
Cellular Origins and Pathogenesis of Gastrointestinal NK- and T-Cell Lymphoproliferative Disorders


Abstract
:Simple Summary
Abstract
1. Introduction
2. Immune Cells in the Gastrointestinal Mucosa
2.1. Innate Lymphoid Cells (ILCs)
2.1.1. Lamina Propria ILCs
2.1.2. Intraepithelial Innate Lymphoid Cells (ILCs)
- (a)
- (b)
- (c)
- Intraepithelial ILCs that are Id2-independent and express intracytoplasmic CD3 as innate CD3+ IEL (iCD3+ IEL) [25];
- (d)
2.2. Natural Killer (NK) Cells
2.2.1. CD56dim versus CD56bright NK-Cells
2.2.2. CD4 and CD8 Expression in NK-Cells
2.2.3. Tissue-Resident NK Cells
2.2.4. NK Cells in the GI Tract
3. T-Cell Populations in the Gastrointestinal Tract
3.1. Conventional T-Lymphocytes
3.2. Unconventional T-Lymphocytes
4. Intraepithelial Lymphocytes (IELs)
4.1. Type ‘a’ (Induced TCR + IELs)
4.2. Type ‘b’ (Natural TCR + IEL)
4.3. Innate Lymphoid Cells (ILCs)
4.4. CD8αα Expression in IELs
5. Lymphomas and Lymphoproliferative Disorders Derived from NK-Cells
5.1. Lymphomatoid Gastropathy (LG)/NK-Cell Enteropathy (NKCE)
5.1.1. Incidence and Prevalence
5.1.2. Clinical Features
5.1.3. Pathological Features
5.1.4. Immunophenotype
5.1.5. Cellular Origin from CD56bright Subset of NK-cells
5.1.6. Pathogenetic Mechanisms
5.2. Extranodal NK/T Cell Lymphoma, Nasal Type (ENKTL)
5.2.1. Incidence and Prevalence
5.2.2. Clinical Features
5.2.3. Pathological Findings
5.2.4. Immunophenotype
5.2.5. Cellular Origin from Activated CD56bright NK-Cells
5.2.6. Cellular Origin from CD56+ T-Cells
5.2.7. Pathogenetic Mechanisms
6. Lymphomas and Lymphoproliferative Disorders Derived from T-Cells
6.1. Indolent T-Cell Lymphoproliferative Disorder of the GI Tract
6.1.1. Incidence and Prevalence
6.1.2. Clinical Features
6.1.3. Pathological Findings
6.1.4. Immunophenotype
6.1.5. Cellular Origin of Indolent T-Cell LPD of GI Tract
CD4+ Indolent T-cell LPD
CD8+ Indolent T-cell LPD
6.1.6. Pathogenetic Mechanisms
6.2. Celiac Disease
6.2.1. Epidemiology
6.2.2. Clinical Features
6.2.3. Pathological Features
6.3. Refractory Celiac Disease (RCD)
6.4. Enteropathy Associated T-Cell Lymphoma
6.4.1. Epidemiology
6.4.2. Clinical Findings
6.4.3. Pathological Findings
6.4.4. Immunophenotype
6.4.5. Cellular Origins
Cellular Origin of EATLs Arising from RCD2
Cellular Origins of EATL Arising from RCD1 and CD
6.4.6. Pathogenetic Mechanisms in Celiac Disease, RCD and EATL
6.5. Monomorphic Epitheliotropic Intestinal T-Cell Lymphoma (MEITL)
6.5.1. Incidence and Prevalence
6.5.2. Clinical Features
6.5.3. Pathological Findings
6.5.4. Immunophenotype
6.5.5. Cellular Origin from Non-Conventional CD8αα+ T-Cells
6.5.6. Pathogenetic Mechanisms
6.6. Intestinal T-Cell Lymphoma, NOS
6.6.1. Incidence and Prevalence
6.6.2. Clinical Features
6.6.3. Pathological Findings
6.6.4. Cellular Origins from Conventional and Non-Conventional T-Cells
6.6.5. Pathogenetic Mechanisms
7. Treatment and Clinical Trials
7.1. Current and Novel Treatment in ENKTL
7.2. Current and Novel Treatment in EATL, MEITL and ITCL, NOS
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kohno, S.; Ohshima, K.; Yoneda, S.; Kodama, T.; Shirakusa, T.; Kikuchi, M. Clinicopathological analysis of 143 primary malignant lymphomas in the small and large intestines based on the new WHO classification. Histopathology 2003, 43, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Matsumoto, T.; Iida, M.; Yao, T.; Tsuneyoshi, M. Primary gastrointestinal lymphoma in Japan: A clinicopathologic analysis of 455 patients with special reference to its time trends. Cancer 2003, 97, 2462–2473. [Google Scholar] [CrossRef] [PubMed]
- Spits, H.; Artis, D.; Colonna, M.; Diefenbach, A.; Di Santo, J.P.; Eberl, G.; Koyasu, S.; Locksley, R.M.; McKenzie, A.N.; Mebius, R.E.; et al. Innate lymphoid cells—A proposal for uniform nomenclature. Nat. Rev. Immunol. 2013, 13, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Spits, H.; Bernink, J.H.; Lanier, L. NK-cells and type 1 innate lymphoid cells: Partners in host defense. Nat. Immunol. 2016, 17, 758–764. [Google Scholar] [CrossRef]
