
LAG3 and Its Ligands Show Increased Expression in High-Risk Uveal Melanoma

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Chromosome Status
2.3. Illumina Array
2.4. T Lymphocyte Fraction
2.5. Fluidigm qPCR
2.6. Statistical Analysis
3. Results
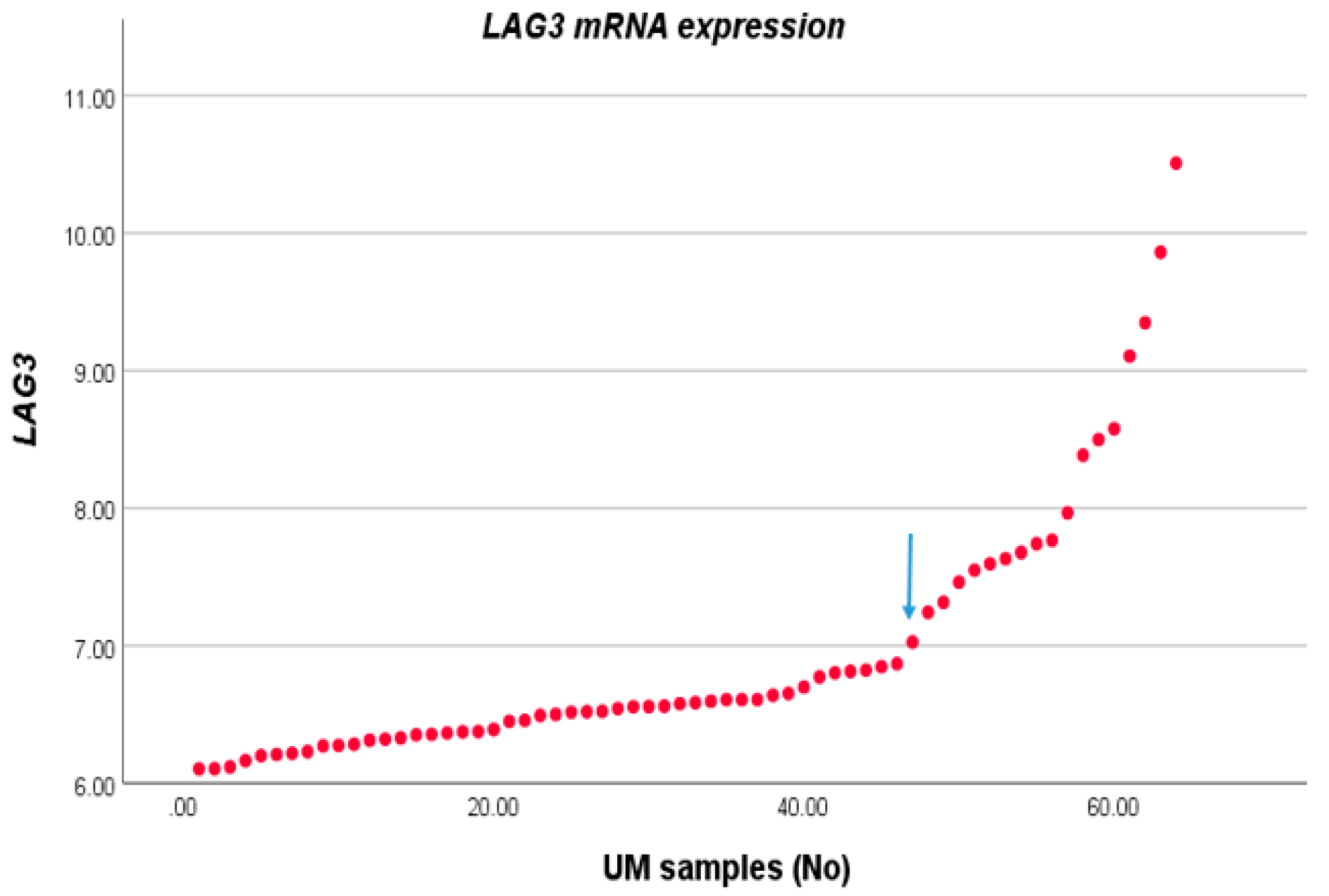
3.1. Association between LAG3 and High-Risk Characteristics of UM
3.2. Association between LAG3 and Cell Surface Ligands
3.3. LAG3 and Inflammatory Phenotype
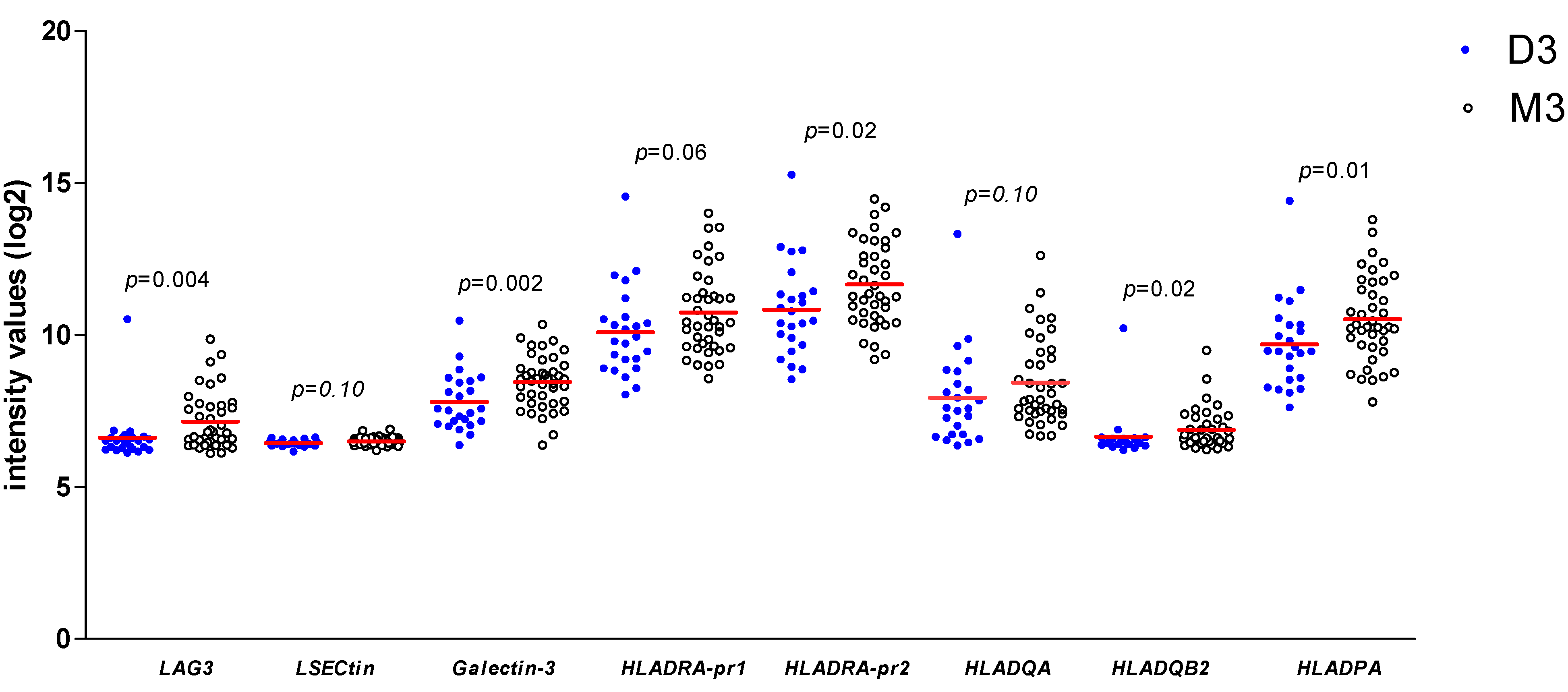
3.4. UM with Monosomy 3 Express Higher Levels of LAG3 Ligands Than with Disomy 3
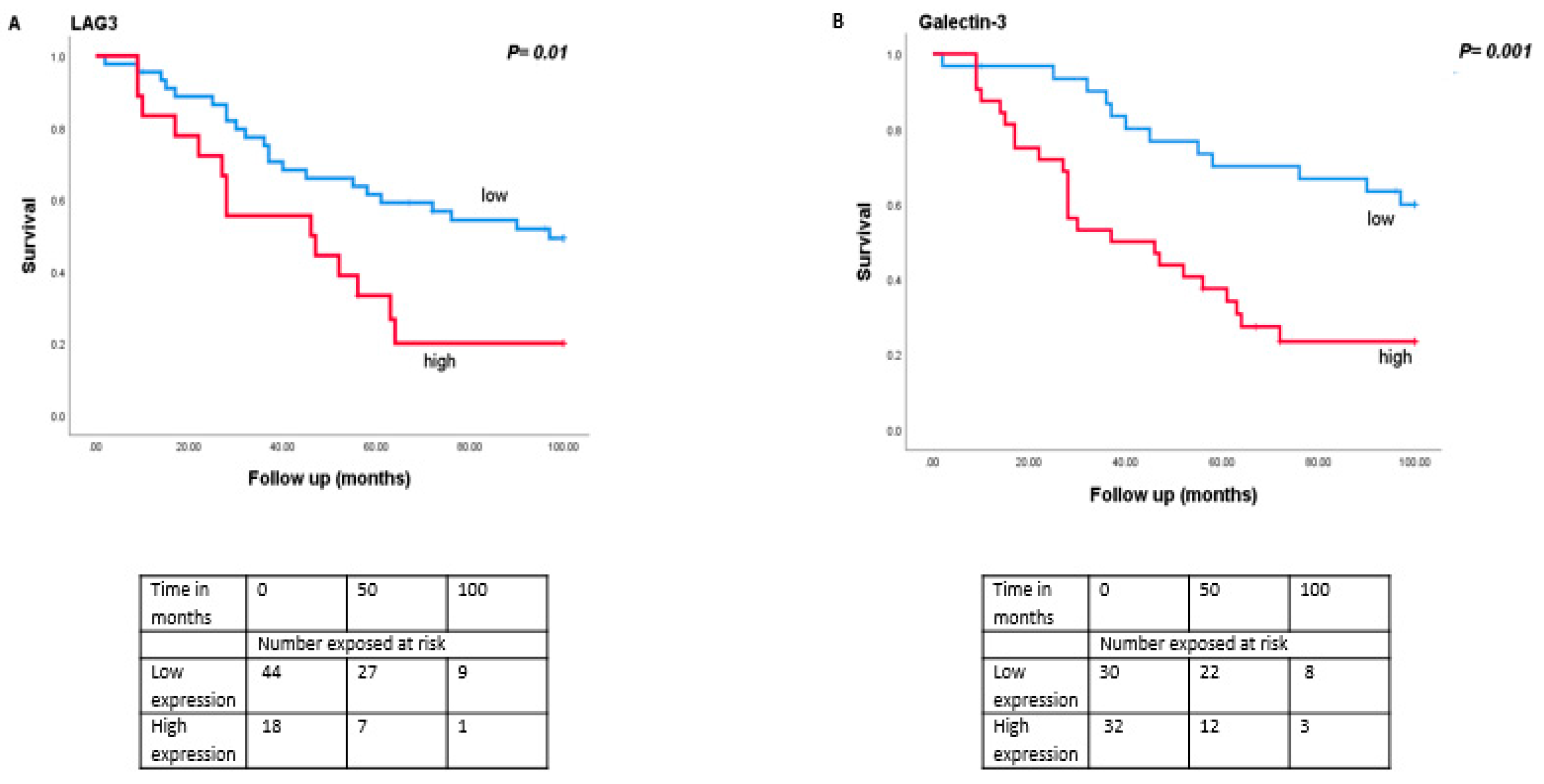
3.5. Association between LAG3 and Its Ligands with Survival
3.6. LAG3 and Immune Modulators
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Metzelaar-Blok, J.A.; Ter Huurne, J.A.; Hurks, H.M.; Keunen, J.E.; Jager, M.J.; Gruis, N. Characterization of melanocortin-1 receptor gene variants in uveal melanoma patients. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1951–1954. [Google Scholar]
- Houtzagers, L.E.; Wierenga, A.P.A.; Ruys, A.A.M.; Luyten, G.P.M.; Jager, M.J. Iris colour and the risk of developing Uveal Melanoma. Int. J. Mol. Sci. 2020, 21, 7172. [Google Scholar] [CrossRef] [PubMed]
- Kujala, E.; Mäkitie, T.; Kivelä, T. Very long-term prognosis of patients with malignant uveal melanoma. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4651–4660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roelofsen, C.D.; Wierenga, A.P.; van Duinen, S.; Verdijk, R.M.; Bleeker, J.; Marinkovic, M.; Luyten, G.P.; Jager, M.J. Five decades of enucleations for uveal melanoma in one center: More tumours with high risk factors, no improvement in survival over time. Ocul. Oncol. Pathol. 2020, 7, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Blom, D.J.; Luyten, G.P.; Mooy, C.; Kerkvliet, S.; Zwinderman, A.H.; Jager, M.J. Human leukocyte antigen class I expression. Marker of poor prognosis in uveal melanoma. Investig. Ophthalmol. Vis. Sci. 1997, 38, 1865–1872. [Google Scholar]
- Ericsson, C.; Seregard, S.; Bartolazzi, A.; Levitskaya, E.; Ferrone, S.; Kiessling, R.; Larsson, O. Association of HLA class I and class II antigen expression and mortality in uveal melanoma. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2153–2156. [Google Scholar]
