar] [CrossRef] [Green Version]
- Ham, B.; Wang, N.; D’Costa, Z.; Fernandez, M.C.; Bourdeau, F.; Auguste, P.; Illemann, M.; Eefsen, R.L.; Høyer-Hansen, G.; Vainer, B.; et al. TNF Receptor-2 Facilitates an Immunosuppressive Microenvironment in the Liver to Promote the Colonization and Growth of Hepatic Metastases. Cancer Res. 2015, 75, 5235–5247. [Google Scholar] [CrossRef] [Green Version]
- Wohlfeil, S.A.; Häfele, V.; Dietsch, B.; Schledzewski, K.; Winkler, M.; Zierow, J.; Leibing, T.; Mohammadi, M.M.; Heineke, J.; Sticht, C.; et al. Hepatic Endothelial Notch Activation Protects against Liver Metastasis by Regulating Endothelial-Tumor Cell Adhesion Independent of Angiocrine Signaling. Cancer Res. 2019, 79, 598–610. [Google Scholar] [CrossRef] [Green Version]
- Witz, I.P. The selectin–selectin ligand axis in tumor progression. Cancer Metastasis Rev. 2008, 27, 19–30. [Google Scholar] [CrossRef]
- Elliott, V.A.; Rychahou, P.; Zaytseva, Y.Y.; Evers, B.M. Activation of c-Met and upregulation of CD44 expression are associated with the metastatic phenotype in the colorectal cancer liver metastasis model. PLoS ONE 2014, 9, e97432. [Google Scholar] [CrossRef] [PubMed]
- Aychek, T.; Miller, K.; Sagi-Assif, O.; Levy-Nissenbaum, O.; Israeli-Amit, M.; Pasmanik-Chor, M.; Jacob-Hirsch, J.; Amariglio, N.; Rechavi, G.; Witz, I.P. E-selectin regulates gene expression in metastatic colorectal carcinoma cells and enhances HMGB1 release. Int. J. Cancer 2008, 123, 1741–1750. [Google Scholar] [CrossRef]
- Ou, J.; Peng, Y.; Deng, J.; Miao, H.; Zhou, J.; Zha, L.; Zhou, R.; Yu, L.; Shi, H.; Liang, H. Endothelial cell-derived fibronectin extra domain A promotes colorectal cancer metastasis via inducing epithelial-mesenchymal transition. Carcinogenesis 2014, 35, 1661–1670. [Google Scholar] [CrossRef] [Green Version]
- Gardner, C.R.; Wasserman, A.J.; Laskin, D.L. Liver macrophage-mediated cytotoxicity toward mastocytoma cells involves phagocytosis of tumor targets. Hepatology 1991, 14, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Seki, S.; Nakashima, H.; Nakashima, M.; Kinoshita, M. Antitumor Immunity Produced by the Liver Kupffer Cells, NK Cells, NKT Cells, and CD8+CD122+T Cells. Clin. Dev. Immunol. 2011, 2011, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.H.; McIntosh, A.R.; Hasinoff, B.B.; Rector, E.S.; Ahmed, N.; Nance, D.M.; Orr, F.W. B16 melanoma cell arrest in the mouse liver induces nitric oxide release and sinusoidal cytotoxicity: A natural hepatic defense against metastasis. Cancer Res. 2000, 60, 5862–5869. [Google Scholar] [PubMed]
- Bayon, L.G.; Izquierdo, M.A.; Sirovich, I.; van Rooijen, N.; Beelen, R.H.; Meijer, S. Role of Kupffer cells in arresting circulating tumor cells and controlling metastatic growth in the liver. Hepatology 1996, 23, 1224–1231. [Google Scholar] [CrossRef]
- Heymann, F.; Peusquens, J.; Ludwig-Portugall, I.; Kohlhepp, M.; Ergen, C.; Niemietz, P.; Martin, C.; Van Rooijen, N.; Ochando, J.C.; Randolph, G.J.; et al. Liver inflammation abrogates immunological tolerance induced by Kupffer cells. Hepatology 2015, 62, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Fridlender, Z.G.; Sun, J.; Kim, S.; Kapoor, V.; Cheng, G.; Ling, L.; Worthen, G.S.; Albelda, S.M. Polarization of tumor-associated neutrophil phenotype by TGF-beta: “N1” versus “N2” TAN. Cancer Cell 2009, 16, 183–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaul, M.E.; Fridlender, Z.G. Neutrophils as active regulators of the immune system in the tumor microenvironment. J. Leukoc. Biol. 2017, 102, 343–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seubert, B.; Grünwald, B.; Kobuch, J.; Cui, H.; Schelter, F.; Schaten, S.; Siveke, J.T.; Lim, N.H.; Nagase, H.; Simonavicius, N.; et al. Tissue inhibitor of metalloproteinases (TIMP)-1 creates a premetastatic niche in the liver through SDF-1/CXCR4-dependent neutrophil recruitment in mice. Hepatology 2015, 61, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, V.; Lazaris, A.; Mayer, T.Z.; Petrillo, S.K.; Alamri, H.; Rada, M.; Jarrouj, G.; Park, W.; Gao, Z.; McDonald, P.P.; et al. Neutrophils expressing lysyl oxidase-like 4 protein are present in colorectal cancer liver metastases resistant to anti-angiogenic therapy. J. Pathol. 2020, 251, 213–223. [Google Scholar] [CrossRef]
