
Extracellular Vesicle-Associated miRNAs and Chemoresistance: A Systematic Review



Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
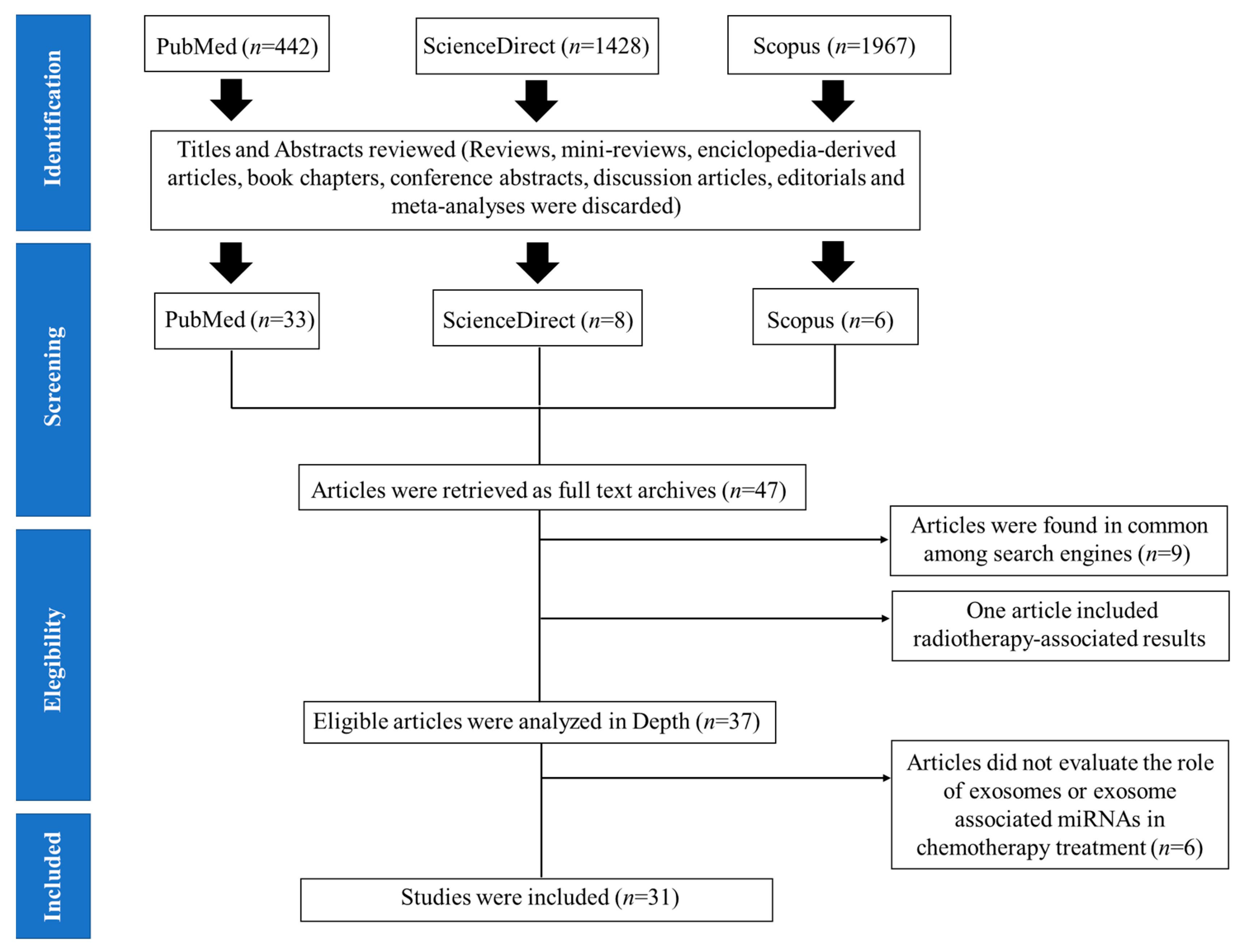
2.1. Search Strategy
2.2. Eligibility Criteria
2.3. Data Extraction
3. Results
3.1. Literature Search and Characteristics
3.2. Candidate Exosomal-Associated miRNAs Involved in Chemotherapy Response
3.2.1. Colorectal Cancer
3.2.2. Ovarian Cancer
3.2.3. Breast Cancer
3.2.4. Renal Cell Carcinoma
3.2.5. Lung Cancer
3.2.6. Multiple Myeloma
3.2.7. Lymphoma
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ma, Y.; Yuwen, D.; Chen, J.; Zheng, B.; Gao, J.; Fan, M.; Xue, W.; Wang, Y.; Li, W.; Shu, Y.; et al. Exosomal Transfer of Cisplatin-Induced miR-425-3p Confers Cisplatin Resistance in NSCLC through Activating Autophagy. Int. J. Nanomed. 2019, 14, 8121–8132. [Google Scholar] [CrossRef] [Green Version]
- Strauss, J.; Figg, W.D. Using Epigenetic Therapy to Overcome Chemotherapy Resistance. Anticancer. Res. 2016, 36, 1–4. [Google Scholar]
- Iorio, M.; Croce, C.M. MicroRNA dysregulation in cancer: Diagnostics, monitoring and therapeutics. A comprehensive review. EMBO Mol. Med. 2012, 4, 143–159. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Kuhlmann, J.D.; Chebouti, I.; Kimmig, R.; Buderath, P.; Reuter, M.; Puppel, S.-H.; Wimberger, P.; Kasimir-Bauer, S. Extracellular vesicle-associated miRNAs in ovarian cancer—Design of an integrated NGS-based workflow for the identification of blood-based biomarkers for platinum-resistance. Clin. Chem. Lab. Med. 2018, 57, 1053–1062. [Google Scholar] [CrossRef]
- Lagos-Quintana, M.; Rauhut, R.; Lendeckel, W.; Tuschl, T. Identification of Novel Genes Coding for Small Expressed RNAs. Science 2001, 294, 853–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuji, T.; Umeda, Y.; Nyuya, A.; Taniguchi, F.; Kawai, T.; Yasui, K.; Toshima, T.; Yoshida, K.; Fujiwara, T.; Goel, A.; et al. Detection of circulating microRNAs with Ago2 complexes to monitor the tumor dynamics of colorectal cancer patients during chemotherapy. Int. J. Cancer 2018, 144, 2169–2180. [Google Scholar] [CrossRef] [PubMed]
- Mir, B.; Goettsch, C. Extracellular Vesicles as Delivery Vehicles of Specific Cellular Cargo. Cells 2020, 9, 1601. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.; Chevillet, J.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.; Bennett, C.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [Green Version]
- Shao, H.; Im, H.; Castro, C.M.; Breakefield, X.; Weissleder, R.; Lee, H. New Technologies for Analysis of Extracellular Vesicles. Chem. Rev. 2018, 118, 1917–1950. [Google Scholar] [CrossRef]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef]
- Qin, X.; Yu, S.; Zhou, L.; Shi, M.; Hu, Y.; Xu, X.; Shen, B.; Liu, S.; Yan, D.; Feng, J. Cisplatin-resistant lung cancer cell–derived exosomes increase cisplatin resistance of recipient cells in exosomal miR-100–5p-dependent manner. Int. J. Nanomed. 2017, 12, 3721–3733. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Yuan, T.; Liang, M.; Du, M.; Xia, S.; Dittmar, R.; Wang, D.; See, W.; Costello, B.A.; Quevedo, F.; et al. Exosomal miR-1290 and miR-375 as Prognostic Markers in Castration-resistant Prostate Cancer. Eur. Urol. 2014, 67, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuwen, D.-L.; Sheng, B.-B.; Liu, J.; Wenyu, W.; Shu, Y.-Q. MiR-146a-5p level in serum exosomes predicts therapeutic effect of cisplatin in non-small cell lung cancer. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2650–2658. [Google Scholar] [PubMed]