- Maat, W.; Ly, L.V.; Jordanova, E.S.; De Wolff-Rouendaal, D.; Schalij-Delfos, N.E.; Jager, M.J. Monosomy of chromosome 3 and an inflammatory phenotype occur together in uveal melanoma. Investig. Ophthalmol. Vis. Sci. 2008, 49, 505–510. [Google Scholar] [CrossRef]
- Bronkhorst, I.H.G.; Vu, T.H.K.; Jordanova, E.S.; Luyten, G.P.M.; van der Burg, S.H.; Jager, M.J. Different subsets of tumour-infiltrating lymphocytes correlate with macrophage influx and monosomy 3 in uveal melanoma. Investig. Ophthalmol. Vis. Sci. 2012, 53, 5370–5378. [Google Scholar] [CrossRef] [Green Version]
- Robertson, A.G.; Shih, J.; Yau, C.; Gibb, E.A.; Oba, J.; Mungall, K.L.; Hess, J.M.; Uzunangelov, V.; Walter, V.; Danilova, L.; et al. Integrative analysis identifies four molecular and clinical subsets in Uveal Melanoma. Cancer Cell 2017, 32, 204–220. [Google Scholar] [CrossRef] [Green Version]
- Niederkorn, J.Y. Immune escape mechanisms of intraocular tumours. Prog. Retin. Eye Res. 2009, 28, 329–347. [Google Scholar] [CrossRef] [Green Version]
- Ly, L.V.; Baghat, A.; Versluis, M.; Jordanova, E.S.; Luyten, G.P.M.; Van Rooijen, N.; van Hall, T.; Van Der Velden, P.A.; Jager, M.J. In aged mice, outgrowth of intraocular melanoma depends on proangiogenic M2-type macrophages. J. Immunol. 2010, 185, 3481–3488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lange, M.J.; Nell, R.; Lalai, R.N.; Versluis, M.; Jordanova, E.S.; Luyten, G.P.; Jager, M.J.; Van Der Burg, S.H.; Zoutman, W.H.; van Hall, T.; et al. Digital PCR-Based T-cell Quantification-Assisted deconvolution of the microenvironment reveals that activated macrophages drive tumour inflammation in Uveal Melanoma. Mol. Cancer Res. 2018, 16, 1902–1911. [Google Scholar] [CrossRef] [Green Version]
- Jager, M.J.; Shields, C.L.; Cebulla, C.M.; Abdel-Rahman, M.H.; Grossniklaus, H.E.; Stern, M.-H.; Carvajal, R.D.; Belfort, R.N.; Jia, R.; Shields, J.A.; et al. Uveal melanoma. Nat. Rev. Dis. Primers 2020, 6, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Gezgin, G.; Dogrusöz, M.; Van Essen, T.H.; Kroes, W.G.M.; Luyten, G.P.M.; Van Der Velden, P.A.; Walter, V.; Verdijk, R.M.; van Hall, T.; Van Der Burg, S.H.; et al. Genetic evolution of uveal melanoma guides the development of an inflammatory microenvironment. Cancer Immunol. Immunother. 2017, 66, 903–912. