
Neuronal Differentiation-Related Epigenetic Regulator ZRF1 Has Independent Prognostic Value in Neuroblastoma but Is Functionally Dispensable In Vitro

 , , , , and
, , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Analysis of Neuroblastoma Gene Expression Datasets
2.2. Cell Lines
2.3. Western Blot Analysis
2.4. Proliferation and Colony Formation Assays
2.5. Migration and Invasion Assays
2.6. Differentiation Assays
2.7. Vectors and Lentiviral Infection
2.8. ZRF1 Overexpression Experiments
2.9. ZRF1 Knockdown Experiments
3. Results
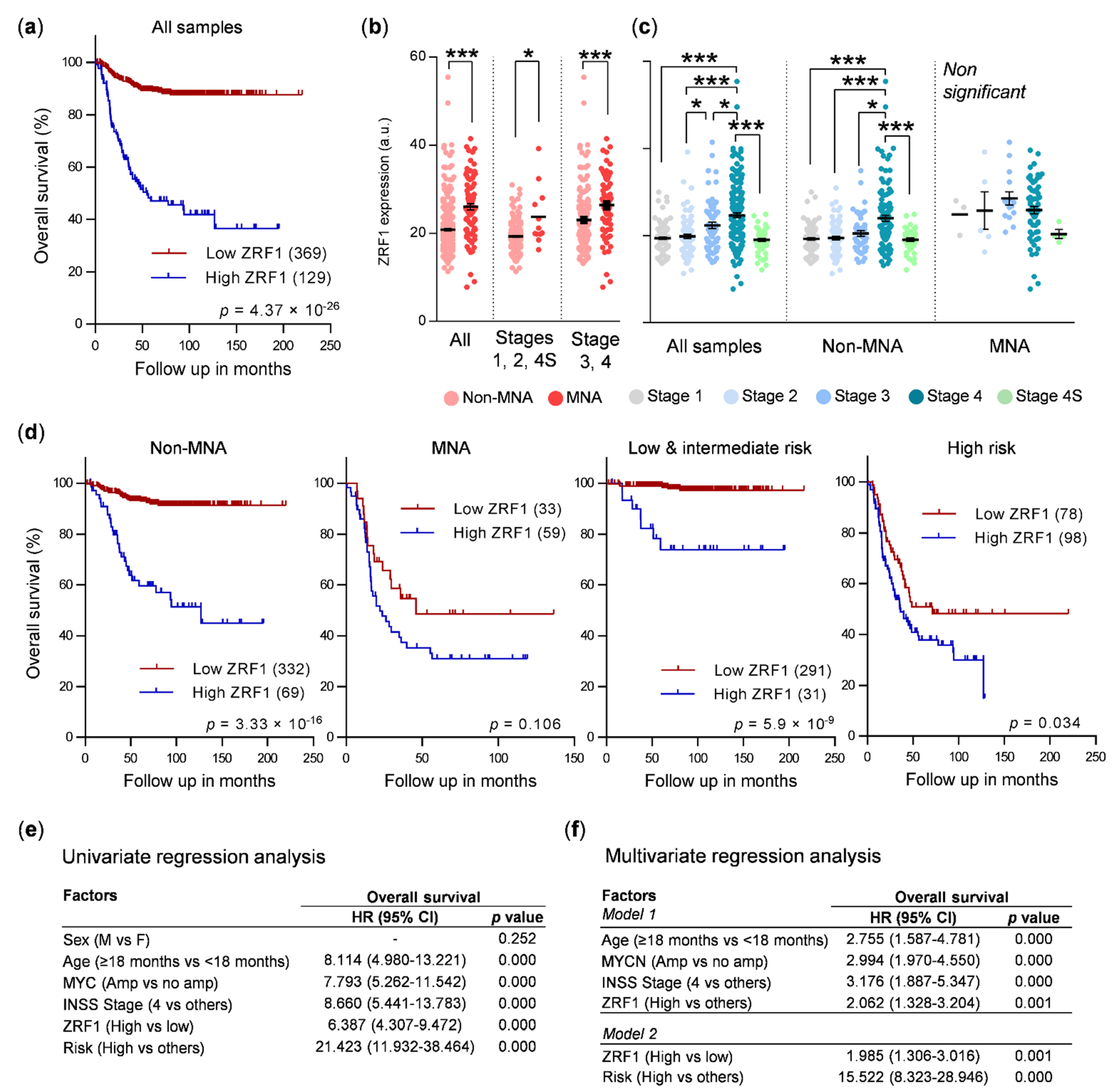
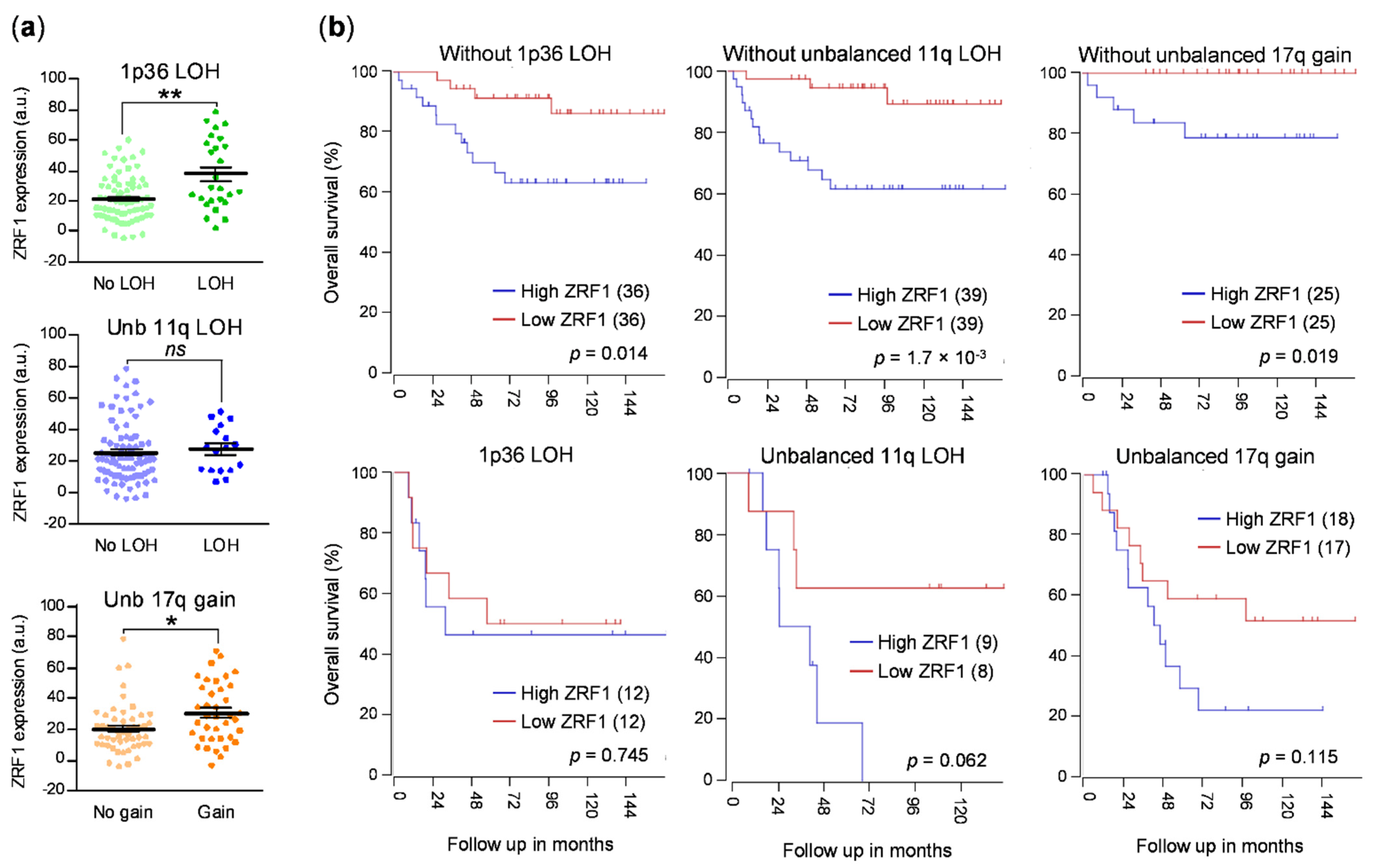
3.1. ZRF1 Is an Independent Prognostic Factor in Neuroblastoma
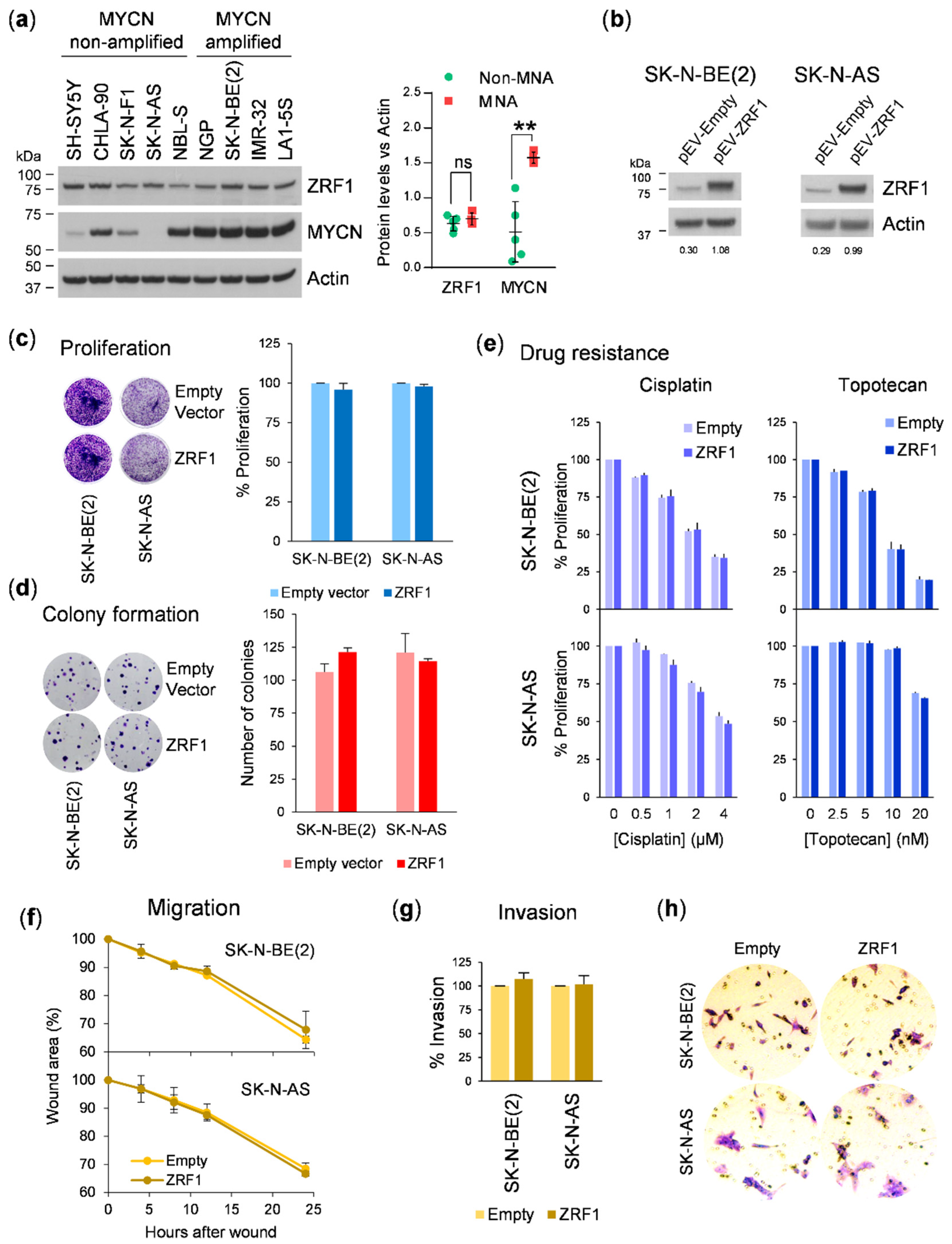
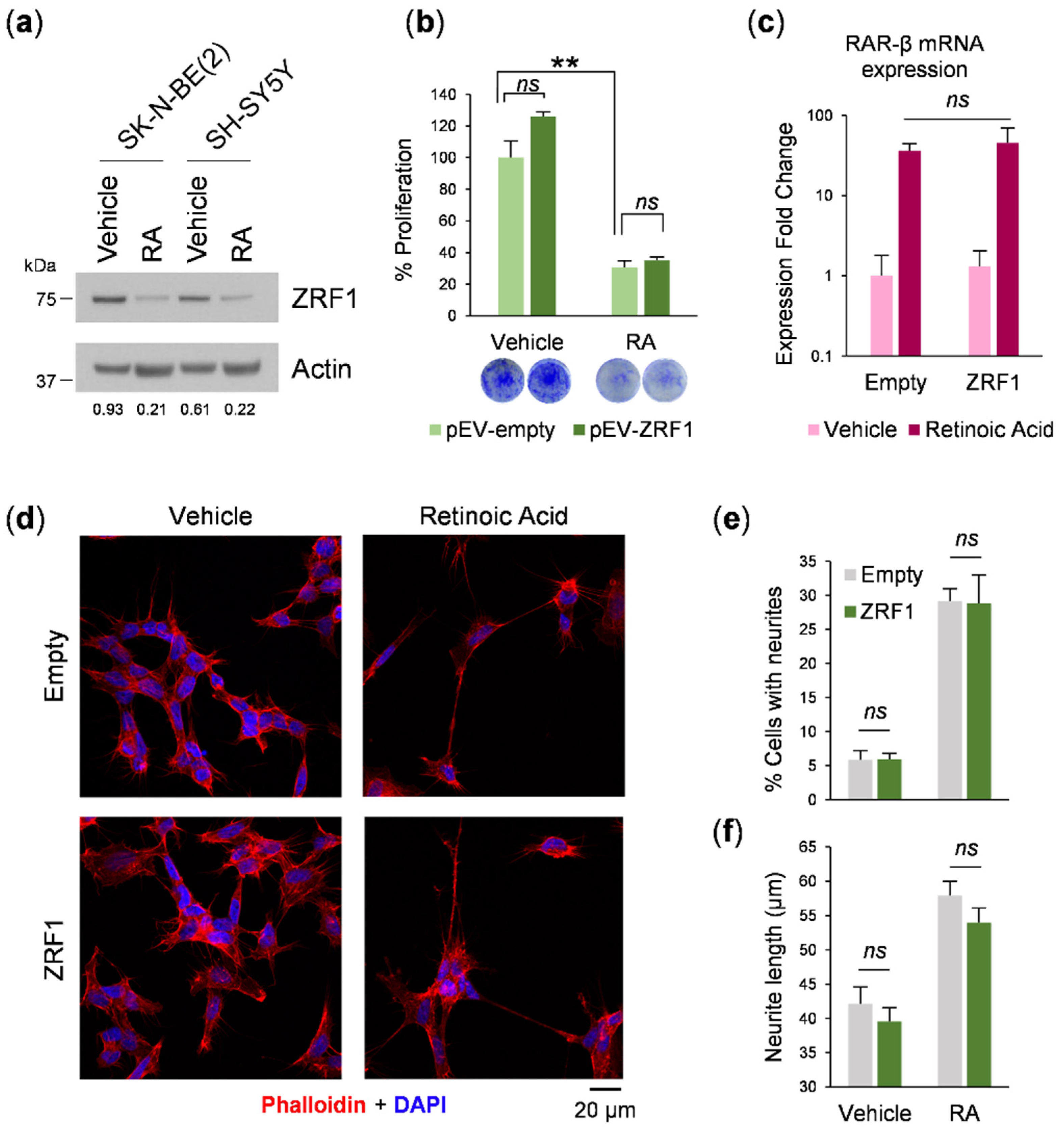
3.2. ZRF1 Is Not Sufficient to Enhance Neuroblastoma Aggressiveness
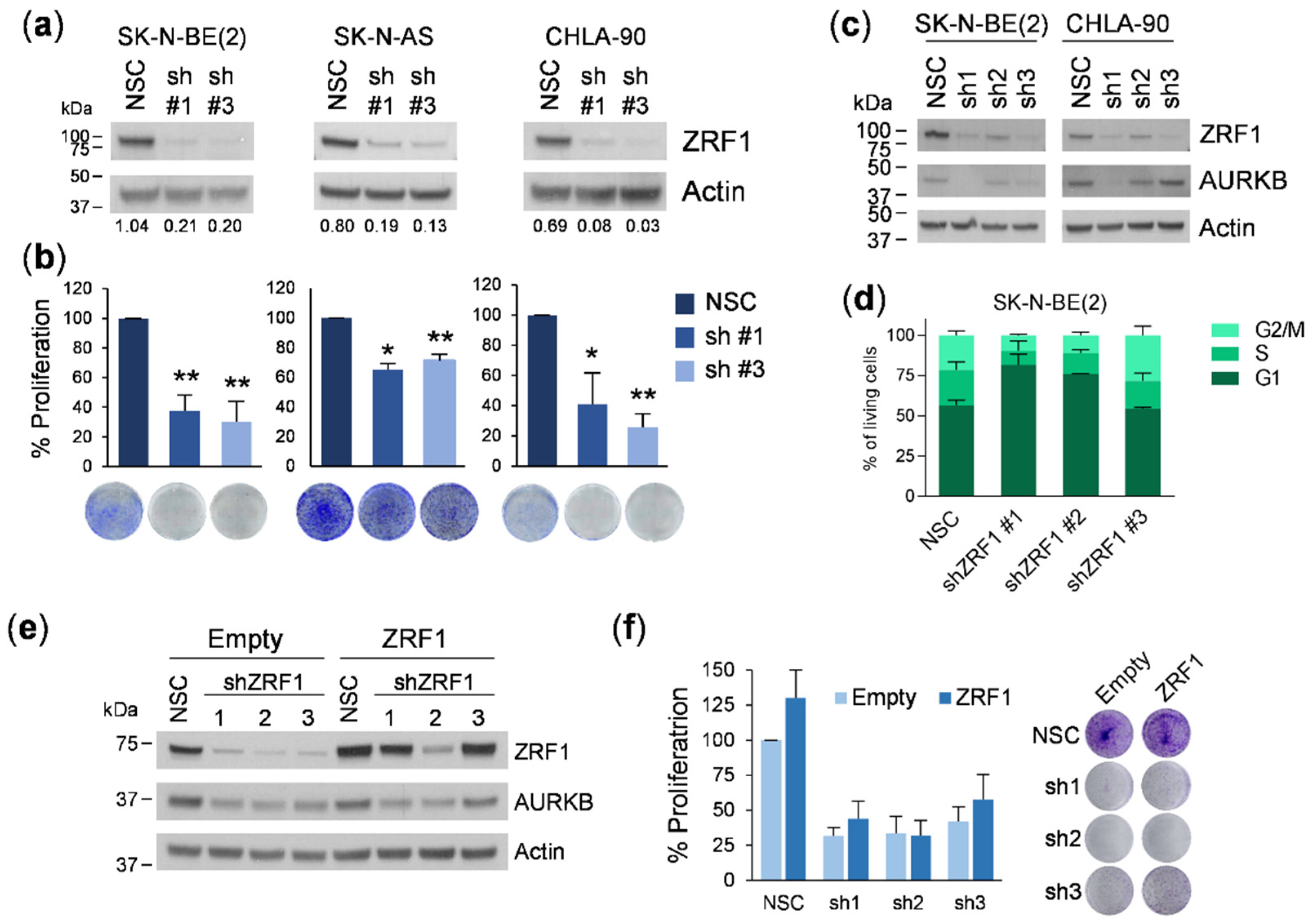
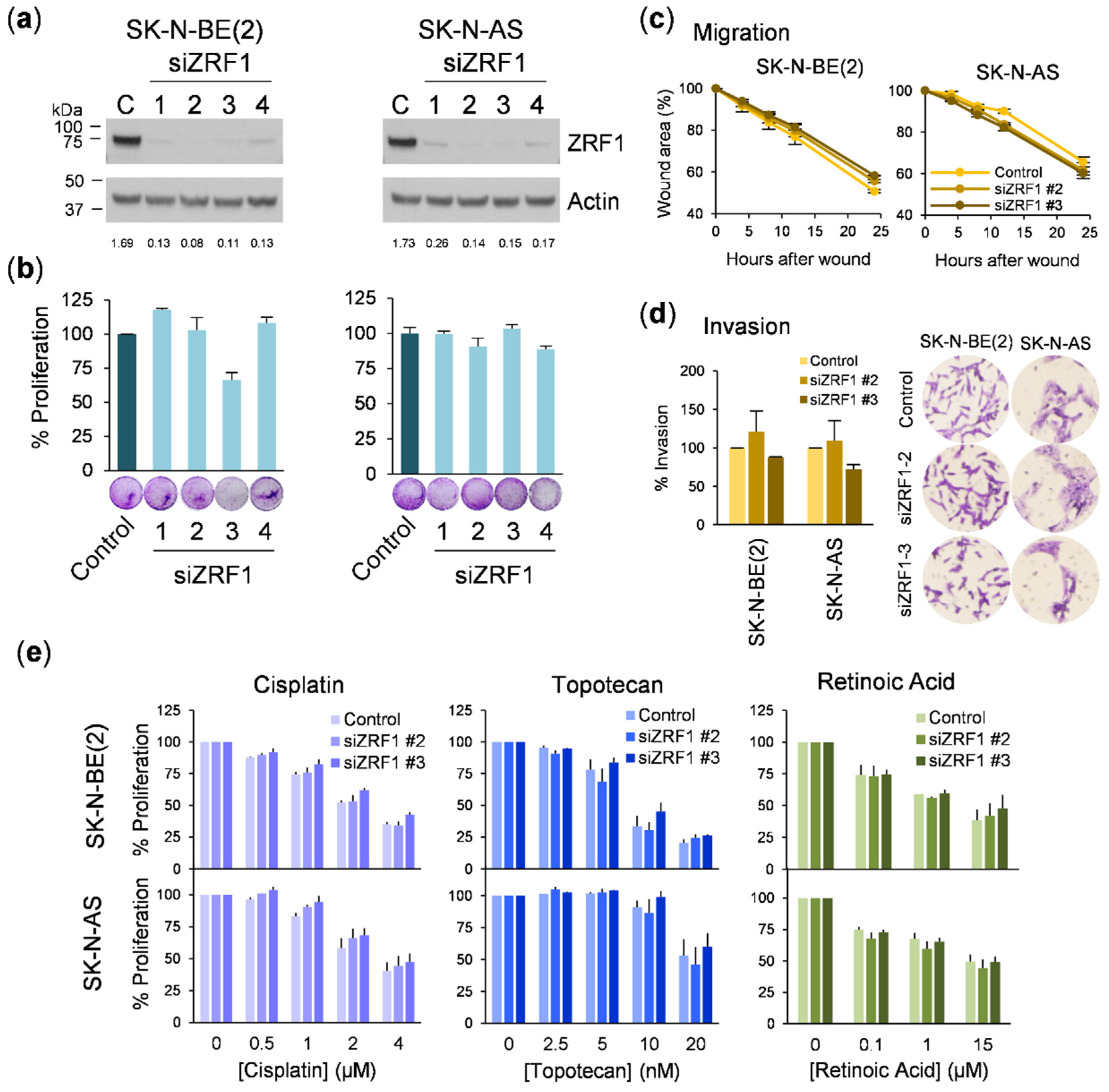
3.3. ZRF1 Knockdown Does Not Impair Neuroblastoma Proliferation and Reveals Inconsistencies between Different Gene Silencing Methodologies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maris, J.M. Recent Advances in Neuroblastoma. N. Engl. J. Med. 2010, 362, 2202–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, N.