
Liquid Biopsy Targeting Monocarboxylate Transporter 1 on the Surface Membrane of Tumor-Derived Extracellular Vesicles from Synovial Sarcoma

 , , , ,
, , , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Preparation of Conditioned Medium
2.3. Patient Serum and Tissue Samples
2.4. Isolation of EVs from Cell CM
2.5. Isolation of EVs from Human Serum
2.6. Liquid Chromatography-Tandem Mass Spectrometry (LC/MS) Analysis
2.7. Transmission Electron Microscopy (TEM)
2.8. Particle Diameter Analysis
2.9. Animal Experiments
2.10. Immunohistochemistry
2.11. Immunoblot Analysis
2.12. Enzyme-Linked Immunosorbent Assay
2.13. RNA Extraction and RT-qPCR
2.14.1. Cell Proliferation Assay
2.14.2. Wound Healing Assay
2.14.3. Transwell Assay
2.15. Statistical Analysis
3. Results
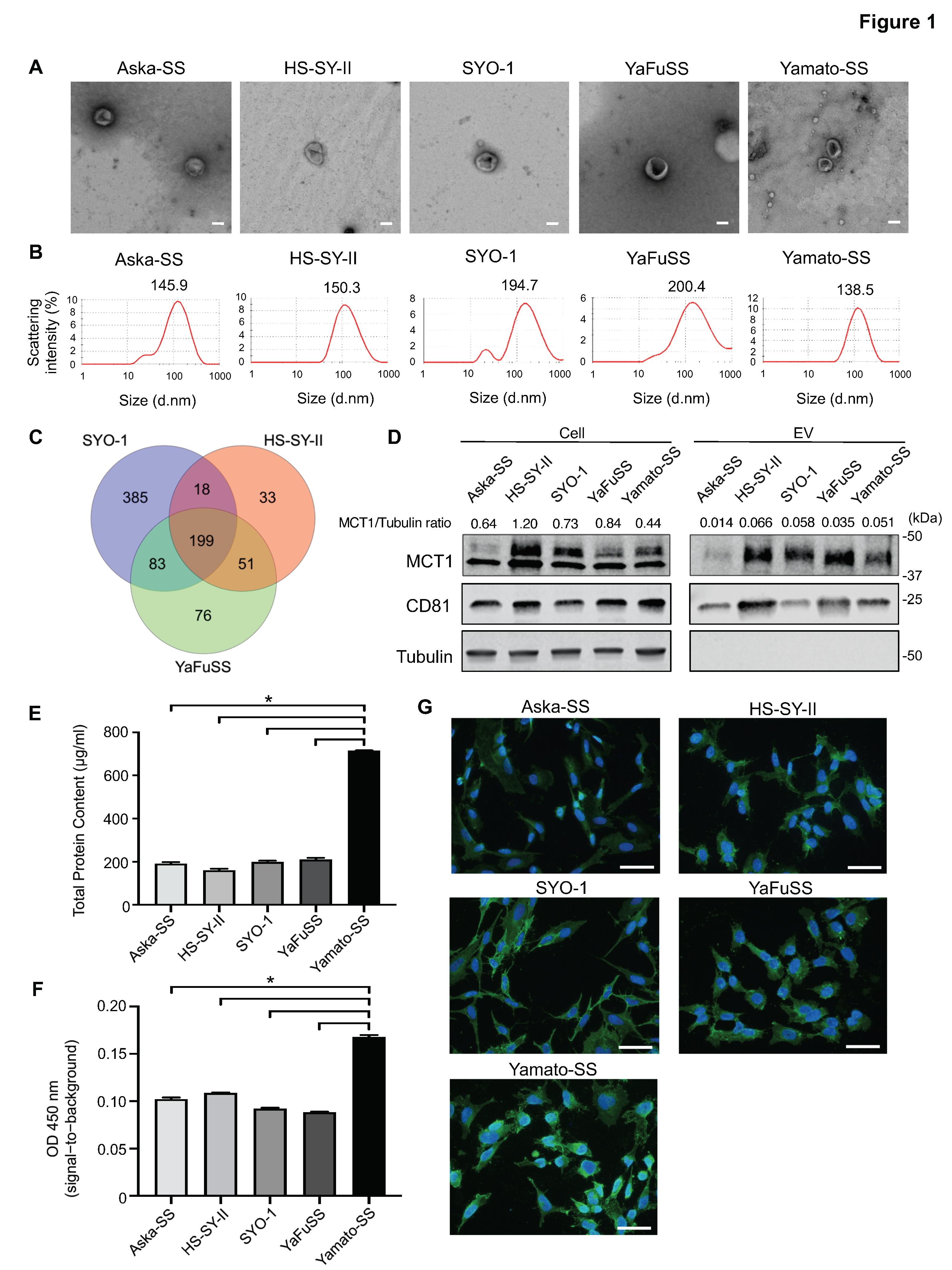
3.1. Isolation and Visualization of EVs from SS cells
3.2. Enrichment of MCT1 on EVs Secreted from SS Cell Lines
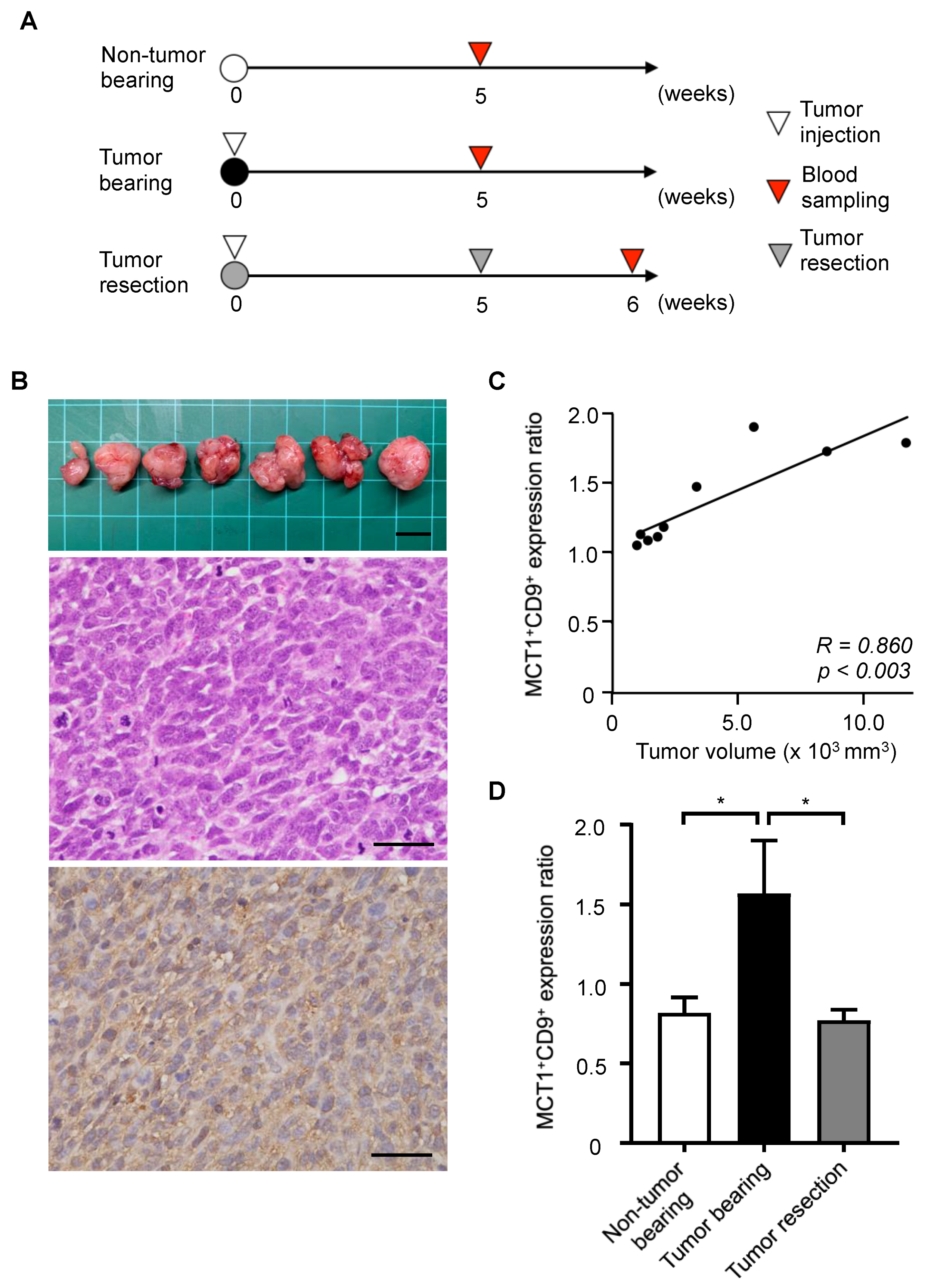
3.3. Tumor Monitoring Using MCT1 and CD9 Double-Positive EVs in a SS Mouse Model
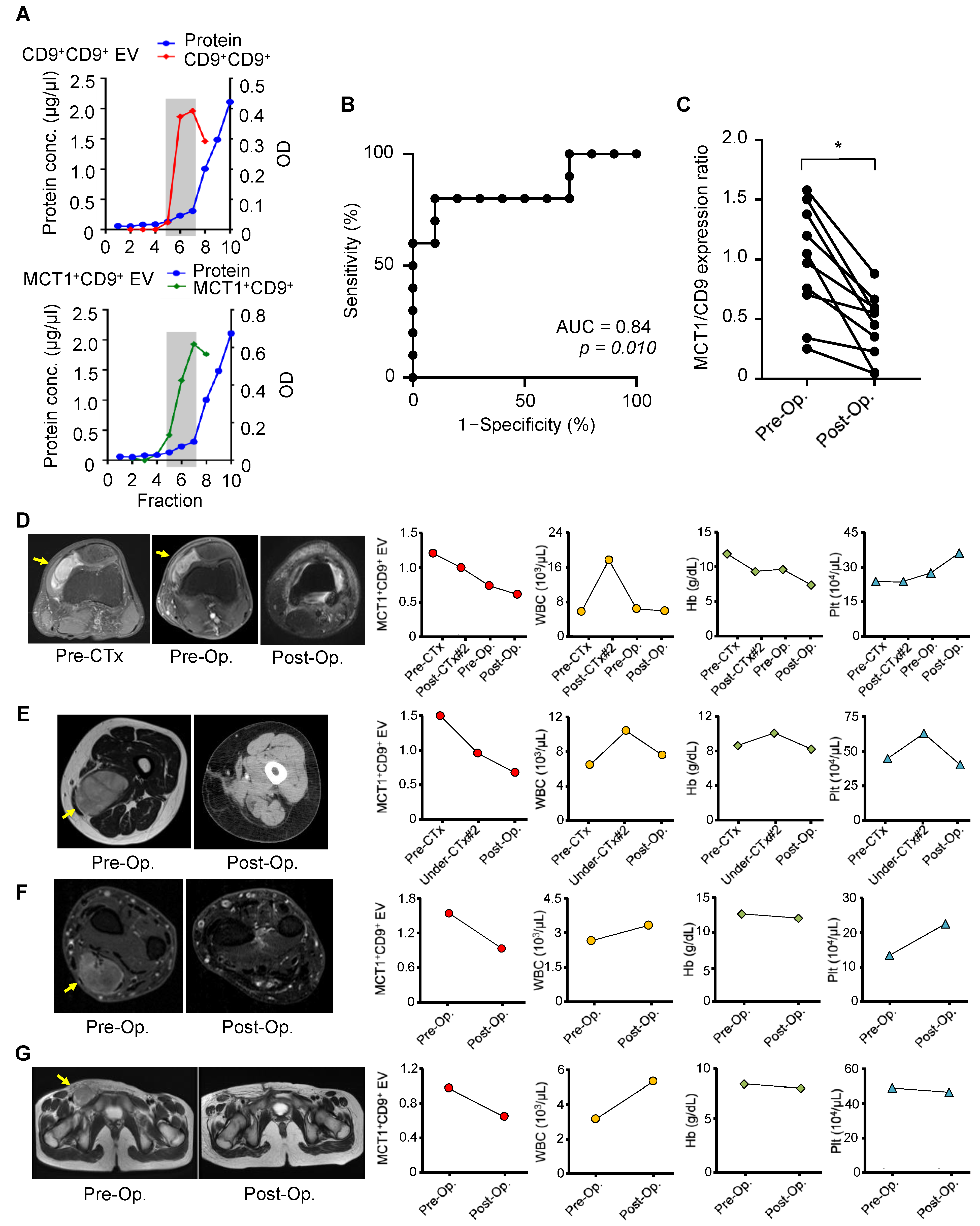
3.4. Tumor Monitoring by Serum MCT1+CD9+ EVs in Patients with SS
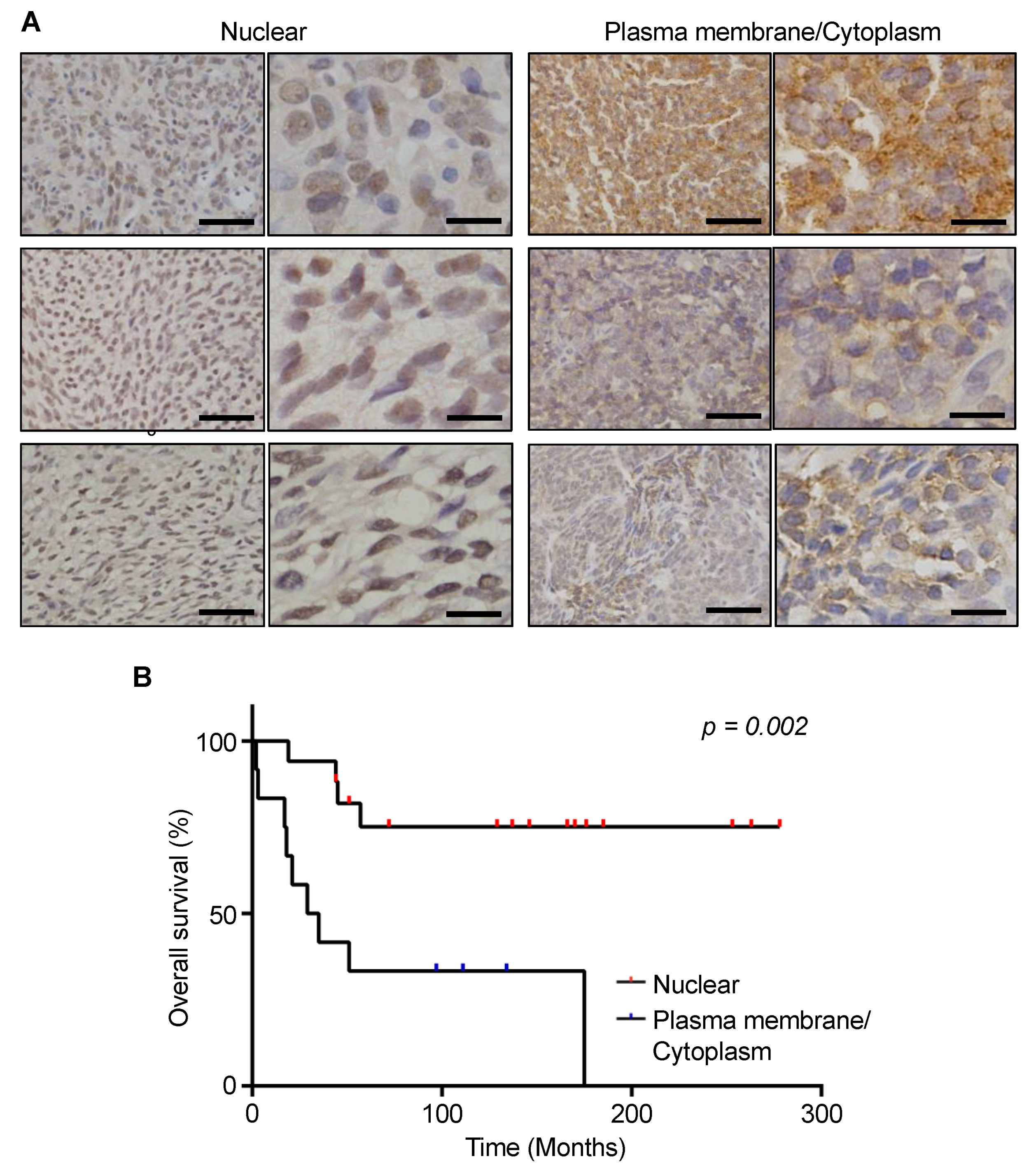
3.5. Prognostic Significance of MCT1 Expression Levels in SS Tumor Specimens
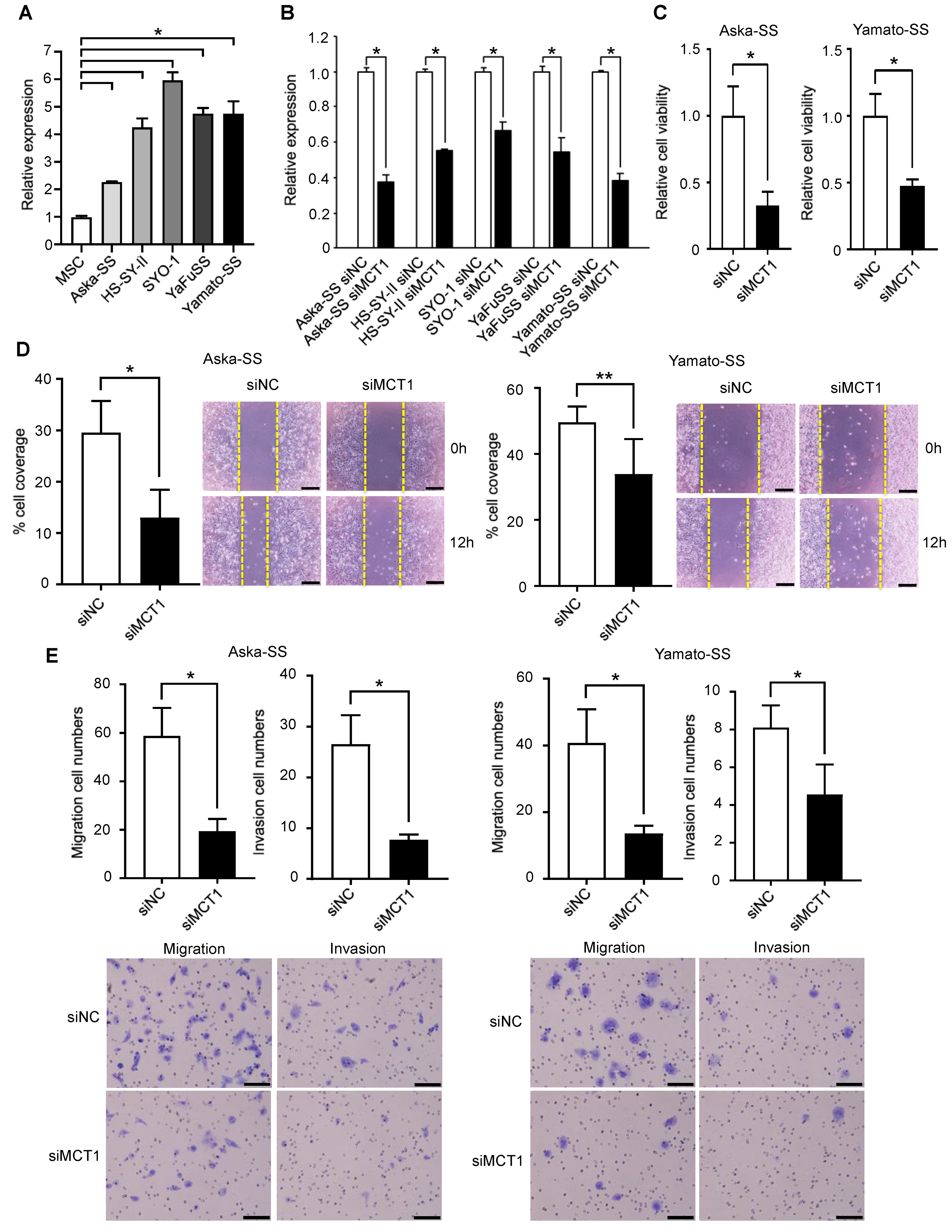
3.6. Silencing of MCT1 Inhibits Tumor Cell Proliferation, Migration, and Invasion of SS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clark, M.A.; Fisher, C.; Judson, I.; Thomas, J.M. Soft-tissue sarcomas in adults. N. Engl. J. Med. 2005, 353, 701–711. [Google Scholar] [CrossRef] [Green Version]
- Christopher, D.; Fletcher, J.A.; Bridge, P. WHO Classification of Tumours of Soft Tissue and Bone, 4th ed.; International Agency for Research on Cancer: Lyon, France, 2013; pp. 110–111. [Google Scholar]
- Hollander, N.; Noone, A.; Krapcho, M.; Garshell, J.; Miller, D.; Altekruse, S. SEER Cancer Statistics Review 1975–2011; National Cancer Institute: Bethesda, MD, USA, 2015. [Google Scholar]
- Jemal, A.; Siegel, R.; Ward, E.; Hao, Y.; Xu, J.; Thun, M. Cancer statistics. CA Cancer J Clin 2009, 59, 225–249. [Google Scholar] [CrossRef]
- Burningham, Z.; Hashibe, M.; Spector, L.; Schiffman, J.D. The epidemiology of sarcoma. Clin. Sarcoma Res. 2012, 2, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dangoor, A.; Seddon, B.; Gerrand, C.; Grimer, R.; Whelan, J.; Judson, I. UK guidelines for the management of soft tissue sarcomas. Clin. Sarcoma Res. 2016, 6, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Mehren, M.; Randall, R.L.; Benjamin, R.S.; Boles, S.; Bui, M.M.; Conrad, E.U.; Ganjoo, K.N.; George, S.; Gonzalez, R.J.; Heslin, M.J. Soft tissue sarcoma, version 2.2016, NCCN clinical practice guidelines in oncology. J. Natl. Compr. Cancer Netw. 2016, 14, 758–786. [Google Scholar] [CrossRef] [Green Version]