- Li, Y.; Liang, Y.; Sang, Y.; Song, X.; Zhang, H.; Liu, Y.; Jiang, L.; Yang, Q. MiR-770 suppresses the chemo-resistance and metastasis of triple negative breast cancer via direct targeting of STMN1. Cell Death Dis. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Wang, G.; Zhao, W.; Wang, H.; Qiu, G.; Jiang, Z.; Wei, G.; Li, X. Exosomal MiR-744 Inhibits Proliferation and Sorafenib Chemoresistance in Hepatocellular Carcinoma by Targeting PAX2. Med. Sci. Monit. 2019, 25, 7209–7217. [Google Scholar] [CrossRef]
- Santos, P.; Almeida, F. Role of Exosomal miRNAs and the Tumor Microenvironment in Drug Resistance. Cells 2020, 9, 1450. [Google Scholar] [CrossRef]
- Guo, Q.-R.; Wang, H.; Yan, Y.-D.; Liu, Y.; Su, C.-Y.; Chen, H.-B.; Yan, Y.-Y.; Adhikari, R.; Wu, Q.; Zhang, J.-Y. The Role of Exosomal microRNA in Cancer Drug Resistance. Front. Oncol. 2020, 10. [Google Scholar] [CrossRef]
- Lefebvre, C.; Duffy, S. Peer review of searches for studies for health technology assessments, systematic reviews, and other evidence syntheses. Int. J. Technol. Assess. Health Care 2021, 37, e64. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; The PRISMA Group. Reprint—Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. Phys. Ther. 2009, 89, 873–880. [Google Scholar] [CrossRef] [PubMed]
- He, J.; He, J.; Min, L.; He, Y.; Guan, H.; Wang, J.; Peng, X. Extracellular vesicles transmitted miR-31-5p promotes sorafenib resistance by targeting MLH1 in renal cell carcinoma. Int. J. Cancer 2019, 146, 1052–1063. [Google Scholar] [CrossRef] [PubMed]
- Yuwen, D.; Ma, Y.; Wang, D.; Gao, J.; Li, X.; Xue, W.; Fan, M.; Xu, Q.; Shen, Y.; Shu, Y. Prognostic Role of Circulating Exosomal miR-425-3p for the Response of NSCLC to Platinum-Based Chemotherapy. Cancer Epidemiology Biomarkers Prev. 2018, 28, 163–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Zhong, M.; Zeng, S.; Wang, L.; Liu, P.; Xiao, X.; Liu, Y. Exosome-derived miRNAs as predictive biomarkers for diffuse large B-cell lymphoma chemotherapy resistance. Epigenomics 2019, 11, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, X.; Du, L.; Wang, Y.; Liu, X.; Tian, H.; Wang, L.; Li, P.; Zhao, Y.; Duan, W.; et al. Exosome-transmitted miR-128-3p increase chemosensitivity of oxaliplatin-resistant colorectal cancer. Mol. Cancer 2019, 18, 1–17. [Google Scholar] [CrossRef] [Green Version]
- de Miguel Pérez, D.; Martínez, A.R.; Palomo, A.O.; Ureña, M.D.; Puche, J.L.G.; Remacho, A.R.; Hernandez, J.E.; Acosta, J.A.L.; Sánchez, F.G.O. Extracellular vesicle-miRNAs as liquid biopsy biomarkers for disease identification and prog-nosis in meta-static colorectal cancer patients. Sci. Rep. 2020, 10, 3974. [Google Scholar] [CrossRef] [Green Version]
- Jin, G.; Liu, Y.; Zhang, J.; Bian, Z.; Yao, S.; Fei, B.; Zhou, L.; Yin, Y.; Huang, Z. A panel of serum exosomal microRNAs as predictive markers for chemoresistance in advanced colorectal cancer. Cancer Chemother. Pharmacol. 2019, 84, 315–325. [Google Scholar] [CrossRef]
- Yang, J.-K.; Yang, J.-P.; Tong, J.; Jing, S.-Y.; Fan, B.; Wang, F.; Sun, G.-Z.; Jiao, B.-H. Exosomal miR-221 targets DNM3 to induce tumor progression and temozolomide resistance in glioma. J. Neuro-Oncol. 2016, 131, 255–265. [Google Scholar] [CrossRef]
- Hu, Y.; Qi, C.; Liu, X.; Zhang, C.; Gao, J.; Wu, Y.; Yang, J.; Zhao, Q.; Li, J.; Wang, X.; et al. Malignant ascites-derived exosomes promote peritoneal tumor cell dissemination and reveal a distinct miRNA signature in advanced gastric cancer. Cancer Lett. 2019, 457, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, C.; Rani, S.; O’Driscoll, L. miR-34a is an intracellular and exosomal predictive biomarker for response to docetaxel with clinical relevance to prostate cancer progression. Prostate 2014, 74, 1320–1334. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Liu, M.; Qu, S.; Ma, J.; Zhang, Y.; Shi, T.; Wen, H.; Yang, Y.; Wang, S.; Wang, J.; et al. Exosomal microRNA-32-5p induces multidrug resistance in hepatocellular carcinoma via the PI3K/Akt pathway. J. Exp. Clin. Cancer Res. 2018, 37, 52. [Google Scholar] [CrossRef] [Green Version]
