
Blood-Derived Extracellular Vesicle-Associated miR-3182 Detects Non-Small Cell Lung Cancer Patients

 , , , , , ,
, , , , , ,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Sample Collection
2.2. Cell Culture
2.3. SEV Isolation
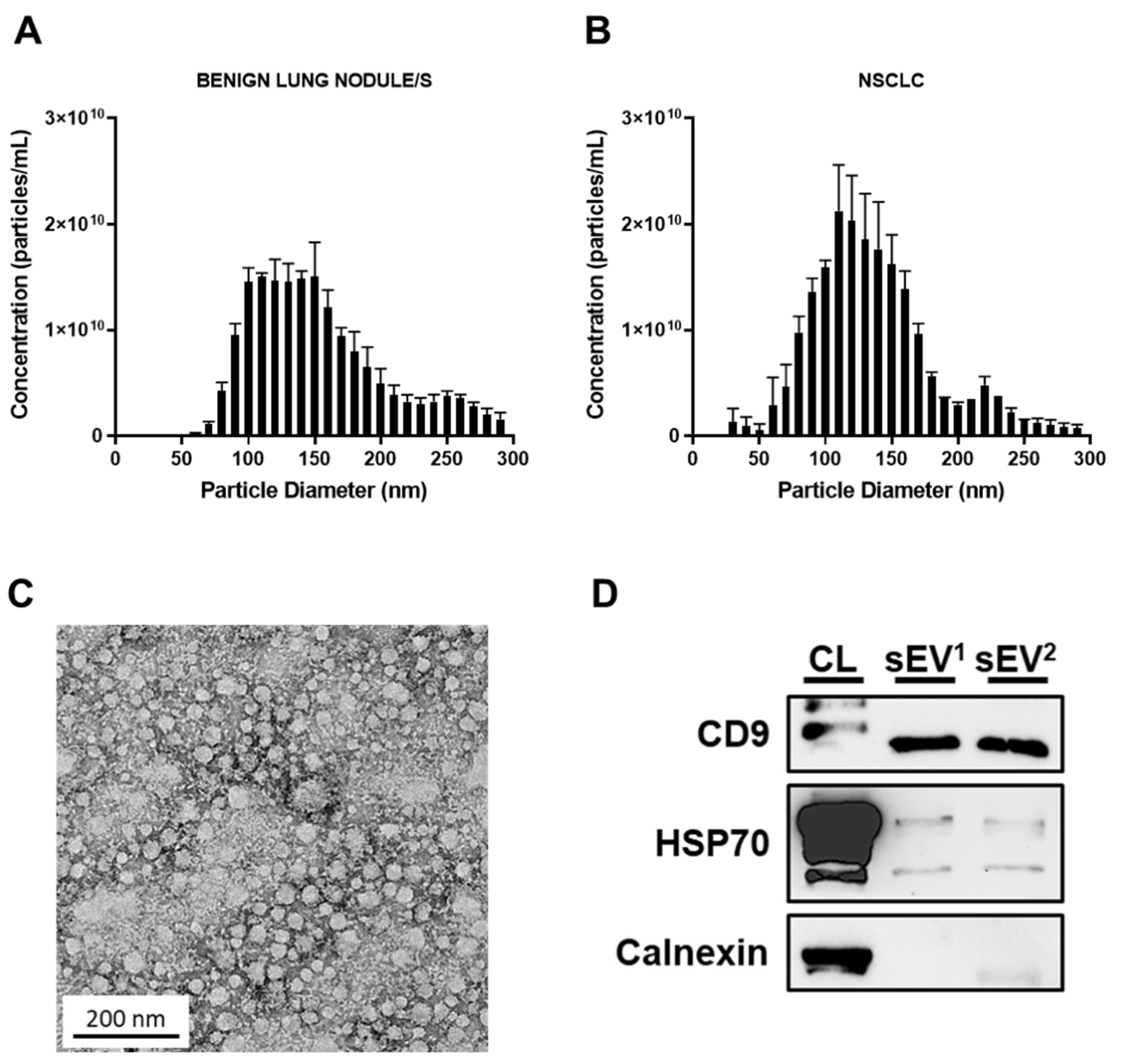
2.4. Western Blot
2.5. Nanoparticle Tracking Analysis
2.6. Transmission Electron Microscopy
2.7. Small RNA Sequencing and Analysis
2.8. SEV miRNA Detection by Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR)
2.9. Statistical Analysis
3. Results
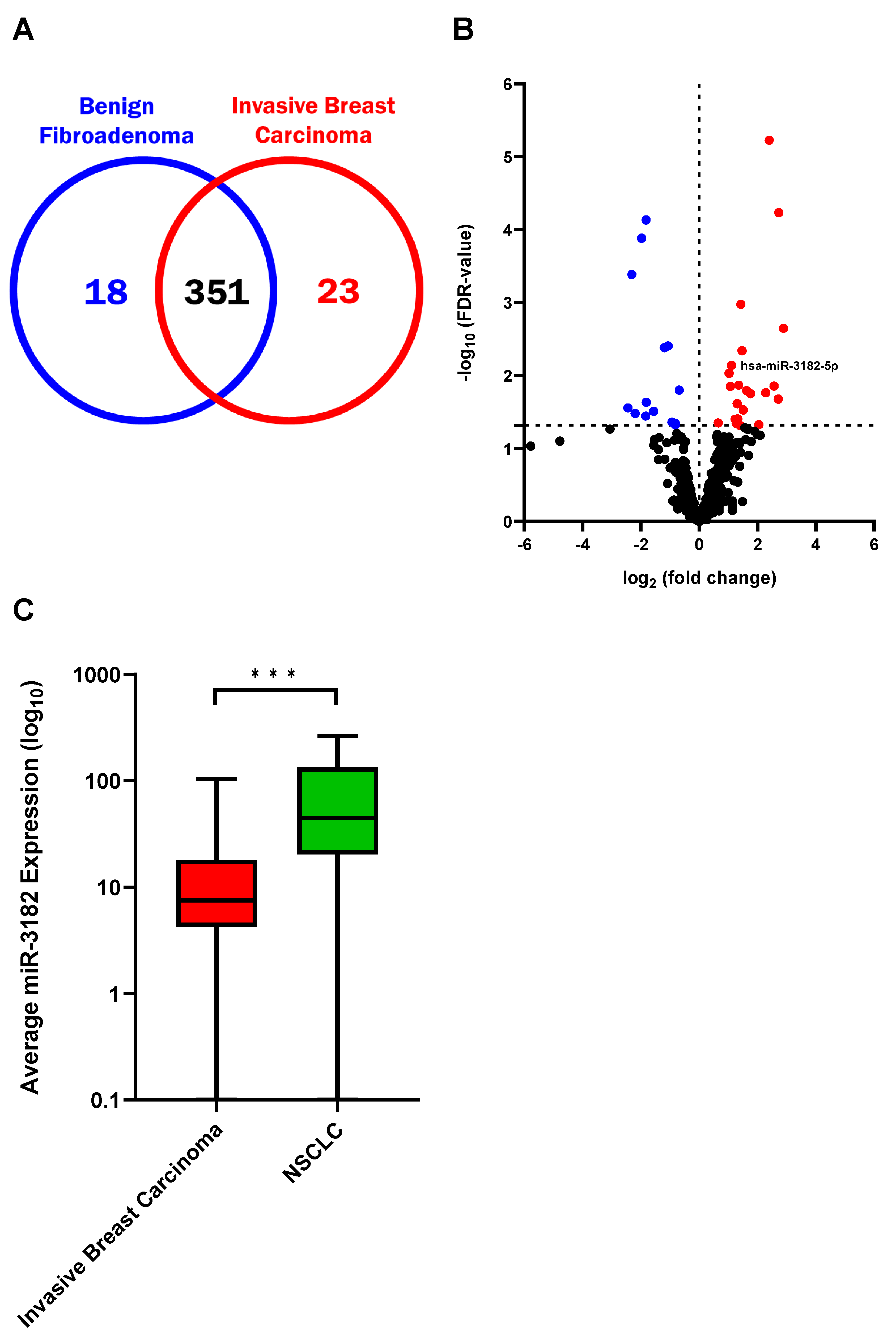
3.1. Circulating sEV miRNAs Differentiate Invasive Breast Carcinoma from Benign Fibroadenoma, and Identify NSCLC and Oncogenic Transformation of Lung Cells
3.2. MiR-3182 Distinguishes Early-Stage NSCLC Patients from Subjects with Benign Pathology in Individuals with Lung Nodules Presenting for Investigation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics. Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Birring, S.S.; Peake, M.D. Symptoms and the early diagnosis of lung cancer. Thorax 2005, 60, 268–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobb, R.J.; Hastie, M.L.; Norris, E.L.; van Amerongen, R.; Gorman, J.J.; Möller, A. Oncogenic transformation of lung cells results in distinct exosome protein profile similar to the cell of origin. Proteomics 2017, 17, 23–24. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Zhang, C.; Du, T.; Gabriel, A.N.A.; Wang, X.; Li, X.; Sun, L.; Wang, N.; Jiang, X.; Zhang, Y. Progress of exosomes in the diagnosis and treatment of lung cancer. Biomed. Pharmacother. 2021, 134, 111111. [Google Scholar] [CrossRef]
- Testa, A.; Venturelli, E.; Brizzi, M. Extracellular Vesicles as a Novel Liquid Biopsy-Based Diagnosis for the Central Nervous System, Head and Neck, Lung, and Gastrointestinal Cancers: Current and Future Perspectives. Cancers 2021, 13, 2792. [Google Scholar] [CrossRef]
- Health AIo, Welfare. Cancer Data in Australia; AIHW: Canberra, Australia, 2021. [Google Scholar]
- Socinski, M.A.; Evans, T.; Gettinger, S.; Hensing, T.A.; VanDam Sequist, L.; Ireland, B.; Stinchcombe, T.E. Treatment of stage IV non-small cell lung cancer: Diagnosis and management of lung cancer, 3rd ed: American College of Chest Physicians evidence-based clinical practice guidelines. Chest 2013, 143, 341S–368S. [Google Scholar] [CrossRef] [Green Version]