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, C.R.; Kalirai, H.; Sacco, J.J.; Azevedo, R.A.; Duckworth, A.; Slupsky, J.R.; Coulson, J.M.; Coupland, S.E. Loss of BAP1 expression is associated with an immunosuppressive microenvironment in uveal melanoma, with implications for immunotherapy development. J. Pathol. 2020, 250, 420–439. [Google Scholar] [CrossRef] [Green Version]
- Ishida, Y.; Agata, Y.; Shibahara, K.; Honjo, T. Induced expression of PD-1, a novel member of the immunoglobulin gene superfamily, upon programmed cell death. EMBO J. 1992, 11, 3887–3895. [Google Scholar] [CrossRef]
- Wierenga, A.P.; Cao, J.; Luyten, G.P.; Jager, M.J. Immune checkpoint inhibitors in uveal and conjunctival melanoma. Int. Ophthalmol. Clin. 2019, 59, 53–63. [Google Scholar] [CrossRef]
- Triebel, F.; Jitsukawa, S.; Baixeras, E.; Roman-Roman, S.; Genevee, C.; Viegas-Pequignot, E.; Hercend, T. LAG-3, a novel lymphocyte activation gene closely related to CD4. J. Exp. Med. 1990, 171, 1393–1405. [Google Scholar] [CrossRef] [Green Version]
- Huard, B.; Mastrangeli, R.; Prigent, P.; Bruniquel, D.; Donini, S.; El-Tayar, N.; Triebel, F. Characterization of the major histocompatibility complex class II binding site on LAG-3 protein. Proc. Natl. Acad. Sci. USA 1997, 94, 5744–5749. [Google Scholar] [CrossRef] [Green Version]
- Maruhashi, T.; Okazaki, I.-M.; Sugiura, D.; Takahashi, S.; Maeda, T.K.; Shimizu, K.; Okazaki, T. LAG-3 inhibits the activation of CD4+ T cells that recognize stable pMHCII through its conformation-dependent recognition of pMHCII. Nat. Immunol. 2018, 19, 1415–1426. [Google Scholar] [CrossRef] [PubMed]
- Andrews, L.P.; Marciscano, A.E.; Drake, C.G.; Vignali, D.A.A. LAG3 (CD223) as a cancer immunotherapy target. Immunol. Rev. 2017, 276, 80–96. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.-R.; Turnis, M.E.; Goldberg, M.V.; Bankoti, J.; Selby, M.; Nirschl, C.; Bettini, M.L.; Gravano, D.M.; Vogel, P.; Liu, C.L.; et al. Immune inhibitory molecules LAG-3 and PD-1 synergistically regulate T-cell function to promote tumoural immune escape. Cancer Res. 2012, 72, 917–927. [Google Scholar] [CrossRef] [Green Version]
