
Bovine Meat and Milk Factors (BMMFs): Their Proposed Role in Common Human Cancers and Type 2 Diabetes Mellitus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction:
- (1)
- Persistence of whole genomes or specific genes in certain cancer cells;
- (2)
- Transformed or malignant phenotype of the latter, dependent on the expression and/or function of those genes;
- (3)
- Induction of malignant growth in susceptible animal systems.
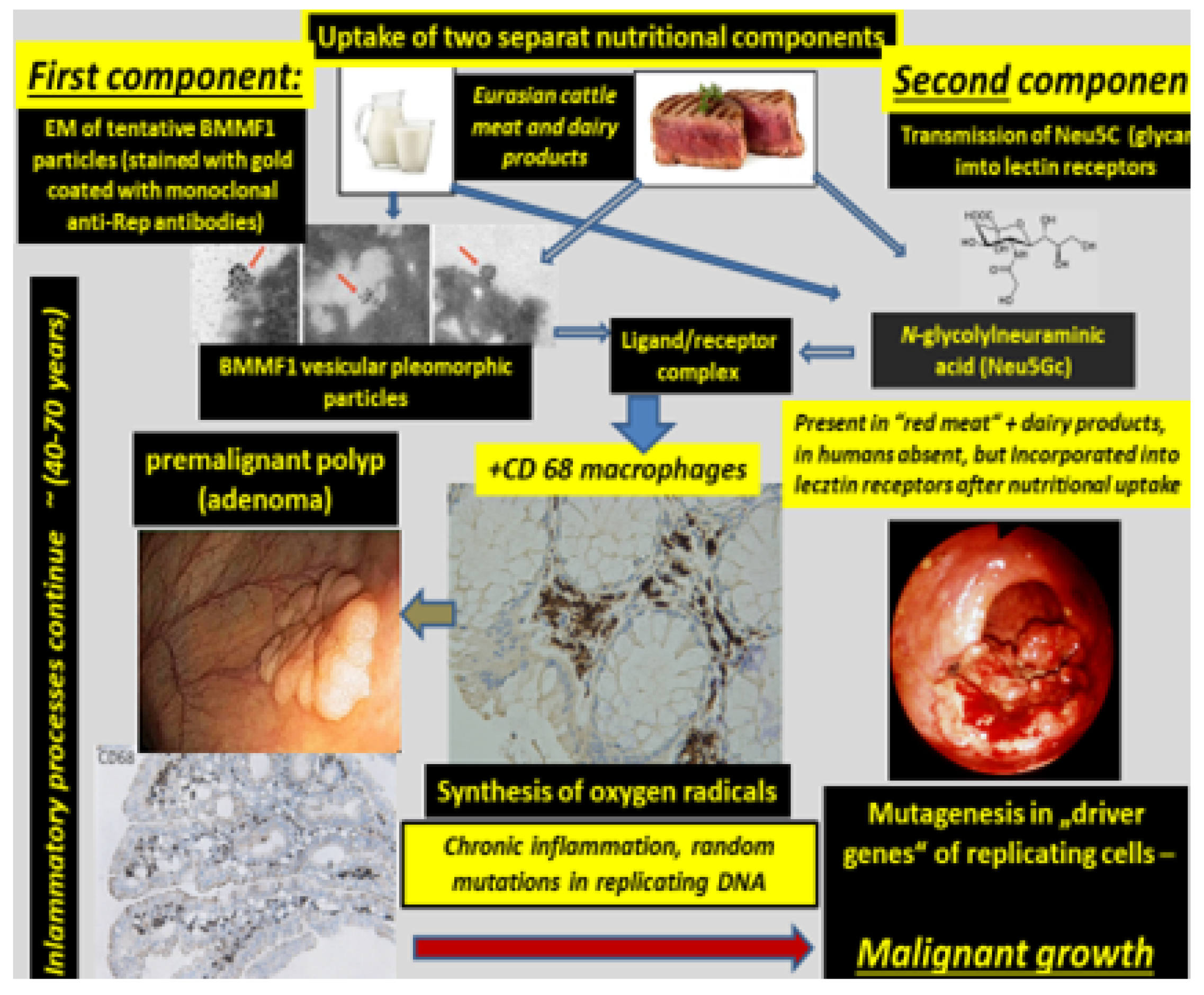
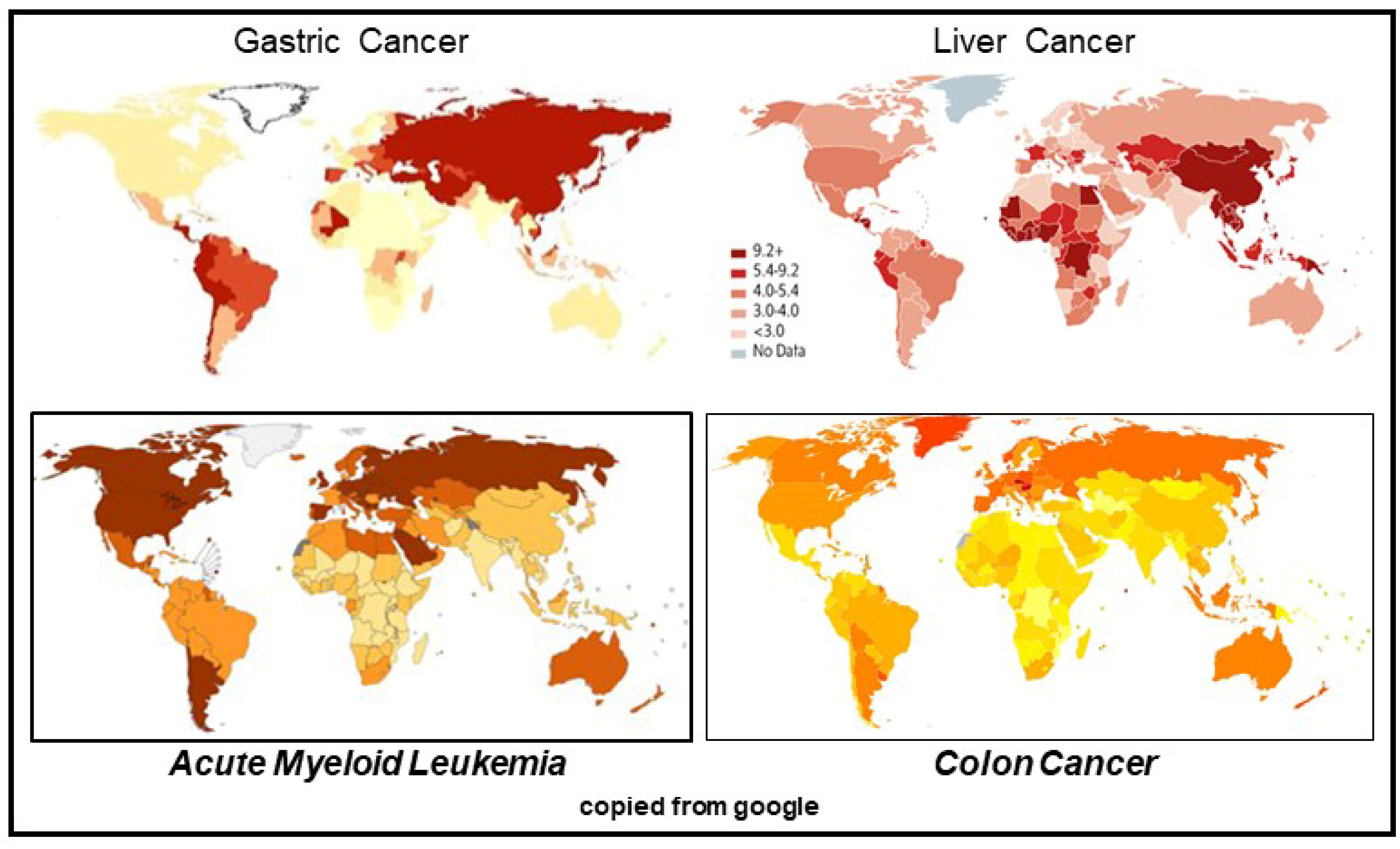
2. Bovine Meat and Milk Factors (BMMFs)
3. Potential Consequences of Indirect Carcinogenesis for Strategies in Prevention and Therapy of BMMF-Linked Cancers
- Uptake of infectious agents by nutrition through dairy products and/or meat of Eurasian cattle.