- Gajra, A.; Zemla, T.J.; Jatoi, A.; Feliciano, J.L.; Wong, M.L.; Chen, H.; Maggiore, R.; McMurray, R.P.; Hurria, A.; Muss, H.B.; et al. Time-to-Treatment-Failure and Related Outcomes Among 1000+ Advanced Non–Small Cell Lung Cancer Patients: Comparisons Between Older Versus Younger Patients (Alliance A151711). J. Thorac. Oncol. 2018, 13, 996–1003. [Google Scholar] [CrossRef] [Green Version]
- Frangioni, J.V. New Technologies for Human Cancer Imaging. J. Clin. Oncol. 2008, 26, 4012–4021. [Google Scholar] [CrossRef] [PubMed]
- Moller, A.; Visan, K.S.; Lobb, R.J. The role of exosomes in the promotion of epithelial-to-mesenchymal transition and metastasis. Front. Biosci. 2020, 25, 1022–1057. [Google Scholar] [CrossRef] [PubMed]
- Shyamala, K.; Girish, H.C.; Murgod, S. Risk of tumor cell seeding through biopsy and aspiration cytology. J. Int. Soc. Prev. Community Dent. 2014, 4, 5–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guix, F.X. The Potential Use of Exosomes as a Diagnostic and Prognostic Tool. J. Biomed. Sci. 2017, 6, 24. [Google Scholar] [CrossRef] [Green Version]
- Möller, A.; Lobb, R.J. The evolving translational potential of small extracellular vesicles in cancer. Nat. Rev. Cancer 2020, 20, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Lima, L.G.; Ham, S.; Shin, H.; Chai, E.P.Z.; Lek, E.S.H.; Lobb, R.J.; Müller, A.F.; Mathivanan, S.; Yeo, B.; Choi, Y.; et al. Tumor microenvironmental cytokines bound to cancer exosomes determine uptake by cytokine receptor-expressing cells and biodistribution. Nat. Commun. 2021, 12, 3543. [Google Scholar] [CrossRef] [PubMed]
- Soung, Y.H.; Ford, S.; Zhang, V.; Chung, J. Exosomes in Cancer Diagnostics. Cancers 2017, 9, 8. [Google Scholar] [CrossRef] [Green Version]
- Lobb, R.; Lima, L.G.; Möller, A. Exosomes: Key mediators of metastasis and pre-metastatic niche formation. Semin. Cell Dev. Biol. 2017, 67, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Słomka, A.; Mocan, T.; Wang, B.; Nenu, I.; Urban, S.K.; Gonzalez-Carmona, M.A.; Schmidt-Wolf, I.G.H.; Lukacs-Kornek, V.; Strassburg, C.P.; Spârchez, Z.; et al. EVs as Potential New Therapeutic Tool/Target in Gastrointestinal Cancer and HCC. Cancers 2020, 12, 3019. [Google Scholar] [CrossRef]
- Sohel, M.H. Extracellular/Circulating MicroRNAs: Release Mechanisms, Functions and Challenges. Achiev. Life Sci. 2016, 10, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Schwarzenbach, H. Methods for quantification and characterization of microRNAs in cell-free plasma/serum, normal exosomes and tumor-derived exosomes. Transl. Cancer Res. 2018, 7, S253–S263. [Google Scholar] [CrossRef]
