
The Antiviral Drug Efavirenz in Breast Cancer Stem Cell Therapy

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Cell Cultures and Drug Treatments
2.2. XTT Cell Viability Assay
2.3. Immunofluorescence (IF)
2.4. Cancer Stem Cell (CSC) Flow Cytometry
2.5. mRNA-Seq Gene Expression Profiling
2.6. microRNA Expression Taqman Assays and microRNA Profiling by Microarrays
3. Results
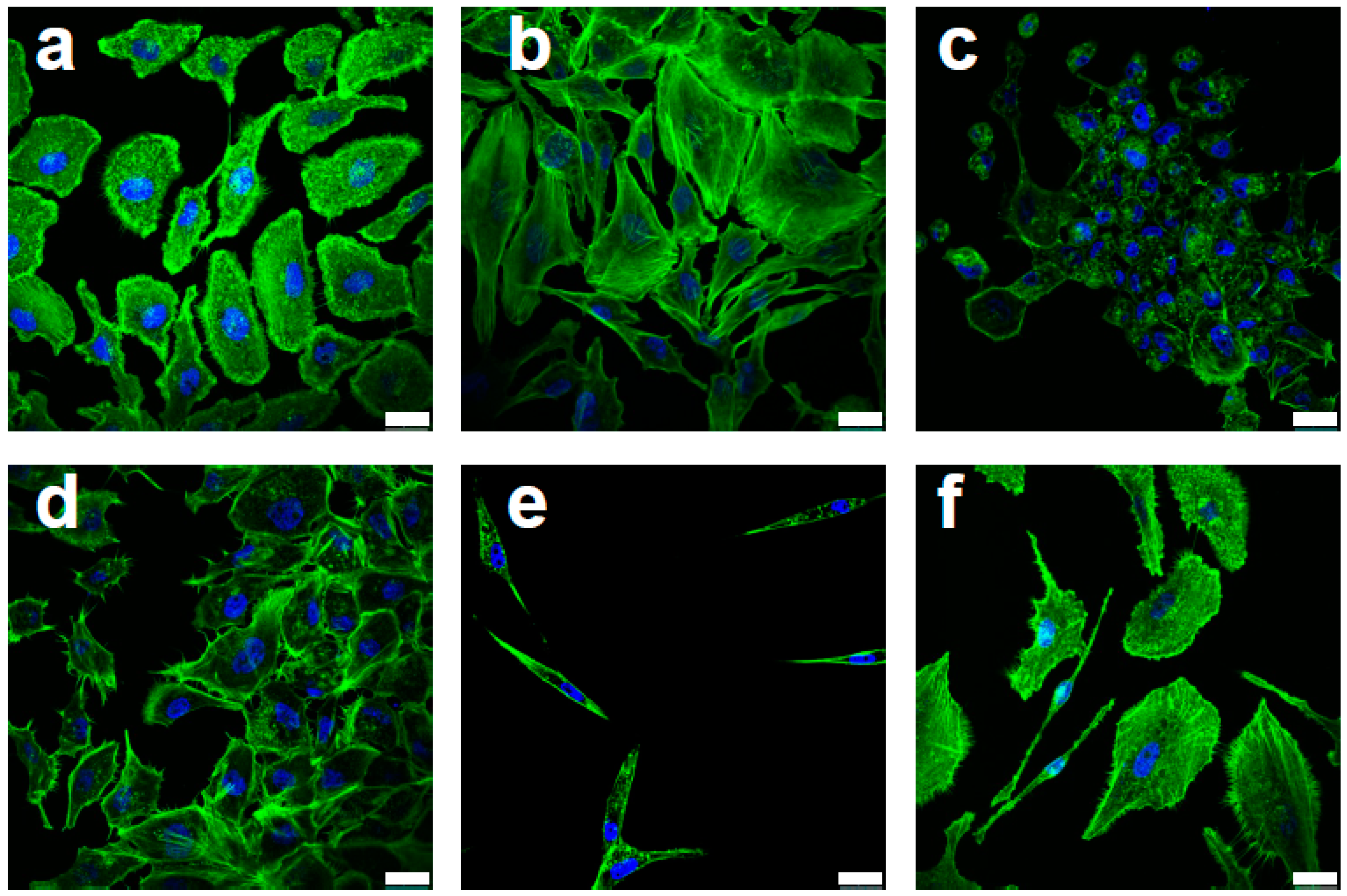
3.1. Changes in Cancer Stem Cell Regulators/Indicators Observed in Efavirenz-Treated Breast Cancer Cells
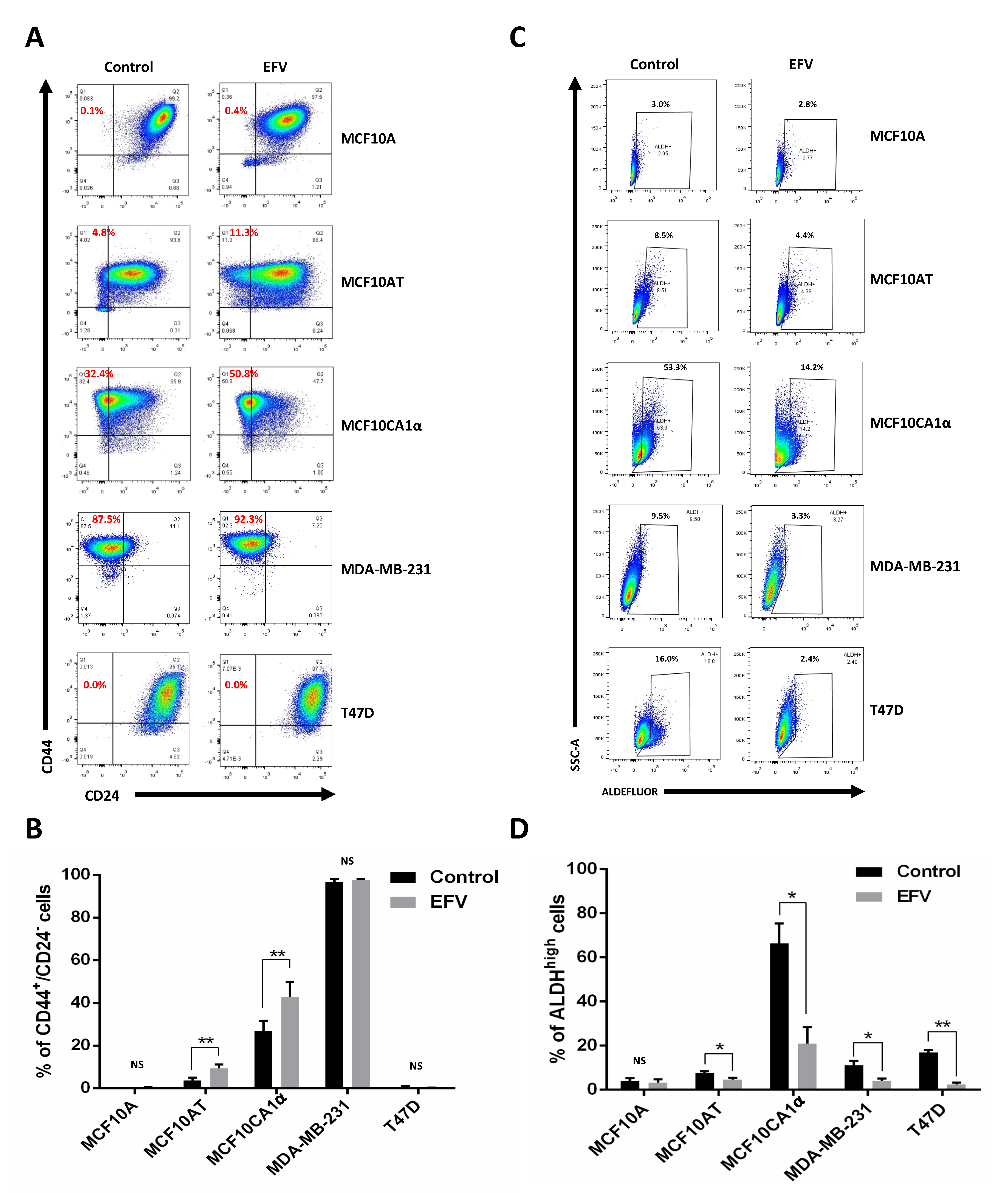
3.2. Breast CSC Population Are Altered by Efavirenz Treatment
3.3. Efavirenz Can Effectively Reduce Functional Breast CSCs
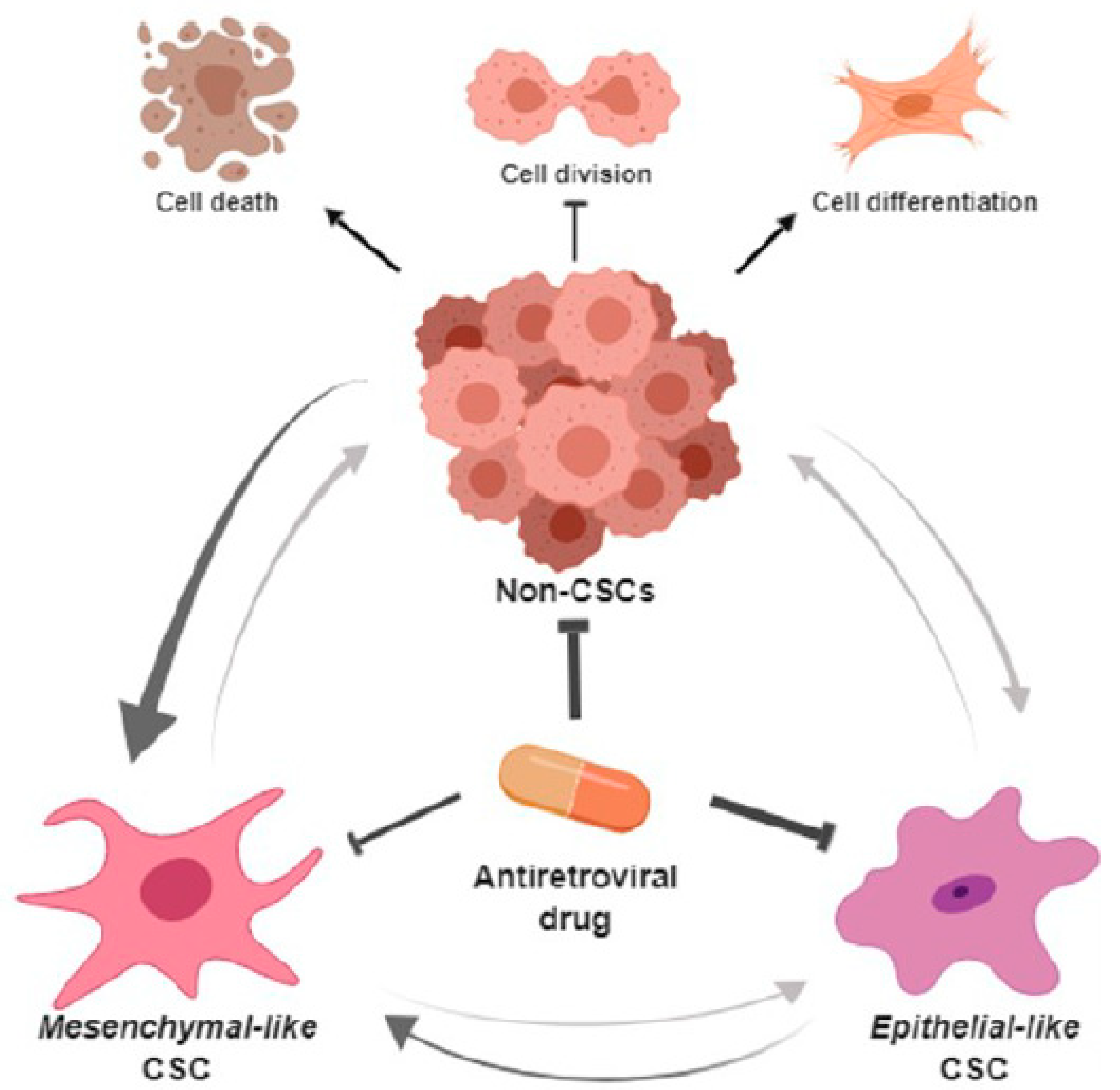
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Batlle, E.; Clevers, H. Cancer stem cells revisited. Nat. Med. 2017, 23, 1124–1134. [Google Scholar] [CrossRef]
- Clevers, H. The cancer stem cell: Premises, promises and challenges. Nat. Med. 2011, 17, 313–319. [Google Scholar] [CrossRef]
- Lapidot, T.; Sirard, C.; Vormoor, J.; Murdoch, B.; Hoang, T.; Caceres-Cortes, J.; Minden, M.; Paterson, B.; Caligiuri, M.A.; Dick, J.E. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature 1994, 367, 645–648. [Google Scholar] [CrossRef]
- Iseghohi, S.O. Cancer stem cells may contribute to the difficulty in treating cancer. Genes Dis. 2016, 3, 7–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charafe-Jauffret, E.; Ginestier, C.; Birnbaum, D. Breast cancer stem cells: Tools and models to rely on. BMC Cancer 2009, 9, 202. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Dong, J.; Haiech, J.; Kilhoffer, M.C.; Zeniou, M. Cancer Stem Cell Quiescence and Plasticity as Major Challenges in Cancer Therapy. Stem Cells Int. 2016, 2016, 1740936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kai, K.; Arima, Y.; Kamiya, T.; Saya, H. Breast cancer stem cells. Breast Cancer 2010, 17, 80–85. [Google Scholar] [CrossRef]