- Spits, H.; Di Santo, J.P. The expanding family of innate lymphoid cells: Regulators and effectors of immunity and tissue remodeling. Nat. Immunol. 2011, 12, 21–27. [Google Scholar] [CrossRef]
- Sun, J.C.; Lanier, L.L. NK-cell development, homeostasis and function: Parallels with CD8(+) T cells. Nat. Rev. Immunol. 2011, 11, 645–657. [Google Scholar] [CrossRef]
- Chen, L.; Youssef, Y.; Robinson, C.; Ernst, G.F.; Carson, M.Y.; Young, K.A.; Scoville, S.D.; Zhang, X.; Harris, R.; Sekhri, P.; et al. CD56 Expression Marks Human Group 2 Innate Lymphoid Cell Divergence from a Shared NK-cell and Group 3 Innate Lymphoid Cell Developmental Pathway. Immunity 2018, 49, 464–476.e4. [Google Scholar] [CrossRef] [Green Version]
- Klose, C.S.; Blatz, K.; d’Hargues, Y.; Hernandez, P.P.; Kofoed-Nielsen, M.; Ripka, J.F.; Ebert, K.; Arnold, S.J.; Diefenbach, A.; Palmer, E.; et al. The transcription factor T-bet is induced by IL-15 and thymic agonist selection and controls CD8αα(+) intraepithelial lymphocyte development. Immunity 2014, 41, 230–243. [Google Scholar] [CrossRef] [Green Version]
- Vivier, E.; Artis, D.; Colonna, M.; Diefenbach, A.; Di Santo, J.P.; Eberl, G.; Koyasu, S.; Locksley, R.M.; McKenzie, A.N.J.; Mebius, R.E.; et al. Innate Lymphoid Cells: 10 Years On. Cell 2018, 174, 1054–1066. [Google Scholar] [CrossRef] [Green Version]
- Bernink, J.H.; Krabbendam, L.; Germar, K.; de Jong, E.; Gronke, K.; Kofoed-Nielsen, M.; Munneke, J.M.; Hazenberg, M.D.; Villaudy, J.; Buskens, C.J.; et al. Interleukin-12 and -23 Control Plasticity of CD127(+) Group 1 and Group 3 Innate Lymphoid Cells in the Intestinal Lamina Propria. Immunity 2015, 43, 146–160. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, A.; Vermi, W.; Lee, J.S.; Lonardi, S.; Gilfillan, S.; Newberry, R.D.; Cella, M.; Colonna, M. Intraepithelial type 1 innate lymphoid cells are a unique subset of IL-12- and IL-15-responsive IFN-gamma-producing cells. Immunity 2013, 38, 769–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, S.H.; Walker, J.A.; Jolin, H.E.; Drynan, L.F.; Hams, E.; Camelo, A.; Barlow, J.L.; Neill, D.R.; Panova, V.; Koch, U.; et al. Transcription factor RORα is critical for nuocyte development. Nat. Immunol. 2012, 13, 229–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mjosberg, J.M.; Trifari, S.; Crellin, N.K.; Peters, C.P.; van Drunen, C.M.; Piet, B.; Fokkens, W.J.; Cupedo, T.; Spits, H. Human IL-25- and IL-33-responsive type 2 innate lymphoid cells are defined by expression of CRTH2 and CD161. Nat. Immunol. 2011, 12, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Moro, K.; Kabata, H.; Tanabe, M.; Koga, S.; Takeno, N.; Mochizuki, M.; Fukunaga, K.; Asano, K.; Betsuyaku, T.; Koyasu, S. Interferon and IL-27 antagonize the function of group 2 innate lymphoid cells and type 2 innate immune responses. Nat. Immunol. 2016, 17, 76–86. [Google Scholar] [CrossRef]
- Neill, D.R.; Wong, S.H.; Bellosi, A.; Flynn, R.J.; Daly, M.; Langford, T.K.; Bucks, C.; Kane, C.M.; Fallon, P.G.; Pannell, R.; et al. Nuocytes represent a new innate effector leukocyte that mediates type-2 immunity. Nature 2010, 464, 1367–1370. [Google Scholar] [CrossRef] [Green Version]
- Cella, M.; Fuchs, A.; Vermi, W.; Facchetti, F.; Otero, K.; Lennerz, J.K.; Doherty, J.M.; Mills, J.C.; Colonna, M. A human natural killer cell subset provides an innate source of IL-22 for mucosal immunity. Nature 2009, 457, 722–725. [Google Scholar] [CrossRef]
- Cupedo, T.; Crellin, N.K.; Papazian, N.; Rombouts, E.J.; Weijer, K.; Grogan, J.L.; Fibbe, W.E.; Cornelissen, J.J.; Spits, H. Human fetal lymphoid tissue-inducer cells are interleukin 17-producing precursors to RORC+ CD127+ natural killer-like cells. Nat. Immunol. 2009, 10, 66–74. [Google Scholar] [CrossRef]
- Hughes, T.; Becknell, B.; McClory, S.; Briercheck, E.; Freud, A.G.; Zhang, X.; Mao, H.; Nuovo, G.; Yu, J.; Caligiuri, M.A. Stage 3 immature human natural killer cells found in secondary lymphoid tissue constitutively and selectively express the TH 17 cytokine interleukin-22. Blood 2009, 113, 4008–4010. [Google Scholar] [CrossRef] [Green Version]