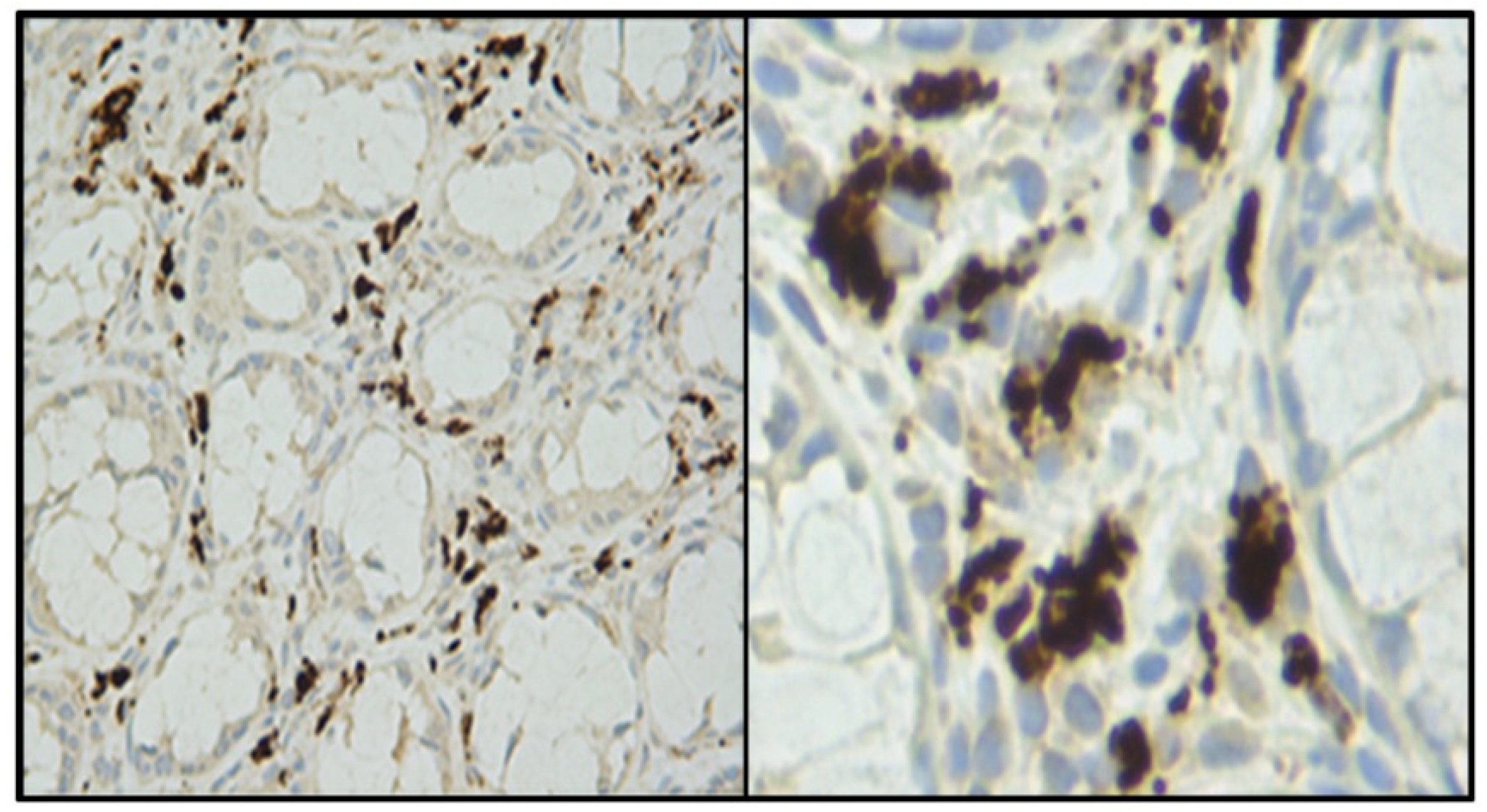
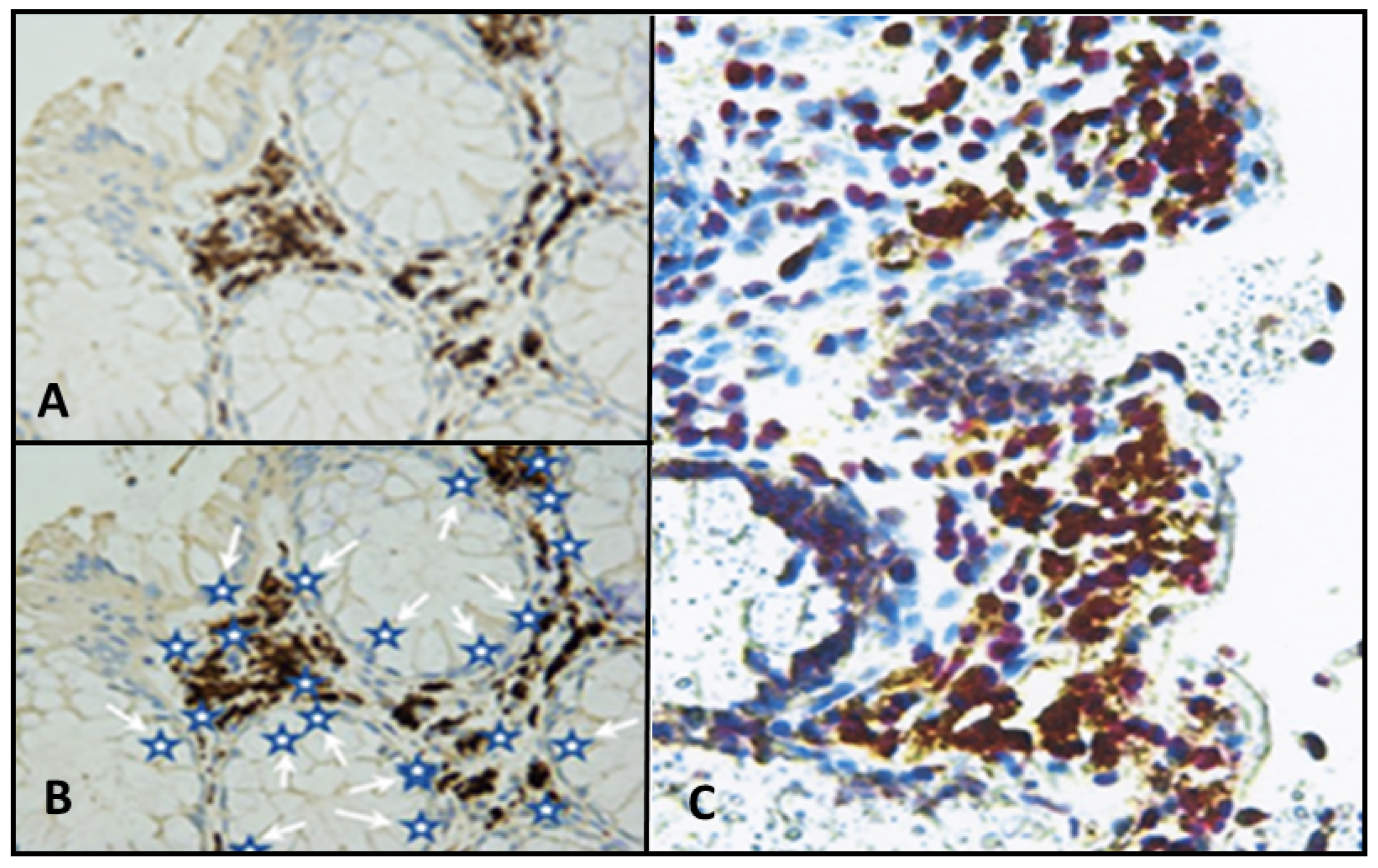
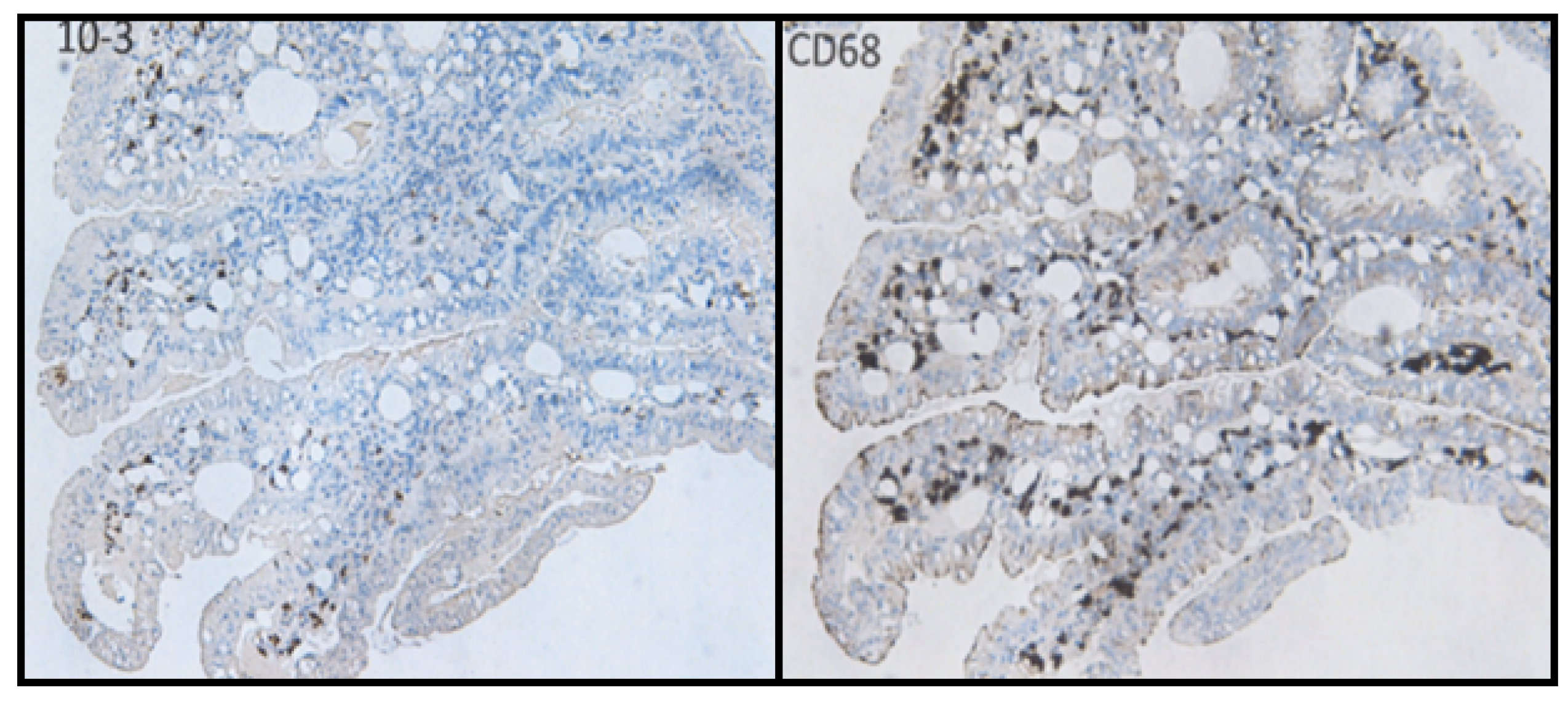
- Infection by these agents and expression of their antigens in lamina propria cells (stromal mesenchymal cells and CD68-positive macrophages).