- Snyder, V.; Reed-Newman, T.C.; Arnold, L.; Thomas, S.M.; Anant, S. Cancer Stem Cell Metabolism and Potential Therapeutic Targets. Front. Oncol. 2018, 8, 203. [Google Scholar] [CrossRef] [PubMed]
- Eun, K.; Ham, S.W.; Kim, H. Cancer stem cell heterogeneity: Origin and new perspectives on CSC targeting. BMB Rep. 2017, 50, 117–125. [Google Scholar] [CrossRef]
- Patel, A.P.; Tirosh, I.; Trombetta, J.J.; Shalek, A.K.; Gillespie, S.M.; Wakimoto, H.; Cahill, D.P.; Nahed, B.V.; Curry, W.T.; Martuza, R.L.; et al. Single-cell RNA-seq highlights intratumoral heterogeneity in primary glioblastoma. Science 2014, 344, 1396–1401. [Google Scholar] [CrossRef] [Green Version]
- Mcdermott, S.P.; Wicha, M.S. Targeting breast cancer stem cells. Mol. Oncol. 2010, 4, 404–419. [Google Scholar] [CrossRef] [Green Version]
- Velasco-Velazquez, M.A.; Homsi, N.; De La Fuente, M.; Pestell, R.G. Breast cancer stem cells. Int. J. Biochem. Cell Biol. 2012, 44, 573–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginestier, C.; Hur, M.H.; Charafe-Jauffret, E.; Monville, F.; Dutcher, J.; Brown, M.; Jacquemier, J.; Viens, P.; Kleer, C.G.; Liu, S.; et al. ALDH1 Is a Marker of Normal and Malignant Human Mammary Stem Cells and a Predictor of Poor Clinical Outcome. Cell Stem Cell 2007, 1, 555–567. [Google Scholar] [CrossRef] [Green Version]
- Ponti, D.; Costa, A.; Zaffaroni, N.; Pratesi, G.; Petrangolini, G.; Coradini, D.; Pilotti, S.; Pierotti, M.A.; Daidone, M.G. Isolation and In vitro Propagation of Tumorigenic Breast Cancer Cells with Stem/Progenitor Cell Properties. Cancer Res. 2005, 65, 5506–5511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saygin, C.; Matei, D.; Majeti, R.; Reizes, O.; Lathia, J.D. Targeting Cancer Stemness in the Clinic: From Hype to Hope. Cell Stem Cell 2019, 24, 25–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simard, E.P.; Engels, E.A. Cancer as a Cause of Death among People with AIDS in the United States. Clin. Infect. Dis. 2010, 51, 