
SSTR2 in Nasopharyngeal Carcinoma: Relationship with Latent EBV Infection and Potential as a Therapeutic Target

 , , ,
, , , 
 , , , , , ,
, , , , , ,  and
and 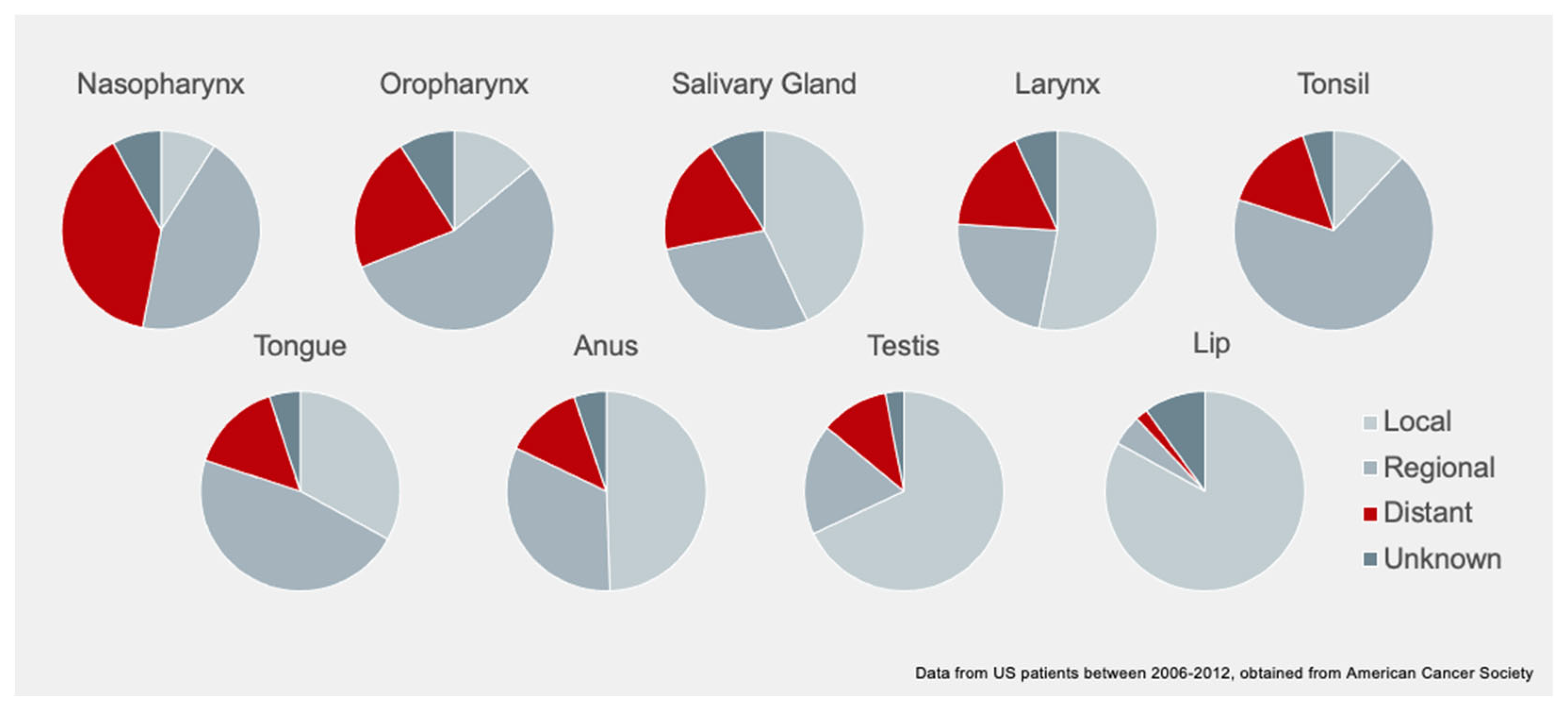{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Non-Viral Risk Factors
2.1. Environmental Risk Factors
2.2. Host Genetic Susceptibility
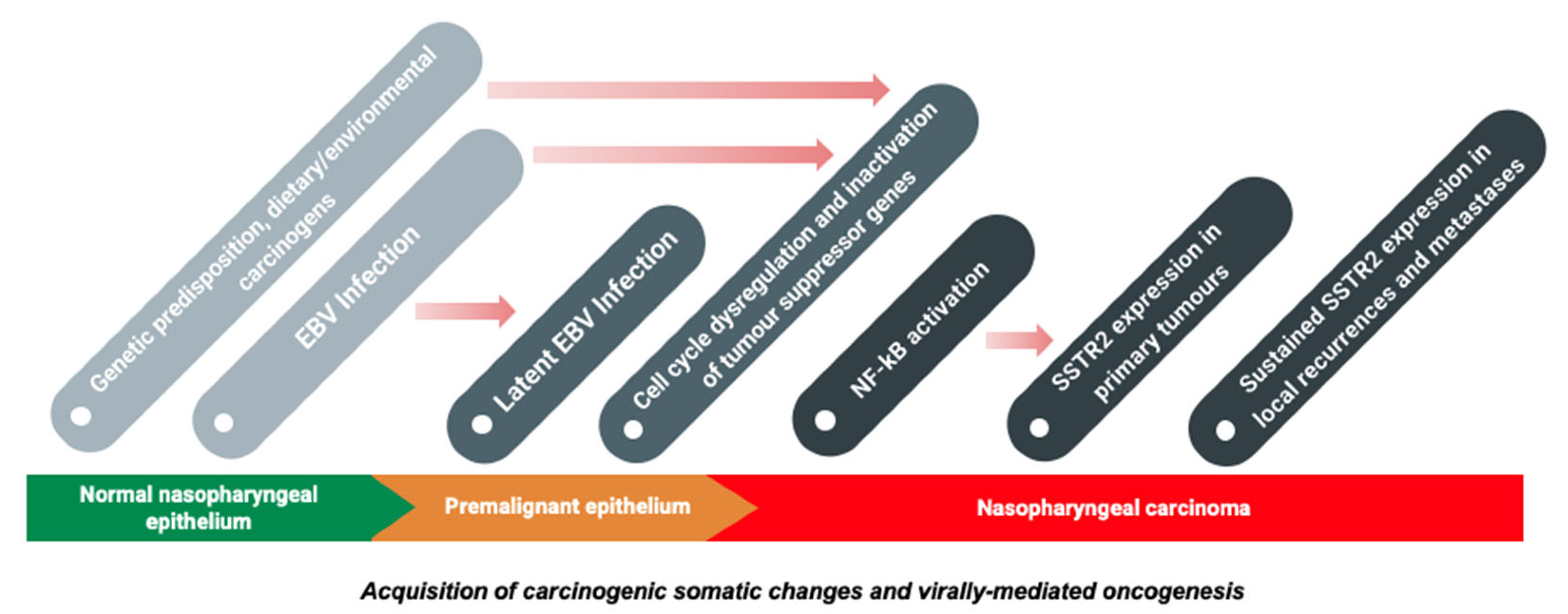
3. Molecular Mechanisms of Oncogenesis
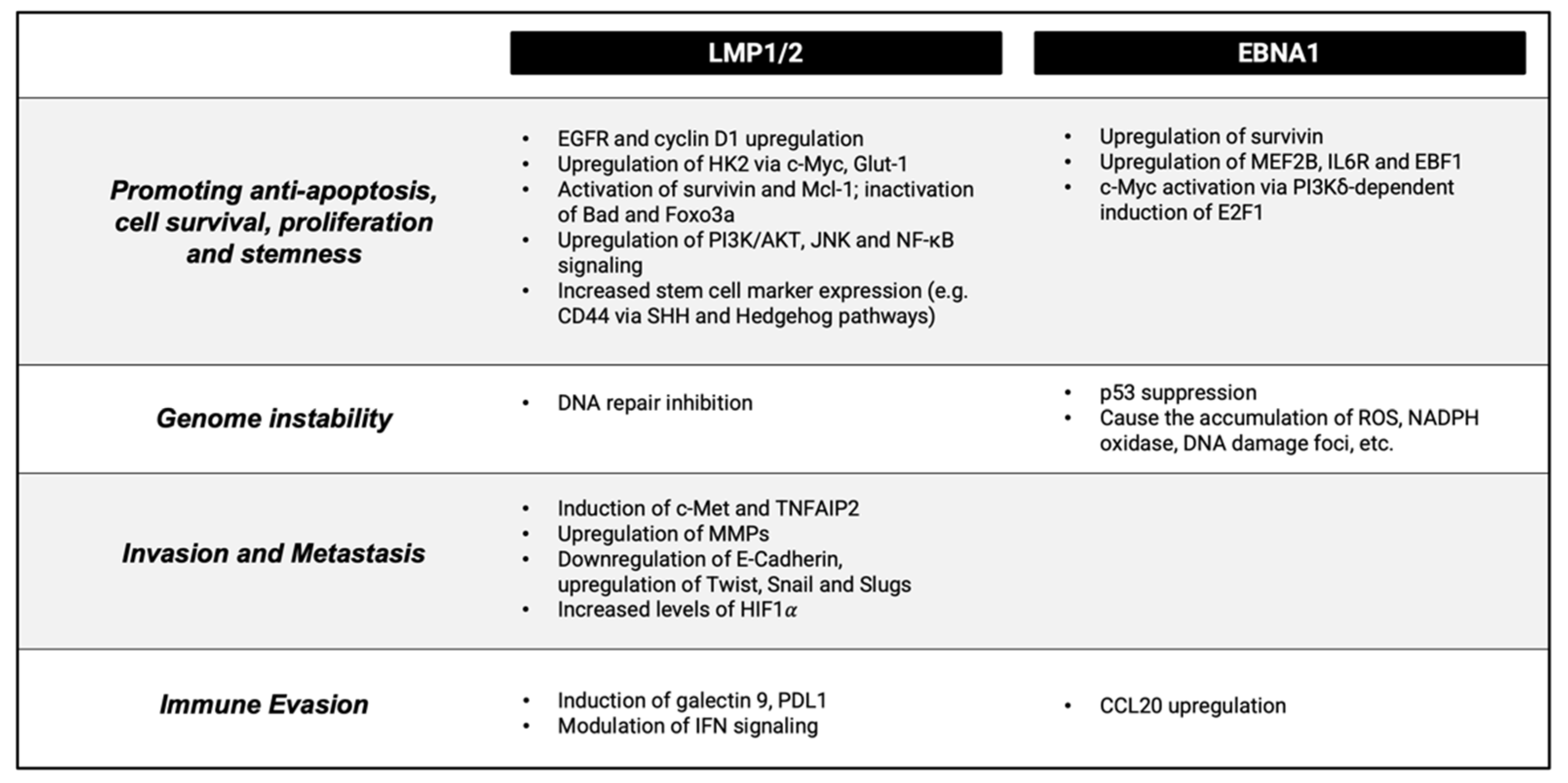