- Macrophage-mediated inflammatory response (reactive oxygen production).
- Random mutagenesis in DNA-replicating Lieberkühn crypt cells adjacent to infected cells, as well as in replicating single-stranded BMMF-DNA.
- After long latency periods (commonly more than three decades) “driver”-mutations in genes of specific cells are established and enhanced growth of such clones occurs. These clones undergo further mutations in two or more additional steps, leading to the development of premalignant polyps. Final transformation of these polyps into malignant tumors occurs due to continuing mutagenic activity. This follows a pattern outlined previously and summarized by Greaves and Maley [26] in a quote: “Cancers evolve by a reiterative process of clonal expansion, genetic diversification and clonal selection within the adaptive landscapes of tissue ecosystems”.
- It is evident from the sequence of events described above, that BMMF DNA itself will not be present in the precursor epithelial cells, nor in their malignant progeny. No evidence exists for the frequently quoted “Hit and Run” mechanism [27,28], since infected cells persist in the lamina propria, and oxygen radical production continues. The “Hits” in colon polyps continue from adjacent BMMF-infected cells (Figure 6) [29].
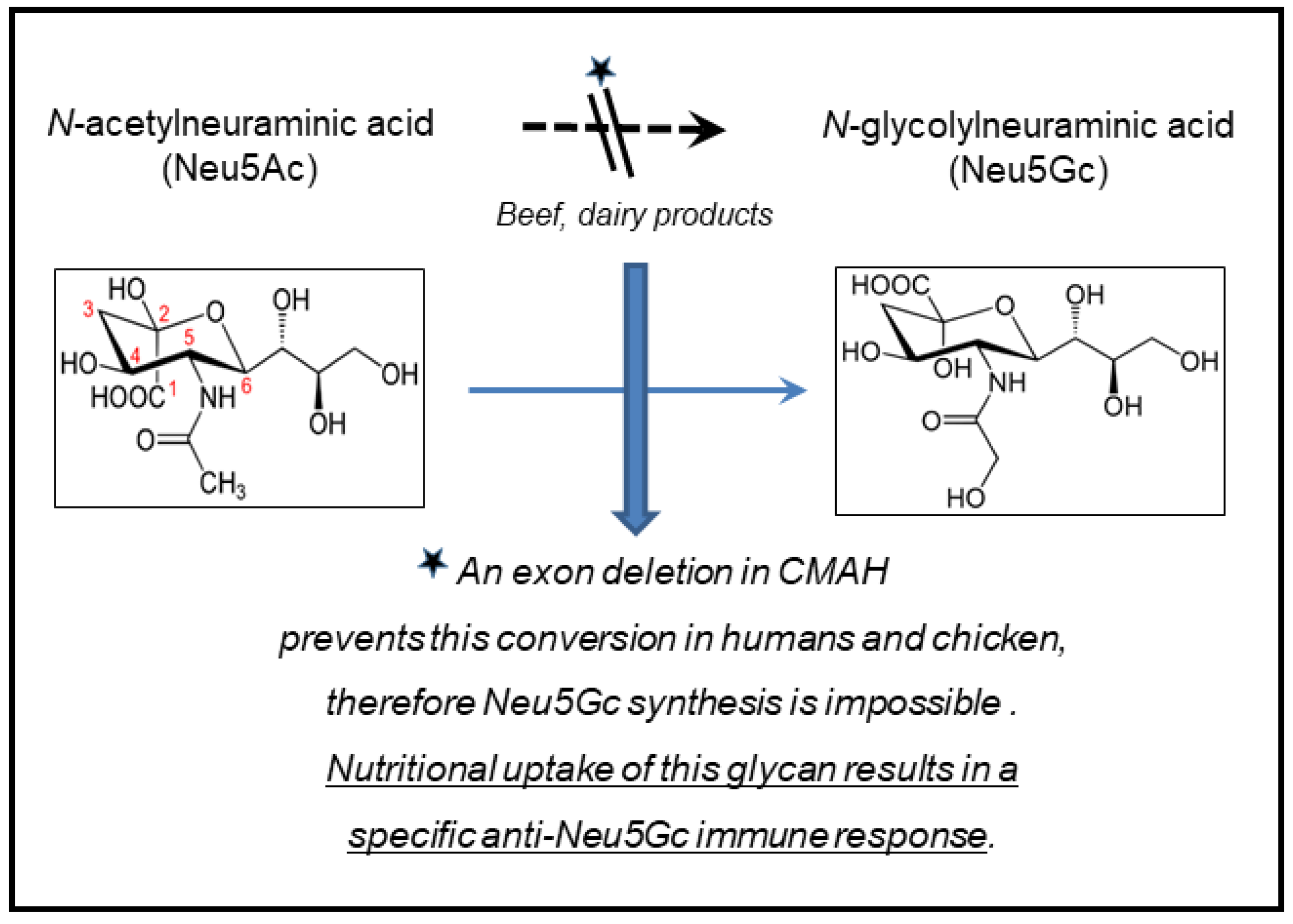
4. N-glycolylneuraminic Acid (Neu5Gc)
5. Non-Steroidal Antiphlogistica: Preventive and Immunosuppressive Events
6. Type II Diabetes Mellitus and BMMF
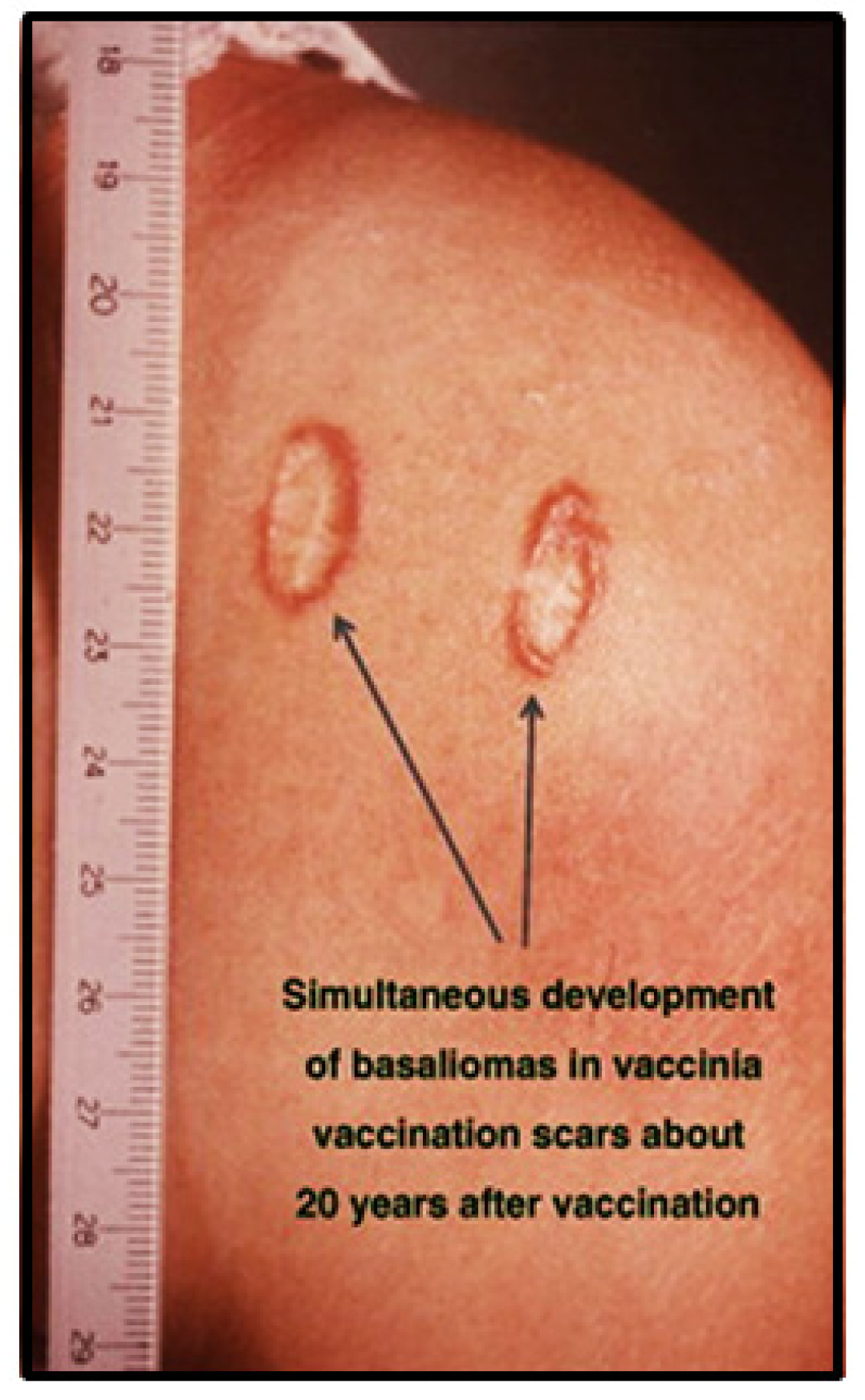
7. Basal Cell Carcinomas in Pox Vaccination Scars
8. Acute Myeloid Leukemia
- To our knowledge, this is the first direct link between a human leukemia and a specific BMMF infection;
- Since the respective BMMF types have been identified in dairy products, it is likely that the human infection was acquired by consumption of infected nutritional components;
9. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Zur Hausen, H. Infections Causing Human Cancer; Wiley-VCH: Weinheim, Germany, 2006; pp. 1–517. [Google Scholar]
- De Martel, C.; Georges, D.; Bray, F.; Ferlay, J.; Clifford, G.M. Global burden of cancer attributable to infections in 2018: A worldwide incidence analysis. Lancet Glob. Health 2020, 8, e180–e190. [Google Scholar] [CrossRef] [Green Version]
