Genetic Screening for Potential New Targets in Chronic Myeloid Leukemia Based on Drosophila Transgenic for Human BCR-ABL1

 ,
,  ,
, 

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
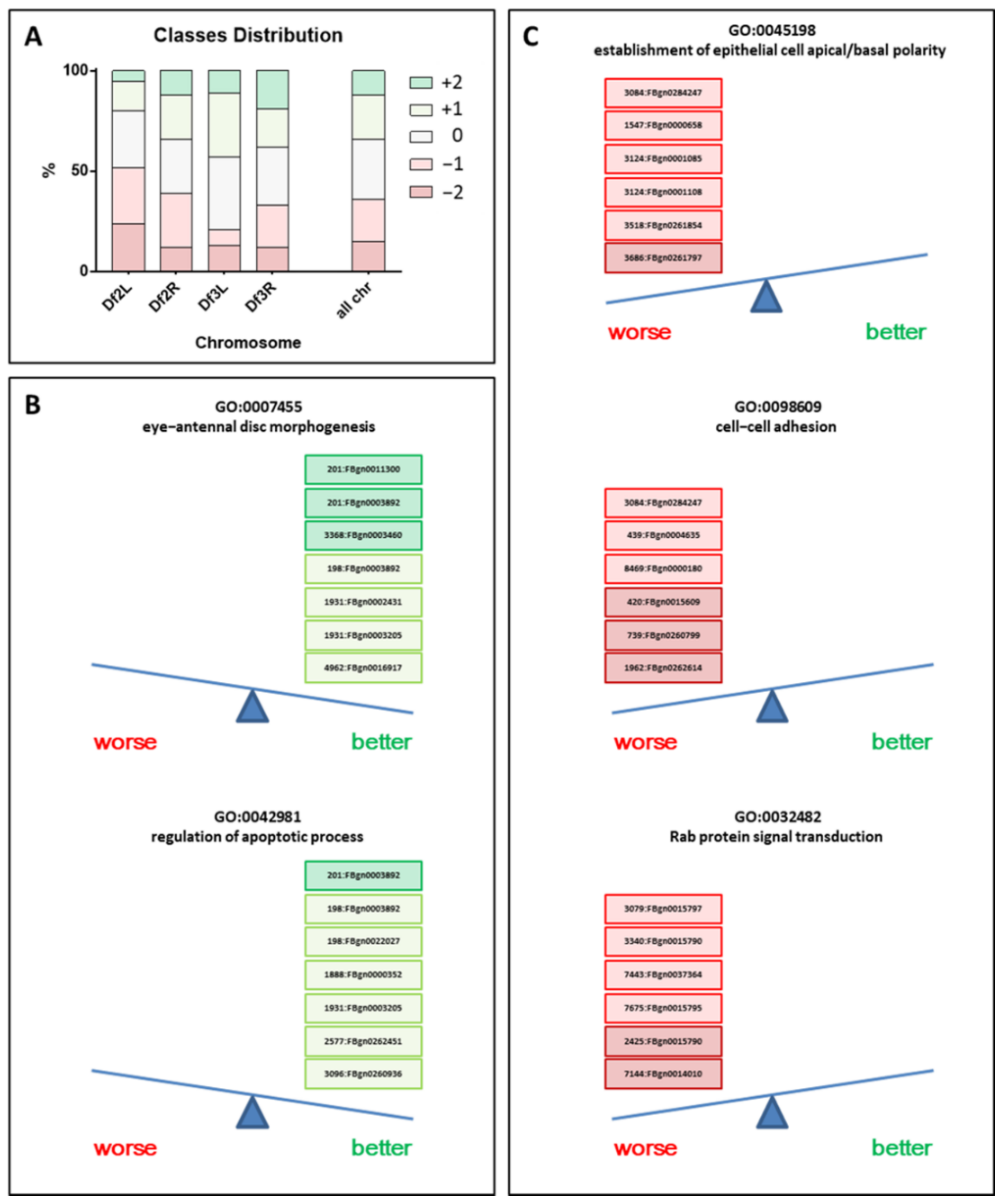
2. Results
2.1. Drosophila BCR-ABL1 Phenotype Was Altered Upon Deletion Stocks Crossing
2.2. Rab Genes as a New Potential Family Involved in Pathological BCR-ABL1 Mechanism
3. Discussion
4. Materials and Methods
4.1. Drosophila Stocks and Screening
4.2. Samples from CML Patients
4.3. Bioinformatics and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chereda, B.; Melo, J.V. Natural course and biology of CML. Ann. Hematol. 2015, 94, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Quintás-Cardama, A.; Cortes, J. Molecular biology of bcr-abl1–positive chronic myeloid leukemia. Blood 2009, 113, 1619–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, T.; Medeiros, L.J.; Hu, S. Chronic Myeloid Leukemia: Beyond BCR-ABL1. Curr. Hematol. Malign. Rep. 2018, 13, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Shuai, K.; Halpern, J.; Hoeve, J.T.; Rao, X.; Sawyers, C.L. Constitutive activation of STAT5 by the BCR-ABL oncogene in chronic myelogenous leukemia. Oncogene 1996, 13, 247–254. [Google Scholar] [PubMed]
- Jain, S.K.; Susa, M.; Keeler, M.L.; Carlesso, N.; Druker, B.; Varticovski, L. PI 3-kinase activation in BCR/abl-transformed hematopoietic cells does not require interaction of p85 SH2 domains with p210 BCR/abl. Blood 1996, 88, 1542–1550. [Google Scholar] [CrossRef] [PubMed]
- Varticovski, L.; Daley, G.Q.; Jackson, P.; Baltimore, D.; Cantley, L.C. Activation of phosphatidylinositol 3-kinase in cells expressing abl oncogene variants. Mol. Cell. Biol. 1991, 11, 1107–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birge, R.B.; Kalodimos, C.; Inagaki, F.; Tanaka, S. Crk and CrkL adaptor proteins: Networks for physiological and pathological signaling. Cell Commun. Signal. 2009, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- Wee, P.; Wang, Z. Epidermal Growth Factor Receptor Cell Proliferation Signaling Pathways. Cancers 2017, 9, 52. [Google Scholar] [CrossRef] [Green Version]
- Ilaria, R.L.; Van Etten, R.A. P210 and P190(BCR/ABL) induce the tyrosine phosphorylation and DNA binding activity of multiple specific STAT family members. J. Biol. Chem. 1996, 271, 31704–31710. [Google Scholar] [CrossRef] [Green Version]
