
Circular RNA in Chemonaive Lymph Node Negative Colon Cancer Patients

 ,
, 
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population and Patient Selection
2.2. Sample Collection and Processing
2.2.1. Microsatellite Instability
2.2.2. Consensus Molecular Subtypes
2.2.3. Identification of circRNAs
2.2.4. Multiple Correspondence Analysis (MCA)
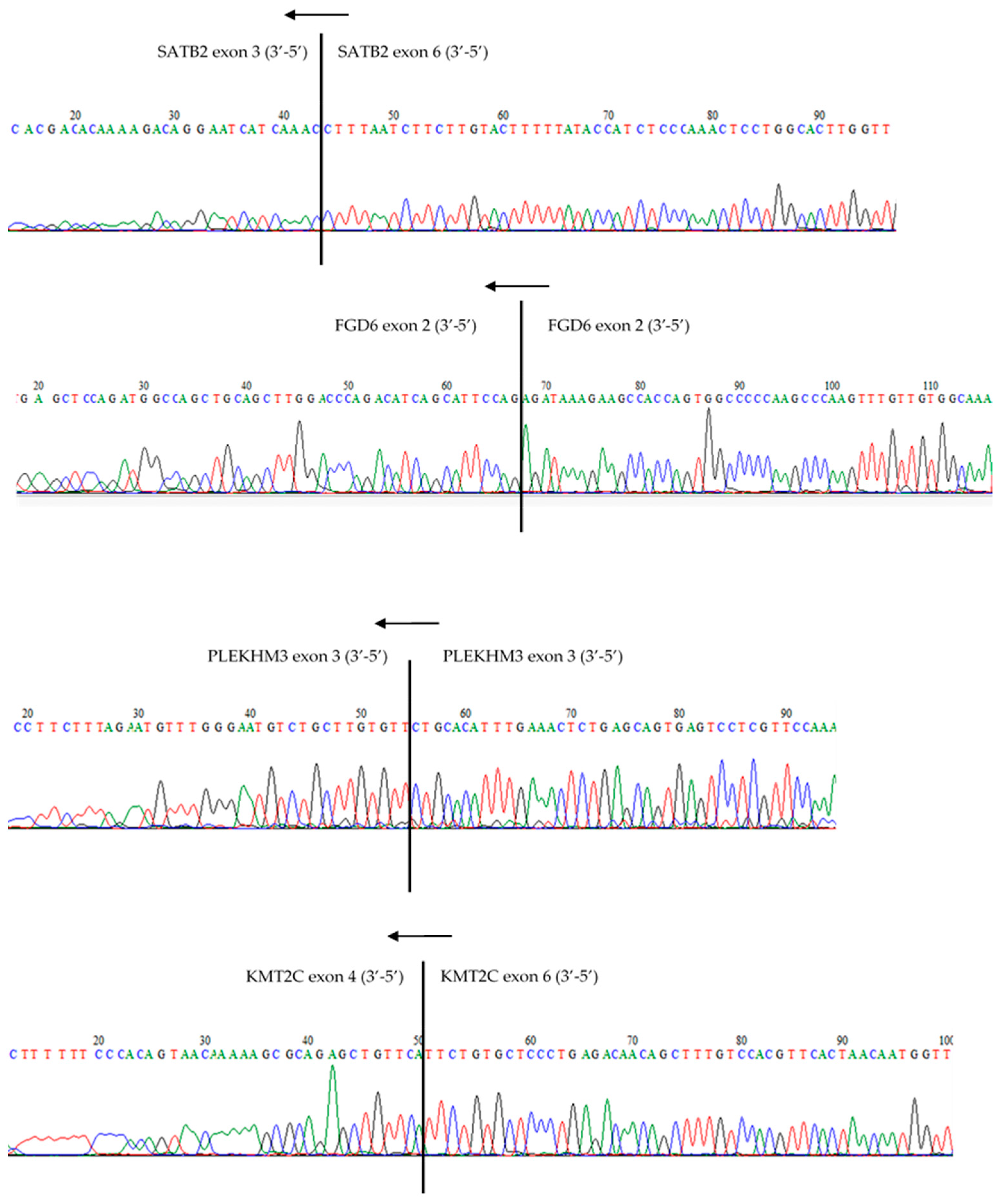
2.2.5. Reverse Transcription, Quantitative PCR, and Sanger Sequencing
2.3. Statistical Analyses
