
The RXR Agonist MSU42011 Is Effective for the Treatment of Preclinical HER2+ Breast Cancer and Kras-Driven Lung Cancer

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Drugs
2.2. Animal Experiments
2.3. Flow Cytometry
2.4. Immunohistochemistry
2.5. Ultrasound
2.6. Lipid Levels in Plasma
2.7. Patient Survival Analysis
2.8. Statistical Analysis
3. Results
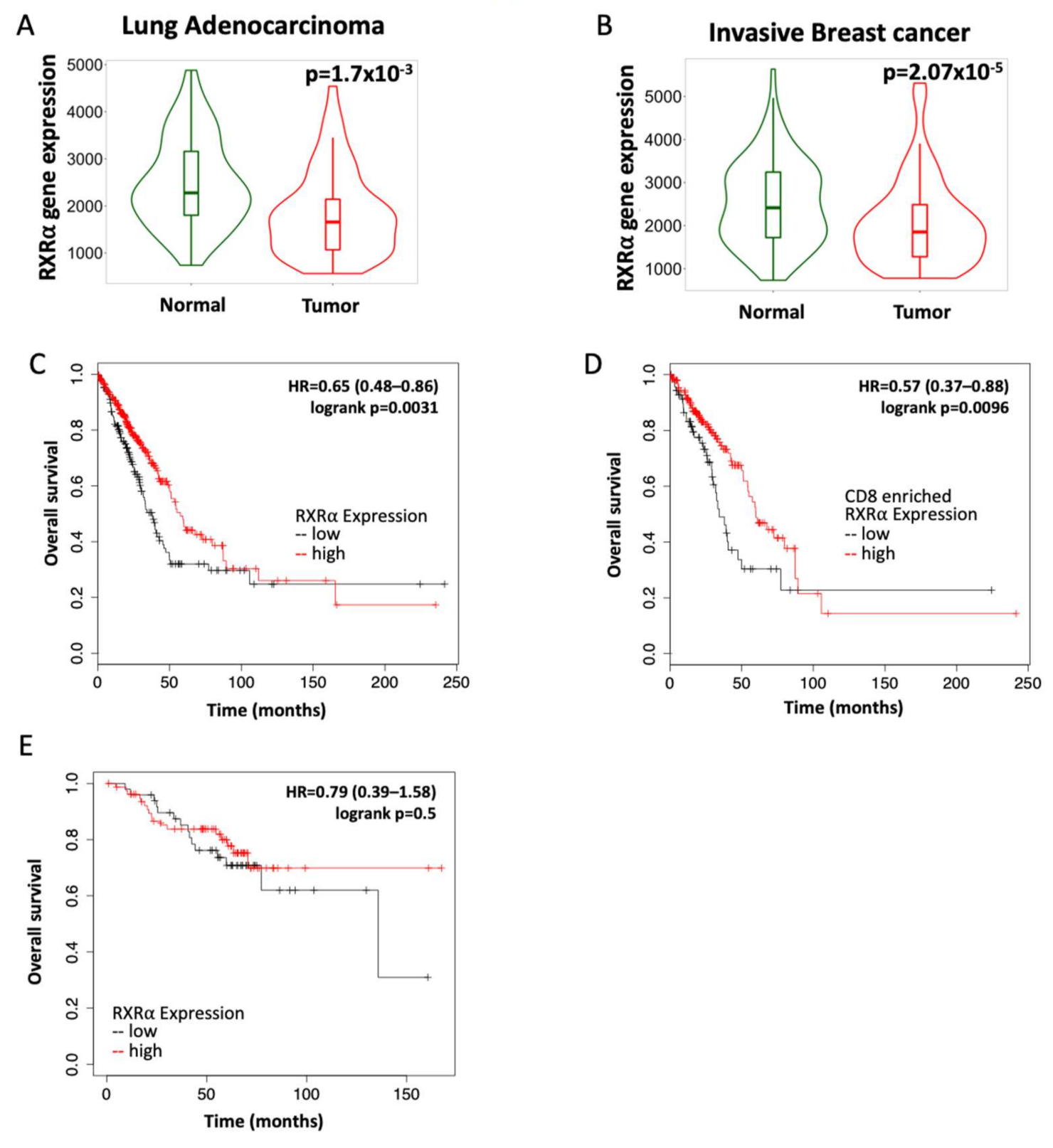
3.1. RXR Expression Correlates with Increased Survival in Lung and Breast Cancers
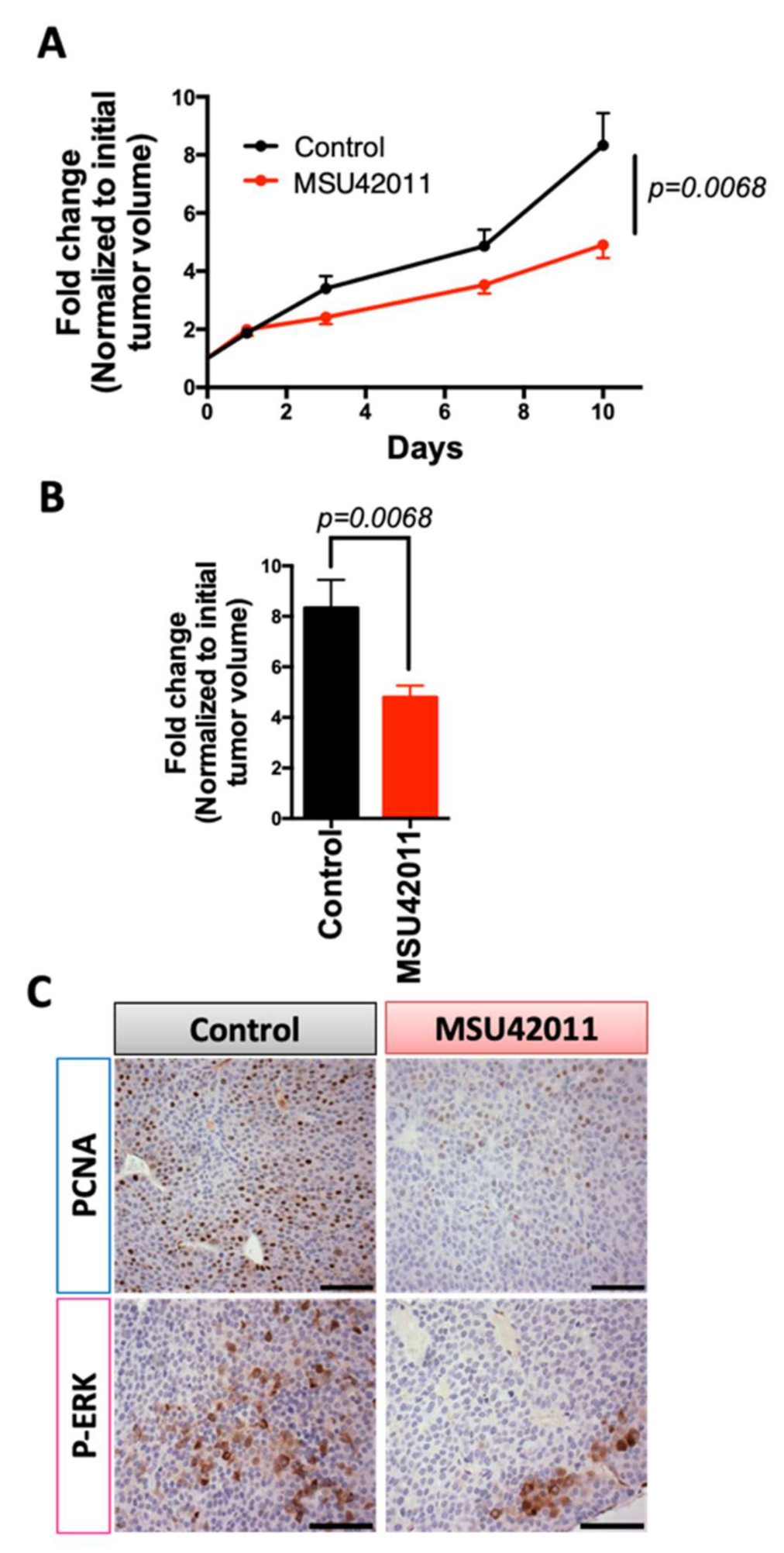
3.2. MSU42011 Reduced Tumor Growth in the MMTV-Neu Model of Breast Cancer and the A/J Mouse Model of Lung Cancer
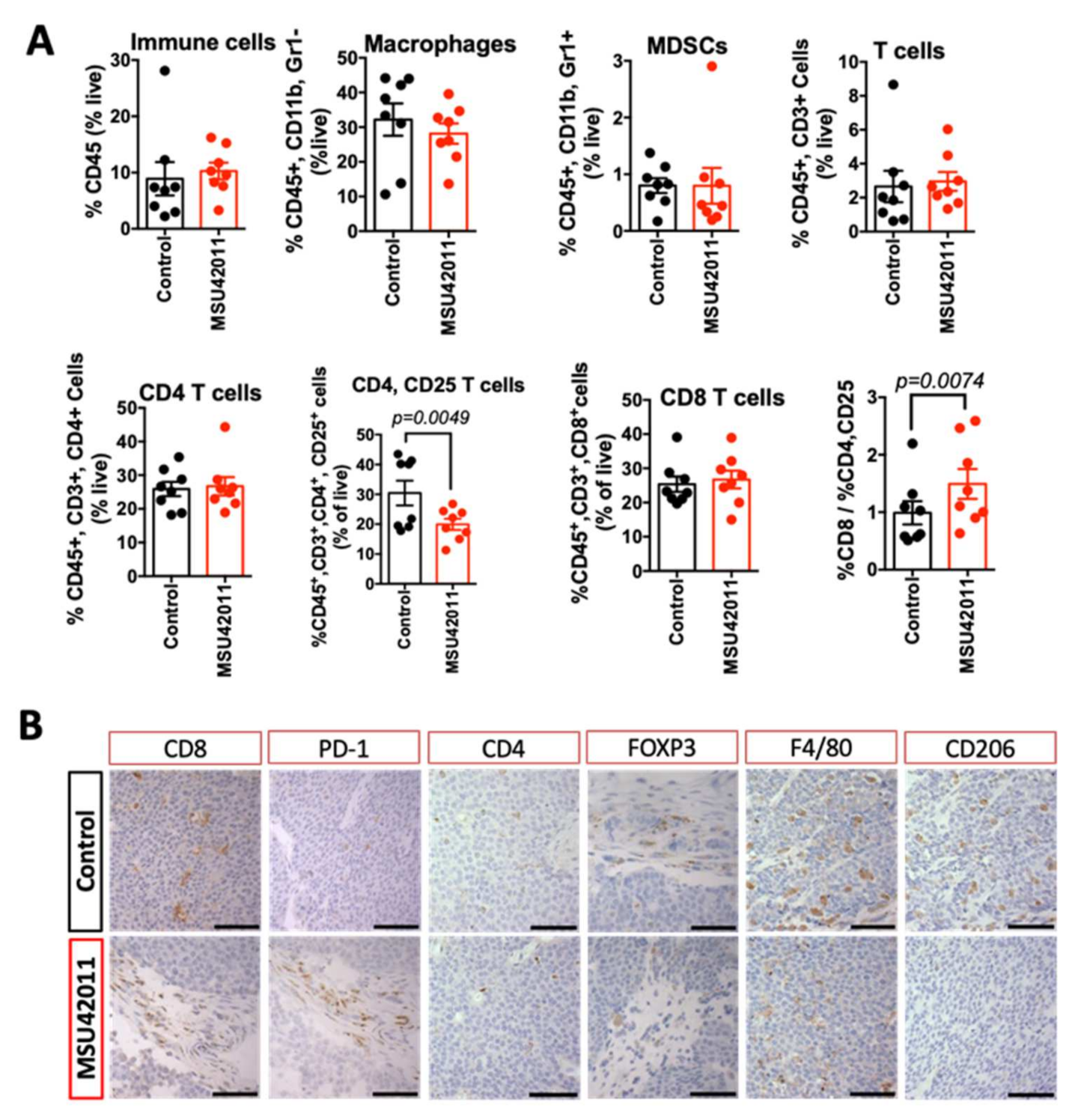
3.3. MSU42011 Induced an Anti-Tumor Immune Response in the Tumor Microenvironment
3.4. MSU42011 Was Ineffective for Suppressing Tumor Growth in a Human A549 Lung Cancer Xenograft Model
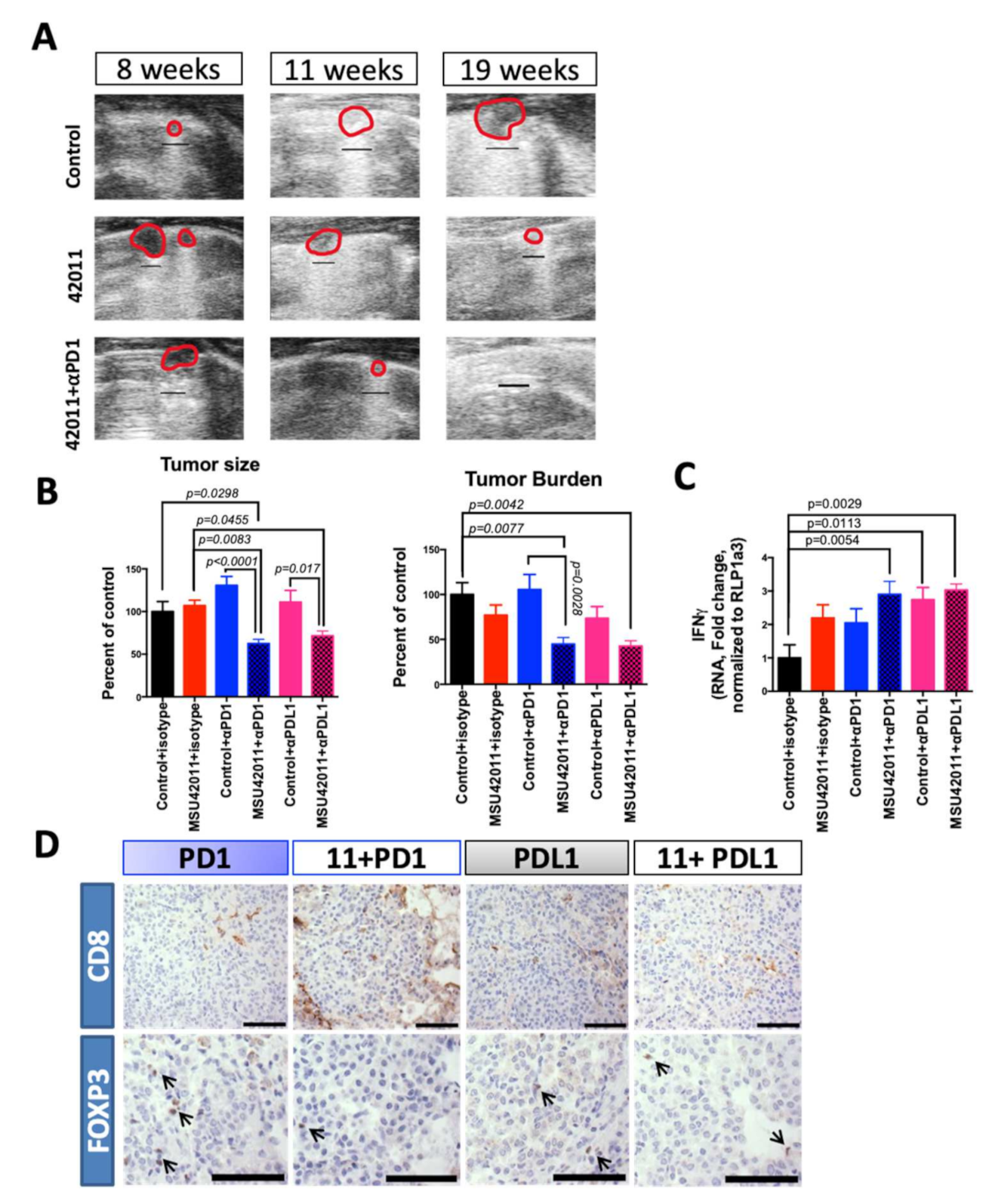
3.5. MSU42011 in Combination with Anti-PD1 or Anti-PDL1 Antibodies Was Effective for Treating Established Lung Tumors in the A/J Mouse Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71. [Google Scholar] [CrossRef] [PubMed]
- Amanam, I.; Mambetsariev, I.; Gupta, R.; Achuthan, S.; Wang, Y.; Pharaon, R.; Massarelli, E.; Koczywas, M.; Reckamp, K.; Salgia, R. Role of immunotherapy and co-mutations on KRAS-mutant nonsmall cell lung cancer survival. J. Thorac. Dis. 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Fridman, W.H.; Pagès, F.; Sautès-Fridman, C.; Galon, J. The immune contexture in human tumours: Impact on clinical outcome. Nat. Rev. Cancer 2012, 12, 298–306. [Google Scholar] [CrossRef]
- Albini, A.; Sporn, M.B. The tumor microenvironment as a target for chemoprevention. Nat. Rev. Cancer 2007, 7, 139–147. [Google Scholar] [CrossRef]
- Klemm, F.; Joyce, J.A. Microenvironmental regulation of therapeutic response in cancer. Trends Cell Biol. 2015, 25, 198–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittal, D.; Gubin, M.M.; Schreiber, R.D.; Smyth, M.J. New insights into cancer immunoediting and its three component phases-elimination, equilibrium and escape. Curr. Opin. Immunol. 2014, 27, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Leal, A.S.; Zydeck, K.; Carapellucci, S.; Reich, L.A.; Zhang, D.; Moerland, J.A.; Sporn, M.B.; Liby, K.T. Retinoid X receptor agonist LG100268 modulates the immune microenvironment in preclinical breast cancer models. NPJ Breast Cancer 2019, 5, 39. [Google Scholar] [CrossRef]
- Roszer, T.; Menéndez-Gutiérrez, M.P.; Cedenilla, M.; Ricote, M. Retinoid X receptors in macrophage biology. Trends Endocrinol. Metab. 2013, 24, 460–468. [Google Scholar] [CrossRef]
- Kiss, M.; Czimmerer, Z.; Nagy, G.; Bieniasz-Krzywiec, P.; Ehling, M.; Pap, A.; Poliska, S.; Boto, P.; Tzerpos, P.; Horvath, A.; et al. Retinoid X receptor suppresses a metastasis-promoting transcriptional program in myeloid cells via a ligand-insensitive mechanism. Proc. Natl. Acad. Sci. USA 2017, 114, 10725–10730. [Google Scholar] [CrossRef] [Green Version]