-L.V.; Dyer, M.A. Neuroblastoma: Developmental biology, cancer genomics and immunotherapy. Nat. Rev. Cancer 2013, 13, 397–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthay, K.K.; Maris, J.M.; Schleiermacher, G.; Nakagawara, A.; Mackall, C.L.; Diller, L.; Weiss, W.A. Neuroblastoma. Nat. Rev. Dis. Prim. 2016, 2, 16078. [Google Scholar] [CrossRef] [PubMed]
- Pudela, C.; Balyasny, S.; Applebaum, M.A. Nervous system: Embryonal tumors: Neuroblastoma. Atlas Genet. Cytogenet. Oncol. Haematol. 2020, 24, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Maris, J.M.; Hogarty, M.D.; Bagatell, R.; Cohn, S.L. Neuroblastoma. Lancet 2007, 369, 2106–2120. [Google Scholar] [CrossRef]
- Reynolds, C.P. Detection and treatment of minimal residual disease in high-risk neuroblastoma. Pediatr. Transplant. 2004, 8, 56–66. [Google Scholar] [CrossRef]
- Durinck, K.; Speleman, F. Epigenetic regulation of neuroblastoma development. Cell Tissue Res. 2018, 372, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Aloia, L.; Di Stefano, B.; Sessa, A.; Morey, L.; Santanach, A.; Gutierrez, A.; Cozzuto, L.; Benitah, S.A.; Graf, T.; Broccoli, V.; et al. ZRF1 is required to establish and maintain neural progenitor identity. Genes Dev. 2014, 28, 182–197. [Google Scholar] [CrossRef] [Green Version]