- van de Pavert, S.A. Lymphoid Tissue inducer (LTi) cell ontogeny and functioning in embryo and adult. Biomed. J. 2021, 44, 123–132. [Google Scholar] [CrossRef]
- Wang, S.; Xia, P.; Chen, Y.; Qu, Y.; Xiong, Z.; Ye, B.; Du, Y.; Tian, Y.; Yin, Z.; Xu, Z.; et al. Regulatory Innate Lymphoid Cells Control Innate Intestinal Inflammation. Cell 2017, 171, 201–216.e18. [Google Scholar] [CrossRef] [Green Version]
- Hoorweg, K.; Peters, C.P.; Cornelissen, F.; Aparicio-Domingo, P.; Papazian, N.; Kazemier, G.; Mjosberg, J.M.; Spits, H.; Cupedo, T. Functional Differences between Human NKp44(-) and NKp44(+) RORC(+) Innate Lymphoid Cells. Front. Immunol. 2012, 3, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simoni, Y.; Fehlings, M.; Kløverpris, H.N.; McGovern, N.; Koo, S.L.; Loh, C.Y.; Lim, S.; Kurioka, A.; Fergusson, J.R.; Tang, C.L.; et al. Human Innate Lymphoid Cell Subsets Possess Tissue-Type Based Heterogeneity in Phenotype and Frequency. Immunity 2017, 46, 148–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talayero, P.; Mancebo, E.; Calvo-Pulido, J.; Rodríguez-Muñoz, S.; Bernardo, I.; Laguna-Goya, R.; Cano-Romero, F.L.; García-Sesma, A.; Loinaz, C.; Jiménez, C.; et al. Innate Lymphoid Cells Groups 1 and 3 in the Epithelial Compartment of Functional Human Intestinal Allografts. Am. J. Transplant. 2016, 16, 72–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Acker, A.; Gronke, K.; Biswas, A.; Martens, L.; Saeys, Y.; Filtjens, J.; Taveirne, S.; Van Ammel, E.; Kerre, T.; Matthys, P.; et al. A Murine Intestinal Intraepithelial NKp46-Negative Innate Lymphoid Cell Population Characterized by Group 1 Properties. Cell Rep. 2017, 19, 1431–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ettersperger, J.; Montcuquet, N.; Malamut, G.; Guegan, N.; Lopez-Lastra, S.; Gayraud, S.; Reimann, C.; Vidal, E.; Cagnard, N.; Villarese, P.; et al. Interleukin-15-Dependent T-Cell-like Innate Intraepithelial Lymphocytes Develop in the Intestine and Transform into Lymphomas in Celiac Disease. Immunity 2016, 45, 610–625. [Google Scholar] [CrossRef]
- Olivares-Villagómez, D.; Van Kaer, L. iCD8α cells: Living at the edge of the intestinal immune system. Oncotarget 2015, 6, 19964–19965. [Google Scholar] [CrossRef]
- Van Kaer, L.; Algood, H.M.S.; Singh, K.; Parekh, V.V.; Greer, M.J.; Piazuelo, M.B.; Weitkamp, J.H.; Matta, P.; Chaturvedi, R.; Wilson, K.T.; et al. CD8αα(+) innate-type lymphocytes in the intestinal epithelium mediate mucosal immunity. Immunity 2014, 41, 451–464. [Google Scholar] [CrossRef] [Green Version]
- Lanier, L.L.; Chang, C.; Spits, H.; Phillips, J.H. Expression of cytoplasmic CD3 epsilon proteins in activated human adult natural killer (NK) cells and CD3 gamma, delta, epsilon complexes in fetal NK-cells. Implications for the relationship of NK and T lymphocytes. J. Immunol. 1992, 149, 1876–1880. [Google Scholar]
- Lopez-Verges, S.; Milush, J.M.; Pandey, S.; York, V.A.; Arakawa-Hoyt, J.; Pircher, H.; Norris, P.J.; Nixon, D.F.; Lanier, L.L. CD57 defines a functionally distinct population of mature NK-cells in the human CD56dimCD16+ NK-cell subset. Blood 2010, 116, 3865–3874. [Google Scholar] [CrossRef] [Green Version]
- Beziat, V.; Duffy, D.; Quoc, S.N.; Le Garff-Tavernier, M.; Decocq, J.; Combadiere, B.; Debre, P.; Vieillard, V. CD56brightCD16+ NK-cells: A functional intermediate stage of NK-cell differentiation. J. Immunol. 2011, 186, 6753–6761. [Google Scholar] [CrossRef] [Green Version]
- Montaldo, E.; Del Zotto, G.; Della Chiesa, M.; Mingari, M.C.; Moretta, A.; De Maria, A.; Moretta, L. Human NK-cell receptors/markers: A tool to analyze NK-cell development, subsets and function. Cytom. Part A 2013, 83, 702–713. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, C.; Juelke, K.; Falco, M.; Morandi, B.; D’Agostino, A.; Costa, R.; Ratto, G.; Forte, G.; Carrega, P.; Lui, G.; et al. CD56brightCD16- killer Ig-like receptor- NK-cells display longer telomeres and acquire features of CD56dim NK-cells upon activation. J. Immunol. 2007, 178, 4947–4955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abel, A.M.; Yang, C.; Thakar, M.S.; Malarkannan, S. Natural Killer Cells: Development, Maturation, and Clinical Utilization. Front. Immunol. 2018, 9, 1869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scoville, S.D.; Freud, A.G.; Caligiuri, M.A. Modeling Human Natural Killer Cell Development in the Era of Innate Lymphoid Cells. Front. Immunol. 2017, 8, 360. [Google Scholar] [CrossRef] [Green Version]