- Zur Hausen, H. Oncogenic DNA viruses. Oncogene 2001, 20, 7820–7823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zur Hausen, H.; de Villiers, E.M. Cancer “causation” by infections—Individual contributions and synergistic networks. Semin. Oncol. 2014, 41, 860–875. [Google Scholar] [CrossRef]
- Lin, S.; Li, Y.; Zamyatnin, A.A.; Werner, J.; Bazhin, A.V. Reactive oxygen species and colorectal cancer. Review J. Cell Physiol. 2018, 233, 5119–5132. [Google Scholar] [CrossRef]
- Zur Hausen, H.; Bund, T.; de Villiers, E.M. Specific nutritional infections early in life as risk factors for human colon and breast cancers several decades later. Int. J. Cancer. 2019, 144, 1574–1583. [Google Scholar] [CrossRef]
- Zur Hausen, H. The search for infectious causes of human cancers: Where and why. Virology 2009, 392, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Coghill, A.E.; Engels, E.A.; Schymura, M.J.; Mahale, P.; Shiels, M.S. Risk of breast, prostate, and colorectal cancer diagnoses among HIV-infected individuals in the United States. J. Natl. Cancer Inst. 2018, 110, 959–966. [Google Scholar] [CrossRef] [Green Version]
- De Villiers, E.M.; Gunst, K.; Chakraborty, D.; Ernst, C.; Bund, T.; zur Hausen, H. A specific class of infectious agents isolated from bovine serum and dairy products and peritumoral colon cancer tissue. Emerg. Microbes Infect. 2019, 8, 1205–1218. [Google Scholar] [CrossRef] [Green Version]
- Manuelidis, L. Nuclease resistant circular DNAs copurify with infectivity in scrapie and CJD. J. Neurovirol. 2011, 17, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Tae, H.; Karunasena, E.; Bavarva, J.H.; McIver, L.J.; Garner, H.R. Large scale comparison of non-human sequences in human sequencing data. Genomics 2014, 104, 453–458. [Google Scholar] [CrossRef]
- Chen, X.; Kost, J.; Sulovari, A.; Wong, N.; Liang, W.S.; Jian Cao, J.; Li, D. A virome-wide clonal integration analysis platform for discovering cancer viral etiology. Genome Res. 2019, 29, 819–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumata, R.; Ito, J.; Tahashi, K.; Suzuki, T.; Sato, K. A tissue level atlas of the healthy human virome. Version 2. BMC Biol. 2020, 18, 55. [Google Scholar] [CrossRef] [PubMed]