- Durante, M.A.; Rodriguez, D.A.; Kurtenbach, S.; Kuznetsov, J.N.; Sanchez, M.I.; Decatur, C.L.; Snyder, H.; Feun, L.G.; Livingstone, A.S.; Harbour, J.W. Single-cell analysis reveals new evolutionary complexity in uveal melanoma. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ascierto, P.; Ferrucci, P.; Stephens, R.; Del Vecchio, M.; Atkinson, V.; Schmidt, H.; Schachter, J.; Queirolo, P.; Long, G.; Di Giacomo, A.; et al. Phase II multicenter, single arm, open label study of nivolumab in combination with ipilimumab in untreated patients with metastatic uveal melanoma. Ann. Oncol. 2018, 29, viii443. [Google Scholar] [CrossRef]
- Rozeman, E.A.; Prevoo, W.; Meier, M.A.; Sikorska, K.; Van, T.M.; Van De Wiel, B.A.; Van Der Wal, J.E.; Mallo, H.A.; Grijpink-Ongering, L.G.; Broeks, A.; et al. Phase Ib/II trial testing combined radiofrequency ablation and ipilimumab in uveal melanoma (SECIRA-UM). Melanoma Res. 2020, 30, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Duhoux, F.P.; Dirix, L.Y.; Huizing, M.T. Combination of paclitaxel and a LAG-3 fusion protein (eftilagimod alpha), as a first-line chemoimmunotherapy in patients with metastatic breast carcinoma (MBC): Final results from the run-in phase of a placebo-controlled randomized phase II. J. Clin. Oncol. 2018, 36, 1050. [Google Scholar] [CrossRef]
- Ascierto, P.A.; Melero, I.; Bhatia, S.; Bono, P.; Sanborn, R.E.; Lipson, E.J.; Callahan, M.K.; Gajewski, T.; Gomez-Roca, C.A.; Hodi, F.S.; et al. Initial efficacy of anti-lymphocyte activation gene-3 (anti–LAG-3; BMS-986016) in combination with nivolumab (nivo) in PTS with melanoma (MEL) previously treated with anti–PD-1/PD-L1 therapy. J. Clin. Oncol. 2017, 35, 9520. [Google Scholar] [CrossRef]
- Khan, S.; Carvajal, R.D. Dual Immunological Checkpoint Blockade for Uveal Melanoma. J. Clin. Oncol. 2021, 39, 586–599. [Google Scholar] [CrossRef] [PubMed]
- Versluis, M.; De Lange, M.J.; Van Pelt, S.I.; Ruivenkamp, C.A.L.; Kroes, W.G.M.; Cao, J.; Jager, M.J.; Luyten, G.P.M.; Van Der Velden, P.A. Digital PCR Validates 8q Dosage as Prognostic Tool in Uveal Melanoma. PLoS ONE 2015, 10, e0116371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Essen, T.H.; Van Pelt, S.I.; Bronkhorst, I.H.G.; Versluis, M.; Némati, F.; Laurent, C.; Luyten, G.P.M.; Van Hall, T.; Elsen, P.J.V.D.; Van Der Velden, P.A.; et al. Upregulation of HLA Expression in Primary Uveal Melanoma by Infiltrating Leukocytes. PLoS ONE 2016, 11, e0164292. [Google Scholar] [CrossRef]