- Blay, J.Y. Management of sarcoma patients: Centralization in reference centers to fragmentation of systemic treatment. Curr. Opin. Oncol. 2018, 30, 240–242. [Google Scholar] [CrossRef] [PubMed]
- Casali, P.G.; Abecassis, N.; Aro, H.T.; Bauer, S.; Biagini, R.; Bielack, S.; Bonvalot, S.; Boukovinas, I.; Bovee, J.; Brodowicz, T.; et al. Soft tissue and visceral sarcomas: ESMO-EURACAN Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2018, 29, iv51–iv67. [Google Scholar] [CrossRef] [PubMed]
- Blay, J.Y.; Honore, C.; Stoeckle, E.; Meeus, P.; Jafari, M.; Gouin, F.; Anract, P.; Ferron, G.; Rochwerger, A.; Ropars, M.; et al. Surgery in reference centers improves survival of sarcoma patients: A nationwide study. Ann. Oncol. 2019, 30, 1143–1153. [Google Scholar] [CrossRef]
- Lewis, J.J.; Antonescu, C.R.; Leung, D.H.; Blumberg, D.; Healey, J.H.; Woodruff, J.M.; Brennan, M.F. Synovial sarcoma: A multivariate analysis of prognostic factors in 112 patients with primary localized tumors of the extremity. J. Clin. Oncol. 2000, 18, 2087–2094. [Google Scholar] [CrossRef]
- Kadoch, C.; Crabtree, G.R. Reversible disruption of mSWI/SNF (BAF) complexes by the SS18-SSX oncogenic fusion in synovial sarcoma. Cell 2013, 153, 71–85. [Google Scholar] [CrossRef] [Green Version]
- Vader, P.; Breakefield, X.O.; Wood, M.J. Extracellular vesicles: Emerging targets for cancer therapy. Trends Mol. Med. 2014, 20, 385–393. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, Y.; Kosaka, N.; Konishi, Y.; Ohta, H.; Okamoto, H.; Sonoda, H.; Nonaka, R.; Yamamoto, H.; Ishii, H.; Mori, M.; et al. Ultra-sensitive liquid biopsy of circulating extracellular vesicles using ExoScreen. Nat. Commun. 2014, 5, 3591. [Google Scholar] [CrossRef]
- Al-Nedawi, K.; Meehan, B.; Micallef, J.; Lhotak, V.; May, L.; Guha, A.; Rak, J. Intercellular transfer of the oncogenic receptor EGFRvIII by microvesicles derived from tumour cells. Nat. Cell Biol. 2008, 10, 619–624. [Google Scholar] [CrossRef]
- Iero, M.; Valenti, R.; Huber, V.; Filipazzi, P.; Parmiani, G.; Fais, S.; Rivoltini, L. Tumour-released exosomes and their implications in cancer immunity. Cell Death Differ. 2008, 15, 80–88. [Google Scholar] [CrossRef]