- Mattiola, I.; Pesant, M.; Tentorio, P.F.; Molgora, M.; Marcenaro, E.; Lugli, E.; Locati, M.; Mavilio, D. Priming of Human Resting NK-cells by Autologous M1 Macrophages via the Engagement of IL-1beta, IFN-beta, and IL-15 Pathways. J. Immunol. 2015, 195, 2818–2828. [Google Scholar] [CrossRef] [Green Version]
- Vitale, M.; Bottino, C.; Sivori, S.; Sanseverino, L.; Castriconi, R.; Marcenaro, E.; Augugliaro, R.; Moretta, L.; Moretta, A. NKp44, a novel triggering surface molecule specifically expressed by activated natural killer cells, is involved in non-major histocompatibility complex-restricted tumor cell lysis. J. Exp. Med. 1998, 187, 2065–2072. [Google Scholar] [CrossRef]
- Cooper, M.A.; Fehniger, T.A.; Turner, S.C.; Chen, K.S.; Ghaheri, B.A.; Ghayur, T.; Carson, W.E.; Caligiuri, M.A. Human natural killer cells: A unique innate immunoregulatory role for the CD56(bright) subset. Blood 2001, 97, 3146–3151. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, R.; Hintzen, G.; Kemper, A.; Beul, K.; Kempf, S.; Behrens, G.; Sykora, K.W.; Schmidt, R.E. CD56bright cells differ in their KIR repertoire and cytotoxic features from CD56dim NK-cells. Eur. J. Immunol. 2001, 31, 3121–3127. [Google Scholar] [CrossRef]
- Chan, A.; Hong, D.L.; Atzberger, A.; Kollnberger, S.; Filer, A.D.; Buckley, C.D.; McMichael, A.; Enver, T.; Bowness, P. CD56bright human NK-cells differentiate into CD56dim cells: Role of contact with peripheral fibroblasts. J. Immunol. 2007, 179, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Loza, M.J.; Perussia, B. The IL-12 signature: NK-cell terminal CD56+high stage and effector functions. J. Immunol. 2004, 172, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Geng, J.; Raghavan, M. CD8αα homodimers function as a coreceptor for KIR3DL1. Proc. Natl. Acad. Sci. USA 2019, 116, 17951–17956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernstein, H.B.; Plasterer, M.C.; Schiff, S.E.; Kitchen, C.M.; Kitchen, S.; Zack, J.A. CD4 expression on activated NK-cells: Ligation of CD4 induces cytokine expression and cell migration. J. Immunol. 2006, 177, 3669–3676. [Google Scholar] [CrossRef] [PubMed]
- Lugthart, G.; Melsen, J.E.; Vervat, C.; van Ostaijen-Ten Dam, M.M.; Corver, W.E.; Roelen, D.L.; van Bergen, J.; van Tol, M.J.; Lankester, A.C.; Schilham, M.W. Human Lymphoid Tissues Harbor a Distinct CD69+CXCR6+ NK-cell Population. J. Immunol. 2016, 197, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Hudspeth, K.; Donadon, M.; Cimino, M.; Pontarini, E.; Tentorio, P.; Preti, M.; Hong, M.; Bertoletti, A.; Bicciato, S.; Invernizzi, P.; et al. Human liver-resident CD56(bright)/CD16(neg) NK-cells are retained within hepatic sinusoids via the engagement of CCR5 and CXCR6 pathways. J. Autoimmun. 2016, 66, 40–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melsen, J.E.; Lugthart, G.; Lankester, A.C.; Schilham, M.W. Human Circulating and Tissue-Resident CD56(bright) Natural Killer Cell Populations. Front. Immunol. 2016, 7, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juelke, K.; Killig, M.; Luetke-Eversloh, M.; Parente, E.; Gruen, J.; Morandi, B.; Ferlazzo, G.; Thiel, A.; Schmitt-Knosalla, I.; Romagnani, C. CD62L expression identifies a unique subset of polyfunctional CD56dim NK-cells. Blood 2010, 116, 1299–1307. [Google Scholar] [CrossRef] [Green Version]
- Poggi, A.; Benelli, R.; Vene, R.; Costa, D.; Ferrari, N.; Tosetti, F.; Zocchi, M.R. Human Gut-Associated Natural Killer Cells in Health and Disease. Front. Immunol. 2019, 10, 961. [Google Scholar] [CrossRef] [Green Version]