- Zoutman, W.H.; Nell, R.J.; Versluis, M.; van Steenderen, D.; Lalai, R.N.; Out-Luiting, J.J.; de Lange, M.J.; Vermeer, M.H.; Langerak, A.W.; van der Velden, P.A. Accurate quantification of T cells by measuring loss of germline T-cell receptor loci with generic single duplex droplet digital PCR assays. J. Mol. Diagn. 2017, 19, 236–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wierenga, A.P.A.; Gezgin, G.; Van Beelen, E.; Eikmans, M.; Spruyt-Gerritse, M.; Brouwer, N.J.; Versluis, M.; Verdijk, R.M.; Van Duinen, S.G.; Marinkovic, M.; et al. Soluble HLA in the aqueous humour of uveal melanoma is associated with unfavourable tumour characteristics. Cancers 2019, 11, 1202. [Google Scholar] [CrossRef] [Green Version]
- Jochems, A.; Van Der Kooij, M.K.; Fiocco, M.; Schouwenburg, M.G.; Aarts, M.J.; Van Akkooi, A.C.; Berkmortel, F.W.V.D.; Blank, C.U.; Eertwegh, A.J.V.D.; Franken, M.G.; et al. Metastatic uveal melanoma: Treatment strategies and survival—Results from the Dutch melanoma treatment registry. Cancers 2019, 11, 1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, M.; De Koning, L.; Coupland, S.E.; Jochemsen, A.G.; Marais, R.; Stern, M.-H.; Valente, A.; Barnhill, R.; Cassoux, N.; Evans, A.; et al. So Close, yet so Far: Discrepancies between Uveal and Other Melanomas. A Position Paper from UM Cure 2020. Cancers 2019, 11, 1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eggermont, A.M.; Sileni, V.C.; Grob, J.-J.; Dummer, R.; Wolchok, J.D.; Schmidt, H.; Hamid, O.; Robert, C.; Ascierto, P.A.; Richards, J.M.; et al. Prolonged survival in Stage III melanoma with ipilimumab adjuvant therapy. N. Engl. J. Med. 2016, 375, 1845–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souri, Z.; Wierenga, A.P.A.; Van Weeghel, C.; Van Der Velden, P.A.; Kroes, W.G.M.; Luyten, G.P.M.; Van Der Burg, S.H.; Jochemsen, A.G.; Jager, M.J. Loss of BAP1 is associated with upregulation of the NFKB pathway and increased HLA class I expression in uveal melanoma. Cancers 2019, 11, 1102. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.-Z.; Kim, H.J.; Villasboas, J.C.; Chen, Y.-P.; Price-Troska, T.; Jalali, S.; Wilson, M.; Novak, A.J.; Ansell, S.M. Expression of LAG-3 defines exhaustion of intratumoural PD-1+ T cells and correlates with poor outcome in follicular lymphoma. Oncotarget 2017, 8, 61425–61439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoefsmit, E.P.; Rozeman, E.A.; Van, T.M.; Dimitriadis, P.; Krijgsman, O.; Conway, J.W.; da Silva, I.P.; Van der Wal, J.E.; Ketelaars, S.L.C.; Bresser, K.; et al. Comprehensive analysis of cutaneous and uveal melanoma liver metastases. J. Immunother. Cancer 2020, 8, e001501. [Google Scholar] [CrossRef]