- Aloia, L.; Gutierrez, A.; Caballero, J.M.; Di Croce, L. Direct interaction between Id1 and ZRF1 controls neural differentiation of embryonic stem cells. EMBO Rep. 2015, 16, 63–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto-Taniura, N.; Pirollet, F.; Monroe, R.; Gerace, L.; Westendorf, J.M. Identification of novel M phase phosphoproteins by expression cloning. Mol. Biol. Cell 1996, 7, 1455–1469. [Google Scholar] [CrossRef] [Green Version]
- Hundley, H.A.; Walter, W.; Bairstow, S.; Craig, E.A. Human Mpp11 J protein: Ribosome-tethered molecular chaperons are ubiquitous. Science 2005, 308, 1032–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, H.; Conz, C.; Maier, P.; Wölfle, T.; Suzuki, C.K.; Jenö, P.; Rücknagel, P.; Stahl, J.; Rospert, S. The chaperones MPP11 and Hsp70L1 form the mammalian ribosome-associated complex. Proc. Natl. Acad. Sci. USA 2005, 102, 10064–10069. [Google Scholar] [CrossRef] [Green Version]
- Richly, H.; Rocha-Viegas, L.; Ribeiro, J.D.; Demajo, S.; Gundem, G.; Lopez-Bigas, N.; Nakagawa, T.; Rospert, S.; Ito, T.; Di Croce, L. Transcriptional activation of polycomb-repressed genes by ZRF1. Nature 2010, 468, 1124–1130. [Google Scholar] [CrossRef] [PubMed]