3.1. Virally-Mediated Oncogenesis
3.2. Contribution of Cellular Factors to Oncogenesis
4. SSTR2 and the Pathogenesis of Nasopharyngeal Carcinoma
4.1. Somatostatin Receptors in Cancer outside the Nasopharynx
4.2. SSTR2 and Nasopharyngeal Cancer
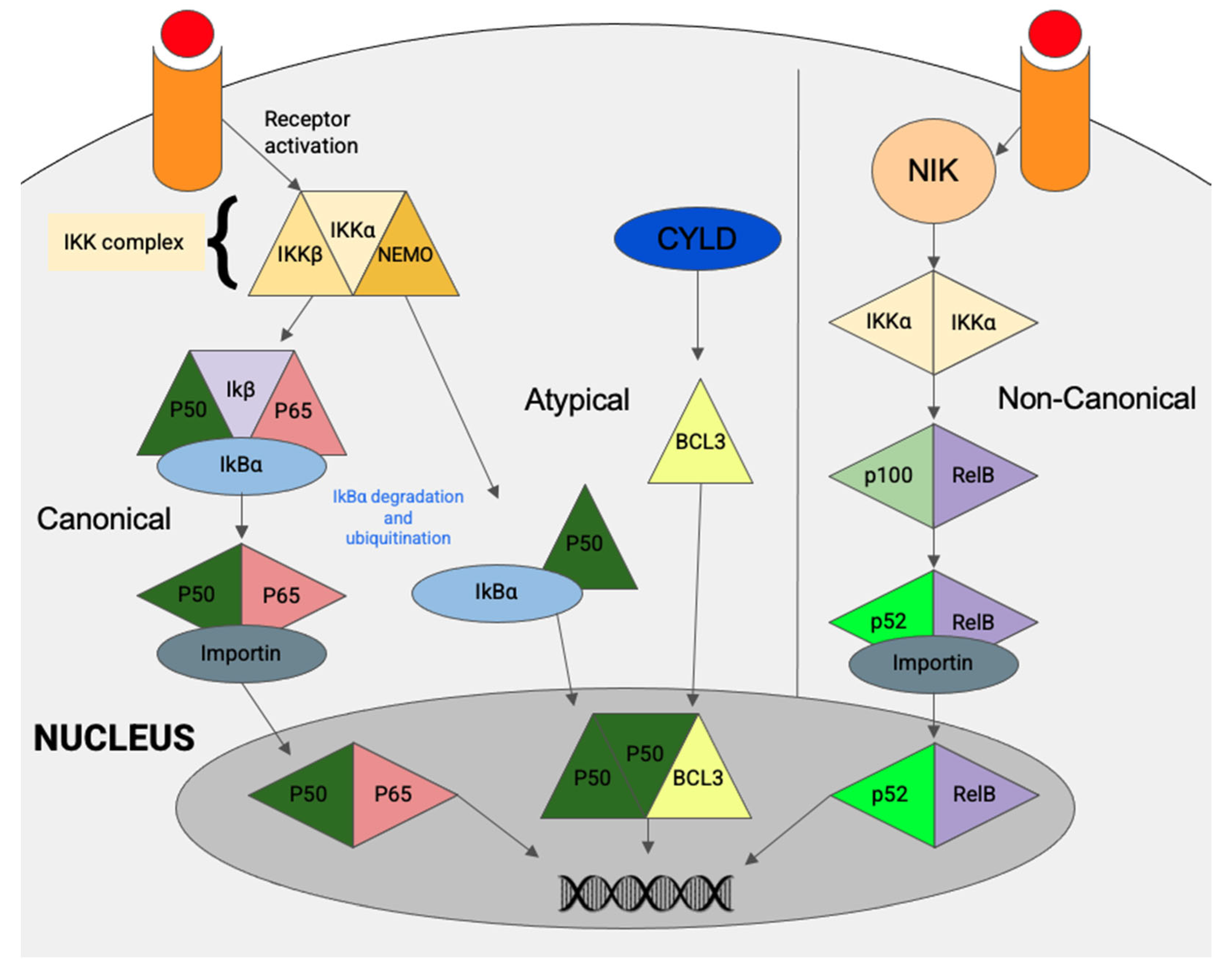
5. The NF-κB Pathway
6. The Role of SSTR2 in Imaging and Targeted Therapeutic Approaches
6.1. The 68Ga-Dotatate Imaging and SSTR2
6.2. Treatment Options
6.3. Radionuclide Therapy
6.4. Immunotherapy
