
Blood Bacterial DNA Load and Profiling Differ in Colorectal Cancer Patients Compared to Tumor-Free Controls

 , ,
, ,  , , , , ,
, , , , ,  and add
Show full author list
and add
Show full author list
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
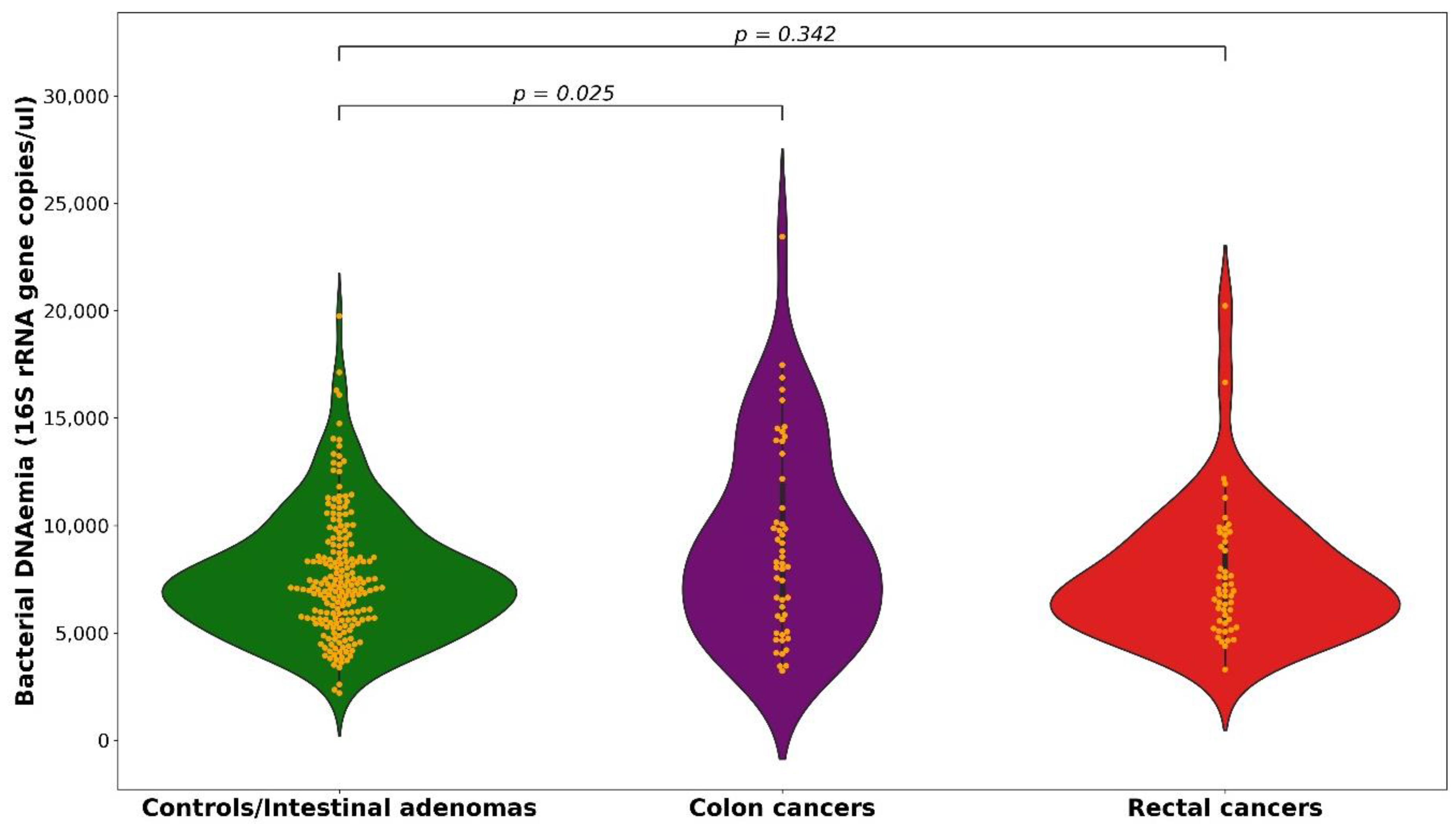
2.1. 16S rRNA Gene Copies
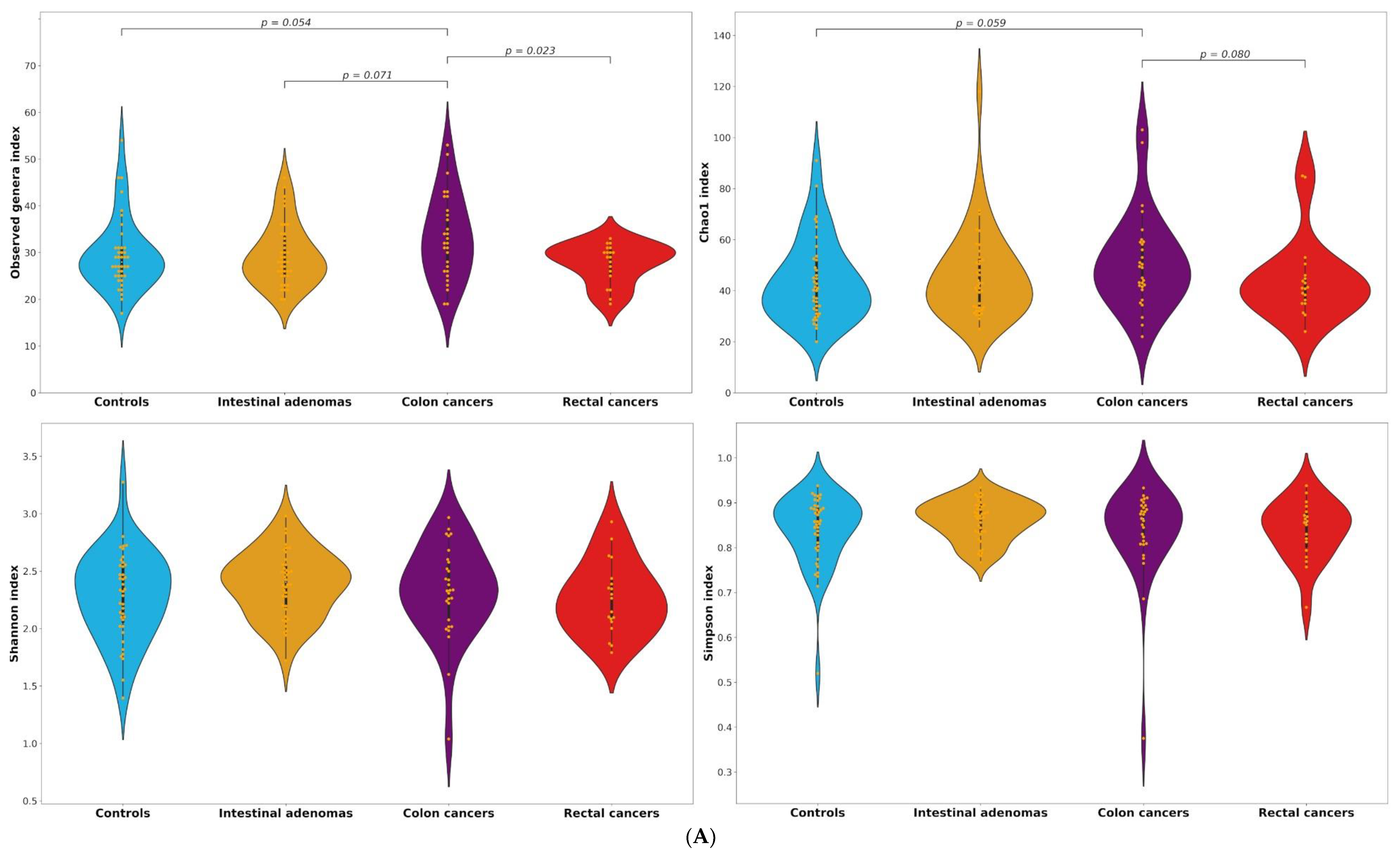
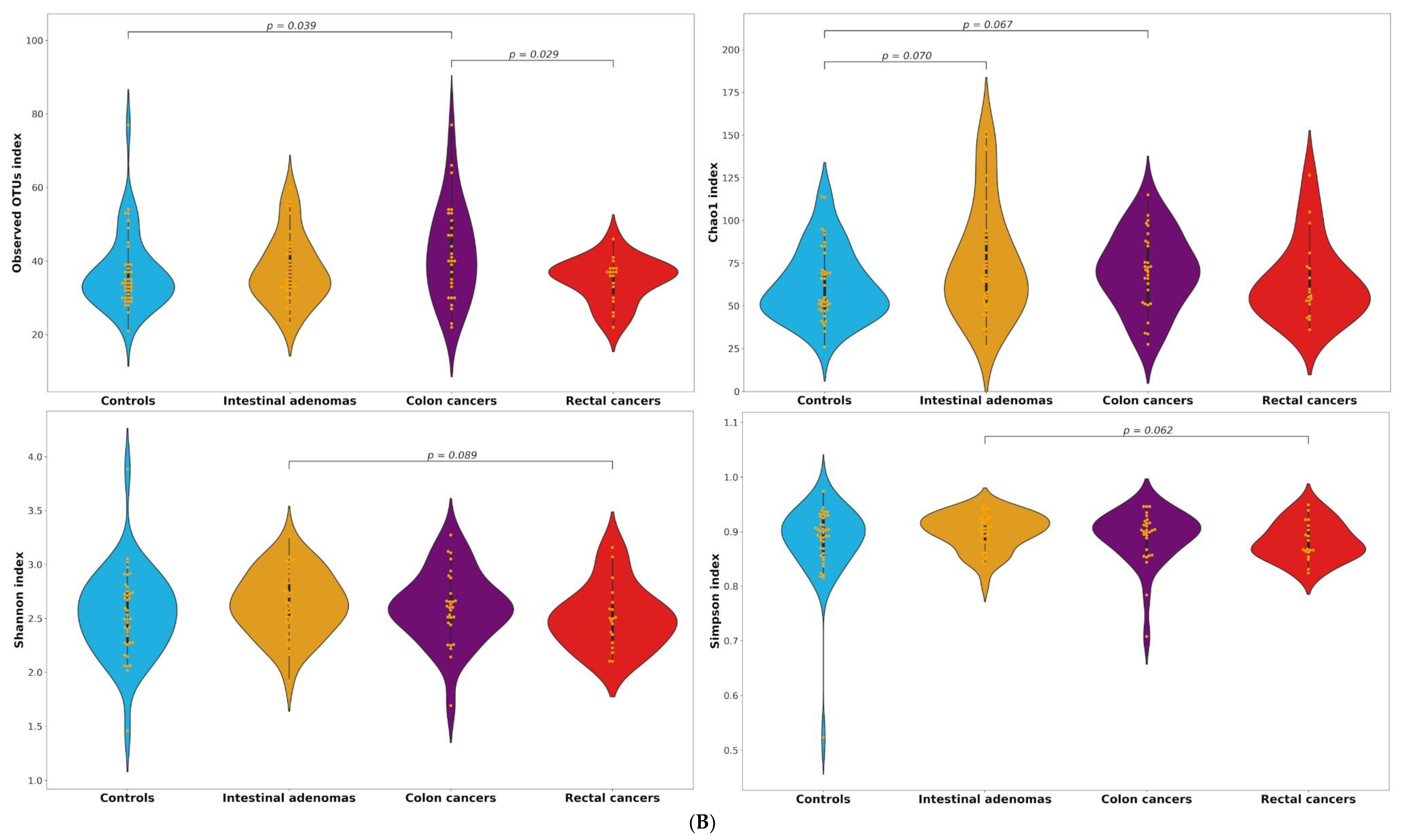
2.2. Alpha and Beta Diversity
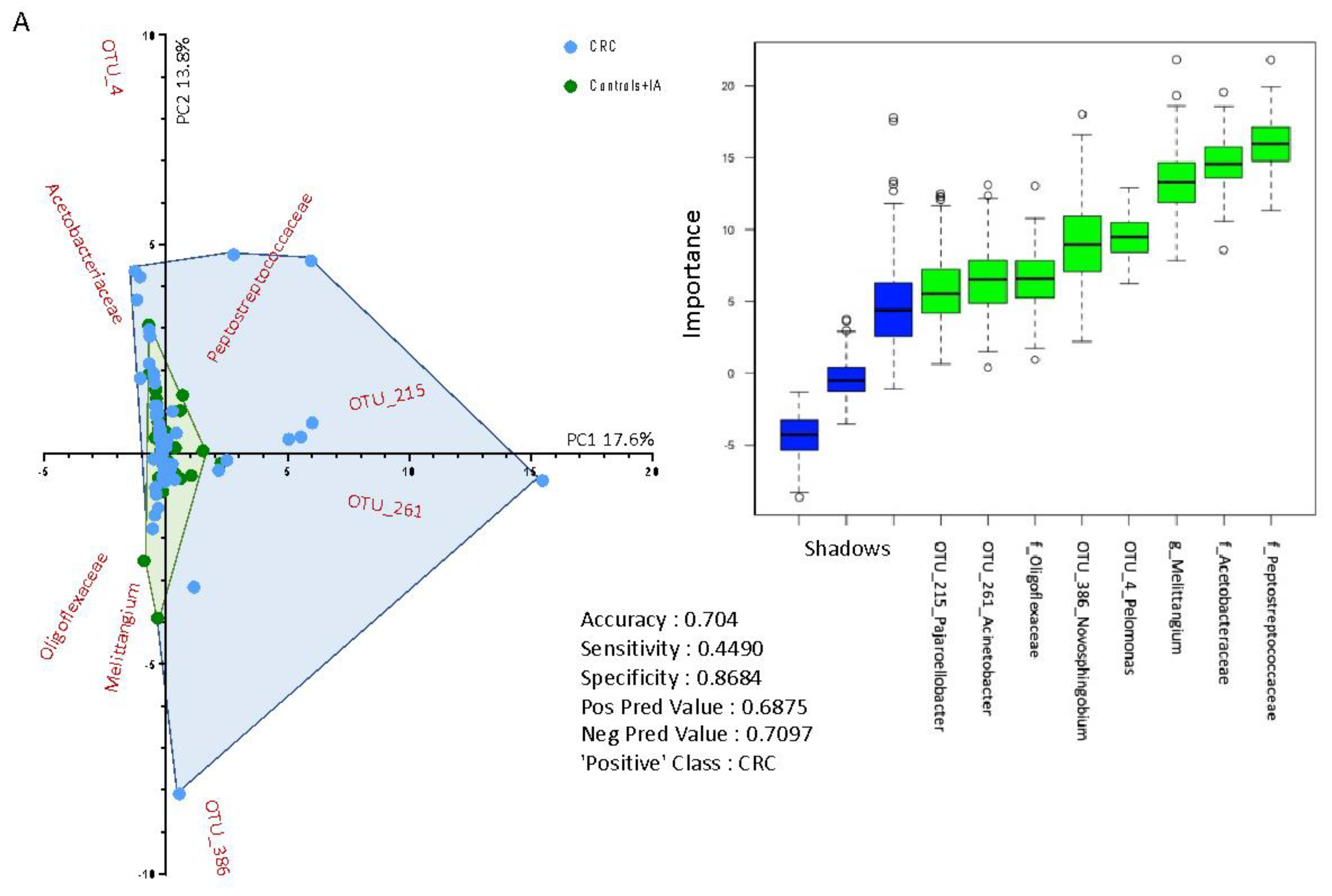
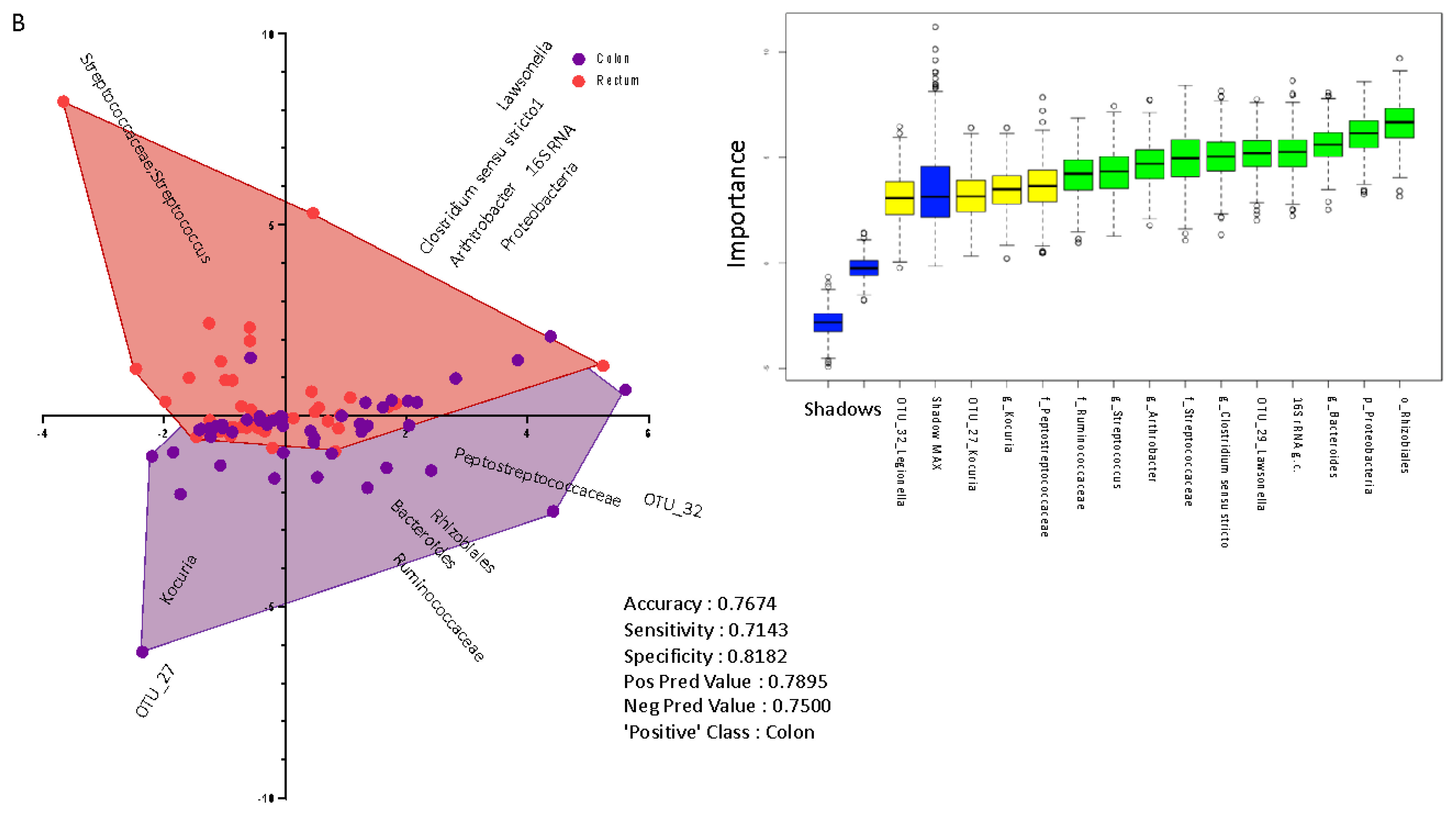
2.3. Taxonomic Profiling of Blood Bacterial DNA between Groups
3. Discussion
4. Materials and Methods
4.1. Interview
4.2. Blood Collection
4.3. DNA Extraction, qPCR Experiments and Sequencing of 16S rRNA Gene Amplicons
4.4. Bacterial DNA Contamination Assessment
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Ait Ouakrim, D.; Pizot, C.; Boniol, M.; Malvezzi, M.; Boniol, M.; Negri, E.; Bota, M.; Jenkins, M.A.; Bleiberg, H.; Autier, P. Trends in colorectal cancer mortality in europe: Retrospective analysis of the who mortality database. BMJ 2015, 351, h4970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carioli, G.; Bertuccio, P.; Boffetta, P.; Levi, F.; La Vecchia, C.; Negri, E.; Malvezzi, M. European cancer mortality predictions for the year 2020 with a focus on prostate cancer. Ann. Oncol. 2020, 31, 650–658. [Google Scholar] [CrossRef] [PubMed]
- Vogelstein, B.; Papadopoulos, N.; Velculescu, V.E.; Zhou, S.B.; Diaz, L.A.; Kinzler, K.W. Cancer genome landscapes. Science 2013, 339, 1546–1558. [Google Scholar] [CrossRef] [PubMed]
- Terzic, J.; Grivennikov, S.; Karin, E.; Karin, M. Inflammation and colon cancer. Gastroenterology 2010, 138, 2101–2114.e5. [Google Scholar] [CrossRef]
- Herrinton, L.J.; Liu, L.; Levin, T.R.; Allison, J.E.; Lewis, J.D.; Velayos, F. Incidence and mortality of colorectal adenocarcinoma in persons with inflammatory bowel disease from 1998 to 2010. Gastroenterology 2012, 143, 382–389. [Google Scholar] [CrossRef]
- Godos, J.; Biondi, A.; Galvano, F.; Basile, F.; Sciacca, S.; Giovannucci, E.L.; Grosso, G. Markers of systemic inflammation and colorectal adenoma risk: Meta-analysis of observational studies. World J. Gastroenterol. 2017, 23, 1909–1919. [Google Scholar] [CrossRef]
- Huang, W.Y.; Berndt, S.I.; Shiels, M.S.; Katki, H.A.; Chaturvedi, A.K.; Wentzensen, N.; Trabert, B.; Kemp, T.J.; Pinto, L.A.; Hildesheim, A.; et al. Circulating inflammation markers and colorectal adenoma risk. Carcinogenesis 2019, 40, 765–770. [Google Scholar] [CrossRef]