957–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sciamanna, I.; Landriscina, M.; Pittoggi, C.; Quirino, M.; Mearelli, C.; Beraldi, R.; Mattei, E.; Serafino, A.; Cassano, A.; Sinibaldi-Vallebona, P.; et al. Inhibition of endogenous reverse transcriptase antagonizes human tumor growth. Oncogene 2005, 24, 3923–3931. [Google Scholar] [CrossRef] [Green Version]
- Sciamanna, I.; Sinibaldi-Vallebona, P.; Serafino, A.; Spadafora, C. LINE-1-encoded reverse Transcriptase as a target in cancer therapy. Front. Biosci. 2018, 23, 1360–1369. [Google Scholar]
- Agliano, A.; Calvo, A.; Box, C. The challenge of targeting cancer stem cells to halt metastasis. Semin. Cancer Biol. 2017, 44, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Bastos, M.M.; Costa, C.C.P.; Bezerra, T.C.; Da Silva, F.C.; Boechat, N. Efavirenz a nonnucleoside reverse transcriptase inhibitor of first-generation: Approaches based on its medicinal chemistry. Eur. J. Med. Chem. 2016, 108, 455–465. [Google Scholar] [CrossRef]
- Rangasamy, D.; Lenka, N.; Ohms, S.; Dahlstrom, J.E.; Blackburn, A.C.; Board, P.G. Activation of LINE-1 Retrotransposon Increases the Risk of Epithelial-Mesenchymal Transition and Metastasis in Epithelial Cancer. Curr. Mol. Med. 2015, 15, 588–597. [Google Scholar] [CrossRef] [Green Version]
- Chiou, P.; Ohms, S.; Board, P.G.; Dahlstrom, J.E.; Rangasamy, D.; Casarotto, M.G. Efavirenz as a potential drug for the treatment of triple-negative breast cancers. Clin. Transl. Oncol. 2021, 23, 353–363. [Google Scholar] [CrossRef]
- Grange, C.; Lanzardo, S.; Cavallo, F.; Camussi, G.; Bussolati, B. Sca-1 identifies the tumor-initiating cells in mammary tumors of BALB-neuT transgenic mice. Neoplasia 2008, 10, 1433–1443. [Google Scholar] [CrossRef] [Green Version]
- Ohms, S.; Rangasamy, D. Silencing of LINE-1 retrotransposons contributes to variation in small noncoding RNA expression in human cancer cells. Oncotarget 2014, 5, 4103–4117. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, H.; Wyckoff, J.; Condeelis, J. Cell migration in tumors. Curr. Opin. Cell Biol. 2005, 17, 559–564. [Google Scholar] [CrossRef]