- Peinado, H.; Aleckovic, M.; Lavotshkin, S.; Matei, I.; Costa-Silva, B.; Moreno-Bueno, G.; Hergueta-Redondo, M.; Williams, C.; Garcia-Santos, G.; Ghajar, C.; et al. Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through MET. Nat. Med. 2012, 18, 883–891. [Google Scholar] [CrossRef] [Green Version]
- Skog, J.; Wurdinger, T.; van Rijn, S.; Meijer, D.H.; Gainche, L.; Sena-Esteves, M.; Curry, W.T., Jr.; Carter, B.S.; Krichevsky, A.M.; Breakefield, X.O. Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat. Cell Biol. 2008, 10, 1470–1476. [Google Scholar] [CrossRef] [PubMed]
- Bobrie, A.; Krumeich, S.; Reyal, F.; Recchi, C.; Moita, L.F.; Seabra, M.C.; Ostrowski, M.; Thery, C. Rab27a supports exosome-dependent and -independent mechanisms that modify the tumor microenvironment and can promote tumor progression. Cancer Res. 2012, 72, 4920–4930. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, Y.; Konishi, Y.; Kosaka, N.; Katsuda, T.; Kato, T.; Ochiya, T. Comparative marker analysis of extracellular vesicles in different human cancer types. J. Extracell. Vesicles 2013, 2, 20424. [Google Scholar] [CrossRef]
- Kawai, A.; Naito, N.; Yoshida, A.; Morimoto, Y.; Ouchida, M.; Shimizu, K.; Beppu, Y. Establishment and characterization of a biphasic synovial sarcoma cell line, SYO-1. Cancer Lett. 2004, 204, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Naka, N.; Takenaka, S.; Araki, N.; Miwa, T.; Hashimoto, N.; Yoshioka, K.; Joyama, S.; Hamada, K.I.; Tsukamoto, Y.; Tomita, Y. Synovial sarcoma is a stem cell malignancy. Stem Cells 2010, 28, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Sonobe, H.; Manabe, Y.; Furihata, M.; Iwata, J.; Oka, T.; Ohtsuki, Y.; Mizobuchi, H.; Yamamoto, H.; Kumano, O.; Abe, S. Establishment and characterization of a new human synovial sarcoma cell line, HS-SY-II. Lab. Investig. 1992, 67, 498–505. [Google Scholar] [PubMed]
- Uotani, K.; Fujiwara, T.; Yoshida, A.; Iwata, S.; Morita, T.; Kiyono, M.; Yokoo, S.; Kunisada, T.; Takeda, K.; Hasei, J.; et al. Circulating MicroRNA-92b-3p as a Novel Biomarker for Monitoring of Synovial Sarcoma. Sci. Rep. 2017, 7, 14634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, K.; Eguchi, T.; Sogawa, C.; Calderwood, S.K.; Futagawa, J.; Kasai, T.; Seno, M.; Okamoto, K.; Sasaki, A.; Kozaki, K.I. HSP-enriched properties of extracellular vesicles involve survival of metastatic oral cancer cells. J. Cell. Biochem. 2018, 119, 7350–7362. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Yuan, Y.; Valenzuela, M.M.; Turay, D.; Ferguson, H.; Wong, S.-F.; Perez, M.; Mirshahidi, S.; Wall, N.R. Early diagnostic value of survivin and its alternative splice variants in breast cancers. Am. Soc. Clin. Oncol. 2012, 14, 176. [Google Scholar] [CrossRef]