3. Results
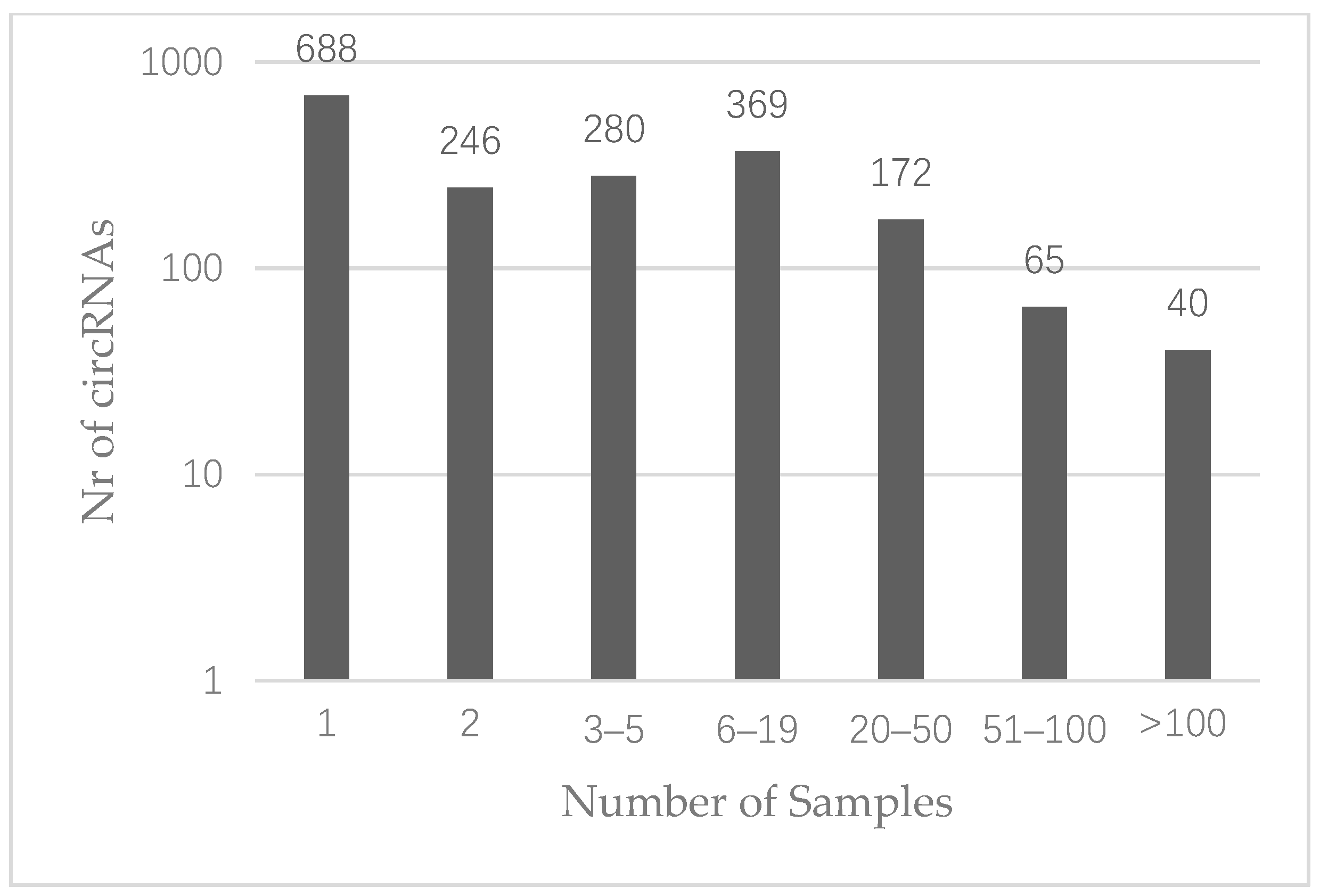
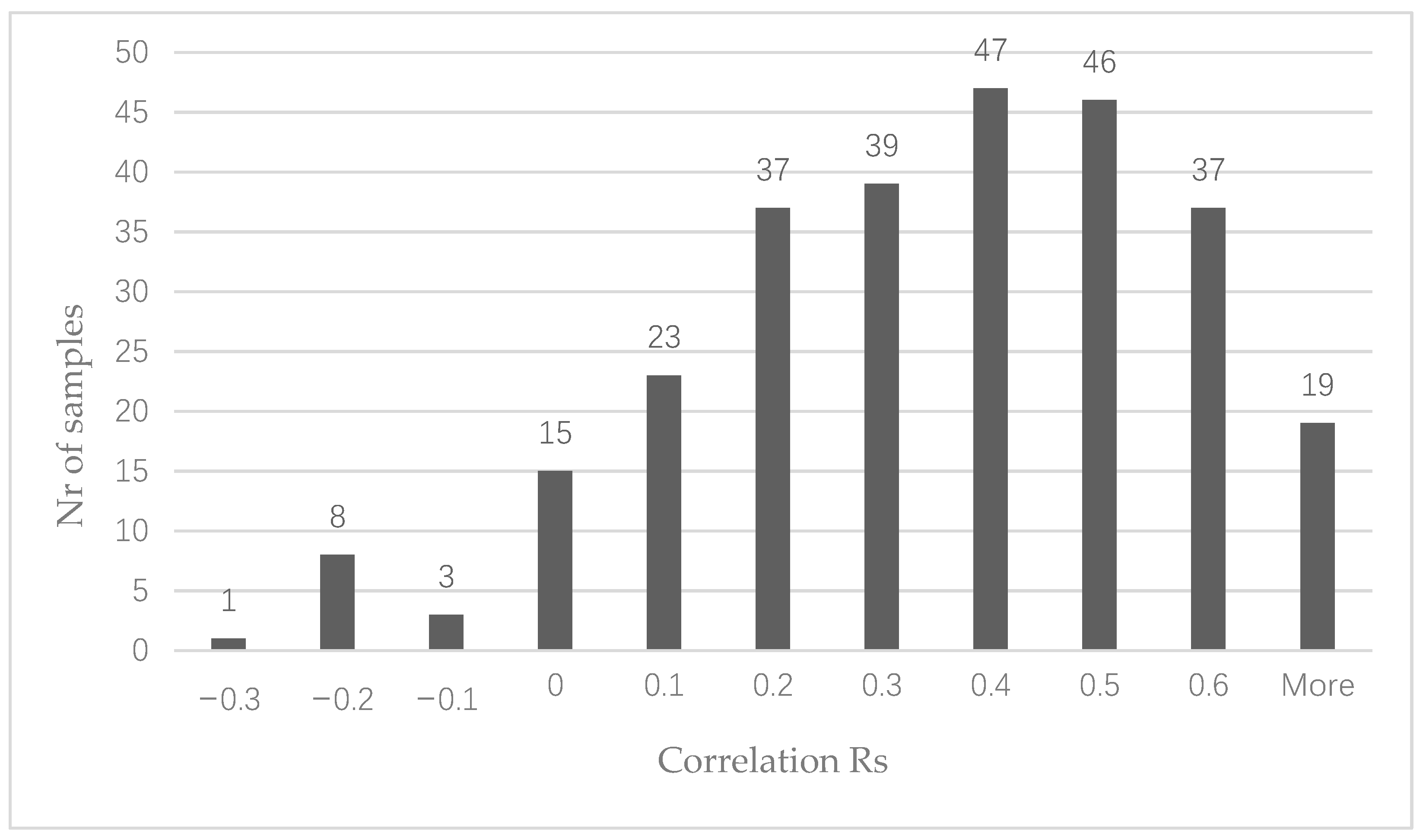
3.1. CircRNA Expression in Colon Cancer
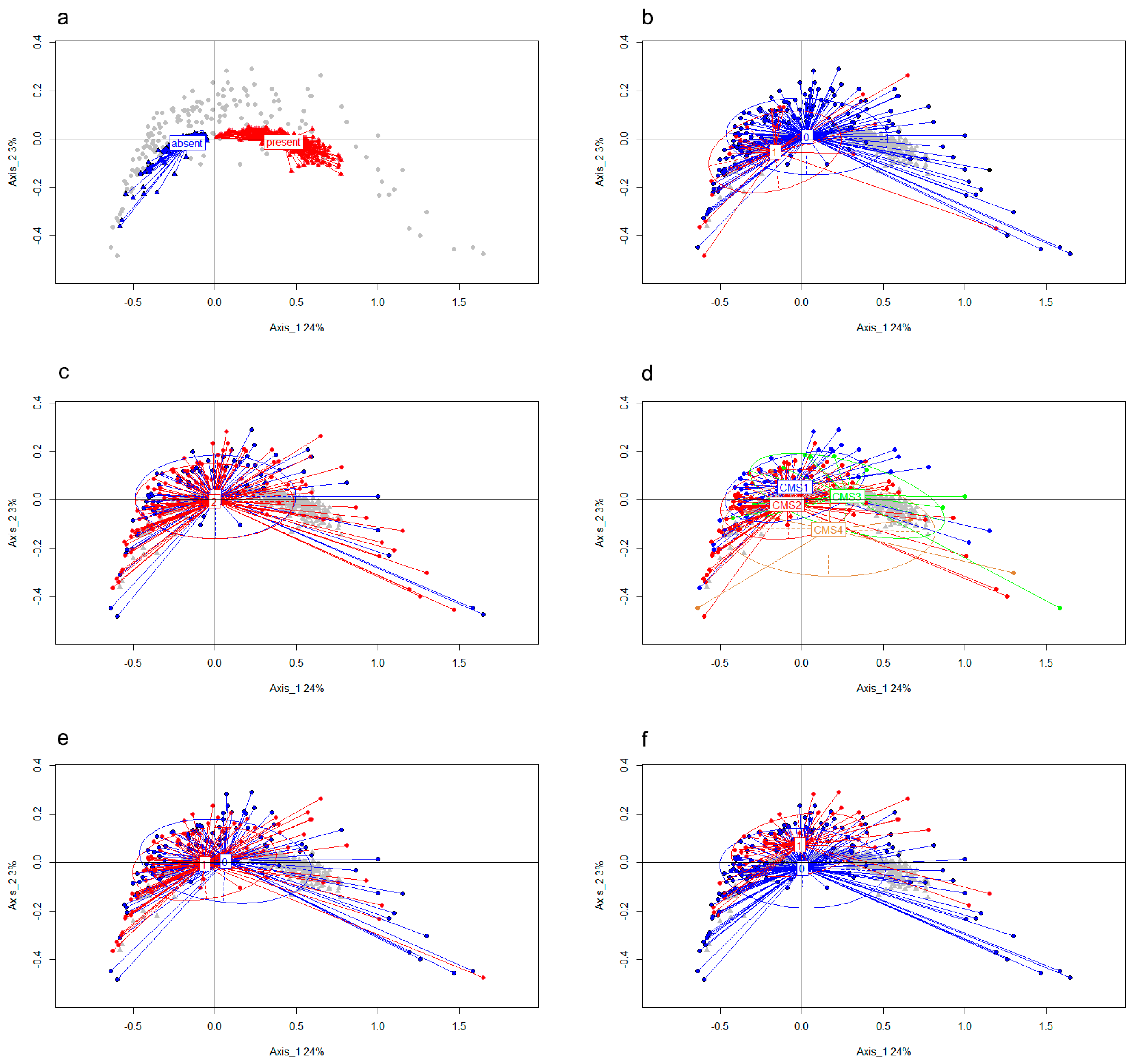
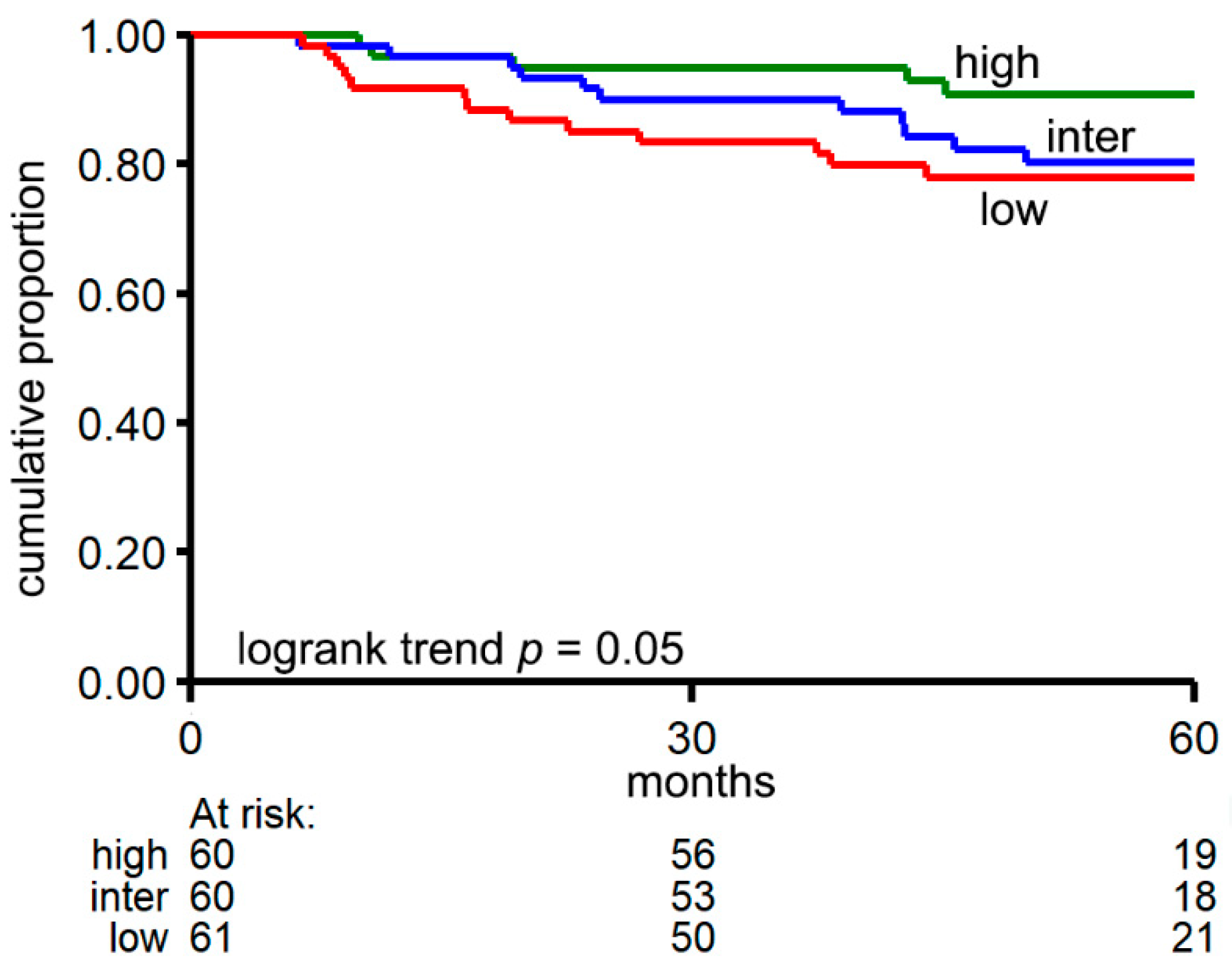
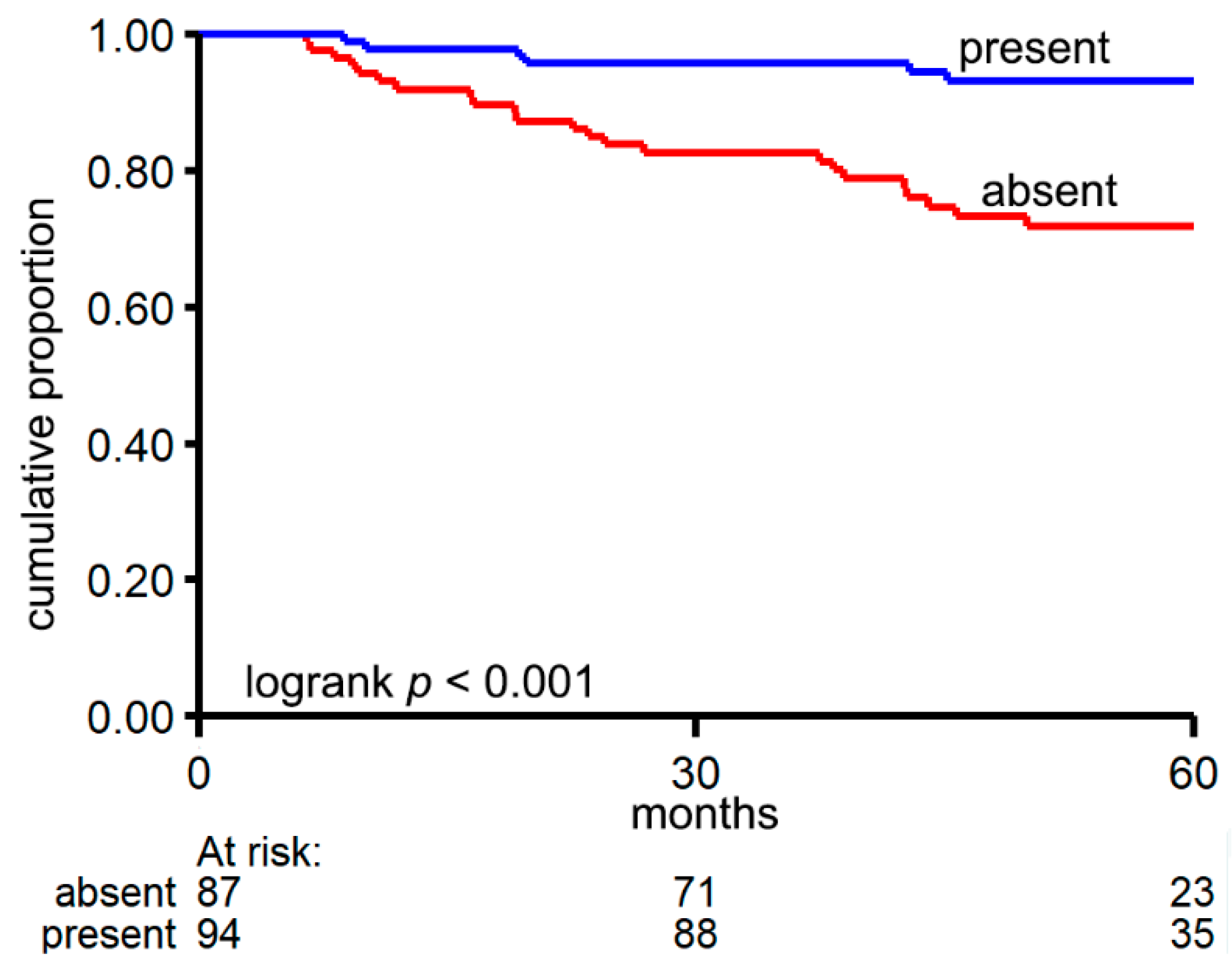
3.2. CircRNA Expression Patterns Are Associated with Relevant Clinical Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cijfers over kanker. Available online: https://iknl.nl/en/ncr/ncr-data-figures (accessed on 23 May 2019).