- Tamura, S.; Isobe, T.; Ariyama, H.; Nakano, M.; Kikushige, Y.; Takaishi, S.; Kusaba, H.; Takenaka, K.; Ueki, T.; Nakamura, M.; et al. E-cadherin regulates proliferation of colorectal cancer stem cells through NANOG. Oncol. Rep. 2018, 40, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Garg, M. Epithelial plasticity and cancer stem cells: Major mechanisms of cancer pathogenesis and therapy resistance. World J. Stem Cells 2017, 9, 118–126. [Google Scholar] [CrossRef]
- Park, J.; Schwarzbauer, J.E. Mammary epithelial cell interactions with fibronectin stimulate epithelial-mesenchymal transition. Oncogene 2014, 33, 1649–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekar, D.; Krishnan, R.; Panagal, M.; Sivakumar, P.; Gopinath, V.; Basam, V. Deciphering the role of microRNA 21 in cancer stem cells (CSCs). Genes Dis. 2016, 3, 277–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, T.; Wang, W.; Zhang, B.; Xu, Y.; Chen, L.; Pan, S.; Hu, H.; Geng, Q. Regulatory mechanisms of microRNAs in lung cancer stem cells. SpringerPlus 2016, 5, 1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, B.; Wang, W.; Meng, X.-x.; Du, G.; Li, J.; Zhang, S.-z.; Zhou, B.-h.; Fu, Z.-h. Let-7 inhibits self-renewal of hepatocellular cancer stem-like cells through regulating the epithelial-mesenchymal transition and the Wnt signaling pathway. BMC Cancer 2016, 16, 863. [Google Scholar] [CrossRef] [Green Version]
- Thammaiah, C.K.; Jayaram, S. Role of let-7 family microRNA in breast cancer. Non-Coding RNA Res. 2016, 1, 77–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang-Verslues, W.W.; Kuo, W.-H.; Chang, P.-H.; Pan, C.-C.; Wang, H.-H.; Tsai, S.-T.; Jeng, Y.-M.; Shew, J.-Y.; Kung, J.T.; Chen, C.