- Wang, D.; Sun, H.; Wei, J.; Cen, B.; Dubois, R.N. CXCL1 Is Critical for Premetastatic Niche Formation and Metastasis in Colorectal Cancer. Cancer Res. 2017, 77, 3655–3665. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Davis, C.; Ryan, J.; Janney, C.; Pena, M.M. Development and characterization of a reliable mouse model of colorectal cancer metastasis to the liver. Clin. Exp. Metastasis 2013, 30, 903–918. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Montero, C.M.; Salem, M.L.; Nishimura, M.I.; Garrett-Mayer, E.; Cole, D.J.; Montero, A.J. Increased circulating myeloid-derived suppressor cells correlate with clinical cancer stage, metastatic tumor burden, and doxorubicin-cyclophosphamide chemotherapy. Cancer Immunol. Immunother 2009, 58, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Spicer, J.D.; McDonald, B.; Cools-Lartigue, J.J.; Chow, S.C.; Giannias, B.; Kubes, P.; Ferri, L.E. Neutrophils promote liver metastasis via Mac-1-mediated interactions with circulating tumor cells. Cancer Res. 2012, 72, 3919–3927. [Google Scholar] [CrossRef] [Green Version]
- Qian, B.Z.; Li, J.; Zhang, H.; Kitamura, T.; Zhang, J.; Campion, L.R.; Kaiser, E.A.; Snyder, L.A.; Pollard, J.W. CCL2 recruits inflammatory monocytes to facilitate breast-tumour metastasis. Nature 2011, 475, 222–225. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, S.R.; Quaranta, V.; Linford, A.; Emeagi, P.; Rainer, C.; Santos, A.; Ireland, L.; Sakai, T.; Sakai, K.; Kim, Y.S.; et al. Macrophage-secreted granulin supports pancreatic cancer metastasis by inducing liver fibrosis. Nat. Cell. Biol. 2016, 18, 549–560. [Google Scholar] [CrossRef] [Green Version]
- Quaranta, V.; Rainer, C.; Nielsen, S.R.; Raymant, M.L.; Ahmed, M.S.; Engle, D.D.; Taylor, A.; Murray, T.; Campbell, F.; Palmer, D.H.; et al. Macrophage-Derived Granulin Drives Resistance to Immune Checkpoint Inhibition in Metastatic Pancreatic Cancer. Cancer Res. 2018, 78, 4253–4269. [Google Scholar] [CrossRef] [Green Version]
- Mitchem, J.B.; Brennan, D.J.; Knolhoff, B.L.; Belt, B.A.; Zhu, Y.; Sanford, D.E.; Belaygorod, L.; Carpenter, D.; Collins, L.; Piwnica-Worms, D.; et al. Targeting tumor-infiltrating macrophages decreases tumor-initiating cells, relieves immunosuppression, and improves chemotherapeutic responses. Cancer Res. 2013, 73, 1128–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Lim, S.Y.; Gordon-Weeks, A.N.; Tapmeier, T.T.; Im, J.H.; Cao, Y.; Beech, J.; Allen, D.; Smart, S.; Muschel, R.J. Recruitment of a myeloid cell subset (CD11b/Gr1 mid) via CCL2/CCR2 promotes the development of colorectal cancer liver metastasis. Hepatology 2013, 57, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Lai, X.; Zhao, Y.; Zhang, Y.; Li, M.; Li, D.; Kong, J.; Zhang, Y.; Jing, P.; Li, H.; et al. Loss of NDRG2 in liver microenvironment inhibits cancer liver metastasis by regulating tumor associate macrophages polarization. Cell Death Dis. 2018, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.W.; Harmon, C.; O’Farrelly, C. Liver immunology and its role in inflammation and homeostasis. Cell. Mol. Immunol. 2016, 13, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Lau, J.K.C.; Yu, J. The role of natural killer cell in gastrointestinal cancer: Killer or helper. Oncogene 2021, 40, 717–730. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef] [PubMed]
- López-Soto, A.; Gonzalez, S.; Smyth, M.J.; Galluzzi, L. Control of Metastasis by NK Cells. Cancer Cell 2017, 32, 135–154. [Google Scholar] [CrossRef]