- Moon, P.-G.; Lee, J.-E.; Cho, Y.-E.; Lee, S.J.; Chae, Y.S.; Jung, J.H.; Kim, I.-S.; Park, H.Y.; Baek, M.-C. Fibronectin on circulating extracellular vesicles as a liquid biopsy to detect breast cancer. Oncotarget 2016, 7, 40189. [Google Scholar] [CrossRef] [Green Version]
- Moon, P.-G.; Lee, J.-E.; Cho, Y.-E.; Lee, S.J.; Jung, J.H.; Chae, Y.S.; Bae, H.-I.; Kim, Y.-B.; Kim, I.-S.; Park, H.Y. Identification of developmental endothelial locus-1 on circulating extracellular vesicles as a novel biomarker for early breast cancer detection. Clin. Cancer Res. 2016, 22, 1757–1766. [Google Scholar] [CrossRef] [Green Version]
- Rupp, A.-K.; Rupp, C.; Keller, S.; Brase, J.C.; Ehehalt, R.; Fogel, M.; Moldenhauer, G.; Marmé, F.; Sültmann, H.; Altevogt, P. Loss of EpCAM expression in breast cancer derived serum exosomes: Role of proteolytic cleavage. Gynecol. Oncol. 2011, 122, 437–446. [Google Scholar] [CrossRef]
- Khan, S.; Jutzy, J.M.; Valenzuela, M.M.A.; Turay, D.; Aspe, J.R.; Ashok, A.; Mirshahidi, S.; Mercola, D.; Lilly, M.B.; Wall, N.R. Plasma-derived exosomal survivin, a plausible biomarker for early detection of prostate cancer. PLoS ONE 2012, 7, e46737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, J.; Skog, J.; Nordstrand, A.; Baranov, V.; Mincheva-Nilsson, L.; Breakefield, X.; Widmark, A. Prostate cancer-derived urine exosomes: A novel approach to biomarkers for prostate cancer. Br. J. Cancer 2009, 100, 1603–1607. [Google Scholar] [CrossRef]
- Melo, S.A.; Luecke, L.B.; Kahlert, C.; Fernandez, A.F.; Gammon, S.T.; Kaye, J.; LeBleu, V.S.; Mittendorf, E.A.; Weitz, J.; Rahbari, N. Glypican-1 identifies cancer exosomes and detects early pancreatic cancer. Nature 2015, 523, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Szajnik, M.; Derbis, M.; Lach, M.; Patalas, P.; Michalak, M.; Drzewiecka, H.; Szpurek, D.; Nowakowski, A.; Spaczynski, M.; Baranowski, W. Exosomes in plasma of patients with ovarian carcinoma: Potential biomarkers of tumor progression and response to therapy. Gynecol. Obstet. 2013. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Yang, Y.; Zeng, Y.; He, M. A microfluidic ExoSearch chip for multiplexed exosome detection towards blood-based ovarian cancer diagnosis. Lab Chip 2016, 16, 489–496. [Google Scholar] [CrossRef] [Green Version]
- Shao, H.; Chung, J.; Balaj, L.; Charest, A.; Bigner, D.D.; Carter, B.S.; Hochberg, F.H.; Breakefield, X.O.; Weissleder, R.; Lee, H. Protein typing of circulating microvesicles allows real-time monitoring of glioblastoma therapy. Nat. Med. 2012, 18, 1835. [Google Scholar] [CrossRef]