- Liu, X.; Cheng, Y.; Shao, L.; Ling, Z. Alterations of the predominant fecal microbiota and disruption of the gut mucosal barrier in patients with early-stage colorectal cancer. Biomed. Res. Int. 2020, 2020, 2948282. [Google Scholar] [CrossRef]
- Ahn, J.; Sinha, R.; Pei, Z.; Dominianni, C.; Wu, J.; Shi, J.; Goedert, J.J.; Hayes, R.B.; Yang, L. Human gut microbiome and risk for colorectal cancer. J. Natl. Cancer Inst. 2013, 105, 1907–1911. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Chan, A.T.; Sun, J. Influence of the gut microbiome, diet, and environment on risk of colorectal cancer. Gastroenterology 2020, 158, 322–340. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.S.; DeSantis, T.Z.; Weinmaier, T.; McMurdie, P.J.; Cope, J.L.; Altrichter, A.; Yamal, J.M.; Hollister, E.B. Leveraging sequence-based faecal microbial community survey data to identify a composite biomarker for colorectal cancer. Gut 2018, 67, 882–891. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Feng, Q.; Wong, S.H.; Zhang, D.; Liang, Q.Y.; Qin, Y.; Tang, L.; Zhao, H.; Stenvang, J.; Li, Y.; et al. Metagenomic analysis of faecal microbiome as a tool towards targeted non-invasive biomarkers for colorectal cancer. Gut 2017, 66, 70–78. [Google Scholar] [CrossRef]
- Thomas, A.M.; Manghi, P.; Asnicar, F.; Pasolli, E.; Armanini, F.; Zolfo, M.; Beghini, F.; Manara, S.; Karcher, N.; Pozzi, C.; et al. Metagenomic analysis of colorectal cancer datasets identifies cross-cohort microbial diagnostic signatures and a link with choline degradation. Nat. Med. 2019, 25, 667–678. [Google Scholar] [CrossRef] [Green Version]
- Wirbel, J.; Pyl, P.T.; Kartal, E.; Zych, K.; Kashani, A.; Milanese, A.; Fleck, J.S.; Voigt, A.Y.; Palleja, A.; Ponnudurai, R.; et al. Meta-analysis of fecal metagenomes reveals global microbial signatures that are specific for colorectal cancer. Nat. Med. 2019, 25, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Castillo, D.J.; Rifkin, R.F.; Cowan, D.A.; Potgieter, M. The healthy human blood microbiome: Fact or fiction? Front. Cell. Infect. Microbiol. 2019, 9, 148. [Google Scholar] [CrossRef] [Green Version]
- Paisse, S.; Valle, C.; Servant, F.; Courtney, M.; Burcelin, R.; Amar, J.; Lelouvier, B. Comprehensive description of blood microbiome from healthy donors assessed by 16s targeted metagenomic sequencing. Transfusion 2016, 56, 1138–1147. [Google Scholar] [CrossRef] [PubMed]