- Kanlikilicer, P.; Bayraktar, R.; Denizli, M.; Rashed, M.H.; Ivan, C.; Aslan, B.; Mitra, R.; Karagoz, K.; Bayraktar, E.; Zhang, X.; et al. Exosomal miRNA confers chemo resistance via targeting Cav1/p-gp/M2-type macrophage axis in ovarian cancer. EBio-Med. 2018, 38, 100–112. [Google Scholar]
- Alharbi, M.; Sharma, S.; Guanzon, D.; Lai, A.; Zuniga, F.A.; Shiddiky, M.J.; Yamauchi, Y.; Salas-Burgos, A.; He, Y.; Pejovic, T.; et al. miRNa signature in small extracellular vesicles and their association with platinum resistance and cancer recurrence in ovarian cancer. Nanomed. Nanotechnol. Biol. Med. 2020, 28, 102207. [Google Scholar] [CrossRef] [PubMed]
- Pink, R.; Samuel, P.; Massa, D.; Caley, D.P.; Brooks, S.A.; Carter, D.R.F. The passenger strand, miR-21-3p, plays a role in mediating cisplatin resistance in ovarian cancer cells. Gynecol. Oncol. 2015, 137, 143–151. [Google Scholar] [CrossRef]
- Yeung, C.L.A.; Co, N.-N.; Tsuruga, T.; Yeung, T.-L.; Kwan, S.Y.; Leung, C.S.; Li, Y.; Lu, E.S.; Kwan, K.; Wong, K.-K.; et al. Exosomal transfer of stroma-derived miR21 confers paclitaxel resistance in ovarian cancer cells through targeting APAF1. Nat. Commun. 2016, 7, 11150. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Shen, H.; Yin, X.; Yang, M.; Wei, H.; Chen, Q.; Feng, F.; Liu, Y.; Xu, W.; Li, Y. Macrophages derived exosomes deliver miR-223 to epithelial ovarian cancer cells to elicit a chemoresistant phenotype. J. Exp. Clin. Cancer Res. 2019, 38, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Stevic, I.; Müller, V.; Weber, K.; Fasching, P.A.; Karn, T.; Marmé, F.; Schem, C.; Stickeler, E.; Denkert, C.; Van Mackelenbergh, M.; et al. Specific microRNA signatures in exosomes of triple-negative and HER2-positive breast cancer patients undergoing neoadjuvant therapy within the GeparSixto trial. BMC Med. 2018, 16, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Martínez, A.; De Miguel-Pérez, D.; Ortega, F.G.; García-Puche, J.L.; Robles-Fernández, I.; Exposito, J.; Martorell-Marugan, J.; Carmona-Sáez, P.; Garrido-Navas, M.D.C.; Rolfo, C.; et al. Exosomal miRNA profile as complementary tool in the diagnostic and prediction of treatment response in localized breast cancer under neoadjuvant chemotherapy. Breast Cancer Res. 2019, 21, 1–9. [Google Scholar] [CrossRef]
- Zhong, S.; Chen, X.; Wang, D.; Zhang, X.; Shen, H.; Yang, S.; Lv, M.; Tang, J.; Zhao, J. MicroRNA expression profiles of drug-resistance breast cancer cells and their exosomes. Oncotarget 2016, 7, 19601–19609. [Google Scholar] [CrossRef]
- Bovy, N.; Blomme, B.; Frères, P.; Dederen, S.; Nivelles, O.; Lion, M.; Carnet, O.; Martial, J.A.; Noël, A.; Thiry, M.; et al. Endo-thelial exosomes contribute to the antitumor response during breast cancer neoadjuvant chemotherapy via mi-croRNA transfer. Oncotarget 2015, 6, 10253. [Google Scholar] [CrossRef] [Green Version]