- Parihar, R.; Dierksheide, J.; Hu, Y.; Carson, W.E. IL-12 enhances the natural killer cell cytokine response to Ab-coated tumor cells. J. Clin. Investig. 2002, 110, 983–992. [Google Scholar] [CrossRef]
- Artis, D.; Spits, H. The biology of innate lymphoid cells. Nature 2015, 517, 293–301. [Google Scholar] [CrossRef]
- Ma, H.; Tao, W.; Zhu, S. T lymphocytes in the intestinal mucosa: Defense and tolerance. Cell Mol. Immunol. 2019, 16, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Leishman, A.J.; Gapin, L.; Capone, M.; Palmer, E.; MacDonald, H.R.; Kronenberg, M.; Cheroutre, H. Precursors of functional MHC class I- or class II-restricted CD8αα(+) T cells are positively selected in the thymus by agonist self-peptides. Immunity 2002, 16, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Peng, K.; Liu, M.; Xiao, W.; Yang, H. CD8αα TCRalphabeta Intraepithelial Lymphocytes in the Mouse Gut. Dig. Dis. Sci. 2016, 61, 1451–1460. [Google Scholar] [CrossRef]
- Bai, L.; Peng, H. Generating CD8αα IELs from two sources of thymic precursors. Cell Mol. Immunol. 2018, 15, 640–641. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Qiu, Y.; Yang, H. Intestinal intraepithelial lymphocytes: Maintainers of intestinal immune tolerance and regulators of intestinal immunity. J. Leukoc. Biol. 2021, 109, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Cheroutre, H.; Lambolez, F.; Mucida, D. The light and dark sides of intestinal intraepithelial lymphocytes. Nat. Rev. Immunol. 2011, 11, 445–456. [Google Scholar] [CrossRef] [Green Version]
- Denning, T.L.; Granger, S.W.; Mucida, D.; Graddy, R.; Leclercq, G.; Zhang, W.; Honey, K.; Rasmussen, J.P.; Cheroutre, H.; Rudensky, A.Y.; et al. Mouse TCRalphabeta+CD8αα intraepithelial lymphocytes express genes that down-regulate their antigen reactivity and suppress immune responses. J. Immunol. 2007, 178, 4230–4239. [Google Scholar] [CrossRef] [Green Version]
- Olivares-Villagomez, D.; Van Kaer, L. Intestinal Intraepithelial Lymphocytes: Sentinels of the Mucosal Barrier. Trends Immunol. 2018, 39, 264–275. [Google Scholar] [CrossRef]
- Van Kaer, L.; Olivares-Villagomez, D. Development, Homeostasis, and Functions of Intestinal Intraepithelial Lymphocytes. J. Immunol. 2018, 200, 2235–2244. [Google Scholar] [CrossRef] [Green Version]
- Reis, B.S.; Hoytema van Konijnenburg, D.P.; Grivennikov, S.I.; Mucida, D. Transcription factor T-bet regulates intraepithelial lymphocyte functional maturation. Immunity 2014, 41, 244–256. [Google Scholar] [CrossRef] [Green Version]
- Das, G.; Augustine, M.M.; Das, J.; Bottomly, K.; Ray, P.; Ray, A. An important regulatory role for CD4+CD8 alpha alpha T cells in the intestinal epithelial layer in the prevention of inflammatory bowel disease. Proc. Natl. Acad. Sci. USA 2003, 100, 5324–5329. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Qiu, Y.; Yang, H. CD4CD8αα IELs: They Have Something to Say. Front. Immunol. 2019, 10, 2269. [Google Scholar] [CrossRef] [PubMed]
- He, X.; He, X.; Dave, V.P.; Zhang, Y.; Hua, X.; Nicolas, E.; Xu, W.; Roe, B.A.; Kappes, D.J. The zinc finger transcription factor Th-POK regulates CD4 versus CD8 T-cell lineage commitment. Nature 2005, 433, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Taniuchi, I.; Osato, M.; Egawa, T.; Sunshine, M.J.; Bae, S.C.; Komori, T.; Ito, Y.; Littman, D.R. Differential requirements for Runx proteins in CD4 repression and epigenetic silencing during T lymphocyte development. Cell 2002, 111, 621–633. [Google Scholar] [CrossRef] [Green Version]
- Cervantes-Barragan, L.; Chai, J.N.; Tianero, M.D.; Di Luccia, B.; Ahern, P.P.; Merriman, J.; Cortez, V.S.; Caparon, M.G.; Donia, M.S.; Gilfillan, S.; et al. Lactobacillus reuteri induces gut intraepithelial CD4(+)CD8αα(+) T cells. Science 2017, 357, 806–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwong, B.; Lazarevic, V. T-bet orchestrates CD8αα IEL differentiation. Immunity 2014, 41, 169–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masopust, D.; Vezys, V.; Wherry, E.J.; Barber, D.L.; Ahmed, R. Cutting edge: Gut microenvironment promotes differentiation of a unique memory CD8 T cell population. J. Immunol. 2006, 176, 2079–2083. [Google Scholar] [CrossRef] [Green Version]