6.5. SSTR2 Targeted Therapies
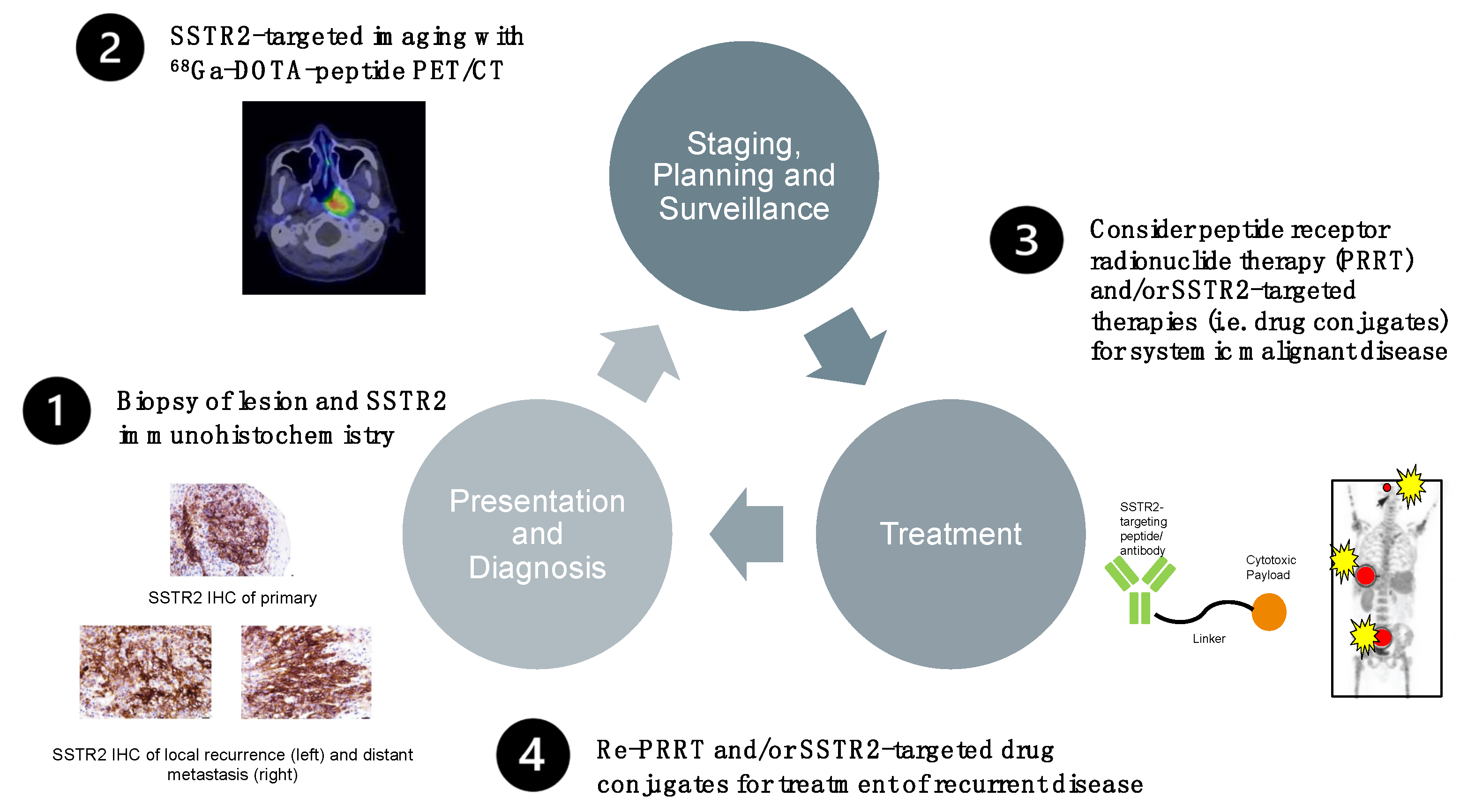
6.6. Prototype Protocol for a Future Management Option of Treatment-Refractory EBV-Positive NPC
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- American Cancer Society. Cancer Facts and Figures 2017. Special Section: Rare Cancers in Adults. Available online: https://www.cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures/cancer-facts-figures-2017.html (accessed on 7 May 2021).