- Mikamori, M.; Yamada, D.; Eguchi, H.; Hasegawa, S.; Kishimoto, T.; Tomimaru, Y.; Asaoka, T.; Noda, T.; Wada, H.; Ka-wamoto, K.; et al. MicroRNA-155 controls exosome synthesis and promotes gemcitabine resistance in pancreatic ductal adenocarci-noma. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Svedman, F.C.; Lohcharoenkal, W.; Bottai, M.; Brage, S.E.; Sonkoly, E.; Hansson, J.; Pivarcsi, A.; Eriksson, H. Extracellular microvesicle microRNAs as predictive biomarkers for targeted therapy in metastastic cutaneous malignant melanoma. PLoS ONE 2018, 13, e0206942. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Ma, C.; Zhou, T.; Dong, X.; Luo, Q.; Geng, L.; Ding, L.; Zhang, Y.; Zhang, L.; Li, N.; et al. Exosomes derived from gemcitabine-resistant cells transfer malignant phenotypic traits via delivery of miRNA-222-3p. Mol. Cancer 2017, 16, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, X.-B.; Gu, Y.; Sun, D.-L.; Ding, L.-Y.; Yuan, X.-G.; Jiang, H.-W.; Wu, Z.-X. Effect of rituximab combined with chemo-therapy on the expression of serum exosome miR-451a in patients with diffuse large b-cell lymphoma. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1620–1625. [Google Scholar] [PubMed]
- Zeng, A.; Wei, Z.; Yan, W.; Yin, J.; Huang, X.; Zhou, X.; Li, R.; Shen, F.; Wu, W.; Wang, X.; et al. Exosomal transfer of miR-151a enhances chemosensitivity to temozolomide in drug-resistant glioblastoma. Cancer Lett. 2018, 436, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Pan, L.; Xiang, B.; Zhu, H.; Wu, Y.; Chen, M.; Guan, P.; Zou, X.; Valencia, C.A.; Dong, B.; et al. Potential role of exosome-associated microRNA panels and in vivo environment to predict drug resistance for patients with multiple myeloma. Oncotarget 2016, 7, 30876–30891. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Peng, Y.-Z.; Li, C.-G.; Jiang, H.-W.; Mei, H.; Hu, Y. Prognostic and Clinicopathological Significance of MiR-155 in Hematologic Malignancies: A Systematic Review and Meta-analysis. J. Cancer 2019, 10, 654–664. [Google Scholar] [CrossRef] [Green Version]
- Zare, N.; Javanmard, S.H.; Mehrzad, V.; Eskandari, N.; Kefayat, A. Evaluation of exosomal miR-155, let-7g and let-7i levels as a potential noninvasive biomarker among refractory/relapsed patients, responsive patients and patients receiving R-CHOP. Leuk. Lymphoma 2019, 60, 1877–1889. [Google Scholar] [CrossRef]
- Zhang, L.; Cao, D.; Tang, L.; Sun, C.; Hu, Y. A panel of circulating miRNAs as diagnostic biomarkers for screening multiple myeloma: A systematic review and meta-analysis. Int. J. Lab Hematol. 2016, 38, 589–599. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Sawada, K.; Yoshimura, A.; Kinose, Y.; Nakatsuka, E.; Kimura, T. Clinical relevance of circulating cell-free microRNAs in ovarian cancer. Mol. Cancer 2016, 15, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Teng, C.; Zheng, H. Low expression of microRNA-1908 predicts a poor prognosis for patients with ovarian cancer. Oncol. Lett. 2017, 14, 4277–4281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staicu, C.E.; Predescu, D.-V.; Rusu, C.M.; Radu, B.M.; Cretoiu, D.; Suciu, N.; Crețoiu, S.M.; Voinea, S.-C. Role of microRNAs as Clinical Cancer Biomarkers for Ovarian Cancer: A Short Overview. Cells 2020, 9, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazici, H. Functions of miRNAs in the Development, Diagnosis, and Treatment of Ovarian Carcinoma. In Current Trends in Cancer Management; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Keniry, M.; Parsons, R. The role of PTEN signaling perturbations in cancer and in targeted therapy. Oncogene 2008, 27, 5477–5485. [Google Scholar] [CrossRef] [Green Version]