-H.; et al. Multiple Lineages of Human Breast Cancer Stem/Progenitor Cells Identified by Profiling with Stem Cell Markers. PLoS ONE 2009, 4, e8377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewi, D.L.; Ishii, H.; Kano, Y.; Nishikawa, S.; Haraguchi, N.; Sakai, D.; Satoh, T.; Doki, Y.; Mori, M. Cancer stem cell theory in gastrointestinal malignancies: Recent progress and upcoming challenges. J. Gastroenterol. 2011, 46, 1145. [Google Scholar] [CrossRef] [PubMed]
- Alison, M.R.; Guppy, N.J.; Lim, S.M.; Nicholson, L.J. Finding cancer stem cells: Are aldehyde dehydrogenases fit for purpose? J. Pathol. 2010, 222, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Faronato, M.; Nguyen, V.T.; Patten, D.K.; Lombardo, Y.; Steel, J.H.; Patel, N.; Woodley, L.; Shousha, S.; Pruneri, G.; Coombes, R.C.; et al. DMXL2 drives epithelial to mesenchymal transition in hormonal therapy resistant breast cancer through Notch hyper-activation. Oncotarget 2015, 6, 22467–22479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Jiang, D.; Chen, Y.; Wei, L.; Zhang, S.; Zhao, F.; Ni, R.; Lu, C.; Wan, C. High CHMP4B expression is associated with accelerated cell proliferation and resistance to doxorubicin in hepatocellular carcinoma. Tumour Biol. 2015, 36, 2569–2581. [Google Scholar] [CrossRef] [PubMed]
- Padanad, M.S.; Konstantinidou, G.; Venkateswaran, N.; Melegari, M.; Rindhe, S.; Mitsche, M.; Yang, C.; Batten, K.; Huffman, K.E.; Liu, J.; et al. Fatty Acid Oxidation Mediated by Acyl-CoA Synthetase Long Chain 3 Is Required for Mutant KRAS Lung Tumorigenesis. Cell Rep. 2016, 16, 1614–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tirinato, L.; Pagliari, F.; Limongi, T.; Marini, M.; Falqui, A.; Seco, J.; Candeloro, P.; Liberale, C.; Di Fabrizio, E. An Overview of Lipid Droplets in Cancer and Cancer Stem Cells. Stem Cells Int. 2017, 2017, 1656053. [Google Scholar] [CrossRef]
- Bielecka, Z.F.; Maliszewska-Olejniczak, K.; Safir, I.J.; Szczylik, C.; Czarnecka, A.M. Three-dimensional cell culture model utilization in cancer stem cell research. Biol. Rev. 2017, 92, 1505–1520. [Google Scholar] [CrossRef]