- Lee, A.W.; Ma, B.B.; Ng, W.T.; Chan, A.T. Management of Nasopharyngeal Carcinoma: Current Practice and Future Perspective. J. Clin. Oncol. 2015, 33, 3356–3364. [Google Scholar] [CrossRef] [PubMed]
- You, B.; Le Tourneau, C.; Chen, E.X.; Wang, L.; Jarvi, A.; Bharadwaj, R.R.; Kamel-Reid, S.; Perez-Ordonez, B.; Mann, V.; Siu, L.L. A Phase II trial of erlotinib as maintenance treatment after gemcitabine plus platinum-based chemotherapy in patients with recurrent and/or metastatic nasopharyngeal carcinoma. Am. J. Clin. Oncol. 2012, 35, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Au, K.H.; Ngan, R.K.C.; Ng, A.W.Y.; Poon, D.M.C.; Ng, W.T.; Yuen, K.T.; Lee, V.H.F.; Tung, S.Y.; Chan, A.T.C.; Sze, H.C.K.; et al. Treatment outcomes of nasopharyngeal carcinoma in modern era after intensity modulated radiotherapy (IMRT) in Hong Kong: A report of 3328 patients (HKNPCSG 1301 study). Oral Oncol. 2018, 77, 16–21. [Google Scholar] [CrossRef]
- Lechner, M.; Schartinger, V.H.; Steele, C.D.; Nei, W.L.; Ooft, M.L.; Schreiber, L.M.; Pipinikas, C.P.; Chung, G.T.; Chan, Y.Y.; Wu, F.; et al. Somatostatin receptor 2 expression in nasopharyngeal cancer is induced by Epstein Barr virus infection: Impact on prognosis, imaging and therapy. Nat. Commun. 2021, 12, 117. [Google Scholar] [CrossRef]
- Reichlin, S. Somatostatin: Historical aspects. Scand. J. Gastroenterol. Suppl. 1986, 21, 1–10. [Google Scholar] [CrossRef]
- Mascardo, R.N.; Sherline, P. Somatostatin inhibits rapid centrosomal separation and cell proliferation induced by epidermal growth factor. Endocrinology 1982, 111, 1394–1396. [Google Scholar] [CrossRef]
- Viswanathan, K.; Sadow, P.M. Somatostatin receptor 2 (SSTR2) is highly sensitive and specific for Epstein-Barr Virus-associated nasopharyngeal carcinoma. Hum. Pathol. 2021. [Google Scholar] [CrossRef]
- Tsao, S.W.; Tsang, C.M.; Lo, K.W. Epstein-Barr virus infection and nasopharyngeal carcinoma. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef]
- Mimi, C.Y.; Ho, J.H.; Lai, S.H.; Henderson, B.E. Cantonese-style salted fish as a cause of nasopharyngeal carcinoma: Report of a case-control study in Hong Kong. Cancer Res. 1986, 46, 956–961. [Google Scholar]
- Cao, S.M.; Simons, M.J.; Qian, C.N. The prevalence and prevention of nasopharyngeal carcinoma in China. Chin. J. Cancer 2011, 30, 114–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, E.T.; Ye, W.; Zeng, Y.X.; Adami, H.O. The Evolving Epidemiology of Nasopharyngeal Carcinoma. Cancer Epidemiol. Biomark. Prev. 2021, 30, 1035–1047. [Google Scholar] [CrossRef]
- Hildesheim, A.; Dosemeci, M.; Chan, C.C.; Chen, C.J.; Cheng, Y.J.; Hsu, M.M.; Chen, I.H.; Mittl, B.F.; Sun, B.; Levine, P.H.; et al. Occupational exposure to wood, formaldehyde, and solvents and risk of nasopharyngeal carcinoma. Cancer Epidemiol. Biomark. Prev. 2001, 10, 1145–1153. [Google Scholar]
- Feng, B.J.; Khyatti, M.; Ben-Ayoub, W.; Dahmoul, S.; Ayad, M.; Maachi, F.; Bedadra, W.; Abdoun, M.; Mesli, S.; Bakkali, H.; et al. Cannabis, tobacco and domestic fumes intake are associated with nasopharyngeal carcinoma in North Africa. Br. J. Cancer 2009, 101, 1207–1212. [Google Scholar] [CrossRef] [Green Version]
- Abdulamir, A.S.; Hafidh, R.R.; Abdulmuhaimen, N.; Abubakar, F.; Abbas, K.A. The distinctive profile of risk factors of nasopharyngeal carcinoma in comparison with other head and neck cancer types. BMC Public Health 2008, 8, 400. [Google Scholar] [CrossRef] [Green Version]
- Feng, B.J.; Huang, W.; Shugart, Y.Y.; Lee, M.K.; Zhang, F.; Xia, J.C.; Wang, H.Y.; Huang, T.B.; Jian, S.W.; Huang, P.; et al. Genome-wide scan for familial nasopharyngeal carcinoma reveals evidence of linkage to chromosome 4. Nat. Genet. 2002, 31, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Zeng, Z.Y.; Xia, J.H.; Xia, K.; Shen, S.R.; Li, X.L.; Hu, D.X.; Tan, C.; Xiang, J.J.; Zhou, J.; et al. A susceptibility locus at chromosome 3p21 linked to familial nasopharyngeal carcinoma. Cancer Res. 2004, 64, 1972–1974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.F.; Qiu, Q.H.; Fu, S.M.; Sun, D.; Magnusson, K.; He, B.; Lindblom, A.; Ernberg, I. A genome-wide scan suggests a susceptibility locus on 5p 13 for nasopharyngeal carcinoma. Eur. J. Hum. Genet. 2008, 16, 343–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tse, K.P.; Su, W.H.; Chang, K.P.; Tsang, N.M.; Yu, C.J.; Tang, P.; See, L.C.; Hsueh, C.; Yang, M.L.; Hao, S.P.; et al. Genome-wide association study reveals multiple nasopharyngeal carcinoma-associated loci within the HLA region at chromosome 6p21.3. Am. J. Hum. Genet. 2009, 85, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Bei, J.X.; Li, Y.; Jia, W.H.; Feng, B.J.; Zhou, G.; Chen, L.Z.; Feng, Q.S.; Low, H.Q.; Zhang, H.; He, F.; et al. A genome-wide association study of nasopharyngeal carcinoma identifies three new susceptibility loci. Nat. Genet. 2010, 42, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Chin, Y.M.; Mushiroda, T.; Takahashi, A.; Kubo, M.; Krishnan, G.; Yap, L.F.; Teo, S.H.; Lim, P.V.; Yap, Y.Y.; Pua, K.C.; et al. HLA-A SNPs and amino acid variants are associated with nasopharyngeal carcinoma in Malaysian Chinese. Int. J. Cancer 2015, 136, 678–687. [Google Scholar] [CrossRef]