- Casanova-Acebes, M.; Menéndez-Gutiérrez, M.P.; Porcuna, J.; Álvarez-Errico, D.; Lavin, Y.; García, A.; Kobayashi, S.; Le Berichel, J.; Núñez, V.; Were, F.; et al. RXRs control serous macrophage neonatal expansion and identity and contribute to ovarian cancer progression. Nat. Commun. 2020, 11, 1655. [Google Scholar] [CrossRef]
- Mangelsdorft, D.J.; Ong, E.S.; Dyck, J.A.; Evans, R.M. Nuclear Receptor that Identifies a Novel Retinoic Acid Response Pathway. Nature 1990, 345, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Dawson, M.I.; Xia, Z. The retinoid X receptors and their ligands. BBA-Mol. Cell Biol. Lipids 2012, 1821, 21–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szeles, L.; Poliska, S.; Nagy, G.; Szatmari, I.; Szanto, A.; Pap, A.; Lindstedt, M.; Sategoets, S.J.A.M.; Ruhl, R.; Dezso, B.; et al. Research Resource: Transcriptome Profiling of Genes Regulated by RXR and Its Permissive and Nonpermissive Partners in Differentiating. Mol. Endocrinol. 2010, 24, 2218–2231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Núñez, V.; Alameda, D.; Rico, D.; Mota, R.; Gonzalo, P.; Cedenilla, M.; Fischer, T. Retinoid X receptor α controls innate in fl ammatory responses through the up-regulation of chemokine expression. Proc. Natl. Acad. Sci. USA 2010, 107, 10626–10631. [Google Scholar] [CrossRef] [Green Version]
- Liby, T.K.; Sporn, B.M. Rexinoids for Prevention and Treatment of Cancer: Opportunities and Challenges. Curr. Top. Med. Chem. 2017, 17, 721–730. [Google Scholar] [CrossRef]
- Qu, L.; Tang, Æ.X. Bexarotene: A promising anticancer agent. Cancer Chemother. Pharmacol. 2010, 65, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Esteva, F.J.; Glaspy, J.; Baidas, S.; Laufman, L.; Hutchins, L.; Dickler, M.; Tripathy, D.; Cohen, R.; DeMichele, A.; Yocum, R.C.; et al. Multicenter phase II study of oral bexarotene for patients with metastatic breast cancer. J. Clin. Oncol. 2003, 21, 999–1006. [Google Scholar] [CrossRef]
- Tyagi, P.; Belani, C.P.; Jain, V.K. Bexarotene in combination with chemotherapy fails to prolong survival in patients with advanced non-small-cell lung cancer: Results from the SPIRIT I and II trials. Clin. Lung Cancer 2005, 7, 17–19. [Google Scholar] [CrossRef]
- Pérez, E.; Bourguet, W.; Gronemeyer, H.; Lera, A.R. De Modulation of RXR function through ligand design. BBA-Mol. Cell Biol. Lipids 2012, 1821, 57–69. [Google Scholar] [CrossRef]
- Boehm, M.F.; Zhang, L.; Badea, B.A.; White, S.K.; Mais, D.E.; Berger, E.; Suto, C.M.; Goldman, M.E.; Heyman, R.A. Synthesis and Structure-Activity Relationships of Novel Retinoid X Receptor-Selective Retinoids. J. Med. Chem. 1994, 37, 2930–2941. [Google Scholar] [CrossRef]