- Fillmore, C.M.; Kuperwasser, C. Human breast cancer cell lines contain stem-like cells that self-renew, give rise to phenotypically diverse progeny and survive chemotherapy. Breast Cancer Res. 2008, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Mendez-Lopez, M.; Sutter, T.; Driessen, C.; Besse, L. HIV protease inhibitors for the treatment of multiple myeloma. Clin. Adv. Hematol. Oncol. 2019, 17, 615–623. [Google Scholar] [PubMed]
- Liu, H.; Lv, L.; Yang, K. Chemotherapy targeting cancer stem cells. Am. J. Cancer Res. 2015, 5, 880–893. [Google Scholar] [PubMed]
- Wu, X.; Zou, X.; Chang, Q.; Zhang, Y.; Li, Y.; Zhang, L.; Huang, J.; Liang, B. The evolutionary pattern and the regulation of stearoyl-CoA desaturase genes. Biomed Res. Int. 2013, 2013, 856521. [Google Scholar] [CrossRef] [Green Version]
- Hecht, M.; Harrer, T.; Büttner, M.; Schwegler, M.; Erber, S.; Fietkau, R.; Distel, L.V. Cytotoxic effect of efavirenz is selective against cancer cells and associated with the cannabinoid system. AIDS 2013, 27, 2031–2040. [Google Scholar] [CrossRef] [PubMed]
- Hecht, M.; Harrer, T.; Korber, V.; Sarpong, E.O.; Moser, F.; Fiebig, N.; Schwegler, M.; Sturzl, M.; Fietkau, R.; Distel, L.V. Cytotoxic effect of Efavirenz in BxPC-3 pancreatic cancer cells is based on oxidative stress and is synergistic with ionizing radiation. Oncol. Lett. 2018, 15, 1728–1736. [Google Scholar] [CrossRef] [PubMed]
- De Cecco, M.; Ito, T.; Petrashen, A.P.; Elias, A.E.; Skvir, N.J.; Criscione, S.W.; Caligiana, A.; Brocculi, G.; Adney, E.M.; Boeke, J.D.; et al. L1 drives IFN in senescent cells and promotes age-associated inflammation. Nature 2019, 566, 73–78. [Google Scholar] [CrossRef]
- Jenike, A.E.; Halushka, M.K. miR-21: A non-specific biomarker of all maladies. Biomark. Res. 2021, 9, 18. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Names | Fold Change | P-Value | References |
|---|---|---|---|
| PROCR | 1.777661 | 7.99 × 10−4 | [33] |
| MED8 | 1.31992 | 9.71 × 10−4 | [34] |