- Ning, L.; Ko, J.M.; Yu, V.Z.; Ng, H.Y.; Chan, C.K.; Tao, L.; Lam, S.Y.; Leong, M.M.; Ngan, R.K.; Kwong, D.L.; et al. Nasopharyngeal carcinoma MHC region deep sequencing identifies HLA and novel non-HLA TRIM31 and TRIM39 loci. Commun. Biol. 2020, 3, 759. [Google Scholar] [CrossRef]
- He, J.F.; Jia, W.H.; Fan, Q.; Zhou, X.X.; Qin, H.D.; Shugart, Y.Y.; Zeng, Y.X. Genetic polymorphisms of TLR3 are associated with Nasopharyngeal carcinoma risk in Cantonese population. BMC Cancer 2007, 7, 194. [Google Scholar] [CrossRef] [Green Version]
- Catarino, R.J.; Breda, E.; Coelho, V.; Pinto, D.; Sousa, H.; Lopes, C.; Medeiros, R. Association of the A870G cyclin D1 gene polymorphism with genetic susceptibility to nasopharyngeal carcinoma. Head Neck 2006, 28, 603–608. [Google Scholar] [CrossRef]
- Lee, M.H.; Huang, Y.H.; Coghill, A.E.; Liu, Z.; Yu, K.J.; Hsu, W.L.; Chien, Y.C.; Wang, C.P.; Chen, T.C.; Chen, C.J.; et al. Epstein-Barr Virus-Based Nasopharyngeal Carcinoma (NPC) Risk Prediction Scores Are Elevated in NPC Multiplex Family Members in Taiwan. J. Infect. Dis. 2021, 223, 441–444. [Google Scholar] [CrossRef]
- Liu, Z.; Derkach, A.; Yu, K.J.; Yeager, M.; Chang, Y.S.; Chen, C.J.; Gyllensten, U.; Lan, Q.; Lee, M.H.; McKay, J.D.; et al. Patterns of Human Leukocyte Antigen Class I and Class II Associations and Cancer. Cancer Res. 2021, 81, 1148–1152. [Google Scholar] [CrossRef]
- Yu, G.; Hsu, W.L.; Coghill, A.E.; Yu, K.J.; Wang, C.P.; Lou, P.J.; Liu, Z.; Jones, K.; Vogt, A.; Wang, M.; et al. Whole-Exome Sequencing of Nasopharyngeal Carcinoma Families Reveals Novel Variants Potentially Involved in Nasopharyngeal Carcinoma. Sci. Rep. 2019, 9, 9916. [Google Scholar] [CrossRef]
- Dai, W.; Zheng, H.; Cheung, A.K.; Tang, C.S.; Ko, J.M.; Wong, B.W.; Leong, M.M.; Sham, P.C.; Cheung, F.; Kwong, D.L.; et al. Whole-exome sequencing identifies MST1R as a genetic susceptibility gene in nasopharyngeal carcinoma. Proc. Natl. Acad. Sci. USA 2016, 113, 3317–3322. [Google Scholar] [CrossRef] [Green Version]
- Tsao, S.W.; Yip, Y.L.; Tsang, C.M.; Pang, P.S.; Lau, V.M.; Zhang, G.; Lo, K.W. Etiological factors of nasopharyngeal carcinoma. Oral Oncol. 2014, 50, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, W.; Zhang, W.; Wang, W.; Song, Y.; Xiao, H.; Luo, B. Constitutive activation of the canonical NF-kappaB signaling pathway in EBV-associated gastric carcinoma. Virology 2019, 532, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Frappier, L. Role of EBNA1 in NPC tumourigenesis. Semin. Cancer Biol. 2012, 22, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jia, L.; Tsang, C.M.; Tsao, S.W. EBV Infection and Glucose Metabolism in Nasopharyngeal Carcinoma. Adv. Exp. Med. Biol. 2017, 1018, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, R.J.A.; Tong, S.; Zong, J.; Chen, Y.; Tsao, S.W.; Pan, J.; Chen, H. NF-kappaB Signaling Regulates Epstein-Barr Virus BamHI-Q-Driven EBNA1 Expression. Cancers 2018, 10, 119. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, G.; Komano, J.; Sugden, B. Epstein-Barr virus provides a survival factor to Burkitt’s lymphomas. Proc. Natl. Acad. Sci. USA 2003, 100, 14269–14274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasimuzzaman, M.; Kuroda, M.; Dohno, S.; Yamamoto, T.; Iwatsuki, K.; Matsuzaki, S.; Mohammad, R.; Kumita, W.; Mizuguchi, H.; Hayakawa, T.; et al. Eradication of Epstein-Barr virus episome and associated inhibition of infected tumor cell growth by adenovirus vector-mediated transduction of dominant-negative EBNA1. Mol. Ther. 2005, 11, 578–590. [Google Scholar] [CrossRef] [PubMed]
- Imai, S.; Kuroda, M.; Yamashita, R.; Ishiura, Y. Therapeutic inhibition of Epstein-Barr virus-associated tumor cell growth by dominant-negative EBNA1. Uirusu 2005, 55, 239–249. [Google Scholar] [CrossRef] [Green Version]
- Roth, G.; Curiel, T.; Lacy, J. Epstein-Barr viral nuclear antigen 1 antisense oligodeoxynucleotide inhibits proliferation of Epstein-Barr virus-immortalized B cells. Blood 1994, 84, 582–587. [Google Scholar] [CrossRef]
- Yin, Q.; Flemington, E.K. siRNAs against the Epstein Barr virus latency replication factor, EBNA1, inhibit its function and growth of EBV-dependent tumor cells. Virology 2006, 346, 385–393. [Google Scholar] [CrossRef] [Green Version]
- Hong, M.; Murai, Y.; Kutsuna, T.; Takahashi, H.; Nomoto, K.; Cheng, C.M.; Ishizawa, S.; Zhao, Q.L.; Ogawa, R.; Harmon, B.V.; et al. Suppression of Epstein-Barr nuclear antigen 1 (EBNA1) by RNA interference inhibits proliferation of EBV-positive Burkitt’s lymphoma cells. J. Cancer Res. Clin. Oncol. 2006, 132, 1–8. [Google Scholar] [CrossRef]
- Pfeffer, S.; Zavolan, M.; Grasser, F.A.; Chien, M.; Russo, J.J.; Ju, J.; John, B.; Enright, A.J.; Marks, D.; Sander, C.; et al. Identification of virus-encoded microRNAs. Science 2004, 304, 734–736. [Google Scholar] [CrossRef]
- Kang, D.; Skalsky, R.L.; Cullen, B.R. EBV BART MicroRNAs Target Multiple Pro-apoptotic Cellular Genes to Promote Epithelial Cell Survival. PLoS Pathog. 2015, 11, e1004979. [Google Scholar] [CrossRef]