- Cao, M.; Royce, D.B.; Risingsong, R.; Williams, C.R.; Sporn, M.B.; Liby, K.T. The rexinoids LG100268 and LG101506 inhibit inflammation and suppress lung carcinogenesis in A/J mice. Cancer Prev. Res. 2016, 9, 105–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moerland, J.A.; Zhang, D.; Reich, L.A.; Carapellucci, S.; Lockwood, B.; Leal, A.S.; Krieger-Burke, T.; Aleiwi, B.; Ellsworth, E.; Liby, K.T. The novel rexinoid MSU-42011 is effective for the treatment of preclinical Kras-driven lung cancer. Sci. Rep. 2020, 10, 22244. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Leal, A.S.; Carapellucci, S.; Shahani, P.H.; Bhogal, J.S.; Ibrahim, S.; Raban, S.; Jurutka, P.W.; Marshall, P.A.; Sporn, M.B.; et al. Testing novel pyrimidinyl rexinoids: A new paradigm for evaluating rexinoids for cancer prevention. Cancer Prev. Res. 2019, 12, 211–224. [Google Scholar] [CrossRef] [Green Version]
- DeNardo, D.G.; Brennan, D.J.; Rexhepaj, E.; Ruffell, B.; Shiao, S.L.; Madden, S.F.; Gallagher, W.M.; Wadhwani, N.; Keil, S.D.; Junaid, S.A.; et al. Leukocyte complexity predicts breast cancer survival and functionally regulates response to chemotherapy. Cancer Discov. 2011, 1, 54–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R. A Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altorki, N.K.; Markowitz, G.J.; Gao, D.; Port, J.L.; Saxena, A.; Stiles, B.; McGraw, T.; Mittal, V. The lung microenvironment: An important regulator of tumour growth and metastasis. Nat. Rev. Cancer 2019, 19, 9–31. [Google Scholar] [CrossRef] [Green Version]
- Leal, A.S.; Reich, L.A.; Moerland, J.A.; Zhang, D.; Liby, K.T. Potential Therapeutic Uses of Rexinoids, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Fry, E.A.; Taneja, P.; Inoue, K. Clinical applications of mouse models for breast cancer engaging HER2/neu. Integr. Cancer Sci. Ther. 2016, 3, 593–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, M.; Candrian, U.; Maronpot, R.R.; Stoner, G.D.; Anderson, M.W. Activation of the Ki-ras protooncogene in spontaneously occurring and chemically induced lung tumors of the strain A mouse. Proc. Natl. Acad. Sci. USA 1989, 86, 3070–3074. [Google Scholar] [CrossRef] [Green Version]
- Massey, T.E.; Devereux, T.R.; Maronpot, R.R.; Foley, J.F.; Anderson, M.W. High frequency of K-ras mutations in spontaneous and vinyl carbamate-induced lung tumors of relatively resistant B6CF1 (C57BL6J x BALCcJ) mice. Lung Cancer 1995, 15, 1065–1069. [Google Scholar] [CrossRef]
- Zhang, D.; Rennhack, J.; Andrechek, E.R.; Rockwell, C.E.; Liby, K.T. Identification of an Unfavorable Immune Signature in Advanced Lung Tumors from Nrf2-Deficient Mice. Antioxid. Redox Signal. 2018, 29, 1535–1552. [Google Scholar] [CrossRef]
- Zhang, D.; Leal, A.S.; Carapellucci, S.; Zydeck, K.; Sporn, M.B.; Liby, K.T. Chemoprevention of Preclinical Breast and Lung Cancer with the Bromodomain Inhibitor I-BET 762. Cancer Prev. Res. 2018, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liby, K.; Risingsong, R.; Royce, D.B.; Williams, C.R.; Yore, M.M.; Honda, T.; Gribble, G.W.; Lamph, W.W.; Vannini, N.; Sogno, I.; et al. Prevention and treatment of experimental estrogen receptor- negative mammary carcinogenesis by the synthetic triterpenoid CDDO-methyl ester and the rexinoid LG100268. Clin. Cancer Res. 2008, 14, 4556–4563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartha, Á.; Győrffy, B. Tnmplot.Com: A web tool for the comparison of gene expression in normal, tumor and metastatic tissues. Int. J. Mol. Sci. 2021, 22, 2622. [Google Scholar] [CrossRef] [PubMed]