- Johnson, J.M.; Cotzia, P.; Fratamico, R.; Mikkilineni, L.; Chen, J.; Colombo, D.; Mollaee, M.; Whitaker-Menezes, D.; Domingo-Vidal, M.; Lin, Z.; et al. MCT1 in Invasive Ductal Carcinoma: Monocarboxylate Metabolism and Aggressive Breast Cancer. Front. Cell Dev. Biol. 2017, 5, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinheiro, C.; Penna, V.; Morais-Santos, F.; Abrahão-Machado, L.F.; Ribeiro, G.; Curcelli, E.C.; Olivieri, M.V.; Morini, S.; Valença, I.; Ribeiro, D. Characterization of monocarboxylate transporters (MCTs) expression in soft tissue sarcomas: Distinct prognostic impact of MCT1 sub-cellular localization. J. Transl. Med. 2014, 12, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diehl, K.; Dinges, L.; Helm, O.; Ammar, N.; Plundrich, D.; Arlt, A.; Röcken, C.; Sebens, S.; Schäfer, H. Nuclear factor E2-related factor-2 has a differential impact on MCT1 and MCT4 lactate carrier expression in colonic epithelial cells: A condition favoring metabolic symbiosis between colorectal cancer and stromal cells. Oncogene 2018, 37, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Latif, A.; Chadwick, A.L.; Kitson, S.J.; Gregson, H.J.; Sivalingam, V.N.; Bolton, J.; McVey, R.J.; Roberts, S.A.; Marshall, K.M.; Williams, K.J. Monocarboxylate Transporter 1 (MCT1) is an independent prognostic biomarker in endometrial cancer. BMC Clin. Pathol. 2017, 17, 27. [Google Scholar] [CrossRef]
- Pinheiro, C.; Longatto-Filho, A.; Azevedo-Silva, J.; Casal, M.; Schmitt, F.C.; Baltazar, F. Role of monocarboxylate transporters in human cancers: State of the art. J. Bioenerg. Biomembr. 2012, 44, 127–139. [Google Scholar] [CrossRef]
- Sanità, P.; Capulli, M.; Teti, A.; Galatioto, G.P.; Vicentini, C.; Chiarugi, P.; Bologna, M.; Angelucci, A. Tumor-stroma metabolic relationship based on lactate shuttle can sustain prostate cancer progression. BMC Cancer 2014, 14, 154. [Google Scholar] [CrossRef]
- Witkiewicz, A.K.; Kline, J.; Queenan, M.; Brody, J.R.; Tsirigos, A.; Bilal, E.; Pavlides, S.; Ertel, A.; Sotgia, F.; Lisanti, M.P. Molecular profiling of a lethal tumor microenvironment, as defined by stromal caveolin-1 status in breast cancers. Cell Cycle 2011, 10, 1794–1809. [Google Scholar] [CrossRef] [Green Version]
- Fiaschi, T.; Marini, A.; Giannoni, E.; Taddei, M.L.; Gandellini, P.; de Donatis, A.; Lanciotti, M.; Serni, S.; Cirri, P.; Chiarugi, P. Reciprocal metabolic reprogramming through lactate shuttle coordinately influences tumor-stroma interplay. Cancer Res. 2012, 72, 5130–5140. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, E.C.; van Houten, B. Metabolic symbiosis in cancer: Refocusing the Warburg lens. Mol. Carcinog. 2013, 52, 329–337. [Google Scholar] [CrossRef]