- Zapatka, M.; Borozan, I.; Brewer, D.S.; Iskar, M.; Grundhoff, A.; Alawi, M.; Desai, N.; Sültmann, H.; Moch, H.; Cooper, C.S.; et al. The landscape of viral associations in human cancers. Nat. Genet. 2020, 52, 320–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grulich, A.E.; van Leeuwen, M.T.; Falster, M.O.; Vajdic, C.M. Incidence of cancers in people with HIV/AIDS compared with immunosuppressed transplant recipients: A meta-analysis. Lancet 2007, 370, 59–67. [Google Scholar] [CrossRef]
- Zur Hausen, H.; Bund, T.; de Villiers, E.M. Infectious agents in bovine red meat and milk and their potential role in cancer and other chronic diseases. Curr. Top. Microbiol. Immunol. 2017, 407, 83–116. [Google Scholar]
- Zur Hausen, H. Red meat consumption and cancer: Reasons to suspect involvement of bovine infectious factors in colorectal cancer. Int. J. Cancer 2012, 130, 2475–2483. [Google Scholar] [CrossRef]
- Huxley, R.R.; Ansary-Moghaddam, A.; Clifton, P.; Czernichow, S.; Parr, C.L.; Woodward, M. The impact of dietary and lifestyle risk factors on risk of colorectal cancer: A quantitative overview of the epidemiological evidence. Int. J. Cancer 2009, 125, 171–180. [Google Scholar] [CrossRef] [PubMed]
- König, M.T.; Fux, R.; Link, E.; Sutter, G.; Märtlbauer, E.; Didier, A. Circular Rep-encoding single-stranded DNA sequences in milk from water buffalos (Bubalus arnee f. bubalis). Viruses 2021, 13, 1088. [Google Scholar] [CrossRef]
- Whitley, C.; Gunst, K.; Müller, H.; Funk, M.; zur Hausen, H.; de Villiers, E.M. Novel replication-competent circular DNA molecules from healthy cattle serum and milk and multiple sclerosis-affected human brain tissue. Genome Announc. 2014, 2, e00849-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funk, M.; Gunst, K.; Lucansky, V.; Müller, H.; zur Hausen, H.; de Villiers, E.M. Isolation of protein-associated circular DNA from healthy cattle serum. Genome Announc. 2014, 2, e00846-14. [Google Scholar] [CrossRef] [Green Version]
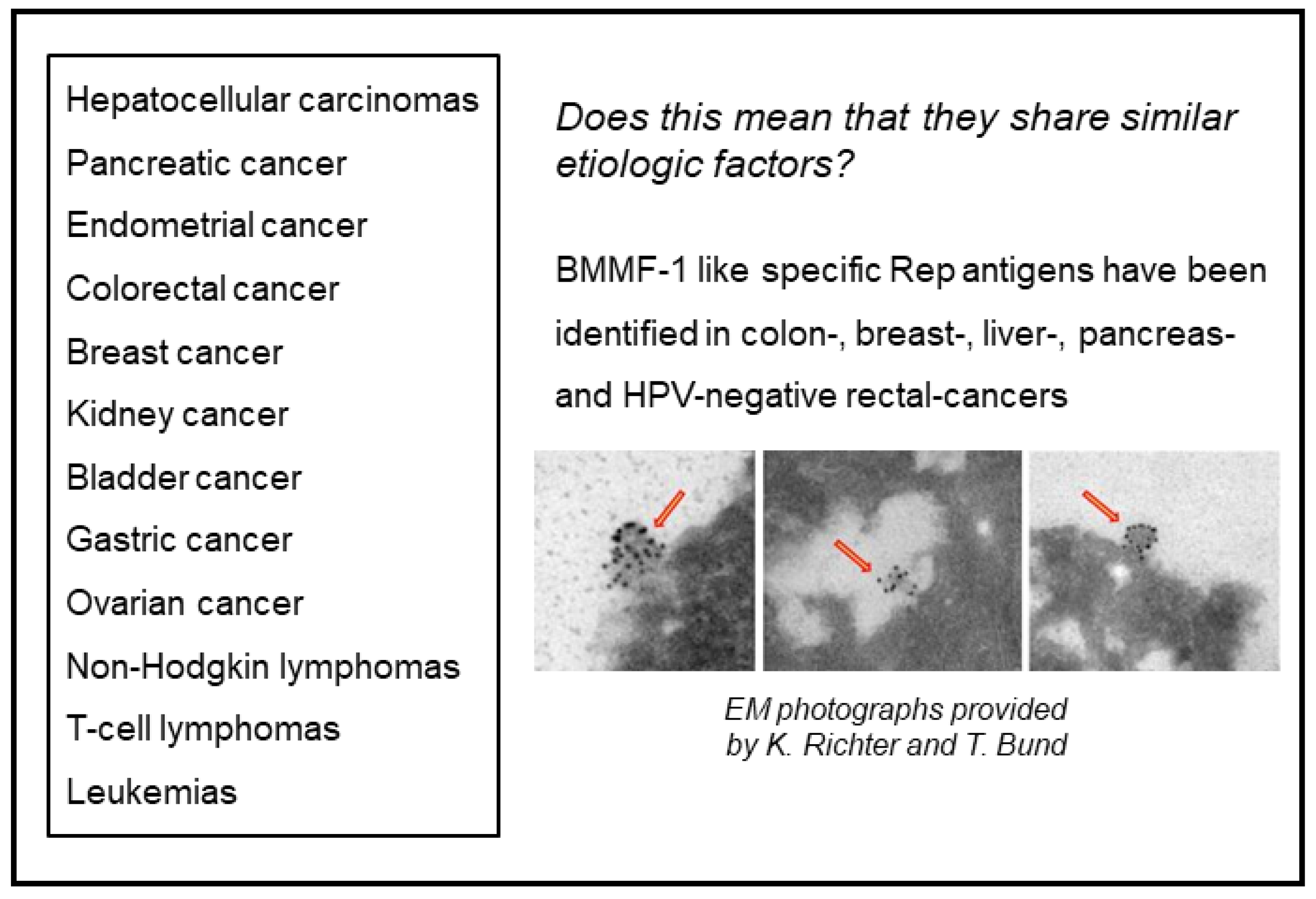
- Bund, T.; Nikitina, E.; Chakraborty, D.; Ernst, C.; Gunst, K.; Boneva, B.; Tessmer, C.; Volk, N.; Brobeil, A.; Weber, A.; et al. Analysis of chronic inflammatory lesions of the colon for BMMF Rep antigen expression and CD68 macrophage interactions. Proc. Nat. Acad. Sci. USA 2021, 118, e2025830118. [Google Scholar] [CrossRef]
- Bund, T.; de Villiers, E.M.; zur Hausen, H. Detection of BMMF antigen in breast- and prostate cancer biopsies. unpublished.