- Nagy, Á.; Munkácsy, G.; Győrffy, B. Pancancer survival analysis of cancer hallmark genes. Sci. Rep. 2021, 11. [Google Scholar] [CrossRef]
- Wu, K.; Zhang, Y.; Xu, X.; Hill, J.; Celestino, J.; Kim, H.; Mohsin, S.K.; Hilsenbeck, S.G.; Lamph, W.W.; Bissonette, R.; et al. The Retinoid X Receptor-Selective Retinoid, LGD1069, Prevents the Development of Estrogen Receptor-Negative Mammary Tumors in Transgenic Mice Estrogen Receptor-Negative Mammary Tumors in Transgenic Mice 1. Cancer Res. 2002, 62, 6376–6380. [Google Scholar]
- Li, Y.; Zhang, Y.; Hill, J.; Kim, H.T.; Shen, Q.; Bissonnette, R.P.; Lamph, W.W.; Brown, P.H. The rexinoid, bexarotene, prevents the development of premalignant lesions in MMTV-erbB2 mice. Br. J. Cancer 2008, 98, 1380–1388. [Google Scholar] [CrossRef] [Green Version]
- Liby, K.; Rendi, M.; Suh, N.; Royce, D.B.; Risingsong, R.; Williams, C.R.; Lamph, W.; Labrie, F.; Krajewski, S.; Xu, X.; et al. The combination of the rexinoid, LG100268, and a selective estrogen receptor modulator, either arzoxifene or acolbifene, synergizes in the prevention and treatment of mammary tumors in an estrogen receptor-negative model of breast cancer. Clin. Cancer Res. 2006, 12, 5902–5909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.-T.; Kong, G.; DeNardo, D.; Li, Y.; Uray, I.; Pal, S.; Mohsin, S.; Hilsenbeck, S.G.; Bissonnette, R.; Lamph, W.W.; et al. Identification of Biomarkers Modulated by the Rexinoid LGD1069 (Bexarotene) in Human Breast Cells Using Oligonucleotide Arrays. Cancer Res. 2006, 66, 12009–12018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roskoski, R. Properties of FDA-approved small molecule protein kinase inhibitors: A 2021 update. Pharmacol. Res. 2021. [Google Scholar] [CrossRef]
- Roskoski, R. Small molecule inhibitors targeting the EGFR/ErbB family of protein-tyrosine kinases in human cancers. Pharmacol. Res. 2019, 139. [Google Scholar] [CrossRef] [PubMed]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA - J. Am. Med. Assoc. 2019, 321, 288–300. [Google Scholar] [CrossRef]
- Harari, D.; Yarden, Y. Molecular mechanisms underlying ErbB2/HER2 action in breast cancer. Oncogene 2000, 19, 6102–6114. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, Y.; Sakimura, A.; Park, C.M.; Tomaru, R.; Tanaka, T.; Ozawa, T.; Zhou, Y.; Narita, K.; Kishi, H.; Muraguchi, A.; et al. Feedback control of ErbB2 via ERK-mediated phosphorylation of a conserved threonine in the juxtamembrane domain. Sci. Rep. 2016, 6, 31502. [Google Scholar] [CrossRef]
- Hernandez, L.G.; Forkert, P.G. Inhibition of vinyl carbamate-induced lung tumors and Kras2 mutations by the garlic derivative diallyl sulfone. Mutat. Res.-Fundam. Mol. Mech. Mutagen. 2009, 662, 16–21. [Google Scholar] [CrossRef]