- Sandforth, L.; Ammar, N.; Dinges, L.A.; Röcken, C.; Arlt, A.; Sebens, S.; Schäfer, H. Impact of the Monocarboxylate Transporter-1 (MCT1)-Mediated Cellular Import of Lactate on Stemness Properties of Human Pancreatic Adenocarcinoma Cells. Cancers 2020, 12, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takada, T.; Takata, K.; Ashihara, E. Inhibition of monocarboxylate transporter 1 suppresses the proliferation of glioblastoma stem cells. J. Physiol. Sci. 2016, 66, 387–396. [Google Scholar] [CrossRef]
- Thakur, A.; Qiu, G.; Xu, C.; Han, X.; Yang, T.; Ng, S.; Chan, K.; Wu, C.; Lee, Y. Label-free sensing of exosomal MCT1 and CD147 for tracking metabolic reprogramming and malignant progression in glioma. Sci. Adv. 2020, 6, eaaz6119. [Google Scholar] [CrossRef]
- Alix-Panabières, C.; Pantel, K. Circulating tumor cells: Liquid biopsy of cancer. Clin. Chem. 2013, 59, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Baccelli, I.; Schneeweiss, A.; Riethdorf, S.; Stenzinger, A.; Schillert, A.; Vogel, V.; Klein, C.; Saini, M.; Bäuerle, T.; Wallwiener, M. Identification of a population of blood circulating tumor cells from breast cancer patients that initiates metastasis in a xenograft assay. Nat. Biotechnol. 2013, 31, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Dive, C.; Brady, G. SnapShot: Circulating tumor cells. Cell 2017, 168, 742–742.e1. [Google Scholar] [CrossRef]
- Lucci, A.; Hall, C.S.; Lodhi, A.K.; Bhattacharyya, A.; Anderson, A.E.; Xiao, L.; Bedrosian, I.; Kuerer, H.M.; Krishnamurthy, S. Circulating tumour cells in non-metastatic breast cancer: A prospective study. Lancet Oncol. 2012, 13, 688–695. [Google Scholar] [CrossRef]
- Plaks, V.; Koopman, C.D.; Werb, Z. Circulating tumor cells. Science 2013, 341, 1186–1188. [Google Scholar] [CrossRef]
- Wang, J.; Ni, J.; Beretov, J.; Thompson, J.; Graham, P.; Li, Y. Exosomal microRNAs as liquid biopsy biomarkers in prostate cancer. Crit. Rev. Oncol. Hematol. 2020, 145, 102860. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Age (years) | Sex | Site | Size (cm) | Subtype | Fusion Gene | CTx | RTx | Viable Cells (%) | Local Recurrence | Distant Metastasis | Follow-Up Period (Months) | Oncologic Outcome |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 21 | M | Knee | 4.5 | Mono-phasic | SY18-SSX1 | Yes | No | 95 | Absence | − | 47 | CDF |