- Konkel, J.E.; Maruyama, T.; Carpenter, A.C.; Xiong, Y.; Zamarron, B.F.; Hall, B.E.; Kulkarni, A.B.; Zhang, P.; Bosselut, R.; Chen, W. Control of the development of CD8αα+ intestinal intraepithelial lymphocytes by TGF-beta. Nat. Immunol. 2011, 12, 312–319. [Google Scholar] [CrossRef] [Green Version]
- Cervantes-Barragan, L.; Colonna, M. AHR signaling in the development and function of intestinal immune cells and beyond. Semin. ImmunoPathol. 2018, 40, 371–377. [Google Scholar] [CrossRef]
- Yu, S.; Bruce, D.; Froicu, M.; Weaver, V.; Cantorna, M.T. Failure of T cell homing, reduced CD4/CD8αα intraepithelial lymphocytes, and inflammation in the gut of vitamin D receptor KO mice. Proc. Natl. Acad. Sci. USA 2008, 105, 20834–20839. [Google Scholar] [CrossRef] [Green Version]
- Mayassi, T.; Jabri, B. Human intraepithelial lymphocytes. Mucosal Immunol. 2018, 11, 1281–1289. [Google Scholar] [CrossRef]
- Bhagat, G.; Naiyer, A.J.; Shah, J.G.; Harper, J.; Jabri, B.; Wang, T.C.; Green, P.H.; Manavalan, J.S. Small intestinal CD8+TCRgammadelta+NKG2A+ intraepithelial lymphocytes have attributes of regulatory cells in patients with celiac disease. J. Clin. Investig. 2008, 118, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Mikulak, J.; Oriolo, F.; Bruni, E.; Roberto, A.; Colombo, F.S.; Villa, A.; Bosticardo, M.; Bortolomai, I.; Lo Presti, E.; Meraviglia, S.; et al. NKp46-expressing human gut-resident intraepithelial Vdelta1 T cell subpopulation exhibits high antitumor activity against colorectal cancer. JCI Insight 2019, 4, e125884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cawthon, A.G.; Alexander-Miller, M.A. Optimal colocalization of TCR and CD8 as a novel mechanism for the control of functional avidity. J. Immunol. 2002, 169, 3492–3498. [Google Scholar] [CrossRef] [Green Version]
- Cawthon, A.G.; Lu, H.; Alexander-Miller, M.A. Peptide requirement for CTL activation reflects the sensitivity to CD3 engagement: Correlation with CD8alphabeta versus CD8αα expression. J. Immunol. 2001, 167, 2577–2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheroutre, H.; Lambolez, F. Doubting the TCR coreceptor function of CD8αα. Immunity 2008, 28, 149–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, D.; Morgan, E.A.; Berger, D.; Pinkus, G.S.; Ferry, J.A.; Zukerberg, L.R. NK-Cell Enteropathy and Similar Indolent Lymphoproliferative Disorders: A Case Series With Literature Review. Am. J. Clin. Pathol. 2019, 151, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, K.; Yokoyama, M.; Ishizawa, S.; Terui, Y.; Nomura, K.; Marutsuka, K.; Nunomura, M.; Fukushima, N.; Yagyuu, T.; Nakamine, H.; et al. Lymphomatoid gastropathy: A distinct clinicopathologic entity of self-limited pseudomalignant NK-cell proliferation. Blood 2010, 116, 5631–5637. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Matsuzono, E.; Yokoyama, F.; Ohara, Y.; Sugai, N.; Seki, H.; Miura, A.; Fujita, J.; Suzuki, J.; Fujisawa, T.; et al. A case of lymphomatoid gastropathy: A self-limited pseudomalignant natural killer (NK)-cell proliferative disease mimicking NK/T-cell lymphomas. Clin. J. Gastroenterol. 2013, 6, 287–290. [Google Scholar] [CrossRef]
- Tanaka, T.; Megahed, N.; Takata, K.; Asano, N.; Niwa, Y.; Hirooka, Y.; Goto, H. A case of lymphomatoid gastropathy: An indolent CD56-positive atypical gastric lymphoid proliferation, mimicking aggressive NK/T cell lymphomas. Pathol. Res. Pract. 2011, 207, 786–789. [Google Scholar] [CrossRef]
- Terai, T.; Sugimoto, M.; Uozaki, H.; Kitagawa, T.; Kinoshita, M.; Baba, S.; Yamada, T.; Osawa, S.; Sugimoto, K. Lymphomatoidgastropathy mimicking extranodal NK/T cell lymphoma, nasal type: A case report. World J. Gastroenterol. 2012, 18, 2140–2144. [Google Scholar] [CrossRef]