- Choy, E.Y.; Siu, K.L.; Kok, K.H.; Lung, R.W.; Tsang, C.M.; To, K.F.; Kwong, D.L.; Tsao, S.W.; Jin, D.Y. An Epstein-Barr virus-encoded microRNA targets PUMA to promote host cell survival. J. Exp. Med. 2008, 205, 2551–2560. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Yao, Y.; Chen, H.; Zhang, S.; Cao, S.M.; Zhang, Z.; Luo, B.; Liu, Z.; Li, Z.; Xiang, T.; et al. Genome sequencing analysis identifies Epstein-Barr virus subtypes associated with high risk of nasopharyngeal carcinoma. Nat. Genet. 2019, 51, 1131–1136. [Google Scholar] [CrossRef] [PubMed]
- Takada, K. Role of EBER and BARF1 in nasopharyngeal carcinoma (NPC) tumorigenesis. Semin. Cancer Biol. 2012, 22, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Nanbo, A.; Inoue, K.; Adachi-Takasawa, K.; Takada, K. Epstein-Barr virus RNA confers resistance to interferon-alpha-induced apoptosis in Burkitt’s lymphoma. EMBO J. 2002, 21, 954–965. [Google Scholar] [CrossRef]
- Tsang, C.M.; Lui, V.W.Y.; Bruce, J.P.; Pugh, T.J.; Lo, K.W. Translational genomics of nasopharyngeal cancer. Semin. Cancer Biol. 2020, 61, 84–100. [Google Scholar] [CrossRef] [PubMed]
- Tsang, C.M.; Yip, Y.L.; Lo, K.W.; Deng, W.; To, K.F.; Hau, P.M.; Lau, V.M.; Takada, K.; Lui, V.W.; Lung, M.L.; et al. Cyclin D1 overexpression supports stable EBV infection in nasopharyngeal epithelial cells. Proc. Natl. Acad. Sci. USA 2012, 109, E3473–E3482. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Dai, W.; Cheung, A.K.; Ko, J.M.; Kan, R.; Wong, B.W.; Leong, M.M.; Deng, M.; Kwok, T.C.; Chan, J.Y.; et al. Whole-exome sequencing identifies multiple loss-of-function mutations of NF-kappaB pathway regulators in nasopharyngeal carcinoma. Proc. Natl. Acad. Sci. USA 2016, 113, 11283–11288. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.Y.; Chung, G.T.; Lui, V.W.; To, K.F.; Ma, B.B.; Chow, C.; Woo, J.K.; Yip, K.Y.; Seo, J.; Hui, E.P.; et al. Exome and genome sequencing of nasopharynx cancer identifies NF-kappaB pathway activating mutations. Nat. Commun. 2017, 8, 14121. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.C.; Meng, X.; Hazawa, M.; Nagata, Y.; Varela, A.M.; Xu, L.; Sato, Y.; Liu, L.Z.; Ding, L.W.; Sharma, A.; et al. The genomic landscape of nasopharyngeal carcinoma. Nat. Genet. 2014, 46, 866–871. [Google Scholar] [CrossRef]
- Bruce, J.P.; To, K.F.; Lui, V.W.Y.; Chung, G.T.Y.; Chan, Y.Y.; Tsang, C.M.; Yip, K.Y.; Ma, B.B.Y.; Woo, J.K.S.; Hui, E.P.; et al. Whole-genome profiling of nasopharyngeal carcinoma reveals viral-host co-operation in inflammatory NF-kappaB activation and immune escape. Nat. Commun. 2021, 12, 4193. [Google Scholar] [CrossRef]
- Lo, K.W.; Chung, G.T.; To, K.F. Deciphering the molecular genetic basis of NPC through molecular, cytogenetic, and epigenetic approaches. Semin. Cancer Biol. 2012, 22, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Leong, M.M.L.; Cheung, A.K.L.; Dai, W.; Tsao, S.W.; Tsang, C.M.; Dawson, C.W.; Mun Yee Ko, J.; Lung, M.L. EBV infection is associated with histone bivalent switch modifications in squamous epithelial cells. Proc. Natl. Acad. Sci. USA 2019, 116, 14144–14153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, L.; Zhou, H.; Wang, C.; Xiong, G.; Xiang, Y.; Ling, Y.; Khabir, A.; Tsao, G.S.; Zeng, Y.; Zeng, M.; et al. Nasopharyngeal carcinoma super-enhancer-driven ETV6 correlates with prognosis. Proc. Natl. Acad. Sci. USA 2017, 114, 9683–9688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, A.K.; OuYang, C.N.; Liu, H.; Chao, M.; Luo, J.D.; Lee, C.Y.; Lu, Y.J.; Chung, I.C.; Chen, L.C.; Wu, S.M.; et al. Targeted sequencing of cancer-related genes in nasopharyngeal carcinoma identifies mutations in the TGF-beta pathway. Cancer Med. 2019, 8, 5116–5127. [Google Scholar] [CrossRef] [Green Version]
- Msaouel, P.; Nandikolla, G.; Pneumaticos, S.G.; Koutsilieris, M. Bone microenvironment-targeted manipulations for the treatment of osteoblastic metastasis in castration-resistant prostate cancer. Expert Opin. Investig. Drugs 2013, 22, 1385–1400. [Google Scholar] [CrossRef]
- Schartinger, V.H.; Dudas, J.; Url, C.; Reinold, S.; Virgolini, I.J.; Kroiss, A.; Riechelmann, H.; Uprimny, C. (68)Ga-DOTA (0)-Tyr (3)-octreotide positron emission tomography in nasopharyngeal carcinoma. Eur. J. Nucl. Med. Mol. Imaging 2015, 42, 20–24. [Google Scholar] [CrossRef]
- Bennink, R.J.; van der Meulen, F.W.; Freling, N.J.; Booij, J. Somatostatin receptor scintigraphy in nasopharyngeal carcinoma. Clin. Nucl. Med. 2008, 33, 558–561. [Google Scholar] [CrossRef]
- Pokuri, V.K.; Fong, M.K.; Iyer, R. Octreotide and Lanreotide in Gastroenteropancreatic Neuroendocrine Tumors. Curr. Oncol. Rep. 2016, 18, 7. [Google Scholar] [CrossRef]
- Chung, G.T.; Lou, W.P.; Chow, C.; To, K.F.; Choy, K.W.; Leung, A.W.; Tong, C.Y.; Yuen, J.W.; Ko, C.W.; Yip, T.T.; et al. Constitutive activation of distinct NF-kappaB signals in EBV-associated nasopharyngeal carcinoma. J. Pathol. 2013, 231, 311–322. [Google Scholar] [CrossRef]