- Rath, S.K.; Deb, M.; Sengupta, D.; Kari, V.; Kar, S.; Parbin, S.; Pradhan, N.; Patra, S.K. Silencing of ZRF1 impedes survival of estrogen receptor positive MCF-7 cells and potentiates the effect of curcumin. Tumor Biol. 2016, 37, 12535–12546. [Google Scholar] [CrossRef] [PubMed]
- Imamura, T.; Komatsu, S.; Ichikawa, D.; Miyamae, M.; Okajima, W.; Ohashi, T.; Kiuchi, J.; Nishibeppu, K.; Kosuga, T.; Konishi, H.; et al. Overexpression of ZRF1 is related to tumor malignant potential and a poor outcome of gastric carcinoma. Carcinogenesis 2018, 39, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Demajo, S.; Uribesalgo, I.; Gutiérrez, A.; Ballaré, C.; Capdevila, S.; Roth, M.; Zuber, J.; Martín-Caballero, J.; Di Croce, L. ZRF1 controls the retinoic acid pathway and regulates leukemogenic potential in acute myeloid leukemia. Oncogene 2014, 33, 5501–5510. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Naldini, L.; Blömer, U.; Gallay, P.; Ory, D.; Mulligan, R.; Gage, F.H.; Verma, I.M.; Trono, D.; Naldini, L.; Blomer, U.; et al. In Vivo Gene Delivery and Stable Transduction of Nondividing Cells by a Lentiviral Vector. Science 1996, 272, 263–267. [Google Scholar] [CrossRef] [Green Version]