- Ducimetière, L.; Lucchiari, G.; Litscher, G.; Nater, M.; Heeb, L.; Nuñez, N.G.; Wyss, L.; Burri, D.; Vermeer, M.; Gschwend, J.; et al. Conventional NK cells and tissue-resident ILC1s join forces to control liver metastasis. BioRxiv 2020. [Google Scholar] [CrossRef]
- Harmon, C.; Robinson, M.W.; Hand, F.; AlMuaili, D.; Mentor, K.; Houlihan, D.D.; Hoti, E.; Lynch, L.; Geoghegan, J.; O’Farrelly, C. Lactate-Mediated Acidification of Tumor Microenvironment Induces Apoptosis of Liver-Resident NK Cells in Colorectal Liver Metastasis. Cancer Immunol. Res. 2018, 7, 335–346. [Google Scholar] [CrossRef] [Green Version]
- Dupaul-Chicoine, J.; Arabzadeh, A.; Dagenais, M.; Douglas, T.; Champagne, C.; Morizot, A.; Rodrigue-Gervais, I.G.; Breton, V.; Colpitts, S.L.; Beauchemin, N.; et al. The Nlrp3 Inflammasome Suppresses Colorectal Cancer Metastatic Growth in the Liver by Promoting Natural Killer Cell Tumoricidal Activity. Immunity 2015, 43, 751–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, K.; Hayakawa, Y.; Smyth, M.J.; Kayagaki, N.; Yamaguchi, N.; Kakuta, S.; Iwakura, Y.; Yagita, H.; Okumura, K. Involvement of tumor necrosis factor-related apoptosis-inducing ligand in surveillance of tumor metastasis by liver natural killer cells. Nat. Med. 2001, 7, 94–100. [Google Scholar] [CrossRef]
- Donadon, M.; Hudspeth, K.; Cimino, M.; Di Tommaso, L.; Preti, M.; Tentorio, P.; Roncalli, M.; Mavilio, D.; Torzilli, G. Increased Infiltration of Natural Killer and T Cells in Colorectal Liver Metastases Improves Patient Overall Survival. J. Gastrointest. Surg. 2017, 21, 1226–1236. [Google Scholar] [CrossRef]
- Krijgsman, D.; Hokland, M.; Kuppen, P.J.K. The Role of Natural Killer T Cells in Cancer-A Phenotypical and Functional Approach. Front. Immunol. 2018, 9, 367. [Google Scholar] [CrossRef]
- Kee, J.-Y.; Ito, A.; Hojo, S.; Hashimoto, I.; Igarashi, Y.; Tsukada, K.; Irimura, T.; Shibahara, N.; Nakayama, T.; Yoshie, O.; et al. Chemokine CXCL16 suppresses liver metastasis of colorectal cancer via augmentation of tumor-infiltrating natural killer T cells in a murine model. Oncol. Rep. 2013, 29, 975–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Han, M.; Heinrich, B.; Fu, Q.; Zhang, Q.; Sandhu, M.; Agdashian, D.; Terabe, M.; Berzofsky, J.A.; Fako, V.; et al. Gut microbiome-mediated bile acid metabolism regulates liver cancer via NKT cells. Science 2018, 360, 876–885. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Li, H.; Mayhew, E.; Mellon, J.; Chen, P.W.; Niederkorn, J.Y. NKT Cell Exacerbation of Liver Metastases Arising from Melanomas Transplanted into Either the Eyes or Spleens of Mice. Investig. Opthalmology Vis. Sci. 2011, 52, 3094–3102. [Google Scholar] [CrossRef] [Green Version]
- Katz, S.C.; Bamboat, Z.M.; Maker, A.V.; Shia, J.; Pillarisetty, V.G.; Yopp, A.C.; Hedvat, C.V.; Gonen, M.; Jarnagin, W.R.; Fong, Y.; et al. Regulatory T Cell Infiltration Predicts Outcome Following Resection of Colorectal Cancer Liver Metastases. Ann. Surg. Oncol. 2013, 20, 946–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, R.N.; Riba, R.D.; Zacharoulis, S.; Bramley, A.H.; Vincent, L.; Costa, C.; Macdonald, D.D.; Jin, D.K.; Shido, K.; Kerns, S.A.; et al. VEGFR1-positive haematopoietic bone marrow progenitors initiate the pre-metastatic niche. Nature 2005, 438, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Hiratsuka, S.; Nakamura, K.; Iwai, S.; Murakami, M.; Itoh, T.; Kijima, H.; Shipley, J.M.; Senior, R.M.; Shibuya, M. MMP9 induction by vascular endothelial growth factor receptor-1 is involved in lung-specific metastasis. Cancer Cell 2002, 2, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Bosman, F.T.; Stamenkovic, I. Functional structure and composition of the extracellular matrix. J. Pathol. 2003, 200, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Voss, H.; Wurlitzer, M.; Smit, D.J.; Ewald, F.; Alawi, M.; Spohn, M.; Indenbirken, D.; Omidi, M.; David, K.; Juhl, H.; et al. Differential regulation of extracellular matrix proteins in three recurrent liver metastases of a single patient with colorectal cancer. Clin. Exp. Metastasis 2020, 37, 649–656. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, X.; Lu, J.; Salfenmoser, M.; Wirsik, N.M.; Schleussner, N.; Imle, A.; Valls, A.F.; Radhakrishnan, P.; Liang, J.; et al. Reduction of Liver Metastasis Stiffness Improves Response to Bevacizumab in Metastatic Colorectal Cancer. Cancer Cell 2020, 37, 800–817.e7. [Google Scholar] [CrossRef]