- Bagnoli, M.; De Cecco, L.; Granata, A.; Nicoletti, R.; Marchesi, E.; Alberti, P.; Valeri, B.; Libra, M.; Barbareschi, M.; Raspagliesi, F.; et al. Identification of a chrXq27.3 microRNA cluster associated with early relapse in advanced stage ovarian cancer pa-tients. Oncotarget 2011, 2, 1265–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, K.; Lowry, M.C.; Corcoran, C.; Martinez, V.G.; Daly, M.; Rani, S.; Gallagher, W.; Radomski, M.W.; MacLeod, R.A.; O’Driscoll, L. miR-134 in extracellular vesicles reduces triple-negative breast cancer aggression and increases drug sensitivity. Oncotarget 2015, 6, 32774–32789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasanpour, P.; Sandoughdaran, S.; Mosavi-Jarrahi, A.; Malekzadeh, M. Predictors of Pathological Complete Response to Neoadjuvant Chemotherapy in Iranian Breast Cancer Patients. Asian Pac. J. Cancer Prev. 2018, 19, 2423–2427. [Google Scholar] [CrossRef]
- Qu, R.; Hu, C.; Tang, Y.; Yu, Q.; Shi, G. Long non-coding RNA BLACAT1 induces tamoxifen resistance in human breast cancer by regulating miR-503/Bcl-2 axis. Cancer Manag. Res. 2020, 12, 1771. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Barrueco, R.; Nekritz, E.A.; Bertucci, F.; Yu, J.; Sanchez-Garcia, F.; Zeleke, T.Z.; Gorbatenko, A.; Birnbaum, D.; Ezhkova, E.; Cordon-Cardo, C.; et al. miR-424 (322)/503 is a breast cancer tumor suppressor whose loss promotes resistance to chemotherapy. Genes Dev. 2017, 31, 553–566. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.-L.; Wang, H.; Liu, J.; Wang, Z.-X. MicroRNA-21 (miR-21) expression promotes growth, metastasis, and chemo- or radioresistance in non-small cell lung cancer cells by targeting PTEN. Mol. Cell. Biochem. 2012, 372, 35–45. [Google Scholar] [CrossRef]
- Shi, G.-H.; Ye, D.-W.; Yao, X.-D.; Zhang, S.-L.; Dai, B.; Zhang, H.-L.; Shen, Y.-J.; Zhu, Y.; Zhu, Y.-P.; Xiao, W.-J.; et al. Involvement of microRNA-21 in mediating chemo-resistance to docetaxel in androgen-independent prostate cancer PC3 cells. Acta Pharmacol. Sin. 2010, 31, 867–873. [Google Scholar] [CrossRef]
- Vandewalle, V.; Essaghir, A.; Bollaert, E.; Lenglez, S.; Graux, C.; Schoemans, H.; Saussoy, P.; Michaux, L.; Valk, P.J.; Demoulin, J.B.; et al. miR-15a-5p and miR-21-5p contribute to chemoresistance in cytogenetically normal acute myeloid leukaemia by targeting PDCD4, ARL2 and BTG2. J. Cell. Mol. Med. 2020, 25, 575–585. [Google Scholar] [CrossRef]
- Wang, P.; Zhuang, L.; Zhang, J.; Fan, J.; Luo, J.; Chen, H.; Wang, K.; Liu, L.; Chen, Z.; Meng, Z. The serum miR-21 level serves as a predictor for the chemosensitivity of advanced pancreatic cancer, and miR-21 expression confers chemoresistance by targeting FasL. Mol. Oncol. 2012, 7, 334–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, L.-Q.; Yang, X.-W.; Chen, Y.-B.; Zhang, D.-W.; Jiang, X.-F.; Xue, P. Exosomal miR-21 regulates the TETs/PTENp1/PTEN pathway to promote hepatocellular carcinoma growth. Mol. Cancer 2019, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.-H.; Tian, D.; Yang, Z.-C.; Li, J.-L. Exosomal miR-21 promotes proliferation, invasion and therapy resistance of colon adenocarcinoma cells through its target PDCD4. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bliss, S.A.; Sinha, G.; Sandiford, O.A.; Williams, L.M.; Engelberth, D.J.; Guiro, K.; Isenalumhe, L.L.; Greco, S.J.; Ayer, S.; Bryan, M.; et al. Mesenchymal Stem Cell–Derived Exosomes Stimulate Cycling Quiescence and Early Breast Cancer Dormancy in Bone Marrow. Cancer Res. 2016, 76, 5832–5844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Tang, J. Circulating exosomal microRNA profiling to depict mechanisms of chemotherapy resistance among triple negative breast cancer. J. Clin. Oncol. 2018, 36, e13124. [Google Scholar] [CrossRef]
- Wang, D.-D.; Yang, S.-J.; Chen, X.; Shen, H.-Y.; Luo, L.-J.; Zhang, X.-H.; Zhong, S.-L.; Zhao, J.-H.; Tang, J.-H. miR-222 induces Adriamycin resistance in breast cancer through PTEN/Akt/p27kip1 pathway. Tumor Biol. 2016, 37, 15315–15324. [Google Scholar] [CrossRef]