- Zufferey, R.; Dull, T.; Mandel, R.J.; Bukovsky, A.; Quiroz, D.; Naldini, L.; Trono, D. Self-Inactivating Lentivirus Vector for Safe and Efficient In Vivo Gene Delivery. J. Virol. 1998, 72, 9873–9880. [Google Scholar] [CrossRef] [Green Version]
- Caron, H. Allelic loss of chromosome 1 and additional chromosome 17 material are both unfavourable prognostic markers in neuroblastoma. Med. Pediatr. Oncol. 1995, 24, 215–221. [Google Scholar] [CrossRef]
- Attiyeh, E.F.; London, W.B.; Mossé, Y.P.; Wang, Q.; Winter, C.; Khazi, D.; McGrady, P.W.; Seeger, R.C.; Look, A.T.; Shimada, H.; et al. Chromosome 1p and 11q Deletions and Outcome in Neuroblastoma. N. Engl. J. Med. 2005, 353, 2243–2253. [Google Scholar] [CrossRef]
- Reynolds, C.P.; Lemons, R.S. Retinoid therapy of childhood cancer. Hematol. Oncol. Clin. N. Am. 2001, 15, 867–910. [Google Scholar] [CrossRef]
- He, S.; Liu, Z.; Oh, D.Y.; Thiele, C.J. MYCN and the epigenome. Front. Oncol. 2013, 3, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnsen, J.I.; Dyberg, C.; Wickström, M. Neuroblastoma—A neural crest derived embryonal malignancy. Front. Mol. Neurosci. 2019, 12, 9. [Google Scholar] [CrossRef] [PubMed]