| 2 | 44 | F | Thigh | 9.4 | Mono-phasic | SY18-SSX1 | Yes | No | 50 | Absence | + | 23 | DOD |
| 3 | 64 | M | Forearm | 5.7 | Mono-phasic | Unknown | No | No | - | Absence | − | 53 | CDF |
| 4 | 48 | M | Thigh | 14.8 | Bi-phasic | SY18-SSX1 | Yes | No | 95 | Absence | − | 55 | CDF |
| 5 | 11 | M | Lumbar | 4.3 | Mono-phasic | SY18-SSX1 | Yes | No | 95 | Absence | − | 54 | CDF |
| 6 | 18 | M | Thigh | 13.7 | Mono-phasic | Unknown | Yes | No | 95 | Absence | − | 49 | CDF |
| 7 | 61 | F | Thigh | 6.6 | Bi-phasic | Unknown | Yes | No | - | Absence | + | 40 | AWD |
| 8 | 20 | F | Back | 8.0 | Mono-phasic | Unknown | Yes | No | 75 | Absence | − | 35 | CDF |
| 9 | 55 | M | Abdomen | 4.2 | Mono-phasic | SY18-SSX1 | No | No | - | Absence | − | 11 | CDF |
| 10 | 11 | F | Groin | 9.5 | Mono-phasic | SY18-SSX2 | Yes | No | 100 | Absence | − | 9 | CDF |
| Variables | Nuclear (N) | Cytoplasm/Membrane (N) | p-Value |
|---|---|---|---|
| Age at diagnosis | - | - | 0.053 |
| 0–40 years | 7 | 10 | - |
| 41+ years | 10 | 2 | - |
| Sex | - | - | 0.264 |
| Male | 7 | 8 | - |
| Female | 10 | 4 | - |
| Subtype | - | - | 0.439 |
| Monophasic | 9 | 8 | - |
| Biphasic | 6 | 4 | - |
| Unknown | 2 | 0 | - |
| Size | - | - | 0.519 |
| ≤5cm | 3 | 3 | - |
| >5cm | 13 | 7 | - |
| Unknown | 1 | 2 | - |
| Site | - | - | 0.566 |
| Thigh | 9 | 8 | - |
| Lower leg | 1 | 0 | - |
| Upper arm | 0 | 1 | - |
| Forearm | 3 | 1 | - |
| Others | 4 | 2 | - |
| Fusion gene | - | - | 0.003 |
| SS18-SSX1 | 14 | 2 | - |
| SS18-SSX2 | 3 | 8 | - |
| Unknown | 0 | 2 | - |
| Chemotherapy | - | - | 0.218 |
| Yes | 14 | 7 | - |
| No | 3 | 5 | - |
| Radiation therapy | - | - | 0.370 |
| Yes | 4 | 1 | - |
| No | 13 | 11 | - |
| Local recurrence | - | - | 0.498 |
| Presence | 2 | 0 | - |
| Absence | 15 | 12 | - |
| Distant metastasis | - | - | 0.025 |
| Presence | 5 | 9 | - |
| Absence | 12 | 3 | - |
| Oncologic outcome | - | - | 0.010 |
| DOD | 4 | 9 | - |
| CDF/NED | 13 | 3 | - |
| Variables | N | Univariate | Multivariate | |||
|---|---|---|---|---|---|---|
| 5-Year OS | p-Value | HR | 95% CI | p-Value | ||
| Age at diagnosis | - | - | 0.986 | - | - | - |
| 0–40 years | 17 | 58% | - | - | - | - |
| 41+ years | 12 | 58% | - | - | - | - |
| Sex | - | - | 0.041 | - | - | - |
| Female | 14 | 70% | - | 1 | Reference | - |
| Male | 15 | 47% | - | 2.49 | 0.74–8.39 | 0.141 |
| Subtype | - | - | 0.131 | - | - | - |
| Monophasic | 17 | 40% | - | - | - | - |
| Biphasic | 10 | 80% | - | - | - | - |
| Unknown | 2 | 100% | - | - | - | - |
| Size | - | - | 0.266 | - | - | - |
| ≤5 cm | 6 | 83% | - | - | - | - |