- Crozatier, M.; Meister, M. Drosophila haematopoiesis. Cell. Microbiol. 2007, 9, 1117–1126. [Google Scholar] [CrossRef]
- Bourbon, H.-M.; Gonzy-Treboul, G.; Péronnet, F.; Alin, M.-F.; Ardourel, C.; Benassayag, C.; Cribbs, D.; Deutsch, J.; Ferrer, P.; Haenlin, M.; et al. A P-insertion screen identifying novel X-linked essential genes in Drosophila. Mech. Dev. 2002, 110, 71–83. [Google Scholar] [CrossRef]
- Gonzalez, C. Drosophila melanogaster: A model and a tool to investigate malignancy and identify new therapeutics. Nat. Rev. Cancer 2013, 13, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Osman, D.; Gobert, V.; Ponthan, F.; Heidenreich, O.; Haenlin, M.; Waltzer, L. A Drosophila model identifies calpains as modulators of the human leukemogenic fusion protein AML1-ETO. Proc. Natl. Acad. Sci. USA 2009, 106, 12043–12048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breig, O.; Bras, S.; Soria, N.M.; Osman, D.; Heidenreich, O.; Haenlin, M.; Waltzer, L. Pontin is a critical regulator for AML1-ETO-induced leukemia. Leukemia 2013, 28, 1271–1279. [Google Scholar] [CrossRef] [PubMed]
- Bernardoni, R.; Giordani, G.; Signorino, E.; Monticelli, S.; Messa, F.; Pradotto, M.; Rosso, V.; Bracco, E.; Giangrande, A.; Perini, G.; et al. A new BCR-ABL1 Drosophila model as a powerful tool to elucidate the pathogenesis and progression of chronic myeloid leukemia. Haematologica 2018, 104, 717–728. [Google Scholar] [CrossRef]
- Villavicencio, E.H.; Walterhouse, D.O.; Iannaccone, P.M. The Sonic Hedgehog–Patched–Gli Pathway in Human Development and Disease. Am. J. Hum. Genet. 2000, 67, 1047–1054. [Google Scholar] [CrossRef]
- Queiroz, K.C.S.; Ruela-De-Sousa, R.R.; Fuhler, G.M.; Aberson, H.L.; Ferreira, C.V.; Peppelenbosch, M.P.; Spek, C.A. Hedgehog signaling maintains chemoresistance in myeloid leukemic cells. Oncogene 2010, 29, 6314–6322. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Chen, A.; Jamieson, C.H.; Fereshteh, M.P.; Abrahamsson, A.; Blum, J.; Kwon, H.Y.; Kim, J.; Chute, J.P.; Rizzieri, D.A.; et al. Hedgehog signalling is essential for maintenance of cancer stem cells in myeloid leukaemia. Nat. Cell Biol. 2009, 458, 776–779. [Google Scholar] [CrossRef] [Green Version]
- Elrhman, H.E.A.; Ebian, H.F. Patched homolog 1 (PTCHI) gene mutations can predict the outcome of chronic myeloid leukemia patients? Am. J. Blood Res. 2019, 9, 15–24. [Google Scholar]