- Weiss, W.A.; Aldape, K.; Mohapatra, G.; Feuerstein, B.G.; Bishop, J.M. Targeted expression of MYCN causes neuroblastoma in transgenic mice. EMBO J. 1997, 16, 2985–2995. [Google Scholar] [CrossRef] [PubMed]
- Heukamp, L.C.; Thor, T.; Schramm, A.; De Preter, K.; Kumps, C.; De Wilde, B.; Odersky, A.; Peifer, M.; Lindner, S.; Spruessel, A.; et al. Targeted expression of mutated ALK induces neuroblastoma in transgenic mice. Sci. Transl. Med. 2012, 4, 141ra91. [Google Scholar] [CrossRef]
- Kamijo, T. Role of stemness-related molecules in neuroblastoma. Pediatr. Res. 2012, 71, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Nowak, K.; Kerl, K.; Fehr, D.; Kramps, C.; Gessner, C.; Killmer, K.; Samans, B.; Berwanger, B.; Christiansen, H.; Lutz, W. BMI1 is a target gene of E2F-1 and is strongly expressed in primary neuroblastomas. Nucleic Acids Res. 2006, 34, 1745–1754. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; Hu, B.; Li, T.; Ma, J.; Alam, G.; Gunning, W.T.; Ding, H.F. Bmi-1 is essential for the tumorigenicity of neuroblastoma cells. Am. J. Pathol. 2007, 170, 1370–1378. [Google Scholar] [CrossRef] [Green Version]