- Rahbari, N.N.; Kedrin, D.; Incio, J.; Liu, H.; Ho, W.W.; Nia, H.T.; Edrich, C.M.; Jung, K.; Daubriac, J.; Chen, I.; et al. Anti-VEGF therapy induces ECM remodeling and mechanical barriers to therapy in colorectal cancer liver metastases. Sci. Transl. Med. 2016, 8, 360ra135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cools-Lartigue, J.; Spicer, J.; McDonald, B.; Gowing, S.; Chow, S.; Giannias, B.; Bourdeau, F.; Kubes, P.; Ferri, L. Neutrophil extracellular traps sequester circulating tumor cells and promote metastasis. J. Clin. Investig. 2013, 123, 3446–3458. [Google Scholar] [CrossRef]
- Tohme, S.; Yazdani, H.O.; Al-Khafaji, A.B.; Chidi, A.P.; Loughran, P.; Mowen, K.; Wang, Y.; Simmons, R.L.; Huang, H.; Tsung, A. Neutrophil extracellular traps promote the development and progression of liver metastases after surgical stress. Cancer Res. 2016, 76, 1367–1380. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Wysocki, R.W.; Amoozgar, Z.; Maiorino, L.; Fein, M.R.; Jorns, J.; Schott, A.F.; Kinugasa-Katayama, Y.; Lee, Y.; Won, N.H.; et al. Cancer cells induce metastasis-supporting neutrophil extracellular DNA traps. Sci. Transl. Med. 2016, 8, 361ra138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.L.; Luo, X.; Sheng, H.; Wang, Y.; Chen, D.L.; Li, J.N.; Wang, F.H.; Xu, R.H. PD-L1 expression in liver metastasis: Its clinical significance and discordance with primary tumor in colorectal cancer. J. Transl. Med. 2020, 18, 475. [Google Scholar] [CrossRef]
- Jabbari, N.; Kenerson, H.L.; Lausted, C.; Yan, X.; Meng, C.; Sullivan, K.M.; Baloni, P.; Bergey, D.; Pillarisetty, V.G.; Hood, L.E.; et al. Modulation of Immune Checkpoints by Chemotherapy in Human Colorectal Liver Metastases. Cell Rep. Med. 2020, 1, 100160. [Google Scholar] [CrossRef] [PubMed]
- Fiegle, E.; Doleschel, D.; Koletnik, S.; Rix, A.; Weiskirchen, R.; Borkham-Kamphorst, E.; Kiessling, F.; Lederle, W. Dual CTLA-4 and PD-L1 Blockade Inhibits Tumor Growth and Liver Metastasis in a Highly Aggressive Orthotopic Mouse Model of Colon Cancer. Neoplasia 2019, 21, 932–944. [Google Scholar] [CrossRef]
- Steele, G.; Bleday, R.; Mayer, R.J.; Lindblad, A.; Petrelli, N.; Weaver, D. A prospective evaluation of hepatic resection for colorectal carcinoma metastases to the liver: Gastrointestinal Tumor Study Group Protocol 6584. J. Clin. Oncol. 1991, 9, 1105–1112. [Google Scholar] [CrossRef]
- Scheele, J.; Stang, R.; Altendorf-Hofmann, A.; Paul, M. Resection of colorectal liver metastases. World J. Surg. 1995, 19, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Cummings, L.C.; Payes, J.D.; Cooper, G.S. Survival after hepatic resection in metastatic colorectal cancer: A population-based study. Cancer 2007, 109, 718–726. [Google Scholar] [CrossRef] [PubMed]
- Turrini, O.; Viret, F.; Guiramand, J.; Lelong, B.; Bege, T.; Delpero, J.R. Strategies for the treatment of synchronous liver metastasis. Eur. J. Surg. Oncol. 2007, 33, 735–740. [Google Scholar] [CrossRef]
- Tomlinson, J.S.; Jarnagin, W.R.; DeMatteo, R.P.; Fong, Y.; Kornprat, P.; Gonen, M.; Kemeny, N.; Brennan, M.F.; Blumgart, L.H.; D’Angelica, M. Actual 10-Year Survival After Resection of Colorectal Liver Metastases Defines Cure. J. Clin. Oncol. 2007, 25, 4575–4580. [Google Scholar] [CrossRef]
- Misiakos, E.P.; Karidis, N.P.; Kouraklis, G. Current treatment for colorectal liver metastases. World J. Gastroenterol. 2011, 17, 4067–4475. [Google Scholar] [CrossRef]