- Zhao, C.; Sun, X.; Li, L. Biogenesis and function of extracellular miRNAs. ExRNA 2019, 1, 38. [Google Scholar] [CrossRef] [Green Version]
- Mirzaei, S.; Zarrabi, A.; Hashemi, F.; Zabolian, A.; Saleki, H.; Ranjbar, A.; Saleh, S.H.S.; Bagherian, M.; Sharifzadeh, S.O.; Hushmandi, K.; et al. Regulation of Nuclear Factor-KappaB (NF-κB) sig-naling pathway by non-coding RNAs in cancer: Inhibiting or promoting carcinogenesis? Cancer Letters 2021, 509, 63–80. [Google Scholar] [CrossRef]
- Jung-Hee, K. Exosomes as diagnostic biomarkers in cancer. Mol. Cell. Toxicology. 2018, 14, 113–122. [Google Scholar]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghamloush, F.; Ghayad, S.E.; Rammal, G.; Fahs, A.; Ayoub, A.J.; Merabi, Z.; Harajly, M.; Zalzali, H.; Saab, R. The PAX3-FOXO1 oncogene alters exosome miRNA content and leads to paracrine effects mediated by exosomal miR-486. Sci. Rep. 2019, 9, 14242. [Google Scholar] [CrossRef]
- Abels, E.R.; Maas, S.L.; Nieland, L.; Wei, Z.; Cheah, P.S.; Tai, E.; Kolsteeg, C.-J.; Dusoswa, S.A.; Ting, D.T.; Hickman, S.; et al. Glioblastoma-Associated Microglia Reprogramming Is Mediated by Functional Transfer of Extracellular miR-21. Cell Rep. 2019, 28, 3105–3119. [Google Scholar] [CrossRef] [Green Version]
- Lucero, R.; Zappulli, V.; Sammarco, A.; Murillo, O.D.; Cheah, P.S.; Srinivasan, S.; Tai, E.; Ting, D.T.; Wei, Z.; Roth, M.E.; et al. Glioma-Derived miRNA-Containing Extra-cellular Vesicles Induce Angiogenesis by Reprogramming Brain Endothelial Cells. Cell Rep. 2020, 30, 2065–2074. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Peng, F.; Chen, J. The Role of Exosomal MicroRNAs in the Tumor Microenvironment of Breast Cancer. Int. J. Mol. Sci. 2019, 20, 3884. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Fong, M.Y.; Min, Y.; Somlo, G.; Liu, L.; Palomares, M.R.; Yu, Y.; Chow, A.; O’Connor, S.T.F.; Chin, A.R.; et al. Cancer-secreted miR-105 destroys vascular endothelial barriers to promote metastasis. Cancer Cell 2014, 25, 501–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dilsiz, N. Role of exosomes and exosomal microRNAs in cancer. Futur. Sci. 2020, 6, FSO465. [Google Scholar] [CrossRef] [Green Version]