- Ochiai, H.; Takenobu, H.; Nakagawa, A.; Yamaguchi, Y.; Kimura, M.; Ohira, M.; Okimoto, Y.; Fujimura, Y.; Koseki, H.; Kohno, Y.; et al. Bmi1 is a MYCN target gene that regulates tumorigenesis through repression of KIF1BΒ and TSLC1 in neuroblastoma. Oncogene 2010, 29, 2681–2690. [Google Scholar] [CrossRef] [Green Version]
- Kaymak, A.; Sayols, S.; Papadopoulou, T.; Richly, H. Role for the transcriptional activator ZRF1 in early metastatic events in breast cancer progression and endocrine resistance. Oncotarget 2018, 9, 28666–28690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Esteban, R.; Jiang, X. Differential gene expression in disease: A comparison between high-throughput studies and the literature. BMC Med. Genom. 2017, 10, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez, C.; Antonelli, R.; Masanas, M.; Soriano, A.; Devis-Jauregui, L.; Camacho, J.; Magdaleno, A.; Guillén, G.; Hladun, R.; Jubierre, L.; et al. Neuronal Differentiation-Related Epigenetic Regulator ZRF1 Has Independent Prognostic Value in Neuroblastoma but Is Functionally Dispensable In Vitro. Cancers 2021, 13, 4845. https://doi.org/10.3390/cancers13194845