- Sargent, D.J.; Patiyil, S.; Yothers, G.; Haller, D.G.; Gray, R.; Benedetti, J.; Buyse, M.; Labianca, R.; Seitz, J.F.; O’Callaghan, C.J.; et al. End Points for Colon Cancer Adjuvant Trials: Observations and Recommendations Based on Individual Patient Data From 20,898 Patients Enrolled Onto 18 Randomized Trials From the ACCENT Group. J. Clin. Oncol. 2007, 25, 4569–4574. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Iyer, M.K.; Niknafs, Y.S.; Malik, R.; Singhal, U.; Sahu, A.; Hosono, Y.; Barrette, T.R.; Prensner, J.R.; Evans, J.R.; Zhao, S.; et al. The landscape of long noncoding RNAs in the human transcriptome. Nat. Genet. 2015, 47, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Muers, M.R. Genome-wide views of long non-coding rnas. Nat. Rev. Genet. 2011, 12, 742–743. [Google Scholar] [CrossRef]
- Sanger, H.L.; Klotz, G.; Riesner, D.; Gross, H.J.; Kleinschmidt, A.K. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc. Natl. Acad. Sci. USA 1976, 73, 3852–3856. [Google Scholar] [CrossRef] [Green Version]
- Yan, B.; Gu, W.; Yang, Z.; Gu, Z.; Yue, X.; Gu, Q.; Liu, L. Downregulation of a long noncoding rna-ncrupar contributes to tumor inhibition in colorectal cancer. Tumour Biol. 2014, 35, 11329–11335. [Google Scholar] [CrossRef]
- Cao, Y.; Lin, M.; Bu, Y.; Ling, H.; He, Y.; Huang, C.; Shen, Y.; Song, B.; Cao, D. p53-inducible long non-coding RNA PICART1 mediates cancer cell proliferation and migration. Int. J. Oncol. 2017, 50, 1671–1682. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Zhang, R.; Zhao, J. The Novel Long Noncoding RNA TUSC7 Inhibits Proliferation by Sponging MiR-211 in Colorectal Cancer. Cell. Physiol. Biochem. 2017, 41, 635–644. [Google Scholar] [CrossRef]
- Jacob, H.; Stanisavljevic, L.; Storli, K.E.; Hestetun, K.E.; Dahl, O.; Myklebust, M.P. A four-microrna classifier as a novel prognostic marker for tumor recurrence in stage ii colon cancer. Sci. Rep. 2018, 8, 6157. [Google Scholar] [CrossRef] [PubMed]
- Bullock, M.D.; Pickard, K.; Mitter, R.; Sayan, A.E.; Primrose, J.N.; Ivan, C.; Calin, G.A.; Thomas, G.J.; Packham, G.K.; Mirnezami, A.H. Stratifying risk of recurrence in stage II colorectal cancer using deregulated stromal and epithelial microRNAs. Oncotarget 2015, 6, 7262–7279. [Google Scholar] [CrossRef] [Green Version]
- Grassi, A.; Perilli, L.; Albertoni, L.; Tessarollo, S.; Mescoli, C.; Urso, E.D.L.; Fassan, M.; Rugge, M.; Zanovello, P. A coordinate deregulation of microRNAs expressed in mucosa adjacent to tumor predicts relapse after resection in localized colon cancer. Mol. Cancer 2018, 17. [Google Scholar] [CrossRef] [Green Version]
- Su, M.; Xiao, Y.; Ma, J.; Tang, Y.; Tian, B.; Zhang, Y.; Li, X.; Wu, Z.; Yang, D.; Zhou, Y.; et al. Circular RNAs in Cancer: Emerging functions in hallmarks, stemness, resistance and roles as potential biomarkers. Mol. Cancer 2019, 18, 1–17. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nat. Cell Biol. 2013, 495, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.-O.; Chen, T.; Xiang, J.-F.; Yin, Q.-F.; Xing, Y.-H.; Zhu, S.; Yang, L.; Chen, L.-L. Circular Intronic Long Noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. Circrna biogenesis competes with pre-mrna splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nat. Cell Biol. 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Yang, X.; Li, X.; Wang, J.; Gao, Y.; Shang, R.; Sun, W.; Dou, K.; Li, H. Circular RNA: A new star of noncoding RNAs. Cancer Lett. 2015, 365, 141–148. [Google Scholar] [CrossRef]
- Zhang, L. Circular RNA: The main regulator of energy metabolic reprogramming in cancer cells. Thorac. Cancer 2019, 11, 6–7. [Google Scholar] [CrossRef]
- Artemaki, P.I.; Scorilas, A.; Kontos, C.K. Circular RNAs: A New Piece in the Colorectal Cancer Puzzle. Cancers 2020, 12, 2464. [Google Scholar] [CrossRef]
- Hsu, M.-T.; Coca-Prados, M. Electron microscopic evidence for the circular form of RNA in the cytoplasm of eukaryotic cells. Nat. Cell Biol. 1979, 280, 339–340. [Google Scholar] [CrossRef]