| DMXL2 | 1.161448 | 3.93 × 10−3 | [35,36] |
| CHMP4B | −1.33233 | 1.85 × 10−4 | [37] |
| ACSL3 | −1.73666 | 3.04 × 10−4 | [38] |
| FASN | −1.75391 | 2.82 × 10−4 | [39] |
| SCD | −4.34746 | 2.48 × 10−3 | [39] |
| Mature miRNA Sequence of the Target miRNAs used in microRNA qRT-PCR Target microRNA | Assay ID | Mature miRNA Sequence |
|---|---|---|
| miR-21 | rno481342_mir | UAGCUUAUCAGACUGAUGUUGA |
| miR-182 | 477935_mir | UUUGGCAAUGGUAGAACUCACACU |
| miR-423 | 478090_mir | UGAGGGGCAGAGAGCGAGACUUU |
| MCF10A | MCF10AT | MCF10CA1α | MDA-MB-231 | T47D | ||
|---|---|---|---|---|---|---|
| Epithelial-like CSCs | Control | 4.08 ± 1.17% | 7.56 ± 0.71% | 66.30 ± 8.97% | 11.06 ± 1.96% | 16.76 ± 1.17% |
| EFV | 3.13 ± 1.50% | 4.53 ± 0.79% * | 20.76 ± 7.51% * | 3.87 ± 1.05% * | 2.32 ± 0.86% ** | |
| Mesenchymal-like CSCs | Control | 0.05 ± 0.03% | 3.66 ± 1.14% | 26.77 ± 4.01% | 96.67 ± 1.25% | 0.44 ± 0.41% |
| EFV | 0.38 ± 0.30% | 9.30 ± 1.56% ** | 42.90 ± 5.60% ** | 98.33 ± 0.47% | 0.14 ± 0.11% | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiou, P.-T.; Ohms, S.; Board, P.G.; Dahlstrom, J.E.; Rangasamy, D.; Casarotto, M.G. The Antiviral Drug Efavirenz in Breast Cancer Stem Cell Therapy. Cancers 2021, 13, 6232. https://doi.org/10.3390/cancers13246232
Chiou P-T, Ohms S, Board PG, Dahlstrom JE, Rangasamy D, Casarotto MG. The Antiviral Drug Efavirenz in Breast Cancer Stem Cell Therapy. Cancers. 2021; 13(24):6232. https://doi.org/10.3390/cancers13246232
Chicago/Turabian StyleChiou, Pey-Tsyr, Stephen Ohms, Philip G. Board, Jane E. Dahlstrom, Danny Rangasamy, and Marco G. Casarotto. 2021. "The Antiviral Drug Efavirenz in Breast Cancer Stem Cell Therapy" Cancers 13, no. 24: 6232. https://doi.org/10.3390/cancers13246232
APA StyleChiou, P. -T., Ohms, S., Board, P. G., Dahlstrom, J. E., Rangasamy, D., & Casarotto, M. G. (2021). The Antiviral Drug Efavirenz in Breast Cancer Stem Cell Therapy. Cancers, 13(24), 6232. https://doi.org/10.3390/cancers13246232