Jiménez C, Antonelli R, Masanas M, Soriano A, Devis-Jauregui L, Camacho J, Magdaleno A, Guillén G, Hladun R, Jubierre L, et al. Neuronal Differentiation-Related Epigenetic Regulator ZRF1 Has Independent Prognostic Value in Neuroblastoma but Is Functionally Dispensable In Vitro. Cancers. 2021; 13(19):4845. https://doi.org/10.3390/cancers13194845
Chicago/Turabian StyleJiménez, Carlos, Roberta Antonelli, Marc Masanas, Aroa Soriano, Laura Devis-Jauregui, Jessica Camacho, Ainara Magdaleno, Gabriela Guillén, Raquel Hladun, Luz Jubierre, and et al. 2021. "Neuronal Differentiation-Related Epigenetic Regulator ZRF1 Has Independent Prognostic Value in Neuroblastoma but Is Functionally Dispensable In Vitro" Cancers 13, no. 19: 4845. https://doi.org/10.3390/cancers13194845
APA StyleJiménez, C., Antonelli, R., Masanas, M., Soriano, A., Devis-Jauregui, L., Camacho, J., Magdaleno, A., Guillén, G., Hladun, R., Jubierre, L., Roma, J., Llobet-Navas, D., Sánchez de Toledo, J., Moreno, L., Gallego, S., & Segura, M. F. (2021). Neuronal Differentiation-Related Epigenetic Regulator ZRF1 Has Independent Prognostic Value in Neuroblastoma but Is Functionally Dispensable In Vitro. Cancers, 13(19), 4845. https://doi.org/10.3390/cancers13194845