- Wilusz, J.E.; Sharp, P.A. A Circuitous Route to Noncoding RNA. Science 2013, 340, 440–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocquerelle, C.; Mascrez, B.; Hétuin, D.; Bailleul, B. Mis-splicing yields circular RNA molecules. FASEB J. 1993, 7, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Salzman, J.; Chen, R.E.; Olsen, M.N.; Wang, P.L.; Brown, P.O. Cell-type specific features of circular rna expression. PLoS Genet. 2013, 9, e1003777. [Google Scholar] [CrossRef]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2012, 19, 141–157. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Xin, Y. Circular RNAs: A new frontier for cancer diagnosis and therapy. J. Hematol. Oncol. 2018, 11. [Google Scholar] [CrossRef] [Green Version]
- Karousi, P.; Artemaki, P.I.; Sotiropoulou, C.D.; Christodoulou, S.; Scorilas, A.; Kontos, C.K. Identification of Two Novel Circular RNAs Deriving from BCL2L12 and Investigation of Their Potential Value as a Molecular Signature in Colorectal Cancer. Int. J. Mol. Sci. 2020, 21, 8867. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, K.; Tan, S.; Xin, J.; Yuan, Q.; Xu, H.; Xu, X.; Liang, Q.; Christiani, D.C.; Wang, M.; et al. Circular RNAs in body fluids as cancer biomarkers: The new frontier of liquid biopsies. Mol. Cancer 2021, 20, 1–10. [Google Scholar] [CrossRef] [PubMed]
- van Vugt, J.L.; Braak, R.R.C.V.D.; Lalmahomed, Z.S.; Vrijland, W.W.; Dekker, J.W.; Zimmerman, D.D.; Vles, W.J.; Coene, P.-P.L.; Ijzermans, J.N. Impact of low skeletal muscle mass and density on short and long-term outcome after resection of stage I-III colorectal cancer. Eur. J. Surg. Oncol. 2018, 44, 1354–1360. [Google Scholar] [CrossRef] [PubMed]
- Sieuwerts, A.M.; Lyng, M.B.; Gelder, M.E.M.-V.; De Weerd, V.; Sweep, F.C.; Foekens, J.A.; Span, P.N.; Martens, J.W.; Ditzel, H.J. Evaluation of the ability of adjuvant tamoxifen-benefit gene signatures to predict outcome of hormone-naive estrogen receptor-positive breast cancer patients treated with tamoxifen in the advanced setting. Mol. Oncol. 2014, 8, 1679–1689. [Google Scholar] [CrossRef] [Green Version]
- Sieuwerts, A.M.; Gelder, M.E.M.-V.; Timmermans, M.; Trapman, A.M.; Garcia, R.R.; Arnold, M.; Goedheer, A.J.; Portengen, H.; Klijn, J.G.; Foekens, J.A. How ADAM-9 and ADAM-11 Differentially from Estrogen Receptor Predict Response to Tamoxifen Treatment in Patients with Recurrent Breast Cancer: A Retrospective Study. Clin. Cancer Res. 2005, 11, 7311–7321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coebergh van den Braak, R.R.J.; Sieuwerts, A.M.; Kandimalla, R.; Lalmahomed, Z.S.; Bril, S.I.; Van Galen, A.; Smid, M.; Biermann, K.; Van Krieken, J.H.J.M.; Kloosterman, W.P.; et al. High mRNA expression of splice variant SYK short correlates with hepatic disease progression in chemonaive lymph node negative colon cancer patients. PLoS ONE 2017, 12, e0185607. [Google Scholar] [CrossRef] [Green Version]
- Smid, M.; Coebergh van den Braak, R.R.J.; van de Werken, H.J.G.; van Riet, J.; van Galen, A.; de Weerd, V.; van der Vlugt-Daane, M.; Bril, S.I.; Lalmahomed, Z.S.; Kloosterman, W.P.; et al. Gene length corrected trimmed mean of m-values (getmm) processing of rna-seq data performs similarly in intersample analyses while improving intrasample comparisons. BMC Bioinform. 2018, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacher, J.W.; Flanagan, L.A.; Smalley, R.L.; Nassif, N.A.; Burgart, L.J.; Halberg, R.B.; Megid, W.M.A.; Thibodeau, S.N. Development of a Fluorescent Multiplex Assay for Detection of MSI-High Tumors. Dis. Markers 2004, 20, 237–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guinney, J.; Dienstmann, R.; Wang, X.; De Reyniès, A.; Schlicker, A.; Soneson, C.; Marisa, L.; Roepman, P.; Nyamundanda, G.; Angelino, P.; et al. The consensus molecular subtypes of colorectal cancer. Nat. Med. 2015, 21, 1350–1356. [Google Scholar] [CrossRef] [PubMed]
- Smid, M.; Wilting, S.M.; Uhr, K.; Rodríguez-González, F.G.; De Weerd, V.; Der Smissen, W.J.P.-V.; Van Der Vlugt-Daane, M.; Van Galen, A.; Nik-Zainal, S.; Butler, A.; et al. The circular RNome of primary breast cancer. Genome Res. 2019, 29, 356–366. [Google Scholar] [CrossRef]
- Lu, H.; Yao, B.; Wen, X.; Jia, B. FBXW7 circular RNA regulates proliferation, migration and invasion of colorectal carcinoma through NEK2, mTOR, and PTEN signaling pathways in vitro and in vivo. BMC Cancer 2019, 19, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glažar, P.; Papavasileiou, P.; Rajewsky, N. circBase: A database for circular RNAs. RNA 2014, 20, 1666–1670. [Google Scholar] [CrossRef] [Green Version]