- Santos, J.C.; Lima, N.; Sarian, L.; Matheu, A.; Ribeiro, M.L.; Derchain, S. Exosome-mediated breast cancer chemoresistance via miR-155 transfer. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Andreu, Z.; Rivas, E.; Sanguino-Pascual, A.; Lamana, A.; Marazuela, M.; González-Alvaro, I.; Sánchez-Madrid, F.; De La Fuente, H.; Yáñez-Mó, M. Comparative analysis of EV isolation procedures for miRNAs detection in serum samples. J. Extracell. Vesicles 2016, 5, 31655. [Google Scholar] [CrossRef]
- Palviainen, M.; Saraswat, M.; Varga, Z.; Kitka, D.; Neuvonen, M.; Puhka, M.; Joenväärä, S.; Renkonen, R.; Nieuwland, R.; Takatalo, M.; et al. Extracellular vesicles from human plasma and serum are carriers of extravesicular cargo—Implications for biomarker discovery. PLoS ONE 2020, 15, e0236439. [Google Scholar] [CrossRef]
- Srinivasan, S.; Yeri, A.; Cheah, P.S.; Chung, A.; Danielson, K.; De Hoff, P.; Filant, J.; Laurent, C.D.; Laurent, L.D.; Magee, R.; et al. Small RNA Sequencing across Diverse Biofluids Identifies Optimal Methods for exRNA Isolation. Cell 2019, 177, 446–462.e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, M.; Wang, H.; Yao, X.; Zhang, D.; Xie, Y.; Cui, R.; Zhang, X. Circulating MicroRNAs in Cancer: Potential and Challenge. Front. Genet. 2019, 10, 626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Peng, R.; Wang, J.; Qin, Z.; Xue, L. Circulating microRNAs as potential cancer biomarkers: The advantage and disadvantage. Clin. Epigenetics 2018, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Publication | Search Criteria |
|---|---|
| Language | English |
| Time period | January 2010–September 2020 |
| Subject | Human |
| Study type | Retrospective, Cohort, and Case-control |
| Excluded | Reviews, Encyclopedia-derived articles, Book chapters, Conference abstracts, Discussion articles, Editorials, Mini-reviews, and Meta-analyses |
| Keywords | Extracellular vesicles, Exosomes, EVs, miRNA, Chemotherapy, Chemoresistance, Cancer recurrence |
| Research Article | Type of Cancer | Drug | Biofluid | Exosomal miRNA | EV Isolation | miRNA Profiling | Findings |
|---|---|---|---|---|---|---|---|
| Bovy et al., 2015 [40] | Breast | Cyclophosphamide or Fluouracil; Epirubicin; Docetaxel or Paclitaxel | Plasma | Plasmatic miR-503 | None | qRT-PCR | ↑ Only after neoadjuvant therapy. |
| Rodriguez-Martínez et al., 2019 [38] | Breast | Doxorubicin/cyclophosphamide Docetaxel, Trastuzumab, Tamoxifen, Anastrozole, Letrozole, Goserelin | Serum | miR-21, miR-105, miR-221 and miR-222 | UC | qRT-PCR | ↑ miR-21 and miR-105 in cancer patients versus healthy donors. Exosomal miRNA-222 levels correlated with clinical and pathological variables such as progesterone receptor status (p = 0.017) and Ki67 (p = 0.05). ↓ miR-221 during neoadjuvant therapy in patients with compromised lymph nodes. |
| Stevic et al., 2018 [37] | Breast | Paclitaxel, Doxorubicin and Carboplatin | Plasma | miR-155, miR-301, miR-27a, miR-376a miR-376c | ExoQuick | Microarray | ↑ miR-155 and miR-301 predicted efficient response towards neoadjuvant therapy. |
| Zhong et al., 2016 [39] | Breast | Docetaxel, Epirubicin, Pemetrexed disodium and, Cytoxan | Tumor tissue | miR-574, miR-210-3p, miR-138-5p, miR-4258, miR-744-5p, miR-7107-5p, miR-6780b-3p, miR-3178, miR-4298, miR-423-5p, miR-7847-3p and miR-4443 | None | qRT-PCR | ↑ miR-574-3p associated with progressive disease. |
| De Miguel Pérez et al., 2020 [26] | Metastatic colorectal cancer | FOLFOX-6m (OXA, Leucovorin, 5-FU) plus Bevacizumab | Serum | miR-21 miR-92a miR-222 miR-19b | UC | qRT-PCR | ↑ marker for diagnosis and associated with low overall survival. |