- Abdalla, E.K.; Vauthey, J.-N.; Ellis, L.M.; Ellis, V.; Pollock, R.; Broglio, K.R.; Hess, K.; Curley, S.A. Recurrence and Outcomes Following Hepatic Resection, Radiofrequency Ablation, and Combined Resection/Ablation for Colorectal Liver Metastases. Ann. Surg. 2004, 239, 818–827. [Google Scholar] [CrossRef]
- Kalofonos, H.; Aravantinos, G.; Kosmidis, P.; Papakostas, P.; Economopoulos, T.; Dimopoulos, M.; Skarlos, D.; Bamias, A.; Pectasides, D.; Chalkidou, S.; et al. Irinotecan or oxaliplatin combined with leucovorin and 5-fluorouracil as first-line treatment in advanced colorectal cancer: A multicenter, randomized, phase II study. Ann. Oncol. 2005, 16, 869–877. [Google Scholar] [CrossRef]
- Seium, Y.; Stupp, R.; Ruhstaller, T.; Gervaz, P.; Mentha, G.; Philippe, M.; Allal, A.; Trembleau, C.; Bauer, J.; Morant, R.; et al. Oxaliplatin combined with irinotecan and 5-fluorouracil/leucovorin (OCFL) in metastatic colorectal cancer: A phase I-II study. Ann Oncol. 2005, 16, 762–766. [Google Scholar] [CrossRef]
- Grivicich, I.; Mans, D.R.; Peters, G.J.; Schwartsmann, G. Irinotecan and oxaliplatin: An overview of the novel chemotherapeutic options for the treatment of advanced colorectal cancer. Braz. J. Med. Biol. Res. 2001, 34, 1087–1103. [Google Scholar] [CrossRef] [Green Version]
- Adam, R.; Avisar, E.; Ariche, A.; Giachetti, S.; Azoulay, D.; Castaing, D.; Kunstlinger, F.; Levi, F.; Bismuth, F. Five-year survival following hepatic resection after neoadjuvant therapy for nonresectable colorectal. Ann. Surg. Oncol. 2001, 8, 347–353. [Google Scholar] [CrossRef]
- Adam, R.; Delvart, V.; Pascal, G.; Valeanu, A.; Castaing, D.; Azoulay, D.; Giacchetti, S.; Paule, B.; Kunstlinger, F.; Ghemard, O.; et al. Rescue surgery for unresectable colorectal liver metastases downstaged by chemotherapy: A model to predict long-term survival. Ann. Surg. 2004, 240, 644–657. [Google Scholar] [CrossRef]
- Raoul, J.-L.; Van Laethem, J.-L.; Peeters, M.; Brezault, C.; Husseini, F.; Cals, L.; Nippgen, J.; Loos, A.-H.; Rougier, P. Cetuximab in combination with irinotecan/5-fluorouracil/folinic acid (FOLFIRI) in the initial treatment of metastatic colorectal cancer: A multicentre two-part phase I/II study. BMC Cancer 2009, 9, 112. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.-X.; Zhang, W.-H.; Jiang, H.-C. Current treatment for liver metastases from colorectal cancer. World J. Gastroenterol. 2003, 9, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Lygidakis, N.J.; Sgourakis, G.; Dedemadi, G.; Safioleus, M.C.; Nestoridis, J. Regional chemoimmunotherapy for nonresectable metastatic liver disease of colorectal origin. A prospective randomized study. Hepatogastroenterology 2001, 48, 1085–1087. [Google Scholar] [PubMed]
- Raval, M.; Bande, D.; Pillai, A.K.; Blaszkowsky, L.S.; Ganguli, S.; Beg, M.S.; Kalva, S.P. Yttrium-90 radioembolization of hepatic metastases from colorectal cancer. Front Oncol. 2014, 4, 120. [Google Scholar] [CrossRef] [Green Version]
- Mok, M.T.; Zhou, J.; Tang, W.; Zeng, X.; Oliver, A.W.; Ward, S.E.; Cheng, A.S. CCRK is a novel signalling hub exploitable in cancer immunotherapy. Pharmacol. Ther. 2018, 186, 138–151. [Google Scholar] [CrossRef] [PubMed]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Yu, Z.; Gao, Y.Q.; Feng, H.; Lee, Y.Y.; Li, M.S.; Tian, Y.; Go, M.Y.; Yu, D.Y.; Cheung, Y.S.; Lai, P.B.; et al. Cell cycle-related kinase mediates viral-host signalling to promote hepatitis B virus-associated hepatocarcinogenesis. Gut 2014, 63, 1793–1804. [Google Scholar] [CrossRef]
- Sun, H.; Yang, W.; Tian, Y.; Zeng, X.; Zhou, J.; Mok, M.T.S.; Tang, W.; Feng, Y.; Xu, L.; Chan, A.W.H.; et al. An inflammatory-CCRK circuitry drives mTORC1-dependent metabolic and immunosuppressive reprogramming in obesity-associated hepatocellular carcinoma. Nat. Commun. 2018, 9, 1–16. [Google Scholar] [CrossRef]