- Koh, J.; Go, H.; Lee, W.A.; Jeon, Y.K. Benign Indolent CD56-Positive NK-Cell Lymphoproliferative Lesion Involving Gastrointestinal Tract in an Adolescent. Korean J. Pathol. 2014, 48, 73–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansoor, A.; Pittaluga, S.; Beck, P.L.; Wilson, W.H.; Ferry, J.A.; Jaffe, E.S. NK-cell enteropathy: A benign NK-cell lymphoproliferative disease mimicking intestinal lymphoma: Clinicopathologic features and follow-up in a unique case series. Blood 2011, 117, 1447–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McElroy, M.K.; Read, W.L.; Harmon, G.S.; Weidner, N. A unique case of an indolent CD56-positive T-cell lymphoproliferative disorder of the gastrointestinal tract: A lesion potentially misdiagnosed as natural killer/T-cell lymphoma. Ann. Diagn. Pathol. 2011, 15, 370–375. [Google Scholar] [CrossRef]
- Vega, F.; Chang, C.C.; Schwartz, M.R.; Preti, H.A.; Younes, M.; Ewton, A.; Verm, R.; Jaffe, E.S. Atypical NK-cell proliferation of the gastrointestinal tract in a patient with antigliadin antibodies but not celiac disease. Am. J. Surg. Pathol. 2006, 30, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Isom, J.A.; Arroyo, M.R.; Reddy, D.; Joshi-Guske, P.; Al-Quran, S.Z.; Li, Y.; Allan, R.W. NK-cell enteropathy: A case report with 10 years of indolent clinical behaviour. Histopathology 2018, 73, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Takata, K.; Noujima-Harada, M.; Miyata-Takata, T.; Ichimura, K.; Sato, Y.; Miyata, T.; Naruse, K.; Iwamoto, T.; Tari, A.; Masunari, T.; et al. Clinicopathologic analysis of 6 lymphomatoid gastropathy cases: Expanding the disease spectrum to CD4-CD8+ cases. Am. J. Surg. Pathol. 2015, 39, 1259–1266. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, J.; Fujishima, F.; Ichinohasama, R.; Imatani, A.; Asano, N.; Harigae, H. A case of benign natural killer cell proliferative disorder of the stomach (gastric manifestation of natural killer cell lymphomatoid gastroenteropathy) mimicking extranodal natural killer/T-cell lymphoma. Leuk. Lymphoma 2011, 52, 1803–1805. [Google Scholar] [CrossRef] [PubMed]
- Montes-Moreno, S.; King, R.L.; Oschlies, I.; Ponzoni, M.; Goodlad, J.R.; Dotlic, S.; Traverse-Glehen, A.; Ott, G.; Ferry, J.A.; Calaminici, M. Update on lymphoproliferative disorders of the gastrointestinal tract: Disease spectrum from indolent lymphoproliferations to aggressive lymphomas. Virchows Arch. 2020, 476, 667–681. [Google Scholar] [CrossRef]
- Freud, A.G.; Zhao, S.; Wei, S.; Gitana, G.M.; Molina-Kirsch, H.F.; Atwater, S.K.; Natkunam, Y. Expression of the activating receptor, NKp46 (CD335), in human natural killer and T-cell neoplasia. Am. J. Clin. Pathol. 2013, 140, 853–866. [Google Scholar] [CrossRef] [Green Version]
- Xiao, W.; Gupta, G.K.; Yao, J.; Jang, Y.J.; Xi, L.; Baik, J.; Sigler, A.; Kumar, A.; Moskowitz, A.J.; Arcila, M.E.; et al. Recurrent somatic JAK3 mutations in NK-cell enteropathy. Blood 2019, 134, 986–991. [Google Scholar] [CrossRef]
- Harabuchi, Y.; Takahara, M.; Kishibe, K.; Nagato, T.; Kumai, T. Extranodal Natural Killer/T-Cell Lymphoma, Nasal Type: Basic Science and Clinical Progress. Front. Pediatr. 2019, 7, 141. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Choi, C.W.; Mun, Y.C.; Oh, S.Y.; Kang, H.J.; Lee, S.I.; Won, J.H.; Kim, M.K.; Kwon, J.H.; Kim, J.S.; et al. Multicenter retrospective analysis of 581 patients with primary intestinal non-hodgkin lymphoma from the Consortium for Improving Survival of Lymphoma (CISL). BMC Cancer 2011, 11, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, B.H.; Shui, R.H.; Sheng, W.Q.; Wang, C.F.; Lu, H.F.; Zhou, X.Y.; Zhu, X.Z.; Li, X.Q. Primary Intestinal Extranodal Natural Killer/T-Cell Lymphoma, Nasal.l Type: A Comprehensive Clinicopathological Analysis of 55 Cases. PLoS ONE 2016, 11, e0161831. [Google Scholar] [CrossRef]
- Li, Z.; Xia, Y.; Feng, L.N.; Chen, J.R.; Li, H.M.; Cui, J.; Cai, Q.Q.; Sim, K.S.; Nairismagi, M.L.; Laurensia, Y.; et al. Genetic risk of extranodal natural killer T-cell lymphoma: A genome-wide association study. Lancet Oncol. 2016, 17, 1240–1247. [Google Scholar] [CrossRef]
- Lin, G.W.; Xu, C.; Chen, K.; Huang, H.Q.; Chen, J.; Song, B.; Chan, J.K.C.; Li, W.; Liu, W.; Shih, L.Y.; et al. Genetic risk of extranodal natural killer T-cell lymphoma: A genome-wide association study in multiple populations. Lancet Oncol. 2020, 21, 306–316. [Google Scholar] [CrossRef]