| Jin et al., 2019 [27] | Advanced colorectal cancer | OXA, 5-FU, (and leucovorin) | Serum | miR-21-5p, miR-1246, miR-1229-5p and miR-96-5p | UC | qRT-PCR | ↑ chemoresistant patients versus chemosensitive patients. |
| Liu et al., 2019 [25] | Colorectal | OXA | Tumor tissue | miR-128-3p | None | qRT-PCR | ↓ relapse after therapy versus patients which responded well to neoadjuvant therapy. |
| Feng et al., 2019 [24] | DLBCL | R-CHOP regimen (cyclophosphamide, doxorubicin, vincristine, and prednisone, combined with the anti-CD20 monoclonal antibody–rituximab). | Serum | miR-99a-5p miR-125b-5p | ExoQuick | qRT-PCR | ↑ chemoresistant group vs chemosensitive group. |
| Xiao et al., 2019 [44] | DLBCL | Rituximab combined with Chemotherapy (R-CHOP regimen) | Serum | miR-451a | ExoQuick | qRT-PCR | ↓ miR-451a in cancer patients versus healthy subjects and may predict treatment efficacy. |
| Zare et al., 2019 [48] | DLBCL | R-CHOP regimen | Plasma | miR-155 | ExoSpin | qRT-PCR | ↑ miR-155 in relapsed patients or non-responsive to R-CHOP regimen. |
| Hu et al., 2019 [29] | Gastric | Non-specified | Ascites | miR-760, miR-6821-5p, miR-4745-5p, miR-200a-5p, miR-4741 and miR-320 | UC | RNA sequencing | ↑ Progressive disease |
| Yang et al., 2017 [28] | GBM | TMZ (only evaluated in vitro) | Serum and tumor tissue | miR-221 | ExoQuick | qRT-PCR | ↑ miR-221 may predict TMZ resistance. |
| Zeng et al., 2018 [45] | GBM | TMZ | Serum and CSF | miR-151a | UC | qRT-PCR | ↓ miR-151a prior to therapy was associated with poor TMZ response and poor prognosis (CSF fluid). |
| Fu et al., 2018 [31] | HCC | 5-FU, OXA, GEM, and Sorafenib | Tumor tissue | miR-32-5p | None | qRT-PCR | ↑ tumor tissue and associated with short overall survival and progression-free survival. |
| Wang et al., 2019 [17] | HCC | Sorafenib (in vitro use only) | Serum and tumor tissue | miR-744 | UC | qRT-PCR | ↓ miR-744 in HCC tissues and exosomes from serum of these patients. |
| Svedman et al., 2018 [42] | Metastatic BRAFV600 mutated CMM | MAPKis | Plasma | let-7g-5p and miR-497-5p | MiRCURY Exosome Isolation Kit | Microarray (MiRCURY) | ↑ let-7g-5p during treatment associated with improved disease control. ↑ miR-497-5p during treatment associated with prolonged progression-free survival. |
| Zhang et al., 2016 [46] | Multiple myeloma | Bz, Thalidomide and lenalidomide | Serum | miR-16-5p, miR-15a-5p, miR-20a-5p, and miR-17-5p | UC | Microarray (MiRCURY) | ↓ miR-16-5p, miR-15a-5p, miR-20a-5p, and miR-17-5p in patients resistant to Bz. |
| Ma et al., 2019 [1] | NSCLC | Cisplatin | Serum | miR-425-3p | ExoQuick | qRT-PCR | ↑ platinum-resistant cancer patients. |
| Wei et al., 2017 [43] | NSCLC | GEM | Plasma | miR-222-3p | UC | qRT-PCR | ↑ miR-222-3p associated with low response towards chemotherapy and progressive disease. |
| Yuwen et al., 2019 [23] | NSCLC | Cisplatin | Serum | miR-425-3p, miR-1273h, miR-4755-5p, miR-9-5p, miR-146a-5p, and miR-215-5p | ExoQuick | RNA sequencing and qRT-PCR | ↑ miR-425-3p associated with low response and poor progression-free survival. |
| Yuwen et al., 2017 [15] | NSCLC | Cisplatin | Serum | miR-146a-5p | ExoQuick | RNA sequencing and qRT-PCR | ↓ miR-146a-5p associated with shorter progression-free survival. |
| Alharbi et al., 2020 [33] | Ovarian | Not Indicated | Plasma | miR-891-5p | UC | qRT-PCR | ↑ relapse |
| Kanlikilicer et al., 2018 [32] | Ovarian | Paclitaxel (in vitro and in vivo use only) | Tumor tissue and TCGA tissue samples | miR-1246 | None | qRT-PCR | ↑ Associated with worse overall prognosis. |