- Lelouvier, B.; Servant, F.; Paisse, S.; Brunet, A.C.; Benyahya, S.; Serino, M.; Valle, C.; Ortiz, M.R.; Puig, J.; Courtney, M.; et al. Changes in blood microbiota profiles associated with liver fibrosis in obese patients: A pilot analysis. Hepatology 2016, 64, 2015–2027. [Google Scholar] [CrossRef]
- Sato, J.; Kanazawa, A.; Ikeda, F.; Yoshihara, T.; Goto, H.; Abe, H.; Komiya, K.; Kawaguchi, M.; Shimizu, T.; Ogihara, T.; et al. Gut dysbiosis and detection of “live gut bacteria” in blood of japanese patients with type 2 diabetes. Diabetes Care 2014, 37, 2343–2350. [Google Scholar] [CrossRef] [Green Version]
- Poore, G.D.; Kopylova, E.; Zhu, Q.; Carpenter, C.; Fraraccio, S.; Wandro, S.; Kosciolek, T.; Janssen, S.; Metcalf, J.; Song, S.J.; et al. Microbiome analyses of blood and tissues suggest cancer diagnostic approach. Nature 2020, 579, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Lu, W.; Kong, X.; Shao, Y.W.; Hu, Y.; Wang, A.; Bao, H.; Cao, R.; Liu, K.; Wang, X.; et al. Alterations of circulating bacterial DNA in colorectal cancer and adenoma: A proof-of-concept study. Cancer Lett. 2021, 499, 201–208. [Google Scholar] [CrossRef]
- Messaritakis, I.; Vogiatzoglou, K.; Tsantaki, K.; Ntretaki, A.; Sfakianaki, M.; Koulouridi, A.; Tsiaoussis, J.; Mavroudis, D.; Souglakos, J. The prognostic value of the detection of microbial translocation in the blood of colorectal cancer patients. Cancers 2020, 12, 1058. [Google Scholar] [CrossRef]
- King, C.H.; Desai, H.; Sylvetsky, A.C.; LoTempio, J.; Ayanyan, S.; Carrie, J.; Crandall, K.A.; Fochtman, B.C.; Gasparyan, L.; Gulzar, N.; et al. Baseline human gut microbiota profile in healthy people and standard reporting template. PLoS ONE 2019, 14, e0206484. [Google Scholar] [CrossRef] [Green Version]
- Shah, N.B.; Allegretti, A.S.; Nigwekar, S.U.; Kalim, S.; Zhao, S.; Lelouvier, B.; Servant, F.; Serena, G.; Thadhani, R.I.; Raj, D.S.; et al. Blood microbiome profile in ckd: A pilot study. Clin. J. Am. Soc. Nephrol. 2019, 14, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Flemer, B.; Warren, R.D.; Barrett, M.P.; Cisek, K.; Das, A.; Jeffery, I.B.; Hurley, E.; O’Riordain, M.; Shanahan, F.; O’Toole, P.W. The oral microbiota in colorectal cancer is distinctive and predictive. Gut 2018, 67, 1454–1463. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.S.; DeSantis, T.; Yamal, J.M.; Weir, T.; Ryan, E.P.; Cope, J.L.; Hollister, E.B. Re-purposing 16s rrna gene sequence data from within case paired tumor biopsy and tumor-adjacent biopsy or fecal samples to identify microbial markers for colorectal cancer. PLoS ONE 2018, 13, e0207002. [Google Scholar] [CrossRef]