- Tan, W.; Zhang, W.; Strasner, A.; Grivennikov, S.; Cheng, J.Q.; Hoffman, R.M.; Karin, M. Tumour-infiltrating regulatory T cells stimulate mammary cancer metastasis through RANKL-RANK signalling. Nature 2011, 470, 548–553. [Google Scholar] [CrossRef] [Green Version]
- Josefowicz, S.Z.; Lu, L.-F.F.; Rudensky, A.Y. Regulatory T Cells: Mechanisms of Differentiation and Function. Annu. Rev. Immunol. 2012, 30, 531–564. [Google Scholar] [CrossRef] [PubMed]
- Hiraoka, N.; Onozato, K.; Kosuge, T.; Hirohashi, S. Prevalence of FOXP3+ regulatory T cells increases during the progression of pancreatic ductal adenocarcinoma and its premalignant lesions. Clin. Cancer Res. 2006, 12, 5423–5434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linde, N.; Casanova-Acebes, M.; Sosa, M.S.; Mortha, A.; Rahman, A.; Farias, E.; Harper, K.; Tardio, E.; Reyes Torres, I.; Jones, J.; et al. Macrophages orchestrate breast cancer early dissemination and metastasis. Nat. Commun. 2018, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bremnes, R.M.; Busund, L.T.; Kilver, T.L.; Andersen, S.; Richardsen, E.; Paulsen, E.E.; Hald, S.; Khanehkenari, M.R.; Cooper, W.A.; Kao, S.C.; et al. The role of tumor-infiltrating lymphocytes in development, progression, and prognosis of non-small cell lung cancer. J. Thorac. Oncol. 2016, 11, 789–800. [Google Scholar] [CrossRef] [Green Version]
- Ireson, C.R.; Alavijeh, M.S.; Palmer, A.M.; Fowler, E.R.; Jones, H.J. The role of mouse tumour models in the discovery and development of anticancer drugs. Br. J. Cancer 2019, 121. [Google Scholar] [CrossRef] [PubMed]
- Jung, J. Human tumor xenograft models for preclinical assessment of anticancer drug development. Toxicol. Res. 2014, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villalba-Orero, M.; López-Olañeta, M.M.; González-López, E.; Padrón-Barthe, L.; Gómez-Salinero, J.M.; García-Prieto, J.; Wai, T.; García-Pavía, P.; Ibáñez, B.; Jiménez-Borreguero, L.J.; et al. Lung ultrasound as a translational approach for non-invasive assessment of heart failure with reduced or preserved ejection fraction in mice. Cardiovasc. Res. 2017, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Shen, Q.; Kim, H.T.; Bissonnette, R.P.; Lamph, W.W.; Yan, B.; Brown, P.H. The rexinoid bexarotene represses cyclin D1 transcription by inducing the DEC2 transcriptional repressor. Breast Cancer Res. Treat. 2011, 128, 667–677. [Google Scholar] [CrossRef]
- Ma, F.; Liu, S.Y.; Razani, B.; Arora, N.; Li, B.; Kagechika, H.; Tontonoz, P.; Núñez, V.; Ricote, M.; Cheng, G. Retinoid X receptor α attenuates host antiviral response by suppressing type I interferon. Nat. Commun. 2014, 5, 5494. [Google Scholar] [CrossRef] [Green Version]
- Cramer, P.E.; Cirrito, R.J.; Wesson, D.W.; Lee, C.Y.D.; Karlo, C.; Zinn, A.E.; Casali, B.T.; Restivo, J.L.; Goebel, W.D.; James, M.J.; et al. ApoE-Directed Therapeutics Rapidly Clear β-Amyloid and Reverse Deficits in AD Mouse Models. Science 2012, 3335, 1503–1506. [Google Scholar] [CrossRef] [Green Version]