- Edwards, R.H.; Marquitz, A.R.; Raab-Traub, N. Changes in expression induced by Epstein-Barr Virus LMP1-CTAR1: Potential role of bcl3. mBio 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Baud, V.; Karin, M. Is NF-kappaB a good target for cancer therapy? Hopes and pitfalls. Nat. Rev. Drug Discov. 2009, 8, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Huen, D.S.; Henderson, S.A.; Croom-Carter, D.; Rowe, M. The Epstein-Barr virus latent membrane protein-1 (LMP1) mediates activation of NF-kappa B and cell surface phenotype via two effector regions in its carboxy-terminal cytoplasmic domain. Oncogene 1995, 10, 549–560. [Google Scholar]
- Xu, Y.J.; Zhou, R.; Zong, J.F.; Lin, W.S.; Tong, S.; Guo, Q.J.; Lin, C.; Lin, S.J.; Chen, Y.X.; Chen, M.R.; et al. Epstein-Barr virus-coded miR-BART13 promotes nasopharyngeal carcinoma cell growth and metastasis via targeting of the NKIRAS2/NF-kappaB pathway. Cancer Lett. 2019, 447, 33–40. [Google Scholar] [CrossRef]
- Gabriel, M.; Decristoforo, C.; Kendler, D.; Dobrozemsky, G.; Heute, D.; Uprimny, C.; Kovacs, P.; Von Guggenberg, E.; Bale, R.; Virgolini, I.J. 68Ga-DOTA-Tyr3-octreotide PET in neuroendocrine tumors: Comparison with somatostatin receptor scintigraphy and CT. J. Nucl. Med. 2007, 48, 508–518. [Google Scholar] [CrossRef]
- Deppen, S.A.; Liu, E.; Blume, J.D.; Clanton, J.; Shi, C.; Jones-Jackson, L.B.; Lakhani, V.; Baum, R.P.; Berlin, J.; Smith, G.T.; et al. Safety and Efficacy of 68Ga-DOTATATE PET/CT for Diagnosis, Staging, and Treatment Management of Neuroendocrine Tumors. J. Nucl. Med. 2016, 57, 708–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, P.; Ginj, M.; Zhang, H.; Waser, B.; Baum, R.P.; Reubi, J.C.; Maecke, H. Are radiogallium-labelled DOTA-conjugated somatostatin analogues superior to those labelled with other radiometals? Eur. J. Nucl. Med. Mol. Imaging 2007, 34, 982–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khor, L.K.; Loi, H.Y.; Sinha, A.K.; Tong, K.T.; Goh, B.C.; Loh, K.S.; Lu, S.J. (68)Ga-DOTA-peptide: A novel molecular biomarker for nasopharyngeal carcinoma. Head Neck 2016, 38, E76–E80. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.C.; Smith, G.T.; Liu, E.; Moore, B.; Clanton, J.; Stabin, M. Measured human dosimetry of 68Ga-DOTATATE. J. Nucl. Med. 2013, 54, 855–860. [Google Scholar] [CrossRef] [Green Version]
- Treatment Options by Stage of Nasopharyngeal Cancer. Available online: https://www.cancer.org/cancer/nasopharyngeal-cancer/treating/by-stage.html (accessed on 28 June 2021).
- Blanchard, P.; Lee, A.; Marguet, S.; Leclercq, J.; Ng, W.T.; Ma, J.; Chan, A.T.; Huang, P.Y.; Benhamou, E.; Zhu, G.; et al. Chemotherapy and radiotherapy in nasopharyngeal carcinoma: An update of the MAC-NPC meta-analysis. Lancet Oncol. 2015, 16, 645–655. [Google Scholar] [CrossRef]
- American Cancer Society. Immunotherapy for Nasopharyngeal Cancer. Available online: https://www.cancer.org/cancer/nasopharyngeal-cancer/treating/immunotherapy.html (accessed on 28 June 2021).
- Zhao, C.; Miao, J.; Shen, G.; Li, J.; Shi, M.; Zhang, N.; Hu, G.; Chen, X.; Hu, X.; Wu, S.; et al. Anti-epidermal growth factor receptor (EGFR) monoclonal antibody combined with cisplatin and 5-fluorouracil in patients with metastatic nasopharyngeal carcinoma after radical radiotherapy: A multicentre, open-label, phase II clinical trial. Ann. Oncol. 2019, 30, 637–643. [Google Scholar] [CrossRef]
- Golden, E.B.; Frances, D.; Pellicciotta, I.; Demaria, S.; Helen Barcellos-Hoff, M.; Formenti, S.C. Radiation fosters dose-dependent and chemotherapy-induced immunogenic cell death. Oncoimmunology 2014, 3, e28518. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yu, X.M.; Sun, R.J.; Pan, X.L. Folate-Functionalized Lipid Nanoemulsion to Deliver Chemo-Radiotherapeutics Together for the Effective Treatment of Nasopharyngeal Carcinoma. AAPS PharmSciTech 2017, 18, 1374–1381. [Google Scholar] [CrossRef]
- van Essen, M.; Krenning, E.P.; Kam, B.L.; de Jong, M.; Valkema, R.; Kwekkeboom, D.J. Peptide-receptor radionuclide therapy for endocrine tumors. Nat. Rev. Endocrinol. 2009, 5, 382–393. [Google Scholar] [CrossRef] [PubMed]
- van der Zwan, W.A.; Bodei, L.; Mueller-Brand, J.; de Herder, W.W.; Kvols, L.K.; Kwekkeboom, D.J. GEPNETs update: Radionuclide therapy in neuroendocrine tumors. Eur. J. Endocrinol. 2015, 172, R1–R8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.; Zhang, J.; Singh, A.; Kulkarni, H.R.; Baum, R.P. Metastatic Nasopharyngeal Carcinoma Treated With Intraarterial Combined With Intravenous Peptide Receptor Radionuclide Therapy. Clin. Nucl. Med. 2019, 44, 989–990. [Google Scholar] [CrossRef]
- Masterson, L.; Howard, J.; Gonzalez-Cruz, J.; Jackson, C.; Barnett, C.; Overton, L.; Liu, H.; Ladwa, R.; Simpson, F.; McGrath, M.; et al. Immune checkpoint inhibitors in advanced nasopharyngeal carcinoma: Beyond an era of chemoradiation? Int. J. Cancer 2020, 146, 2305–2314. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.; Ma, B.B.Y. Targeting the PD-1/ PD-L1 interaction in nasopharyngeal carcinoma. Oral Oncol. 2021, 113, 105127. [Google Scholar] [CrossRef]