- Gurzu, S.; Turdean, S.; Kovecsi, A.; Contac, A.O.; Jung, I. Epithelial-mesenchymal, mesenchymal-epithelial, and endothelial-mesenchymal transitions in malignant tumors: An update. World J. Clin. Cases 2015, 3, 393–404. [Google Scholar] [CrossRef]
- Cai, J.; Chen, Z.; Zuo, X. Circsmarca5 functions as a diagnostic and prognostic biomarker for gastric cancer. Dis. Markers 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Barbagallo, D.; Caponnetto, A.; Brex, D.; Mirabella, F.; Barbagallo, C.; Lauretta, G.; Morrone, A.; Certo, F.; Broggi, G.; Caltabiano, R.; et al. CircSMARCA5 Regulates VEGFA mRNA Splicing and Angiogenesis in Glioblastoma Multiforme Through the Binding of SRSF. Cancers 2019, 11, 194. [Google Scholar] [CrossRef] [Green Version]
- Kong, Z.; Wan, X.; Zhang, Y.; Zhang, P.; Zhang, Y.; Zhang, X.; Qi, X.; Wu, H.; Huang, J.; Li, Y. Androgen-responsive circular RNA circSMARCA5 is up-regulated and promotes cell proliferation in prostate cancer. Biochem. Biophys. Res. Commun. 2017, 493, 1217–1223. [Google Scholar] [CrossRef]
- Zeng, K.; Chen, X.; Xu, M.; Liu, X.; Hu, X.; Xu, T.; Sun, H.; Pan, Y.; He, B.; Wang, S. CircHIPK3 promotes colorectal cancer growth and metastasis by sponging miR. Cell Death Dis. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Vo, J.N.; Cieslik, M.; Zhang, Y.; Shukla, S.; Xiao, L.; Zhang, Y.; Wu, Y.-M.; Dhanasekaran, S.M.; Engelke, C.G.; Cao, X.; et al. The Landscape of Circular RNA in Cancer. Cell 2019, 176, 869–881.e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathsyaraja, H.; Catchpole, J.; Eastwood, E.; Babaeva, E.; Geuenich, M.; Cheng, P.F.; Freie, B.; Ayers, J.; Yu, M.; Wu, N.; et al. Loss of mga mediated polycomb repression promotes tumor progression and invasiveness. bioRxiv 2020. [Google Scholar] [CrossRef]
- Zhang, N.; Nan, A.; Chen, L.; Li, X.; Jia, Y.; Qiu, M.; Dai, X.; Zhou, H.; Zhu, J.; Zhang, H.; et al. Circular RNA circSATB2 promotes progression of non-small cell lung cancer cells. Mol. Cancer 2020, 19, 1–16. [Google Scholar] [CrossRef]
- Wu, L.; Hui, H.; Wang, L.-J.; Wang, H.; Liu, Q.-F.; Han, S.-X. MicroRNA-326 functions as a tumor suppressor in colorectal cancer by targeting the nin one binding protein. Oncol. Rep. 2015, 33, 2309–2318. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhao, Y.; Kong, P.; Han, M.; Li, B. Expression of circZNF609 is Down-Regulated in Colorectal Cancer Tissue and Promotes Apoptosis in Colorectal Cancer Cells by Upregulating p. Med. Sci. Monit. 2019, 25, 5977–5985. [Google Scholar] [CrossRef]
- Dai, J.; Zhuang, Y.; Tang, M.; Qian, Q.; Chen, J.P. Circrna ubap2 facilitates the progression of colorectal cancer by regulating mir-199a/vegfa pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 7963–7971. [Google Scholar] [PubMed]
- Buratin, A.; Paganin, M.; Gaffo, E.; Molin, A.D.; Roels, J.; Germano, G.; Siddi, M.T.; Serafin, V.; De Decker, M.; Gachet, S.; et al. Large-scale circular RNA deregulation in T-ALL: Unlocking unique ectopic expression of molecular subtypes. Blood Adv. 2020, 4, 5902–5914. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Huang, H.; Jin, L.; Zhang, F.; Zeng, M.; Wei, L.; Tang, S.; Chen, D.; Wang, W. CircZNF609 enhances hepatocellular carcinoma cell proliferation, metastasis, and stemness by activating the Hedgehog pathway through the regulation of miR-15a-5p/15b-5p and GLI2 expressions. Cell Death Dis. 2020, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, Y.; Yu, M. CircRNA ZNF609 Knockdown Suppresses Cell Growth via Modulating miR-188/ELF2 Axis in Nasopharyngeal Carcinoma. Oncotargets 2020, 13, 2399–2409. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Pan, H.-M.; Xin, L.; Zhang, Y.; Zhang, W.-M.; Cao, P.; Xu, H.-W. Circ-ZNF609 promotes carcinogenesis of gastric cancer cells by inhibiting miRNA-145-5p expression. Eur. Rev. Med. Pharm. Sci. 2019, 23, 9411–9417. [Google Scholar]
- Zhu, L.; Liu, Y.; Yang, Y.; Mao, X.-M.; Yin, Z.-D. CircRNA ZNF609 promotes growth and metastasis of nasopharyngeal carcinoma by competing with microRNA-150-5p. Eur. Rev. Med. Pharm. Sci. 2019, 23, 2817–2826. [Google Scholar]