- Liu, W.; Tang, L.; Zhang, G.; Wei, H.; Cui, Y.; Guo, L.; Gou, Z.; Chen, X.; Jiang, D.; Zhu, Y.; et al. Characterization of a novel C-type lectin-like gene, LSECtin: Demonstration of carbohydrate binding and expression in sinusoidal endothelial cells of liver and lymph node. J. Biol. Chem. 2004, 279, 18748–18758. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Liu, J.; Liu, D.; Liu, B.; Wang, M.; Hu, Z.; Du, X.; Tang, L.; He, F. LSECtin expressed on melanoma cells promotes tumour progression by inhibiting antitumour T-cell responses. Cancer Res. 2014, 74, 3418–3428. [Google Scholar] [CrossRef] [Green Version]
- Flotte, T.J.; Springer, T.A.; Thorbecke, G.J. Dendritic cell and macrophage staining by monoclonal antibodies in tissue sections and epidermal sheets. Am. J. Pathol. 1983, 111, 111–112. [Google Scholar]
- Kouo, T.S.; Huang, L.; Pucsek, A.B.; Cao, M.; Solt, S.; Armstrong, T.D.; Jaffee, E.M. Galectin-3 shapes antitumour immune responses by suppressing CD8+ T cells via LAG-3 and inhibiting expansion of plasmacytoid dendritic cells. Cancer Immune Res. 2015, 3, 412–423. [Google Scholar] [CrossRef] [Green Version]
- Prieto, V.G.; Mourad-Zeidan, A.A.; Melnikova, V.; Johnson, M.M.; Lopez, A.; Diwan, A.H.; Lazar, A.; Shen, S.S.; Zhang, P.S.; Reed, J.A.; et al. Galectin-3 expression is associated with tumour progression and pattern of sun exposure in melanoma. Clin. Cancer Res. 2006, 12, 6709–6715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, S.; Tian, Y.; Wang, Y.; Zhang, Q.; Zhou, X.; Meng, X.; Song, N. Prognostic role of galectin-3 expression in patients with solid tumours: A meta-analysis of 36 eligible studies. Cancer Cell Internat. 2018, 18, 1–5. [Google Scholar] [CrossRef]
- Markowska, A.I.; Jefferies, K.C.; Panjwani, N. Galectin-3 protein modulates cell surface expression and activation of vascular endothelial growth factor receptor 2 in human endothelial cells. J. Biol. Chem. 2011, 34, 29913–29921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korman, A.J.; Boss, J.M.; Spies, T.; Sorrentino, R.; Okada, K.; Strominger, J.L. Genetic Complexity and Expression of Human Class II Histocompatibility Antigens. Immunol. Rev. 1985, 85, 45–86. [Google Scholar] [CrossRef]