- Feng, H.; Cheng, A.S.; Tsang, D.P.; Li, M.S.; Go, M.Y.; Cheung, Y.S.; Zhao, G.J.; Ng, S.S.; Lin, M.C.; Yu, J.; et al. Cell cycle-related kinase is a direct androgen receptor-regulated gene that drives beta-catenin/T cell factor-dependent hepatocarcinogenesis. J. Clin. Investig. 2011, 121, 3159–3175. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Yu, Z.; Tian, Y.; Lee, Y.-Y.; Li, M.S.; Go, M.Y.; Cheung, Y.-S.; Lai, P.B.; Chan, A.M.; To, K.-F.; et al. A CCRK-EZH2 epigenetic circuitry drives hepatocarcinogenesis and associates with tumor recurrence and poor survival of patients. J. Hepatol. 2015, 62, 1100–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Liu, M.; Sun, H.; Feng, Y.; Xu, L.; Chan, A.W.H.; Tong, J.H.; Wong, J.; Chong, C.C.N.; Lai, P.B.S.; et al. Hepatoma-intrinsic CCRK inhibition diminishes myeloid-derived suppressor cell immunosuppression and enhances immune-checkpoint blockade efficacy. Gut 2018, 67, 931–944. [Google Scholar] [CrossRef] [PubMed]
- Greten, T.F.; Korangy, F. CDK20 inhibition and immune checkpoint blockade: Bringing cancer biology and tumour immunology together to develop novel treatment options for HCC. Gut 2017, 67, 783–784. [Google Scholar] [CrossRef]
- Das, S.; Allen, A.; Berlin, J. Immunotherapy after Immunotherapy: Response Rescue in a Patient With Microsatellite Instability-high Colorectal Cancer Post-Pembrolizumab. Clin. Colorectal. Cancer 2020, 19, 137–140. [Google Scholar] [CrossRef]
- Binnewies, M.; Roberts, E.W.; Kersten, K.; Chan, V.; Fearon, D.F.; Merad, M.; Coussens, L.M.; Gabrilovich, D.I.; Ostrand-Rosenberg, S.; Hedrick, C.C.; et al. Understanding the tumor immune microenvironment (TIME) for effective therapy. Nat. Med. 2018, 24, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhu, L.; Liu, J.; Xie, M.; Chen, J.; Li, J. Emerging Role of Immunotherapy for Colorectal Cancer with Liver Metastasis. OncoTargets Ther. 2020, 13, 11645–11658. [Google Scholar] [CrossRef] [PubMed]
- Sur, D.; Havasi, A.; Cainap, C.; Samasca, G.; Burz, C.; Balacescu, O.; Lupan, I.; Deleanu, D.; Irimie, A. Chimeric Antigen Receptor T-Cell Therapy for Colorectal Cancer. J. Clin. Med. 2020, 9, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina-Echeverz, J.; Eggert, T.; Han, M.; Greten, T.F. Hepatic myeloid-derived suppressor cells in cancer. Cancer Immunol. Immunother. 2015, 64, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Adam, R.; Shimada, H.; Azoulay, D.; Lévi, F.; Bismuth, H. Role of neoadjuvant chemotherapy in the treatment of multiple colorectal metastases to the liver. Brit. J. Surg. 2003, 90, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Power, D.G.; Kemeny, N.E. Chemotherapy for the conversion of unresectable colorectal cancer liver metastases to resection. Crit. Rev. Oncol. Hematol. 2011, 79, 251–264. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Trial ID | Phase | Study Population | Intervention | Recruitment Status |
|---|---|---|---|---|
| Targeted therapy and chemotherapy (Locally or systemically) | ||||
| NCT04509635 | III | Colorectal Cancer Liver Metastasis with Progression After First-line Treatment of Cetuximab | Cetuximab±Chemotherapy | Not yet recruiting |
| NCT03493061 | II | Unresectable Colorectal Cancer Liver Metastases | Irinotecan; Oxaliplatin; Floxuridine | Recruiting |
| NCT04189055 | II | Neo Wild-type RAS/RAF Metastatic Colorectal Cancer with Liver Metastases | Cetuximab; Irinotecan | Recruiting |
| NCT04276090 | NA | Unresectable Colorectal Metastases/Intrahepatic Cholangiocarcinoma | Codman Catheter; Synchromed Pump Hepatic Artery Chemotherapy | Recruiting |
| NCT03697044 | NA | Colorectal Cancer Liver Metastases | Irinotecan Drug-Eluting-Bead Trans Arterial ChemoEmbolisation | Not yet recruiting |
| NCT03125161/ NCT02102789 | III | Unresectable Colorectal Cancer Liver Metastases | HAI; Chemotherapy ± target therapy or mFOLFOX6 | Recruiting |