| Kuhlmann et al., 2019 [5] | Ovarian | Carboplatin and Paclitaxel | Plasma | miR-181a, miR-1908, miR-21, miR-486 and miR-223 | ExoQuick | RNA sequencing | ↑ platinum-resistant cancer patients (preliminary study). |
| Pink et al., 2015 [34] | Ovarian | Cisplatin (in vitro use only) | Tumor tissue | miR-21-3p | None | GEO repository data and TCGA data | ↑ tumor tissue associated with shorter progression-free interval. |
| Yeung et al., 2016 [35] | Ovarian | Paclitaxel (in vitro and in vivo use only) | Primary culture derived from ovarian tissue | miR-21 | UC | Next generation sequencing | miR-21 can ↓ paclitaxel sensitivity in vitro and in vivo. |
| Zhu et al., 2019 [36] | Ovarian | Taxol and Cisplatin | Serum and tumor tissue | miR-223 | ExoQuick | qRT-PCR | ↓ miR-223 associated with longer progression-free survival and decreased cancer relapse. |
| Mikamori et al., 2017 [41] | Pancreatic ductal adenocarcinoma | GEM | Plasma | miR-155 | ExoQuick | qRT-PCR | ↑ associated with low disease-free survival. |
| Corcoran et al., 2014 [30] | Prostate | Docetaxel | Tumor and urine | miR-34a | None | Gene expression dataset (Omnibus) | ↓ cancer tissue vs benign and recurrent cancer vs no recurrence. |
| Huang et al., 2015 [14] | Prostate | Androgen-deprivation therapy (Docetaxel, abiraterone acetate, prednisone, cabazitaxel, and/or mitoxantrone. | Plasma | miR-1290, miR-1246, and miR-375 | ExoQuick | RNA Sequencing | ↑ Associated with poor overall survival. |
| He et al., 2020 [22] | Renal cell carcinoma | Sorafenib | Plasma | miR-31-5p | UC and sucrose cushion | qRT-PCR | ↑ Progressive disease during chemotherapy. |
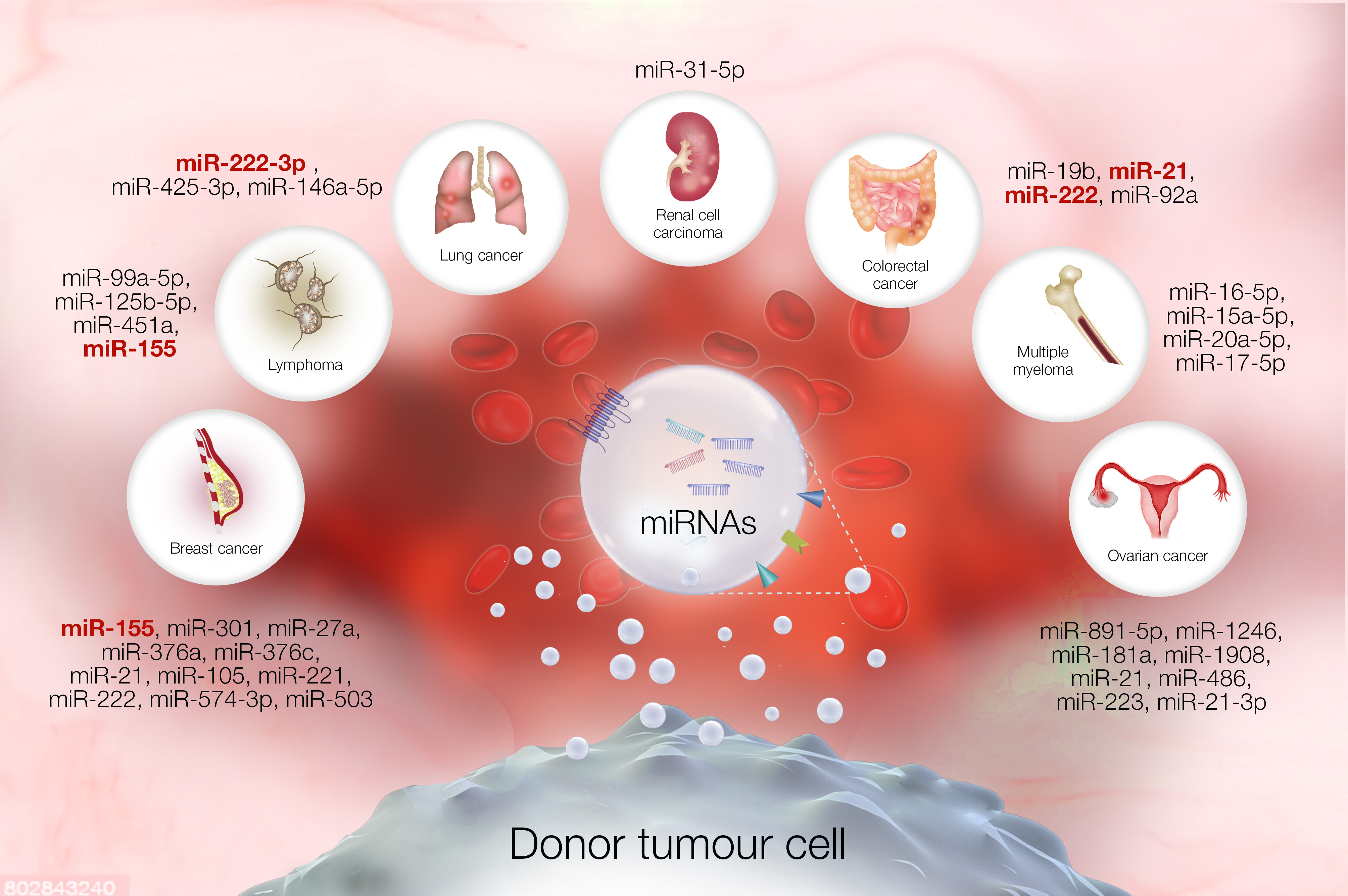
| Tumor Type | Exosomal miRNAs | References |
|---|---|---|
| Colorectal | miR-19b, miR-21, miR-222 and miR-92a | [25,26,27] |
| Ovarian | miR-891-5p, miR-1246, miR-181a, miR-1908, miR-21, miR-486, miR-223, miR-21-3p | [5,32,33,34,35,36] |
| Breast | miR-155, miR-301, mi-27a, miR-376a, miR-376c, miR-21, miR-105, miR-221, miR-222, miR-574-3p, miR-503 | [37,38,39,40] |
| Renal cell carcinoma | miR-31-5p | [22] |
| Lung | miR-222-3p, miR-425-3p, miR-146a-5p | [1,15,23,43] |
| Multiple myeloma | miR-16-5p, miR-15a-5p, miR-20a-5p and miR-17-5p | [46] |
| Lymphoma | miR-99a-5p, miR-125b-5p, miR-451a, miR-155 | [24,44,48] |
| Prostate | miR-34a, miR-1290, miR-1246, miR-375 | [14,30] |
| Hepatocellular carcinoma | miR-32-5p, miR-744 | [17,31] |
| Gastric | miR-760, miR-6821-5p, mi-4745-5p, miR-200a-5p, miR-4741, miR-320 | [29] |
| Pancreatic ductal adenocarcinoma | miR-155 | [41] |
| Metastatic BRAFV600 mutated cutaneous malignant melanoma | Let-7g-5p, miR-497-5p | [42] |
| Glioblastoma | miR-221, miR-151a | [28,45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campos, A.; Sharma, S.; Obermair, A.; Salomon, C. Extracellular Vesicle-Associated miRNAs and Chemoresistance: A Systematic Review. Cancers 2021, 13, 4608. https://doi.org/10.3390/cancers13184608
Campos A, Sharma S, Obermair A, Salomon C. Extracellular Vesicle-Associated miRNAs and Chemoresistance: A Systematic Review. Cancers. 2021; 13(18):4608. https://doi.org/10.3390/cancers13184608
Chicago/Turabian StyleCampos, America, Shayna Sharma, Andreas Obermair, and Carlos Salomon. 2021. "Extracellular Vesicle-Associated miRNAs and Chemoresistance: A Systematic Review" Cancers 13, no. 18: 4608. https://doi.org/10.3390/cancers13184608
APA StyleCampos, A., Sharma, S., Obermair, A., & Salomon, C. (2021). Extracellular Vesicle-Associated miRNAs and Chemoresistance: A Systematic Review. Cancers, 13(18), 4608. https://doi.org/10.3390/cancers13184608