- Alonso-Dominguez, J.M.; Grinfeld, J.; Alikian, M.; Marín, D.; Reid, A.; Daghistani, M.; Hedgley, C.; Brien, S.J.O.; Clark, R.E.; Apperley, J.; et al. PTCH1expression at diagnosis predicts imatinib failure in chronic myeloid leukaemia patients in chronic phase. Am. J. Hematol. 2015, 90, 20–26. [Google Scholar] [CrossRef]
- Cea, M.; Cagnetta, A.; Cirmena, G.; Garuti, A.; Rocco, I.; Palermo, C.; Pierri, I.; Reverberi, D.; Nencioni, A.; Ballestrero, A.; et al. Tracking molecular relapse of chronic myeloid leukemia by measuring Hedgehog signaling status. Leuk. Lymphoma 2013, 54, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Long, B.; Zhu, H.-L.; Zhu, C.; Liu, T.; Meng, W. Activation of the hedgehog pathway in chronic myelogeneous leukemia patients. J. Exp. Clin. Cancer Res. 2011, 30, 8. [Google Scholar] [CrossRef] [Green Version]
- Shyamala, B.V.; Bhat, K.M. A positive role for patched-smoothened signaling in promoting cell proliferation during normal head development in Drosophila. Development 2002, 129, 1839–1847. [Google Scholar] [PubMed]
- Su, E.; Han, X.; Jiang, G. The Transforming Growth Factor Beta 1/SMAD Signaling Pathway Involved in Human Chronic Myeloid Leukemia. Tumori J. 2010, 96, 659–666. [Google Scholar] [CrossRef]
- Liu, N.; Zang, S.; Liu, Y.; Wang, Y.; Li, W.; Liu, Q.; Ji, M.; Ma, D.; Ji, C. FZD7 regulates BMSCs-mediated protection of CML cells. Oncotarget 2015, 7, 6175–6187. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Yu, F.-X. GPCR-Hippo Signaling in Cancer. Cells 2019, 8, 426. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Groffen, J.; Heisterkamp, N. Increased resistance to a farnesyltransferase inhibitor by N-cadherin expression in Bcr/Abl-P190 lymphoblastic leukemia cells. Leukemia 2007, 21, 1189–1197. [Google Scholar] [CrossRef] [Green Version]
- Mu, H.-J.; Xie, P.; Shen, Y.-F.; Jiang, Y.-Q.; Zeng, Y.-J. Cadherin-13 in primary and blast crisis chronic myeloid leukaemia: Declining expression and negative correlation with the BCR/ABL fusion gene. Br. J. Biomed. Sci. 2009, 66, 20–24. [Google Scholar] [CrossRef]
- Yuan, W.; Song, C. The Emerging Role of Rab5 in Membrane Receptor Trafficking and Signaling Pathways. Biochem. Res. Int. 2020, 2020, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Trino, S.; De Luca, L.; Simeon, V.; Laurenzana, I.; Morano, A.; Caivano, A.; La Rocca, F.; Pietrantuono, G.; Bianchino, G.; Grieco, V.; et al. Inverse regulation of bridging integrator 1 and BCR-ABL1 in chronic myeloid leukemia. Tumor Biol. 2015, 37, 217–225. [Google Scholar] [CrossRef]