- Zappasodi, R.; Merghoub, T.; Wolchok, J.D. Emerging Concepts for Immune Checkpoint Blockade-Based Combination Therapies. Cancer Cell 2018, 33, 581–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, T.; Ma, Y.; Yu, L.; Jiang, J.; Shen, S.; Hou, Y.; Wang, T. Cancer immunotherapy: A focus on the regulation of immune checkpoints. Int. J. Mol. Sci. 2018, 19, 1389. [Google Scholar] [CrossRef] [Green Version]
- Beckers, R.K.; Selinger, C.I.; Vilain, R.; Madore, J.; Wilmott, J.S.; Harvey, K.; Holliday, A.; Cooper, C.L.; Robbins, E.; Gillett, D.; et al. Programmed death ligand 1 expression in triple-negative breast cancer is associated with tumour-infiltrating lymphocytes and improved outcome. Histopathology 2016, 69, 25–34. [Google Scholar] [CrossRef]
- Jones, G.S.; Baldwin, D.R. Recent advances in the management of lung cancer. Clin. Med. J. R. Coll. Physicians London 2018, 18. [Google Scholar] [CrossRef]
- Horvath, L.; Thienpont, B.; Zhao, L.; Wolf, D.; Pircher, A. Overcoming immunotherapy resistance in non-small cell lung cancer (NSCLC) - Novel approaches and future outlook. Mol. Cancer 2020, 19, 141. [Google Scholar] [CrossRef]
- Adams, J.L.; Smothers, J.; Srinivasan, R.; Hoos, A. Big opportunities for small molecules in immuno-oncology. Nat. Rev. Drug Discov. 2015, 14, 603–622. [Google Scholar] [CrossRef] [PubMed]
- Schmid, P.; Adams, S.; Rugo, H.S.; Schneeweiss, C.H.; Barrios, H.; Iwata, H.; Diéras, R.; Hegg, R.; Im, S.-A.; Shaw Wright, G.; et al. Atezolizumab and Nab-Paclitaxel in Advanced Triple-Negative Breast Cancer. N. Engl. J. Med. 2018, 379, 2108–2121. [Google Scholar] [CrossRef]
- Tanaka, A.; Sakaguchi, S. Regulatory T cells in cancer immunotherapy. Cell Res. 2017, 27, 109–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyao, T.; Floess, S.; Setoguchi, R.; Luche, H.; Fehling, H.J.; Waldmann, H.; Huehn, J.; Hori, S. Plasticity of Foxp3 + T Cells Reflects Promiscuous Foxp3 Expression in Conventional T Cells but Not Reprogramming of Regulatory T Cells. Immunity 2012, 36, 262–275. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lazarus, J.; Steele, N.G.; Yan, W.; Lee, H.J.; Nwosu, Z.C.; Halbrook, C.J.; Menjivar, R.E.; Kemp, S.B.; Sirihorachai, V.R.; et al. Regulatory T-cell depletion alters the tumor microenvironment and accelerates pancreatic carcinogenesis. Cancer Discov. 2020, 10, 422–439. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Xu, C.; Wang, B.; Niu, Q.; Su, X.; Bai, Y.; Zhu, S.; Zhao, C.; Sun, Y.; Wang, J.; et al. Single-cell transcriptomic analysis reveals disparate effector differentiation pathways in human Treg compartment. Nat. Commun. 2021, 12. [Google Scholar] [CrossRef]
- Farhood, B.; Najafi, M.; Mortezaee, K. CD8+ cytotoxic T lymphocytes in cancer immunotherapy: A review. J. Cell. Physiol. 2019, 234. [Google Scholar] [CrossRef] [PubMed]
- González-Navajas, J.M.; Fan, D.D.; Yang, S.; Yang, F.M.; Lozano-Ruiz, B.; Shen, L.; Lee, J. The Impact of Tregs on the Anticancer Immunity and the Efficacy of Immune Checkpoint Inhibitor Therapies. Front. Immunol. 2021, 12. [Google Scholar]