- Peris-Bondia, F.; Latorre, A.; Artacho, A.; Moya, A.; D’Auria, G. The active human gut microbiota differs from the total microbiota. PLoS ONE 2011, 6, e22448. [Google Scholar] [CrossRef] [Green Version]
- Vital, M.; Karch, A.; Pieper, D.H. Colonic butyrate-producing communities in humans: An overview using omics data. mSystems 2017, 2, e00130-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkataraman, A.; Sieber, J.R.; Schmidt, A.W.; Waldron, C.; Theis, K.R.; Schmidt, T.M. Variable responses of human microbiomes to dietary supplementation with resistant starch. Microbiome 2016, 4, 33. [Google Scholar] [CrossRef] [Green Version]
- Bhat, A.A.; Uppada, S.; Achkar, I.W.; Hashem, S.; Yadav, S.K.; Shanmugakonar, M.; Al-Naemi, H.A.; Haris, M.; Uddin, S. Tight junction proteins and signaling pathways in cancer and inflammation: A functional crosstalk. Front. Physiol. 2018, 9, 1942. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.C. Microbiota dysbiosis and barrier dysfunction in inflammatory bowel disease and colorectal cancers: Exploring a common ground hypothesis. J. Biomed. Sci. 2018, 25, 79. [Google Scholar] [CrossRef] [Green Version]
- Taverniti, V.; Stuknyte, M.; Minuzzo, M.; Arioli, S.; De Noni, I.; Scabiosi, C.; Cordova, Z.M.; Junttila, I.; Hamalainen, S.; Turpeinen, H.; et al. S-layer protein mediates the stimulatory effect of lactobacillus helveticus mimlh5 on innate immunity. Appl. Environ. Microbiol. 2013, 79, 1221–1231. [Google Scholar] [CrossRef] [Green Version]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Wasinger, V.C.; Lu, K.; Yau, Y.Y.; Nash, J.; Lee, J.; Chang, J.; Paramsothy, S.; Kaakoush, N.O.; Mitchell, H.M.; Leong, R.W.L. Spp24 is associated with endocytic signalling, lipid metabolism, and discrimination of tissue integrity for ‘leaky-gut’ in inflammatory bowel disease. Sci. Rep. 2020, 10, 12932. [Google Scholar] [CrossRef] [PubMed]
- Bertocchi, A.; Carloni, S.; Ravenda, P.S.; Bertalot, G.; Spadoni, I.; Lo Cascio, A.; Gandini, S.; Lizier, M.; Braga, D.; Asnicar, F.; et al. Gut vascular barrier impairment leads to intestinal bacteria dissemination and colorectal cancer metastasis to liver. Cancer Cell 2021, 39, 708–724.e711. [Google Scholar] [CrossRef] [PubMed]