| >5 cm | 20 | 53% | - | - | - | - |
| Unknown | 3 | 33% | - | - | - | - |
| Site | - | - | 0.735 | - | - | - |
| Lower extremity | 18 | 60% | - | - | - | - |
| Upper extremity | 5 | 40% | - | - | - | - |
| Others | 6 | 67% | - | - | - | - |
| Fusion gene | - | - | 0.408 | - | - | - |
| SS18-SSX1 | 16 | 68% | - | - | - | - |
| SS18-SSX2 | 11 | 44% | - | - | - | - |
| Unknown | 2 | 50% | - | - | - | - |
| Chemotherapy | - | - | 0.005 | - | - | - |
| Yes | 21 | 80% | - | 1 | Reference | - |
| No | 8 | 25% | - | 7.99 | 1.96–32.54 | 0.004 |
| Radiation therapy | - | - | 0.821 | - | - | - |
| Yes | 5 | 40% | - | - | - | - |
| No | 24 | 62% | - | - | - | - |
| Local recurrence | - | - | 0.240 | - | - | - |
| Absence | 27 | 55% | - | - | - | - |
| Presence | 2 | 100% | - | - | - | - |
| Distant metastasis | - | - | 0.010 | - | - | - |
| Absence | 15 | 79% | - | 1 | Reference | - |
| Presence | 14 | 36% | - | 2.94 | 0.71–12.22 | 0.138 |
| MCT1 expression | - | - | 0.002 | - | - | - |
| Nuclear | 17 | 75% | - | 1 | Reference | - |
| Cytoplasm/membrane | 12 | 33% | - | 5.34 | 1.20–23.89 | 0.028 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yokoo, S.; Fujiwara, T.; Yoshida, A.; Uotani, K.; Morita, T.; Kiyono, M.; Hasei, J.; Nakata, E.; Kunisada, T.; Iwata, S.; et al. Liquid Biopsy Targeting Monocarboxylate Transporter 1 on the Surface Membrane of Tumor-Derived Extracellular Vesicles from Synovial Sarcoma. Cancers 2021, 13, 1823. https://doi.org/10.3390/cancers13081823
Yokoo S, Fujiwara T, Yoshida A, Uotani K, Morita T, Kiyono M, Hasei J, Nakata E, Kunisada T, Iwata S, et al. Liquid Biopsy Targeting Monocarboxylate Transporter 1 on the Surface Membrane of Tumor-Derived Extracellular Vesicles from Synovial Sarcoma. Cancers. 2021; 13(8):1823. https://doi.org/10.3390/cancers13081823
Chicago/Turabian StyleYokoo, Suguru, Tomohiro Fujiwara, Aki Yoshida, Koji Uotani, Takuya Morita, Masahiro Kiyono, Joe Hasei, Eiji Nakata, Toshiyuki Kunisada, Shintaro Iwata, and et al. 2021. "Liquid Biopsy Targeting Monocarboxylate Transporter 1 on the Surface Membrane of Tumor-Derived Extracellular Vesicles from Synovial Sarcoma" Cancers 13, no. 8: 1823. https://doi.org/10.3390/cancers13081823
APA StyleYokoo, S., Fujiwara, T., Yoshida, A., Uotani, K., Morita, T., Kiyono, M., Hasei, J., Nakata, E., Kunisada, T., Iwata, S., Yonemoto, T., Ueda, K., & Ozaki, T. (2021). Liquid Biopsy Targeting Monocarboxylate Transporter 1 on the Surface Membrane of Tumor-Derived Extracellular Vesicles from Synovial Sarcoma. Cancers, 13(8), 1823. https://doi.org/10.3390/cancers13081823