- Lobb, R.; Van Amerongen, R.; Wiegmans, A.; Ham, S.; Larsen, J.E.; Möller, A. Exosomes derived from mesenchymal non-small cell lung cancer cells promote chemoresistance. Int. J. Cancer 2017, 141, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Lobb, R.j.; Becker, M.; Wen, S.W.; Wong, C.S.F.; Wiegmans, A.P.; Leimgruber, A.; Möller, A. Optimized exosome isolation protocol for cell culture supernatant and human plasma. J. Extracell. Vesicles 2015, 4, 27031. [Google Scholar] [CrossRef]
- Möller, A.; House, C.M.; Wong, C.S.F.; Scanlon, D.B.; Liu, M.C.P.; Ronai, Z.; Bowtell, D.D.L. Inhibition of Siah ubiquitin ligase function. Oncogene 2009, 28, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Salomon, C.; Guanzon, D.; Scholz-Romero, K.; Longo, S.; Correa, P.; Illanes, S.E.; Rice, G.E. Placental Exosomes as Early Biomarker of Preeclampsia: Potential Role of Exosomal MicroRNAs Across Gestation. J. Clin. Endocrinol. Metab. 2017, 102, 3182–3194. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Ortiz, A. Not all extracellular vesicles were created equal: Clinical implications. Ann. Transl. Med. 2017, 5, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Diergaarde, B.; Ferrone, S.; Kirkwood, J.M.; Whiteside, T.L. Melanoma cell-derived exosomes in plasma of melanoma patients suppress functions of immune effector cells. Sci. Rep. 2020, 10, 92. [Google Scholar] [CrossRef] [PubMed]
- Boukouris, S.; Mathivanan, S. Exosomes in bodily fluids are a highly stable resource of disease biomarkers. Proteom.–Clin. Appl. 2015, 9, 358–367. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Larsen, J.; Lee, W.; Sun, H.; Shames, D.S.; Dalvi, M.P.; Ramirez, R.D.; Tang, H.; DiMaio, J.M.; Gao, B.; et al. Human Lung Epithelial Cells Progressed to Malignancy through Specific Oncogenic Manipulations. Mol. Cancer Res. 2013, 11, 638–650. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Hu, C.; Liu, P.; Lu, M. Tumor-derived exosomes in cancer metastasis risk diagnosis and metastasis therapy. Clin. Transl. Oncol. 2018, 21, 152–159. [Google Scholar] [CrossRef]
- Huang, W.; Yan, Y.; Liu, Y.; Lin, M.; Ma, J.; Zhang, W.; Dai, W.; Li, J.; Guo, Q.; Chen, H.; et al. Exosomes with low miR-34c-3p expression promote invasion and mi-gration of non-small cell lung cancer by upregulating integrin α2β. Signal Transduct. Target. Ther. 2020, 5, 39. [Google Scholar] [CrossRef]