- Trowsdale, J.; Young, J.A.T.; Kelly, A.R.; Austin, P.J.; Carson, S.; Meunter, H.; So, A.; Erlich, H.A.; Spielman, R.S.; Bodmer, J.; et al. Structure, sequence and polymorphism in the HLA-D region. Immunol. Rev. 1985, 85, 5–43. [Google Scholar] [CrossRef]
- Hemon, P.; Jean-Louis, F.; Ramgolam, K.; Brignone, C.; Viguier, M.; Bachelez, H.; Triebel, F.; Charron, D.; Aoudjit, F.; Al-Daccak, R.; et al. MHC Class II Engagement by Its Ligand LAG-3 (CD223) Contributes to Melanoma Resistance to Apoptosis. J. Immunol. 2011, 186, 5173–5183. [Google Scholar] [CrossRef] [Green Version]
- Jager, M.J.; van der Pol, J.P.; de Wolff-Rouendaal, D.; de Jong, P.T.; Ruiter, D.J. Decreased expression of HLA class II antigens on human uveal melanoma cells after in vivo X-ray irradiation. Am. J. Ophthalmol. 1988, 105, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Karp, D.R.; Teletski, C.L.; Jaraquemada, D.; Maloy, W.L.; Coligan, J.E.; Long, E.O. Structural requirements for pairing of alpha and beta chains in HLA-DR and HLA-DP molecules. J. Exp. Med. 1990, 171, 615–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulvestad, E.; Williams, K.; Bø, L.; Trapp, B.; Antel, J.; Mørk, S. HLA class II molecules (HLA-DR,-DP,-DQ) on cells in the human CNS studied in situ and in vitro. Immunology 1994, 82, 535–541. [Google Scholar] [PubMed]
- Grifoni, A.; Moore, E.; Voic, H.; Sidney, J.; Phillips, E.; Jadi, R.; Mallal, S.; De Silva, A.D.; De Silva, A.M.; Peters, B.; et al. Characterization of magnitude and antigen specificity of HLA-DP, DQ, and DRB3/4/5 restricted DENV-specific CD4+ T cell responses. Front. Immunol. 2019, 10, 1568–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Waard-Siebinga, I.; Creyghton, W.M.; Kool, J.; Jager, M.J. Effects of interferon alfa and gamma on human uveal melanoma cells in vitro. Br. J. Ophthalmol. 1995, 79, 847–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Patients (%) | |||
|---|---|---|---|
| Clinical and Histopathologic Characteristics | |||
| L | H | p | |
| Gender | |||
| Male | 23 (36%) | 10 (16%) | |
| Female | 23 (36%) | 8 (12%) | 0.69 |