| NCT04003792 | II | Unresectable Colorectal Cancer Liver Metastases | oxaliplatin; FOLFIRI Protocol; Bevacizumab | Recruiting |
| NCT00695201 | I | Unresectable Colorectal Cancer Liver Metastases | Floxuridine, Oxaliplatin, CPT-11 | Active, not recruiting |
| NCT04194034 | I/II | Unresectable Colorectal Cancer Liver Metastases | TG6002; Flucytosine (5-FC) | Recruiting |
| NCT04595266 | II | Colorectal Cancer Liver Metastases | FOLFOX regimen; Anti-EGFR or Bevacizumab; LIVERPEARLS-Irinotecan | Not yet recruiting |
| NCT03164655 | II | Unresectable Colorectal Cancer Liver Metastases | Oxaliplatin, Cetuximab, Bevacizumab, Panitumumab, Irinotecan, Leucovorin, 5-Fluorouracil | Recruiting |
| NCT03031444 | II/III | Resectable Colorectal Liver Metastasis | Cetuximab plus FOLFIRI/FOLFOX; FOLFIRI/FOLFOX/CapeOX | Recruiting |
| NCT04525326 | III | Unresectable Colorectal Cancer Liver Metastases | Cetuximab; Bevacizumab; mFOLFOX/FOLFIRI (Standard Chemotherapy) | Not yet recruiting |
| NCT03366155 | II | Colorectal Cancer Liver Metastases | Panitumumab; FUDR-Dex; Oxaliplatin; 5FU; Irinotecan; cetuximab | |
| NCT01312857 | II | Resected Colorectal Cancer Liver Metastasis with Wild Type RAS | panitumumab | Active, not recruiting |
| NCT03732235 | NA | Refractory Colorectal Cancer Liver Metastases | TACE+ systemic Bevacizumab; FOLFIRI+Bevacizumab; TACE | Recruiting |
| NCT04126655 | I/II | Colorectal Cancer Liver Metastases | Arfolitixorin + 5-FU; Calciumfolinate + 5-FU | Recruiting |
| NCT03477019 | I/II | Liver Metastasis from Breast- and Colorectal Cancer | SonoVue; Focused Ultrasound | Recruiting |
| NCT03458975 | II | Colorectal Cancer Liver Metastases | Contrast enhanced ultrasound; Sonoporation | Recruiting |
| NCT04021277 | I | Colorectal Cancer Liver Metastases | PS101-mediated Acoustic Cluster Therapy | Recruiting |
| NCT03493048 | II | RAS wildtype Unresectable Colorectal Cancer Liver Metastases | Irinotecan; Cetuximab; 5-fluorouracil; Oxaliplatin; Leucovorin | Recruiting |
| NCT03801915 | II | Colorectal Cancer Liver Metastases | MVT-5873 | Recruiting |
| NCT02172651 | Early Phase 1 | Stage I-III Colon Cancer or Resectable Colon Cancer Liver Metastases | Vitamin D3 | Recruiting |
| Ablation, radiotherapy, surgery | ||||
| NCT03088150 | NAII | Colorectal Cancer Liver Metastases | Thermal ablation; Surgical resection | Recruiting |
| NCT03963726/ NCT03654131/ NCT04081168 | NA/II | Colorectal Cancer Liver Metastases | stereotactic radiotherapy; microwave ablation | Recruiting |
| NCT02185443 | II | Unresectable Colorectal Cancer Liver Metastases | SBRT | Recruiting |
| NCT04491929 | NA | Refractory Colorectal Cancer Liver Metastases | Selective Internal Radiation Therapy With 90Y Resin | Recruiting |
| NCT03895723 | NA | Colorectal Cancer Liver Metastases | laparoscopic and robotic liver resection or open surgery | Recruiting |
| NCT02954913 | NA | Colorectal Cancer Liver Metastases | Simultaneous Resection | Recruiting |
| NCT03803436 | II | Unresectable Colorectal Cancer Liver Metastases | liver transplantation vs triplet chemotherapy+antiEGFR | Recruiting |
| NCT02864485/ NCT01479608/ NCT02597348 | NA; II; III | Unresectable Colorectal Cancer Liver Metastases | liver transplantation | Recruiting |
| NCT02215889 | I/II | Colorectal Cancer Liver Metastases | Partial Liver Segment 2/3 Transplantation | Recruiting |
| NCT03494946 | NA | Colorectal Cancer Liver Metastases | Liver transplantation vs Chemotherapy | Recruiting |
| NCT04161092 | NA | non-resectable/ non-abatable colorectal liver metastases | Liver transplantation Ltx or best alternative care | Not yet recruiting |
| NCT03488953 | NA | Isolated, Irresectable Colorectal Liver Metastases | Living donor liver transplantation with two-staged hepatectomy | Recruiting |
| NCT03577665 | NA | Colorectal Cancer Liver Metastases | Curative Proton Beam Therapy | Recruiting |