- Paschke, S.; Jafarov, S.; Staib, L.; Kreuser, E.D.; Maulbecker-Armstrong, C.; Roitman, M.; Holm, T.; Harris, C.C.; Link, K.H.; Kornmann, M. Are colon and rectal cancer two different tumor entities? A proposal to abandon the term colorectal cancer. Int. J. Mol. Sci. 2018, 19, 2577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, E.K.; Giovannucci, E.; Wu, K.; Rosner, B.; Fuchs, C.S.; Willett, W.C.; Colditz, G.A. Comparison of risk factors for colon and rectal cancer. Int. J. Cancer 2004, 108, 433–442. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Haines, C.; Watson, A.J.M.; Hart, A.R.; Platt, M.J.; Pardoll, D.M.; Cosgrove, S.E.; Gebo, K.A.; Sears, C.L. Oral antibiotic use and risk of colorectal cancer in the United Kingdom, 1989–2012: A matched case-control study. Gut 2019, 68, 1971–1978. [Google Scholar] [CrossRef]
- Hjartaker, A.; Aagnes, B.; Robsahm, T.E.; Langseth, H.; Bray, F.; Larsen, I.K. Subsite-specific dietary risk factors for colorectal cancer: A review of cohort studies. J. Oncol. 2013, 2013, 703854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, M.; Mascaretti, F.; Parpinel, M.; Serraino, D.; Crispo, A.; Celentano, E.; Giacosa, A.; La Vecchia, C. Dietary intake of branched-chain amino acids and colorectal cancer risk. Br. J. Nutr. 2021, 126, 22–27. [Google Scholar] [CrossRef]
- Lee, G.H.; Malietzis, G.; Askari, A.; Bernardo, D.; Al-Hassi, H.O.; Clark, S.K. Is right-sided colon cancer different to left-sided colorectal cancer?—A systematic review. Eur. J. Surg. Oncol. 2015, 41, 300–308. [Google Scholar] [CrossRef]
- Glebov, O.K.; Rodriguez, L.M.; Nakahara, K.; Jenkins, J.; Cliatt, J.; Humbyrd, C.J.; DeNobile, J.; Soballe, P.; Simon, R.; Wright, G.; et al. Distinguishing right from left colon by the pattern of gene expression. Cancer Epidemiol. Biomark. Prev. 2003, 12, 755–762. [Google Scholar]
- Tamas, K.; Walenkamp, A.M.; de Vries, E.G.; van Vugt, M.A.; Beets-Tan, R.G.; van Etten, B.; de Groot, D.J.; Hospers, G.A. Rectal and colon cancer: Not just a different anatomic site. Cancer Treat. Rev. 2015, 41, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Herath, M.; Hosie, S.; Bornstein, J.C.; Franks, A.E.; Hill-Yardin, E.L. The role of the gastrointestinal mucus system in intestinal homeostasis: Implications for neurological disorders. Front. Cell. Infect. Microbiol. 2020, 10, 248. [Google Scholar] [CrossRef]
- Kamphuis, J.B.J.; Mercier-Bonin, M.; Eutamene, H.; Theodorou, V. Mucus organisation is shaped by colonic content; a new view. Sci. Rep. 2017, 7, 8527. [Google Scholar] [CrossRef] [Green Version]
- Bergstrom, K.; Shan, X.; Casero, D.; Batushansky, A.; Lagishetty, V.; Jacobs, J.P.; Hoover, C.; Kondo, Y.; Shao, B.; Gao, L.; et al. Proximal colon-derived o-glycosylated mucus encapsulates and modulates the microbiota. Science 2020, 370, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, D.F. Evaluating a new cancer screening blood test: Unintended consequences and the need for clarity in policy making. J. Natl. Cancer Inst. 2021, 113, 109–111. [Google Scholar] [CrossRef]