| Age (Years) at Enucleation (SD) | |||
| ≤60 | 26 (41%) | 5 (8%) | |
| >60 | 20 (31%) | 13 (20%) | 0.04 |
| Cell Type | |||
| Spindle | 21 (33%) | 1 (1%) | |
| Mixed/epithelioid | 25 (37%) | 17 (26%) | 0.002 |
| Largest Tumour Diameter (LBD) in mm | |||
| <13.0 (median) | 20 (31%) | 7 (11%) | |
| ≥13.0 (median) | 26 (41%) | 11 (17%) | 0.74 |
| Tumour Prominence in mm | |||
| <8.0 (median) | 24 (37%) | 5 (8%) | |
| ≥8.0 (median) | 22 (34%) | 13 (20%) | 0.08 |
| Ciliary Body Involvement | |||
| Not involved | 31 (48%) | 9 (14%) | |
| Involved | 15 (23%) | 9 (14%) | 0.2 |
| cTNM Stage (n = 62) | |||
| Stage I-IIB | 27 (43%) | 10 (16%) | |
| Stage IIIA-IIIB | 17 (27%) | 8 (13%) | 0.67 |
| Metastasis | |||
| No | 22 (34%) | 4 (6%) | |
| Yes | 24 (37%) | 14 (22%) | 0.06 |
| BAP1 status (n = 55) | |||
| BAP1 staining positive | 24 (44%) | 1 (2%) | |
| BAP1 staining negative | 15 (27%) | 15 (27%) | 0.001 |
| Immune Modulator | D3 (n = 9) | M3 (n = 6) | |
|---|---|---|---|
| Mean ± SD | Mean ± SD | p | |
| LAG3 | 4 ± 3 | 98 ± 80 | 0.004 |
| Galectin-3 | 1179 ± 1489 | 581 ± 345 | 0.61 |
| HLA-DR | 721 ± 606 | 4953 ± 4751 | 0.004 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souri, Z.; Wierenga, A.P.A.; Kroes, W.G.M.; van der Velden, P.A.; Verdijk, R.M.; Eikmans, M.; Luyten, G.P.M.; Jager, M.J. LAG3 and Its Ligands Show Increased Expression in High-Risk Uveal Melanoma. Cancers 2021, 13, 4445. https://doi.org/10.3390/cancers13174445
Souri Z, Wierenga APA, Kroes WGM, van der Velden PA, Verdijk RM, Eikmans M, Luyten GPM, Jager MJ. LAG3 and Its Ligands Show Increased Expression in High-Risk Uveal Melanoma. Cancers. 2021; 13(17):4445. https://doi.org/10.3390/cancers13174445
Chicago/Turabian StyleSouri, Zahra, Annemijn P. A. Wierenga, Wilma G. M. Kroes, Pieter A. van der Velden, Robert M. Verdijk, Michael Eikmans, Gregorius P. M. Luyten, and Martine J. Jager. 2021. "LAG3 and Its Ligands Show Increased Expression in High-Risk Uveal Melanoma" Cancers 13, no. 17: 4445. https://doi.org/10.3390/cancers13174445
APA StyleSouri, Z., Wierenga, A. P. A., Kroes, W. G. M., van der Velden, P. A., Verdijk, R. M., Eikmans, M., Luyten, G. P. M., & Jager, M. J. (2021). LAG3 and Its Ligands Show Increased Expression in High-Risk Uveal Melanoma. Cancers, 13(17), 4445. https://doi.org/10.3390/cancers13174445