- Rossi, F.; Legnini, I.; Megiorni, F.; Colantoni, A.; Santini, T.; Morlando, M.; Di Timoteo, G.; Dattilo, D.; Dominici, C.; Bozzoni, I. Circ-ZNF609 regulates G1-S progression in rhabdomyosarcoma. Oncogene 2019, 38, 3843–3854. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Ma, Y.; Zhang, H.; Lu, Q.-J.; Yang, L.; Jiang, G.-N.; Liao, W.-L. HMGA2 regulates circular RNA ASPH to promote tumor growth in lung adenocarcinoma. Cell Death Dis. 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Yan, D.; Dong, W.; Bi, J.; Huang, L.; Yang, M.; Huang, J.; Qin, H.; Lin, T. circRNA circFUT8 Upregulates Krüpple-like Factor 10 to Inhibit the Metastasis of Bladder Cancer via Sponging miR-570-3p. Mol. Oncol. 2020, 16, 172–187. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Variables | Categories | n = 181 | % |
|---|---|---|---|
| Gender | Female | 92 | (50.9) |
| Male | 89 | (49.2) | |
| Age (median, IQR) | 70 | (63–76) | |
| Tumor stage | Stage I | 66 | (36.5) |
| Stage II | 115 | (63.5) | |
| T status | T2 | 66 | (36.5) |
| T3 | 110 | (60.8) | |
| T4 | 5 | (2.8) | |
| Nodal status | N0 ≥ 10 nodes assessed | 149 | (82.2) |
| N0 < 10 nodes assessed | 32 | (17.3) | |
| Tumor grade | Good | 16 | (8.8) |
| Poor | 10 | (5.5) | |
| Moderate | 152 | (84) | |
| Unknown | 3 | (1.7) | |
| Location | Right | 92 | (50.8) |
| Left | 89 | (49.2) | |
| MSI status | MSI | 44 | (24.3) |
| MSS | 137 | (75.7) | |
| Relapse | No | 152 | (84) |
| Yes | 29 | (16) |
| Circular Region | Ensembl Gene ID | Gene | Exons | Nr. of Samples | R * |
|---|---|---|---|---|---|
| chr4:143543509-143543973 | ENSG00000153147 | SMARCA5 | 15-16 | 177 | 0.296 |
| chr11:33286413-33287512 | ENSG00000110422 | HIPK3 | 2 | 177 | 0.541 |
| chr7:100023419-100024308 | ENSG00000106261 | ZKSCAN1 | 2-3 | 177 | 0.539 |
| chr4:152411303-152412530 | ENSG00000109670 | FBXW7 | 2 | 177 | 0.437 |
| Name | Ensemble Gene ID | Circular Region | Fisher p-Value | FDR * | Comparison |
|---|---|---|---|---|---|
| circSATB2 | ENSG00000119042 | chr2:199368605-199433515 | 2.47 × 10−8 | 6.84 × 10−6 | MSI |
| 2.41 × 10−8 | 6.68 × 10−6 | CMS | |||
| circNAB1 | ENSG00000138386 | chr2:190659158-190673153 | 6.48 × 10−7 | 0.000179 | MSI |
| 0.00007584 | 0.02078 | CMS | |||
| circZBTB44 | ENSG00000196323 | chr11:130260856-130261930 | 3.4 × 10−5 | 0.009347 | MSI |
| circCEP192 | ENSG00000101639 | chr18:12999421-13019207 | 0.000152 | 0.041758 | MSI |
| 0.00007808 | 0.021316 | CMS | |||
| chr18:12999421-13030609 | 0.000233 | 0.063043 | CMS | ||
| circUBAP2 | ENSG00000137073 | chr9:33960826-33973238 | 0.000238 | 0.064846 | MSI |
| circMGA | ENSG00000174197 | chr15:41668828-41669959 | 0.000235 | 0.064979 | DFSI |
| circASPH | ENSG00000198363 | chr8:61618978-61653661 | 1.19 × 10−7 | 3.28 × 10−5 | CMS |
| circCCSER2 | ENSG00000107771 | chr10:84438512-84477665 | 3.9 × 10−5 | 0.010728 | CMS |
| circZNF609 | ENSG00000180357 | chr15:64499293-64500167 | 0.00017 | 0.046128 | CMS |
| circFUT8 | ENSG00000033170 | chr14:65561337-65561767 | 0.00026 | 0.070135 | CMS |
| circMRPS35 | ENSG00000061794 | chr12:27714780-27724187 | 0.000299 | 0.08035 | CMS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van den Berg, I.; Smid, M.; Coebergh van den Braak, R.R.J.; van Deurzen, C.H.M.; de Weerd, V.; Foekens, J.A.; IJzermans, J.N.M.; Martens, J.W.M.; Wilting, S.M. Circular RNA in Chemonaive Lymph Node Negative Colon Cancer Patients. Cancers 2021, 13, 1903. https://doi.org/10.3390/cancers13081903
van den Berg I, Smid M, Coebergh van den Braak RRJ, van Deurzen CHM, de Weerd V, Foekens JA, IJzermans JNM, Martens JWM, Wilting SM. Circular RNA in Chemonaive Lymph Node Negative Colon Cancer Patients. Cancers. 2021; 13(8):1903. https://doi.org/10.3390/cancers13081903
Chicago/Turabian Stylevan den Berg, Inge, Marcel Smid, Robert R. J. Coebergh van den Braak, Carolien H. M. van Deurzen, Vanja de Weerd, John A. Foekens, Jan N. M. IJzermans, John W. M. Martens, and Saskia M. Wilting. 2021. "Circular RNA in Chemonaive Lymph Node Negative Colon Cancer Patients" Cancers 13, no. 8: 1903. https://doi.org/10.3390/cancers13081903
APA Stylevan den Berg, I., Smid, M., Coebergh van den Braak, R. R. J., van Deurzen, C. H. M., de Weerd, V., Foekens, J. A., IJzermans, J. N. M., Martens, J. W. M., & Wilting, S. M. (2021). Circular RNA in Chemonaive Lymph Node Negative Colon Cancer Patients. Cancers, 13(8), 1903. https://doi.org/10.3390/cancers13081903