- Ceresa, B.P. Regulation of EGFR endocytic trafficking by rab proteins. Histol. Histopathol. 2006, 21, 987–993. [Google Scholar] [PubMed]
- Bastin, G.; Heximer, S.P. Rab Family Proteins Regulate the Endosomal Trafficking and Function of RGS4. J. Biol. Chem. 2013, 288, 21836–21849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petiti, J.; Rosso, V.; Iacono, M.L.; Panuzzo, C.; Calabrese, C.; Signorino, E.; Pironi, L.; Cartellà, A.; Bracco, E.; Pergolizzi, B.; et al. Curcumin induces apoptosis in JAK2-mutated cells by the inhibition of JAK2/STAT and mTORC1 pathways. J. Cell. Mol. Med. 2019, 23, 4349–4357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Outa, A.; Abubaker, D.; Bazarbachi, A.; El Sabban, M.; Shirinian, M.; Nasr, R. Validation of a Drosophila model of wild-type and T315I mutated BCR-ABL1 in chronic myeloid leukemia: An effective platform for treatment screening. Haematologica 2019, 105, 387–397. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lo Iacono, M.; Signorino, E.; Petiti, J.; Pradotto, M.; Calabrese, C.; Panuzzo, C.; Caciolli, F.; Pergolizzi, B.; De Gobbi, M.; Rege-Cambrin, G.; et al. Genetic Screening for Potential New Targets in Chronic Myeloid Leukemia Based on Drosophila Transgenic for Human BCR-ABL1. Cancers 2021, 13, 293. https://doi.org/10.3390/cancers13020293
Lo Iacono M, Signorino E, Petiti J, Pradotto M, Calabrese C, Panuzzo C, Caciolli F, Pergolizzi B, De Gobbi M, Rege-Cambrin G, et al. Genetic Screening for Potential New Targets in Chronic Myeloid Leukemia Based on Drosophila Transgenic for Human BCR-ABL1. Cancers. 2021; 13(2):293. https://doi.org/10.3390/cancers13020293
Chicago/Turabian StyleLo Iacono, Marco, Elisabetta Signorino, Jessica Petiti, Monica Pradotto, Chiara Calabrese, Cristina Panuzzo, Francesca Caciolli, Barbara Pergolizzi, Marco De Gobbi, Giovanna Rege-Cambrin, and et al. 2021. "Genetic Screening for Potential New Targets in Chronic Myeloid Leukemia Based on Drosophila Transgenic for Human BCR-ABL1" Cancers 13, no. 2: 293. https://doi.org/10.3390/cancers13020293
APA StyleLo Iacono, M., Signorino, E., Petiti, J., Pradotto, M., Calabrese, C., Panuzzo, C., Caciolli, F., Pergolizzi, B., De Gobbi, M., Rege-Cambrin, G., Fava, C., Giachino, C., Bracco, E., Saglio, G., Frassoni, F., & Cilloni, D. (2021). Genetic Screening for Potential New Targets in Chronic Myeloid Leukemia Based on Drosophila Transgenic for Human BCR-ABL1. Cancers, 13(2), 293. https://doi.org/10.3390/cancers13020293