- Rabinowits, G.; Bowden, M.; Flores, L.M.; Verselis, S.; Vergara, V.; Jo, V.Y.; Chau, N.; Lorch, J.; Hammerman, P.S.; Thomas, T.; et al. Comparative Analysis of MicroRNA Expression among Benign and Malignant Tongue Tissue and Plasma of Patients with Tongue Cancer. Front. Oncol. 2017, 7, 191. [Google Scholar] [CrossRef] [Green Version]
- de Mooij, T.; Peterson, T.E.; Evans, J.; McCutcheon, B.; Parney, I.F. Short non-coding RNA sequencing of glioblastoma extracellular vesicles. J. Neuro-Oncol. 2020, 146, 253–263. [Google Scholar] [CrossRef]
- Wang, X.; Wilkinson, R.; Kildey, K.; Potriquet, J.; Mulvenna, J.; Lobb, R.; Möller, A.; Cloonan, N.; Mukhopadhyay, P.; Kassianos, A.J.; et al. Unique molecular profile of exosomes derived from primary human proximal tubular epithelial cells under diseased conditions. J. Extracell. Vesicles 2017, 6, 1314073. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Sceneay, J.; Godde, N.; Kinwel, T.; Ham, S.; Thompson, E.W.; Humbert, O.P.; Möller, A. Intermittent hypoxia induces a metastatic phenotype in breast cancer. Oncogene 2018, 37, 4214–4225. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.-L.; Hung, J.-Y.; Chang, W.-A.; Lin, Y.-S.; Pan, Y.-C.; Tsai, P.-H.; Wu, C.-Y.; Kuo, P.-L. Hypoxic lung cancer-secreted exosomal miR-23a increased angiogenesis and vascular permeability by targeting prolyl hydroxylase and tight junction protein ZO-1. Oncogene 2017, 36, 4929–4942. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Min, Z.; Zhou, Z.; Linhong, M.; Tao, R.; Yan, L.; Song, H. Hypoxia-induced exosomes promote hepatocellular carcinoma prolif-eration and metastasis via miR-1273f transfer. Exp. Cell Res. 2019, 385, 111649. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, C.; Wang, S.; Wang, Z.; Jiang, J.; Wang, W.; Li, X.; Chen, J.; Liu, K.; Li, C.; et al. Exosomes Derived from Hypoxic Oral Squamous Cell Carcinoma Cells Deliver miR-21 to Normoxic Cells to Elicit a Prometastatic Phenotype. Cancer Res. 2016, 76, 1770–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, G.; Huang, J.; Chen, W.; You, P.; Ding, Y.; Tu, P. circUBAP2 exacerbates malignant capabilities of NSCLC by targeting KLF4 through miR-3182 modulation. Aging 2021, 13, 11083–11095. [Google Scholar] [CrossRef]