- Fang, W.; Zhang, J.; Hong, S.; Zhan, J.; Chen, N.; Qin, T.; Tang, Y.; Zhang, Y.; Kang, S.; Zhou, T.; et al. EBV-driven LMP1 and IFN-gamma up-regulate PD-L1 in nasopharyngeal carcinoma: Implications for oncotargeted therapy. Oncotarget 2014, 5, 12189–12202. [Google Scholar] [CrossRef]
- Yoshizaki, T.; Kondo, S.; Wakisaka, N.; Murono, S.; Endo, K.; Sugimoto, H.; Nakanishi, S.; Tsuji, A.; Ito, M. Pathogenic role of Epstein-Barr virus latent membrane protein-1 in the development of nasopharyngeal carcinoma. Cancer Lett. 2013, 337, 1–7. [Google Scholar] [CrossRef]
- Zheng, H.; Li, L.L.; Hu, D.S.; Deng, X.Y.; Cao, Y. Role of Epstein-Barr virus encoded latent membrane protein 1 in the carcinogenesis of nasopharyngeal carcinoma. Cell Mol. Immunol. 2007, 4, 185–196. [Google Scholar]
- Morris, M.A.; Dawson, C.W.; Young, L.S. Role of the Epstein-Barr virus-encoded latent membrane protein-1, LMP1, in the pathogenesis of nasopharyngeal carcinoma. Future Oncol. 2009, 5, 811–825. [Google Scholar] [CrossRef]
- Wu, Y.; Pfeifer, A.K.; Myschetzky, R.; Garbyal, R.S.; Rasmussen, P.; Knigge, U.; Bzorek, M.; Kristensen, M.H.; Kjaer, A. Induction of Anti-Tumor Immune Responses by Peptide Receptor Radionuclide Therapy with (177)Lu-DOTATATE in a Murine Model of a Human Neuroendocrine Tumor. Diagnostics 2013, 3, 344–355. [Google Scholar] [CrossRef]
- Kasi, P.M.; Sharma, A.; Jain, M.K. Expanding the Indication for Novel Theranostic 177Lu-Dotatate Peptide Receptor Radionuclide Therapy: Proof-of-Concept of PRRT in Merkel Cell Cancer. Case Rep. Oncol. 2019, 12, 98–103. [Google Scholar] [CrossRef]
- Msaouel, P.; Galanis, E.; Koutsilieris, M. Somatostatin and somatostatin receptors: Implications for neoplastic growth and cancer biology. Expert Opin. Investig. Drugs 2009, 18, 1297–1316. [Google Scholar] [CrossRef]
- Caplin, M.E.; Pavel, M.; Cwikla, J.B.; Phan, A.T.; Raderer, M.; Sedlackova, E.; Cadiot, G.; Wolin, E.M.; Capdevila, J.; Wall, L.; et al. Lanreotide in metastatic enteropancreatic neuroendocrine tumors. N. Engl. J. Med. 2014, 371, 224–233. [Google Scholar] [CrossRef] [PubMed]
- White, B.H.; Whalen, K.; Kriksciukaite, K.; Alargova, R.; Au Yeung, T.; Bazinet, P.; Brockman, A.; DuPont, M.; Oller, H.; Lemelin, C.A.; et al. Discovery of an SSTR2-Targeting Maytansinoid Conjugate (PEN-221) with Potent Activity in Vitro and in Vivo. J. Med. Chem. 2019, 62, 2708–2719. [Google Scholar] [CrossRef] [PubMed]
- Si, Y.; Kim, S.; Ou, J.; Lu, Y.; Ernst, P.; Chen, K.; Whitt, J.; Carter, A.M.; Markert, J.M.; Bibb, J.A.; et al. Anti-SSTR2 antibody-drug conjugate for neuroendocrine tumor therapy. Cancer Gene Ther. 2021, 28, 799–812. [Google Scholar] [CrossRef] [PubMed]
- Gauzy-Lazo, L.; Sassoon, I.; Brun, M.P. Advances in Antibody-Drug Conjugate Design: Current Clinical Landscape and Future Innovations. SLAS Discov. 2020, 25, 843–868. [Google Scholar] [CrossRef]
- Whalen, K.A.; White, B.H.; Quinn, J.M.; Kriksciukaite, K.; Alargova, R.; Au Yeung, T.P.; Bazinet, P.; Brockman, A.; DuPont, M.M.; Oller, H.; et al. Targeting the Somatostatin Receptor 2 with the Miniaturized Drug Conjugate, PEN-221: A Potent and Novel Therapeutic for the Treatment of Small Cell Lung Cancer. Mol. Cancer Ther. 2019, 18, 1926–1936. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.L.; Meyer, T.; Halperin, D.M.; Fojo, A.T.; Cook, N.; Blaszkowsky, L.S.; Schlechter, B.L.; Yao, J.C.; Jemiai, Y.; Kriksciukaite, K.; et al. First in human phase 1/2a study of PEN-221 somatostatin analog (SSA)-DM1 conjugate for patients (PTS) with advanced neuroendocrine tumor (NET) or small cell lung cancer (SCLC): Phase 1 results. J. Clin. Oncol. 2018, 36, 4097. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emanuel, O.; Liu, J.; Schartinger, V.H.; Nei, W.L.; Chan, Y.Y.; Tsang, C.M.; Riechelmann, H.; Masterson, L.; Haybaeck, J.; Oppermann, U.; et al. SSTR2 in Nasopharyngeal Carcinoma: Relationship with Latent EBV Infection and Potential as a Therapeutic Target. Cancers 2021, 13, 4944. https://doi.org/10.3390/cancers13194944
Emanuel O, Liu J, Schartinger VH, Nei WL, Chan YY, Tsang CM, Riechelmann H, Masterson L, Haybaeck J, Oppermann U, et al. SSTR2 in Nasopharyngeal Carcinoma: Relationship with Latent EBV Infection and Potential as a Therapeutic Target. Cancers. 2021; 13(19):4944. https://doi.org/10.3390/cancers13194944
Chicago/Turabian StyleEmanuel, Oscar, Jacklyn Liu, Volker H. Schartinger, Wen Long Nei, Yuk Yu Chan, Chi Man Tsang, Herbert Riechelmann, Liam Masterson, Johannes Haybaeck, Udo Oppermann, and et al. 2021. "SSTR2 in Nasopharyngeal Carcinoma: Relationship with Latent EBV Infection and Potential as a Therapeutic Target" Cancers 13, no. 19: 4944. https://doi.org/10.3390/cancers13194944
APA StyleEmanuel, O., Liu, J., Schartinger, V. H., Nei, W. L., Chan, Y. Y., Tsang, C. M., Riechelmann, H., Masterson, L., Haybaeck, J., Oppermann, U., Willems, S. M., Ooft, M. L., Wollmann, G., Howard, D., Vanhaesebroeck, B., Lund, V. J., Royle, G., Chua, M. L. K., Lo, K. W., ... Lechner, M. (2021). SSTR2 in Nasopharyngeal Carcinoma: Relationship with Latent EBV Infection and Potential as a Therapeutic Target. Cancers, 13(19), 4944. https://doi.org/10.3390/cancers13194944