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Rev. Cell. 2010, 140, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Candido, J.; Hagemann, T. Cancer-related inflammation. J. Clin. Immunol. 2013, 1, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Greaves, M.; Maley, C.C. Clonal evolution in cancer. Nature 2012, 481, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.C.; Gadi, I.K.; Anisowicz, A.; Sager, R. Plasmid-induced “hit-and-run” tumorigenesis in Chinese hamster embryo fibroblast (CHEF) cells. Proc. Natl. Acad. Sci. USA 1985, 82, 2839–2843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarisbrick, I.A.; Rodriguez, M. Hit-Hit and hit-Run: Viruses in the playing field of multiple sclerosis. Cur. Neurol. Neurosc. Rep. 2003, 3, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Aarons, C.B.; Shanmugan, S.; Bleier, J.I. Management of malignant colon polyps: Current status and controversies. World J. Gastroenterol. 2014, 20, 16178–16183. [Google Scholar] [CrossRef] [PubMed]
- Akkerman, R.; Faas, M.M.; de Vos, P. Non-digestible carbohydrates in infant formula as substitution for human milk oligosaccharide functions: Effects on microbiota and gut maturation. Crit. Rev. Food Sci. Nutr. 2019, 59, 1486–1497. [Google Scholar] [CrossRef]
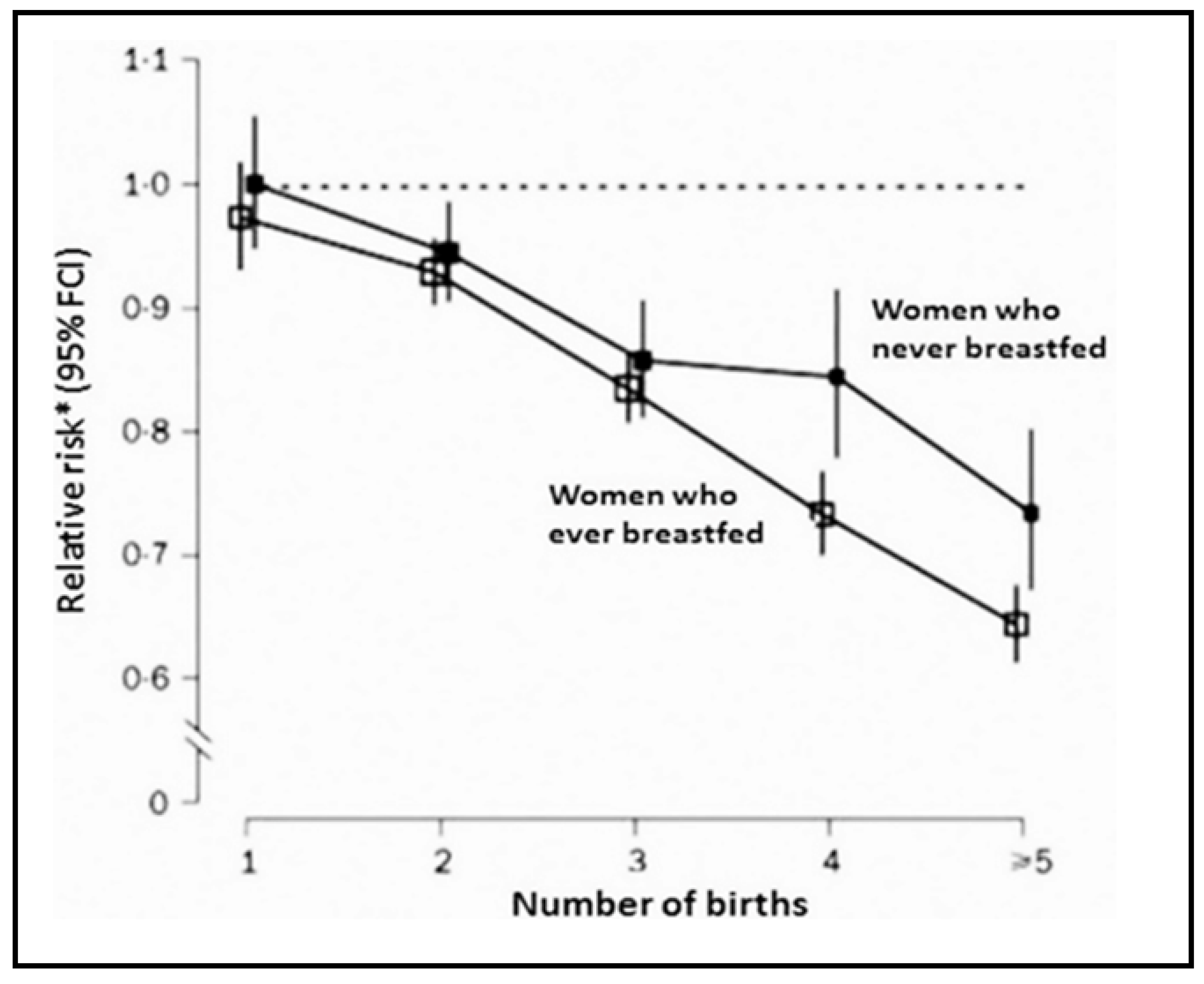
- Beral, V. Collaborative Group on Hormonal Factors in Breast Cancer. Breast cancer and breastfeeding: Collaborative reanalysis of individual data from 47 epidemiological studies in 30 countries, including 50,302 women with breast cancer and 96,973 women without the disease. Lancet 2002, 360, 187–195. [Google Scholar]
- Faupel-Badger, J.M.; Arcaro, K.F.; Balkam, J.J.; Eliassen, A.H.; Hassiotou, F.; Lebrilla, C.B.; Michels, K.B.; Palmer, J.R.; Schedin, P.; Stuebe, A.M.; et al. Postpartum remodeling, lactation, and breast cancer risk: Summary of a National Cancer Institute-sponsored workshop. J. Natl. Cancer Inst. 2013, 105, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Troisi, R.; Bjørge, T.; Gissler, M.; Grotmol, T.; Kitahara, C.M.; Myrtveit Saether, S.M.; Ording, A.G.; Sköld, C.; Sørensen, H.T.; Trabert, B.; et al. The role of pregnancy, perinatal factors and hormones in maternal cancer risk: A review of the evidence. J. Intern. Med. 2018, 283, 430–445. [Google Scholar] [CrossRef] [Green Version]
- Okerblom, J.; Varki, A. Biochemical, cellular, physiological, and pathological consequences of human loss of N-glycolylneuraminic acid. Chembiochem. 2017, 18, 1155–1171. [Google Scholar] [CrossRef] [Green Version]
- Diaz, S.L.; Padler-Karavani, V.; Ghaderi, D.; Hurtado-Ziola, N.; Yu, H.; Chen, X.; Brinkman-Van der Linden, E.C.M.; Varki, A.; Varki, N.M. Sensitive and specific detection of the non-human sialic acid N-glycolylneuraminic acid in human tissues and biotherapeutic products. PLoS ONE 2009, 4, e4241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Karin, M. Tumor-Elicited Inflammation and Colorectal Cancer. Adv. Cancer Res. 2015, 128, 173–196. [Google Scholar]
- Crusz, S.M.; Balkwill, F.R. Inflammation and cancer: Advances and new agents. Nat. Rev. Clin. Oncol. 2015, 12, 584–596. [Google Scholar] [CrossRef]
- Marelli, G.; Sica, A.; Vannucci, L.; Allavena, P. Inflammation as Target in Cancer Therapy. Curr. Opin. Pharmacol. 2017, 35, 57–65. [Google Scholar] [CrossRef]
- Harris, R.E.; Beebe-Donk, J.; Doss, H.; Burr Doss, D. Aspirin, ibuprofen, and other non-steroidal anti-inflammatory drugs in cancer prevention: A critical review of non-selective COX-2 blockade. Oncol. Rep. 2005, 13, 559–583. [Google Scholar] [CrossRef] [PubMed]
- Eddi, R.; Karki, A.; Shah, A.; DeBari, V.A.; DePasquale, J.R. Association of type 2 diabetes and colon adenomas. J. Gastrointest. Cancer. 2012, 43, 87–92. [Google Scholar] [CrossRef]