- Xue, M.; Shi, D.; Xu, G.; Wang, W. The long noncoding RNA linc00858 promotes progress of lung cancer through miR-3182/MMP2 axis. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2091–2097. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zhu, Y.; Zuo, G.; Chen, X.; Cheng, J.; Zhang, S. LINC00858 promotes retinoblastoma cell proliferation, migration and invasion by inhibiting miR-3182. Exp. Ther. Med. 2019, 19, 999–1005. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.-P.; Ma, X.-L.; Zhang, C.-L. LncRNA ODRUL Contributes to Osteosarcoma Progression through the miR-3182/MMP2 Axis. Mol. Ther. J. Am. Soc. Gene Therapy 2017, 25, 2383–2393. [Google Scholar] [CrossRef] [Green Version]
- Tian, F.; Shen, Y.; Chen, Z.; Li, R.; Ge, Q. No Significant Difference between Plasma miRNAs and Plasma-Derived Exosomal miRNAs from Healthy People. BioMed Res. Int. 2017, 2017, 1304816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, D.J.; Franklin, J.L.; Dou, Y.; Liu, Q.; Higginbotham, J.N.; Demory Beckler, M.; Weaver, A.M.; Vickers, K.; Prasad, N.; Levy, S.; et al. KRAS-dependent sorting of miRNA to exo-somes. eLife 2015, 4, e07197. [Google Scholar] [CrossRef] [PubMed]
- Ju, L.; Han, M.; Li, X.; Zhao, C. MicroRNA Signature of Lung Adenocarcinoma with EGFR Exon 19 Deletion. J. Cancer 2017, 8, 1311–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiong, K.-L.; Yeang, C.-H. Explaining cancer type specific mutations with transcriptomic and epigenomic features in normal tissues. Sci. Rep. 2018, 8, 11456. [Google Scholar] [CrossRef] [Green Version]
- Momcilovic, M.; Shackelford, D.B. Targeting LKB1 in cancer—Exposing and exploiting vulnerabilities. Br. J. Cancer 2015, 113, 574–584. [Google Scholar] [CrossRef] [Green Version]
- Mo, X.; Yin, W.; Huang, Y.; Guo, W.; Zhou, M.; Ye, H. Expression of miR-3182 and EBV-miR-BART8-3p in nasopharyngeal carcinoma is correlated with distant metastasis. Int. J. Clin. Exp. Pathol. 2018, 11, 3134–3140. [Google Scholar]
- Husby, S.; Ralfkiaer, U.; Garde, C.; Zandi, R.; Ek, S.; Kolstad, A.; Jerkeman, M.; Laurell, A.; Räty, R.; Pedersen, L.B.; et al. miR-18b overexpression identifies mantle cell lymphoma patients with poor outcome and improves the MIPI-B prognosticator. Blood 2015, 125, 2669–2677. [Google Scholar] [CrossRef] [Green Version]
- Bach, D.; Fuereder, J.; Karbiener, M.; Scheideler, M.; Ress, A.L.; Neureiter, D.; Kemmerling, R.; Dietze, O.; Wiederstein, M.; Berr, F.; et al. Comprehensive analysis of alterations in the miRNome in response to photodynamic treatment. J. Photochem. Photobiol. B Biol. 2013, 120, 74–81. [Google Scholar] [CrossRef]