| NCT04108481 | I/II | Colorectal Cancer Liver Metastases | Durvalumab; Yttrium-90 RadioEmbolization | Recruiting |
| Immunotherapy | ||||
| NCT02754856 | I | Resectable Colorectal Cancer Liver Metastases | Durvalumab; Tremelimumab | Recruiting |
| NCT03370198 | I | Unresectable Liver Metastases from Colorectal Cancer | Hepatic Transarterial Administrations of NKR-2(modified T cells) | Active, not recruiting |
| NCT02850536 | I | Liver Metastases or Pancreas Cancer | anti-CEA CAR-T cells | Active, not recruiting |
| NCT04513431 | Early Phase 1 | Stage III Colorectal Cancer Colorectal Cancer Liver Metastasis | Anti-CEA-CAR T | Not yet recruiting |
| Combination therapy | ||||
| NCT04062721 | Ib/II | Unresectable Colorectal Liver Metastases | radiofrequency ablation (RFA) plus in situ TLR agonist and GM-CSF | Not yet recruiting |
| NCT04202978 | I/II | Colorectal Cancer Liver Metastases | Camrelizumab Combined With Apatinib XELOX RFA | Recruiting |
| NCT03223779 | I/II | Colorectal Cancer Liver Metastases | TAS-102; Photon SBRT | Recruiting |
| NCT02738606 | II | Resectable Colorectal Cancer Liver Metastases and unresectable Colorectal Cancer Lung Metastases | liver surgery and chemotherapy | Recruiting |
| NCT03127072 | IV | Unresectable Colorectal Cancer Liver Metastases | Radiofrequency Ablation (RFA); chemotherapy ± target therapy | Recruiting |
| NCT04562727 | NA/II | Colorectal Cancer Liver Metastases | Microwave Ablation Combined with Chemotherapy | Not yet recruiting |
| NCT03135652 | II | Colorectal Cancer Liver Metastases Receiving Surgery or Radiofrequency Ablation | Adjuvant SBRT; Chemotherapy | Recruiting |
| NCT03101475 | II | Colorectal Cancer Liver Metastases | Durvalumab (MEDI4736); Tremelimumab; Sterotactic body radiation therapy (SBRT); Radiofrequency ablation (RFA) | Recruiting |
| NCT04508140 | II | Colorectal or Gastric/GEJ Cancer with Liver Metastasis | BO-112 with Pembrolizumab | Recruiting |
| NCT03507699 | I | Colorectal Cancer Liver Metastases | Liver radiation therapy; Nivolumab Injection; Ipilimumab Injection; CMP-001 | Recruiting |
| NCT04166383 | II | Colorectal Cancer Liver Metastases | VB-111; Nivolumab | Recruiting |
| NCT03785210 | II | Refractory Primary Hepatocellular Carcinoma or Liver Dominant Metastatic Cancer from Colorectal or Pancreatic Cancers | nivolumab; tadalafil; oral vancomycin | Recruiting |
| NCT04430985 | II | Colorectal Cancer Liver Metastases | Oxaliplatin; 5-Fluorouracil; Leucovorin; Nivolumab; Ipilimumab | Recruiting |
| NCT03698461 | II | Colorectal Cancer Liver Metastases | Atezolizumab; Bevacizumab; Oxaliplatin; Levoleucovorin; 5-FU | Recruiting |
| NCT03310008 | I | Colorectal Cancer with Potentially Resectable Liver Metastases | NKR-2(modified T cells) with FOLFOX | Active, not recruiting |
| NCT04046445 | I/II | Stage IV Colorectal Cancer | ATP128; BI 754091 | Recruiting |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, X.; Ward, S.E.; Zhou, J.; Cheng, A.S.L. Liver Immune Microenvironment and Metastasis from Colorectal Cancer-Pathogenesis and Therapeutic Perspectives. Cancers 2021, 13, 2418. https://doi.org/10.3390/cancers13102418
Zeng X, Ward SE, Zhou J, Cheng ASL. Liver Immune Microenvironment and Metastasis from Colorectal Cancer-Pathogenesis and Therapeutic Perspectives. Cancers. 2021; 13(10):2418. https://doi.org/10.3390/cancers13102418
Chicago/Turabian StyleZeng, Xuezhen, Simon E. Ward, Jingying Zhou, and Alfred S. L. Cheng. 2021. "Liver Immune Microenvironment and Metastasis from Colorectal Cancer-Pathogenesis and Therapeutic Perspectives" Cancers 13, no. 10: 2418. https://doi.org/10.3390/cancers13102418
APA StyleZeng, X., Ward, S. E., Zhou, J., & Cheng, A. S. L. (2021). Liver Immune Microenvironment and Metastasis from Colorectal Cancer-Pathogenesis and Therapeutic Perspectives. Cancers, 13(10), 2418. https://doi.org/10.3390/cancers13102418