- Boleij, A.; Tjalsma, H. The itinerary of streptococcus gallolyticus infection in patients with colonic malignant disease. Lancet Infect. Dis. 2013, 13, 719–724. [Google Scholar] [CrossRef]
- Wang, H.F.; Li, L.F.; Guo, S.H.; Zeng, Q.Y.; Ning, F.; Liu, W.L.; Zhang, G. Evaluation of antibody level against fusobacterium nucleatum in the serological diagnosis of colorectal cancer. Sci. Rep. 2016, 6, 33440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, D.L.; Leighton, J.A.; Gurudu, S.R. Approach to incomplete colonoscopy: New techniques and technologies. Gastroenterol. Hepatol. 2017, 13, 476–483. [Google Scholar]
- Genua, F.; Raghunathan, V.; Jenab, M.; Gallagher, W.M.; Hughes, D.J. The role of gut barrier dysfunction and microbiome dysbiosis in colorectal cancer development. Front. Oncol. 2021, 11, 626349. [Google Scholar] [CrossRef] [PubMed]
- Gargari, G.; Mantegazza, G.; Taverniti, V.; Del Bo, C.; Bernardi, S.; Andres-Lacueva, C.; Gonzalez-Dominguez, R.; Kroon, P.A.; Winterbone, M.S.; Cherubini, A.; et al. Bacterial dnaemia is associated with serum zonulin levels in older subjects. Sci. Rep. 2021, 11, 11054. [Google Scholar] [CrossRef]
- Lluch, J.; Servant, F.; Paisse, S.; Valle, C.; Valiere, S.; Kuchly, C.; Vilchez, G.; Donnadieu, C.; Courtney, M.; Burcelin, R.; et al. The characterization of novel tissue microbiota using an optimized 16s metagenomic sequencing pipeline. PLoS ONE 2015, 10, e0142334. [Google Scholar] [CrossRef]
- Nadkarni, M.A.; Martin, F.E.; Jacques, N.A.; Hunter, N. Determination of bacterial load by real-time PCR using a broad-range (universal) probe and primers set. Microbiology 2002, 148, 257–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escudie, F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauquil, L.; Vidal, K.; Maman, S.; Hernandez-Raquet, G.; Combes, S.; Pascal, G. Frogs: Find, rapidly, otus with galaxy solution. Bioinformatics 2018, 34, 1287–1294. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Controls | IA | CRC |
|---|---|---|---|
| Sex | |||
| Male | 62 (62%) | 62 (62%) | 62 (62%) |
| Female | 38 (38%) | 38 (38%) | 38 (38%) |
| Age group (years) | |||
| <50 | 7 (7%) | 4 (4%) | 10 (10%) |
| 50–59 | 23 (23%) | 20 (20%) | 19 (19%) |
| 60–69 | 26 (26%) | 36 (36%) | 29 (29%) |
| 70–79 | 33 (33%) | 29 (29%) | 31 (42%) |
| ≥80 | 11 (11%) | 11 (11%) | 11 (11%) |