- Pehserl, A.-M.; Ress, A.L.; Stanzer, S.; Resel, M.; Karbiener, M.; Stadelmeyer, E.; Stiegelbauer, V.; Gerger, A.; Mayr, C.; Scheideler, M.; et al. Comprehensive Analysis of miRNome Alterations in Response to Sorafenib Treatment in Colorectal Cancer Cells. Int. J. Mol. Sci. 2016, 17, 2011. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.F.; Liu, Z.; Chaurasiya, S.; Dellinger, T.; Lu, J.; Wu, X.; Qin, H.; Wang, J.; Fong, Y.; Yuan, Y.-C. Identification of core aberrantly expressed microRNAs in serous ovarian carcinoma. Oncotarget 2018, 9, 20451–20466. [Google Scholar] [CrossRef] [Green Version]
- Wei, P.; Xie, Y.; Abel, P.W.; Huang, Y.; Ma, Q.; Li, L.; Hao, J.; Wolff, D.W.; Wie, T.; Tu, Y. Transforming growth factor (TGF)-β1-induced miR-133a inhibits myofi-broblast differentiation and pulmonary fibrosis. Cell Death Disease 2019, 10, 670. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Lee, Y.J.; Park, J.S.; Cho, Y.-J.; Yoon, H.I.; Lee, J.-H.; Lee, C.-T.; Chung, J.-H.; Lee, K.W.; Lee, S.H. Prognosis of non-small-cell lung cancer in patients with idiopathic pulmonary fibrosis. Sci. Rep. 2019, 9, 12561. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Visan, K.S.; Lobb, R.J.; Wen, S.W.; Bedo, J.; Lima, L.G.; Krumeich, S.; Palma, C.; Ferguson, K.; Green, B.; Niland, C.; et al. Blood-Derived Extracellular Vesicle-Associated miR-3182 Detects Non-Small Cell Lung Cancer Patients. Cancers 2022, 14, 257. https://doi.org/10.3390/cancers14010257
Visan KS, Lobb RJ, Wen SW, Bedo J, Lima LG, Krumeich S, Palma C, Ferguson K, Green B, Niland C, et al. Blood-Derived Extracellular Vesicle-Associated miR-3182 Detects Non-Small Cell Lung Cancer Patients. Cancers. 2022; 14(1):257. https://doi.org/10.3390/cancers14010257
Chicago/Turabian StyleVisan, Kekoolani S., Richard J. Lobb, Shu Wen Wen, Justin Bedo, Luize G. Lima, Sophie Krumeich, Carlos Palma, Kaltin Ferguson, Ben Green, Colleen Niland, and et al. 2022. "Blood-Derived Extracellular Vesicle-Associated miR-3182 Detects Non-Small Cell Lung Cancer Patients" Cancers 14, no. 1: 257. https://doi.org/10.3390/cancers14010257
APA StyleVisan, K. S., Lobb, R. J., Wen, S. W., Bedo, J., Lima, L. G., Krumeich, S., Palma, C., Ferguson, K., Green, B., Niland, C., Cloonan, N., Simpson, P. T., McCart Reed, A. E., Everitt, S. J., MacManus, M. P., Hartel, G., Salomon, C., Lakhani, S. R., Fielding, D., & Möller, A. (2022). Blood-Derived Extracellular Vesicle-Associated miR-3182 Detects Non-Small Cell Lung Cancer Patients. Cancers, 14(1), 257. https://doi.org/10.3390/cancers14010257