- Nagy, L.; Szanto, A.; Szatmari, I.; Széles, L. Nuclear hormone receptors enable macrophages and dendritic cells to sense their lipid environment and shape their immune response. Physiol. Rev. 2012, 92. [Google Scholar] [CrossRef] [Green Version]
- Geissmann, F.; Revy, P.; Brousse, N.; Lepelletier, Y.; Folli, C.; Durandy, A.; Chambon, P.; Dy, M. Retinoids regulate survival and antigen presentation by immature dendritic cells. J. Exp. Med. 2003, 198. [Google Scholar] [CrossRef] [Green Version]
- Zapata-Gonzalez, F.; Rueda, F.; Petriz, J.; Domingo, P.; Villarroya, F.; de Madariaga, A.; Domingo, J.C. 9- cis -Retinoic Acid (9cRA), a Retinoid X Receptor (RXR) Ligand, Exerts Immunosuppressive Effects on Dendritic Cells by RXR-Dependent Activation: Inhibition of Peroxisome Proliferator-Activated Receptor γ Blocks Some of the 9cRA Activities, and Precludes Them to Mature Phenotype Development. J. Immunol. 2007, 178. [Google Scholar] [CrossRef] [Green Version]
- Spranger, S.; Dai, D.; Horton, B.; Gajewski, T.F. Tumor-Residing Batf3 Dendritic Cells Are Required for Effector T Cell Trafficking and Adoptive T Cell Therapy. Cancer Cell 2017. [Google Scholar] [CrossRef] [Green Version]
- Hegde, S.; Krisnawan, V.E.; Herzog, B.H.; Zuo, C.; Breden, M.A.; Knolhoff, B.L.; Hogg, G.D.; Tang, J.P.; Baer, J.M.; Mpoy, C.; et al. Dendritic Cell Paucity Leads to Dysfunctional Immune Surveillance in Pancreatic Cancer. Cancer Cell 2020, 37, 289–307.e9. [Google Scholar] [CrossRef]
- Binnewies, M.; Mujal, A.M.; Pollack, J.L.; Combes, A.J.; Hardison, E.A.; Kevin, B.C.; Tsui, J.; Ruhland, M.K.; Kersten, K.; Abushawish, M.A.; et al. Unleashing Type-2 Dendritic Cells to Drive Protective Antitumor CD4 + T Cell Immunity. Cell 2019, 177, 556–571. [Google Scholar] [CrossRef] [PubMed]
- Mayoux, M.; Roller, A.; Pulko, V.; Sammicheli, S.; Chen, S.; Sum, E.; Jost, C.; Fransen, M.F.; Buser, R.B.; Kowanetz, M.; et al. Dendritic cells dictate responses to PD-L1 blockade cancer immunotherapy. Sci. Transl. Med. 2020, 20, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hamarsheh, S.; Groß, O.; Brummer, T.; Zeiser, R. Immune modulatory effects of oncogenic KRAS in cancer. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, S.; Thummalapalli, R.; Barbie, D.A. Inflammation as a driver and vulnerability of KRAS mediated oncogenesis. Semin. Cell Dev. Biol. 2016, 58. [Google Scholar] [CrossRef] [Green Version]
- Drosten, M.; Barbacid, M. Targeting the MAPK Pathway in KRAS-Driven Tumors. Cancer Cell 2020, 37. [Google Scholar] [CrossRef]
- Bar-Sagi, D.; Knelson, E.H.; Sequist, L.V. A bright future for KRAS inhibitors. Nat. Cancer 2020, 1. [Google Scholar] [CrossRef] [Green Version]
- Kerr, W.G.; Chisholm, J.D. The Next Generation of Immunotherapy for Cancer: Small Molecules Could Make Big Waves. J. Immunol. 2018, 202, 11–19. [Google Scholar] [CrossRef] [Green Version]
- van der Zanden, S.Y.; Luimstra, J.J.; Neefjes, J.; Borst, J.; Ovaa, H. Opportunities for Small Molecules in Cancer Immunotherapy. Trends Immunol. 2020, 41. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups Analyzed | Control | MSU42011 100 MG/KG DIET |