| χ2 test, p = 0.76 | |||
| Mean (SD) age (years) * | 66.0 (11.8) | 65.9 (10.9) | 66.1 (11.6) |
| Center | |||
| Niguarda | 65 (65%) | 65 (65%) | 65 (65%) |
| Policlinico | 35 (35%) | 35 (35%) | 35 (35%) |
| Education (years) † | |||
| <7 | 12 (12%) | 19 (19%) | 25 (25%) |
| 7–11 | 24 (24%) | 26 (26%) | 25 (25%) |
| ≥12 | 64 (64%) | 55 (55%) | 49 (50%) |
| χ2 test, p = 0.155 |
| Mean (SD) | Quintile of Number of Gene Copies †, OR (95% CI) | χ2 Trend (p Value) across the 3 Categories | Continuous OR § | |||
|---|---|---|---|---|---|---|
| 1–3 ‡ | 4 | 5 | ||||
| Upper cutpoints (n copies/µL) | 7617.5 | 9707.4 | - | |||
| Control/IA, n (%) | 7606.6 (3895.8) | 120 (60%) | 40 (20%) | 40 (20%) | ||
| Total CRC, n (%) | 8387.1 (2865.4) | 52 (52%) | 20 (20%) | 28 (28%) | ||
| 1 ‡ | 1.16 | 1.59 | 2.40 | 1.39 | ||
| (0.60–2.22) | (0.89–2.82) | (0.121) | (1.00–1.92) | |||
| Colon cancer, n (%) | 9145.4 (4476.2) | 21 (42%) | 10 (20%) | 19 (38%) | ||
| 1 ‡ | 1.96 | 2.62 | 6.21 | 2.02 | ||
| (0.75–5.08) | (1.22–5.65) | (0.013) | (1.26–3.25) | |||
| Rectal cancer, n (%) | 7628.8 (3075.0) | 31 (62%) | 10 (20%) | 9 (18%) | ||
| 1 ‡ | 0.73 | 0.81 | 0.358 | 0.86 | ||
| (0.29–1.84) | (0.32–2.03) | (0.549) | (0.51–1.42) | |||
| χ2 interaction (p value) between colon and rectum | 5.30 (0.021) | 4.34 (0.037) | ||||
| Total | Quintile of Number of Gene Copies *, n (%) | |||
|---|---|---|---|---|
| 1–3 | 4 | 5 | ||
| Control/IA | 200 | 120 (60%) | 40 (20%) | 40 (20%) |
| Tumor subsite | ||||
| Right colon | 21 | 7 (33%) | 3 (14%) | 11 (53%) |
| Cecum | 4 | 2 (50%) | 0 (0%) | 2 (50%) |
| Ascending | 11 | 2 (18%) | 2 (18%) | 7 (64%) |
| Hepatic flexure | 6 | 3 (50%) | 1 (17%) | 2 (33%) |
| Other than right colon | 29 | 14 (48%) | 7 (24%) | 8 (38%) |
| Transverse colon | 2 | 1 (50%) | 1 (50%) | 0 (0%) |
| Splenic flexure | 3 | 1 (33%) | 1 (33%) | 1 (33%) |
| Descending colon | 7 | 4 (57%) | 2 (29%) | 1 (14%) |
| Sigmoid colon | 17 | 8 (47%) | 3 (18%) | 6 (35%) |
| Rectum | 50 | 31 (62%) | 10 (20%) | 9 (18%) |
| Rectosigmoid junction | 3 | 3 (100%) | 0 (0%) | 0 (0%) |
| Rectum | 47 | 28 (60%) | 10 (21%) | 9 (19%) |
| Median (I–III Quartiles) | p * Controls vs IA | p * Colon Cancers vs IA | p * Colon Cancer vs Controls | p * Rectal Cancer vs IA | p * Rectal Cancer vs controls | p * Colon Cancer vs Rectal Cancer | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Alpha-Diversity | Controls | IA | Colon Cancer | Rectal Cancer | |||||||
| Genera | Observed | 28 (25–31) | 28 (25–33.5) | 32 (26–39) | 29 (25–31) | 0.942 | 0.071 | 0.054 | 0.714 | 0.968 | 0.023 |