- Wang, Z.; Lai, S.T.; Xie, L.; Zhao, J.D.; Ma, N.Y.; Zhu, J.; Ren, Z.G.; Jiang, G.L. Metformin is associated with reduced risk of pancreatic cancer in patients with type 2 diabetes mellitus: A systematic review and meta-analysis. Diabetes Res. Clin. Pract. 2014, 106, 19–26. [Google Scholar] [CrossRef]
- Cho, Y.H.; Ko, B.M.; Kim, S.H.; Myung, Y.S.; Choi, J.H.; Han, J.P.; Hong, S.J.; Jeon, S.R.; Kim, H.G.; Kim, J.O.; et al. Does metformin affect the incidence of colonic polyps and adenomas in patients with type 2 diabetes mellitus? Intest. Res. 2014, 12, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Rokkas, T.; Portincasa, P. Colon neoplasia in patients with type 2 diabetes on metformin: A meta-analysis. Eur. J. Intern. Med. 2016, 33, 60–66. [Google Scholar] [CrossRef]
- Miłek, T.; Forysiński, K.; Myrcha, P.; Ciostek, P. Diabetes association of polyps and colon cancer. Pol. Przegl. Chir. 2019, 91, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wen, L.; Zhou, Q.; He, K.; Teng, L. Preventative and therapeutic effects of metformin in gastric cancer: A new contribution of an old friend. Cancer Manag. Res. 2020, 12, 8545–8554. [Google Scholar] [CrossRef]
- Gallagher, E.J.; LeRoith, D. Does a single nucleotide polymorphism in the FGFR explain the connection between diabetes and cancer? Cell Metab. 2013, 17, 808–809. [Google Scholar] [CrossRef] [Green Version]
- Yan, P.; Wang, Y.; Fu, T.; Liu, Y.; Zhang, Z.-Y. The association between type 1 and 2 diabetes mellitus and the risk of leukemia: A systematic review and meta-analysis of 18 cohort studies. Endocr. J. 2021, 68, 281–289. [Google Scholar] [CrossRef]
- Vajdic, C.M.; Grulich, A.E.; Kaldor, J.M.; Fritschi, L.; Benke, G.; Hughes, A.M.; Kricker, A.; Turner, J.J.; Milliken, S.; Armstrong, B.K. Specific infections, infection-related behavior, and risk of non-Hodgkin lymphoma in adults. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1102–1108. [Google Scholar] [CrossRef] [Green Version]
- Hedlund, M.; Tangvoranuntakul, P.; Takematsu, H.; Long, J.M.; Housley, G.D.; Kozutsumi, Y.; Suzuki, A.; Wynshaw-Boris, A.; Ryan, A.F.; Gallo, R.L.; et al. N-glycolylneuraminic acid deficiency in mice: Implications for human biology and evolution. Mol. Cell Biol. 2007, 27, 4340–4346. [Google Scholar] [CrossRef] [Green Version]
- Khalesi, M.; Whiteman, D.C.; Doi, S.A.; Clark, J.; Kimlin, M.G.; Neale, R.E. Cutaneous markers of photo-damage and risk of Basal cell carcinoma of the skin: A meta-analysis. Cancer Epidemiol. Biomarkers Prev. 2013, 22, 1483–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haefele, L.; Feuerbach, L.; Bund, T. RNA-Seq analyses. Unpublished.
- Zur Hausen, H.; de Villiers, E.M. Virus target cell conditioning model to explain some epidemiologic characteristics of childhood leukemias and lymphomas. Int. J. Cancer. 2005, 115, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Zur Hausen, H. Childhood leukemias and other hematopoietic malignancies: Interdependence between an infectious event and chromosomal modifications. Int. J. Cancer 2009, 125, 1764–1770. [Google Scholar] [CrossRef] [PubMed]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Villiers, E.-M.; zur Hausen, H. Bovine Meat and Milk Factors (BMMFs): Their Proposed Role in Common Human Cancers and Type 2 Diabetes Mellitus. Cancers 2021, 13, 5407. https://doi.org/10.3390/cancers13215407
de Villiers E-M, zur Hausen H. Bovine Meat and Milk Factors (BMMFs): Their Proposed Role in Common Human Cancers and Type 2 Diabetes Mellitus. Cancers. 2021; 13(21):5407. https://doi.org/10.3390/cancers13215407
Chicago/Turabian Stylede Villiers, Ethel-Michele, and Harald zur Hausen. 2021. "Bovine Meat and Milk Factors (BMMFs): Their Proposed Role in Common Human Cancers and Type 2 Diabetes Mellitus" Cancers 13, no. 21: 5407. https://doi.org/10.3390/cancers13215407
APA Stylede Villiers, E. -M., & zur Hausen, H. (2021). Bovine Meat and Milk Factors (BMMFs): Their Proposed Role in Common Human Cancers and Type 2 Diabetes Mellitus. Cancers, 13(21), 5407. https://doi.org/10.3390/cancers13215407