|---|---|---|
| # SLIDES/GROUP | 30 | 28 |
| # TUMORS/GROUP | 110 | 75 |
| TOTAL # TUMORS/SLIDE (% OF CONTROL) | 3.67 ± 0.4 (100%) | 2.68 ± 0.36 (73.1%) |
| TOTAL # L AND M GRADE (% TOTAL) | 37 (34%) | 36 (48%) (p < 0.001) |
| TOTAL # H GRADE (% TOTAL) | 73 (66%) | 39 (52%) |
| TOTAL TUMOR VOLUME, MM3 | 30.90 | 11.96 |
| AVE TUMOR SIZE (MM3)/TUMOR (% CONTROL) | 0.28 ± 0.033 (100%) | 0.16 ± 0.02 (56.8%) (p = 0.0036) |
| AVE TUMOR BURDEN (MM3) (% CONTROL) | 1.03 ± 0.15 (100%) | 0.43 ± 0.062 (41.5%) (p = 0.0015) |
| Groups Analyzed | CONTROL + ISO | MSU42011 + ISO | ANTI PD-1 AB | ANTI- PDL1 AB | MSU42011 + ANTI-PD1 | MSU42011 + ANTI-PDL1 |
|---|---|---|---|---|---|---|
| # OF SLIDES/GROUP | 28 | 30 | 30 | 30 | 28 | 30 |
| # OF TUMORS/GROUP | 127 | 98 | 110 | 90 | 91 | 81 |
| TOTAL # TUMORS/SLIDE (% OF CONTROL) | 4.54 ± 0.46 (100%) | 3.27 ± 0.4 (72%) a | 3.67 ± 0.33 (80.8%) | 3.00 ± 0.27 (66.1%) b | 3.25 ± 0.35 (71.7%) c | 2.70 ± 0.25 (59.5%) d |
| TOTAL # L AND M GRADE (% TOTAL) | 40 (32%) | 35 (36%) | 42 (38%) | 35 (39%) | 49 (54%) e, f | 42 (52%) g |
| TOTAL # H GRADE(% TOTAL) | 87 (68%) | 63 (64%) | 68 (62%) | 55 (61%) | 42 (46%) h | 39 (48%) i |
| TOTAL TUMOR VOLUME, MM3 | 28.54 | 23.536 | 32.304 | 22.48 | 12.78 | 13.01 |
| AVE TUMOR SIZE (MM3)/TUMOR (% CONTROL) | 0.22 ± 0.02 (100%) | 0.24 ± 0.023 (106.9%) | 0.29 ± 0.029 (130.7%) | 0.25 ± 0.039 (111.1%) | 0.14 ±0.014 (62.5%) j, k, l | 0.16 ± 0.016 (71.5%) m, n |
| AVE TUMOR BURDEN (MM3) | 1.02 ± 0.13 (100) | 0.78 ± 0.11 (77%) | 1.08 ± 0.17 (105.6%) | 0.75 ± 0.13 (73.5%) | 0.46 ± 0.073 (44.8%) o, p | 0.43 ± 0.061 (42.5%) q |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leal, A.S.; Moerland, J.A.; Zhang, D.; Carapellucci, S.; Lockwood, B.; Krieger-Burke, T.; Aleiwi, B.; Ellsworth, E.; Liby, K.T. The RXR Agonist MSU42011 Is Effective for the Treatment of Preclinical HER2+ Breast Cancer and Kras-Driven Lung Cancer. Cancers 2021, 13, 5004. https://doi.org/10.3390/cancers13195004
Leal AS, Moerland JA, Zhang D, Carapellucci S, Lockwood B, Krieger-Burke T, Aleiwi B, Ellsworth E, Liby KT. The RXR Agonist MSU42011 Is Effective for the Treatment of Preclinical HER2+ Breast Cancer and Kras-Driven Lung Cancer. Cancers. 2021; 13(19):5004. https://doi.org/10.3390/cancers13195004
Chicago/Turabian StyleLeal, Ana S., Jessica A. Moerland, Di Zhang, Sarah Carapellucci, Beth Lockwood, Teresa Krieger-Burke, Bilal Aleiwi, Edmund Ellsworth, and Karen T. Liby. 2021. "The RXR Agonist MSU42011 Is Effective for the Treatment of Preclinical HER2+ Breast Cancer and Kras-Driven Lung Cancer" Cancers 13, no. 19: 5004. https://doi.org/10.3390/cancers13195004
APA StyleLeal, A. S., Moerland, J. A., Zhang, D., Carapellucci, S., Lockwood, B., Krieger-Burke, T., Aleiwi, B., Ellsworth, E., & Liby, K. T. (2021). The RXR Agonist MSU42011 Is Effective for the Treatment of Preclinical HER2+ Breast Cancer and Kras-Driven Lung Cancer. Cancers, 13(19), 5004. https://doi.org/10.3390/cancers13195004