| Chao1 | 40.6 (31.7–52.3) | 42.5 (33.4–52.5) | 49 (41.5–59) | 41 (35–46) | 0.476 | 0.148 | 0.059 | 0.703 | 0.789 | 0.080 | |
| Shannon | 2.33 (2,05–2,55) | 2.41 (2.18–2.56) | 2.33 (2.08–2.58) | 2.26 (2.06–2.44) | 0.254 | 0.535 | 0.614 | 0.119 | 0.715 | 0.442 | |
| Simpson | 0.86 (0.80–0.89) | 0.87 (0.84–0.89) | 0.84 (0.81–0.89) | 0.86 (0.79–0.88) | 0.394 | 0.496 | 0.846 | 0.186 | 0.697 | 0.513 | |
| OTUs | Observed | 34 (30–39) | 35 (32.4–43.5) | 40 (33–51) | 37 (30–38) | 0.413 | 0.154 | 0.039 | 0.570 | 0.820 | 0.029 |
| Chao1 | 53.4 (47.8–70.5) | 66 (51.9–92.8) | 71.1 (52–87) | 56 (51–73) | 0.070 | 0.981 | 0.067 | 0.233 | 0.565 | 0.278 | |
| Shannon | 2.52 (2.27–2.74) | 2.63 (2.46–2.84) | 2.60 (2.46–2.73) | 2.50 (2.27–2.65) | 0.154 | 0.662 | 0.473 | 0.089 | 0.727 | 0.149 | |
| Simpson | 0.90 (0.86–0.93) | 0.91 (0.88–0.93) | 0.90 (0.87–0.92) | 0.87 (0.86–0.91) | 0.233 | 0.473 | 0.653 | 0.062 | 0.403 | 0.229 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mutignani, M.; Penagini, R.; Gargari, G.; Guglielmetti, S.; Cintolo, M.; Airoldi, A.; Leone, P.; Carnevali, P.; Ciafardini, C.; Petrocelli, G.; et al. Blood Bacterial DNA Load and Profiling Differ in Colorectal Cancer Patients Compared to Tumor-Free Controls. Cancers 2021, 13, 6363. https://doi.org/10.3390/cancers13246363
Mutignani M, Penagini R, Gargari G, Guglielmetti S, Cintolo M, Airoldi A, Leone P, Carnevali P, Ciafardini C, Petrocelli G, et al. Blood Bacterial DNA Load and Profiling Differ in Colorectal Cancer Patients Compared to Tumor-Free Controls. Cancers. 2021; 13(24):6363. https://doi.org/10.3390/cancers13246363
Chicago/Turabian StyleMutignani, Massimiliano, Roberto Penagini, Giorgio Gargari, Simone Guglielmetti, Marcello Cintolo, Aldo Airoldi, Pierfrancesco Leone, Pietro Carnevali, Clorinda Ciafardini, Giulio Petrocelli, and et al. 2021. "Blood Bacterial DNA Load and Profiling Differ in Colorectal Cancer Patients Compared to Tumor-Free Controls" Cancers 13, no. 24: 6363. https://doi.org/10.3390/cancers13246363
APA StyleMutignani, M., Penagini, R., Gargari, G., Guglielmetti, S., Cintolo, M., Airoldi, A., Leone, P., Carnevali, P., Ciafardini, C., Petrocelli, G., Mascaretti, F., Oreggia, B., Dioscoridi, L., Cavalcoli, F., Primignani, M., Pugliese, F., Bertuccio, P., Soru, P., Magistro, C., ... Rossi, M. (2021). Blood Bacterial DNA Load and Profiling Differ in Colorectal Cancer Patients Compared to Tumor-Free Controls. Cancers, 13(24), 6363. https://doi.org/10.3390/cancers13246363