- Kim, S.J.; Jung, H.A.; Chuang, S.S.; Hong, H.; Guo, C.C.; Cao, J.; Hong, X.N.; Suzuki, R.; Kang, H.J.; Won, J.H.; et al. Extranodal natural killer/T-cell lymphoma involving the gastrointestinal tract: Analysis of clinical features and outcomes from the Asia Lymphoma Study Group. J. Hematol. Oncol. 2013, 6, 86. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Chen, X.; Yi, Z.; Zhang, X.; Zhang, B.; Luo, F.; Jiang, Y.; Zou, L. Prognostic characteristics of gastrointestinal tract NK/T-cell lymphoma: An analysis of 47 patients in China. J. Clin. Gastroenterol. 2013, 47, e74–e79. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, J.H.; Lee, J.; Oh, S.O.; Chang, D.K.; Rhee, P.L.; Kim, J.J.; Rhee, J.C.; Lee, J.; Kim, W.S.; et al. Primary NK-/T-cell lymphoma of the gastrointestinal tract: Clinical characteristics and endoscopic findings. Endoscopy 2007, 39, 156–160. [Google Scholar] [CrossRef]
- Lee, G.; Ryu, H.J.; Choi, J.W.; Kang, H.; Yang, W.I.; Yang, I.S.; Seo, M.K.; Kim, S.; Yoon, S.O. Characteristic gene alterations in primary gastrointestinal T- and NK-cell lymphomas. Leukemia 2019, 33, 1797–1832. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, R.; Suzumiya, J.; Yamaguchi, M.; Nakamura, S.; Kameoka, J.; Kojima, H.; Abe, M.; Kinoshita, T.; Yoshino, T.; Iwatsuki, K.; et al. Prognostic factors for mature natural killer (NK) cell neoplasms: Aggressive NK-cell leukemia and extranodal NK-cell lymphoma, nasal type. Ann. Oncol. 2010, 21, 1032–1040. [Google Scholar] [CrossRef]
- Tang, X.F.; Yang, L.; Duan, S.; Guo, H.; Guo, Q.N. Intestinal T-cell and NK/T-cell lymphomas: A clinicopathological study of 27 Chinese patients. Ann. Diagn. Pathol. 2018, 37, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.K.C.; Quintanilla-Martinez, L.; Ferry, J.A. Extranodal NK/T-cell lymphoma, nasal type. In WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues; Swerdlow, S.H., Campo, E., Harris, N.L., Jaffe, E.S., Pileri, S.A., Stein, H., Thiele, J., Arber, D.A., Hasserjian, R.P., Le Beau, M.M., et al., Eds.; International Agency for Research on Cancer: Lyon, France, 2017; pp. 368–371. [Google Scholar]
- Ko, Y.H.; Li, G.D.; Takeuchi, K. Extranodal NK/T cell lymphoma. In Digestive System Tumours; WHO Classification of Tumours Editorial Board, Ed.; International Agency for Research on Cancer: Lyon, France, 2019; pp. 420–421. [Google Scholar]
- Dukers, D.F.; Vermeer, M.H.; Jaspars, L.H.; Sander, C.A.; Flaig, M.J.; Vos, W.; Willemze, R.; Meijer, C.J. Expression of killer cell inhibitory receptors is restricted to true NK-cell lymphomas and a subset of intestinal enteropathy-type T cell lymphomas with a cytotoxic phenotype. J. Clin. Pathol. 2001, 54, 224–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uemura, Y.; Isobe, Y.; Uchida, A.; Asano, J.; Nishio, Y.; Sakai, H.; Hoshikawa, M.; Takagi, M.; Nakamura, N.; Miura, I. Expression of activating natural killer-cell receptors is a hallmark of the innate-like T-cell neoplasm in peripheral T-cell lymphomas. Cancer Sci. 2018, 109, 1254–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishibashi, H.; Nimura, S.; Ishitsuka, K.; Mihashi, Y.; Mizoguchi, M.; Nakamura, S.; Okamura, S.; Momosaki, S.; Aoyagi, K.; Sakisaka, S.; et al. High Expression of Intestinal Homing Receptor CD103 in Adult T-Cell Leukemia/Lymphoma, Similar to 2 Other CD8+ T-Cell Lymphomas. Am. J. Surg. Pathol. 2016, 40, 462–470. [Google Scholar] [CrossRef] [PubMed]
- de Mel, S.; Li, J.B.; Abid, M.B.; Tang, T. The utility of flow cytometry in differentiating NK/T cell lymphoma from indolent and reactive NK-cell proliferations. Cytom. Part. B Clin. Cytom. 2018, 94, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Lima, M. Extranodal NK/T cell lymphoma and aggressive NK-cell leukaemia: Evidence for their origin on CD56+bright CD16-/+dim NK-cells. Pathology 2015, 47, 503–514. [Google Scholar] [CrossRef]
- Au, W.Y.; Weisenburger, D.D.; Intragumtornchai, T.; Nakamura, S.; Kim, W.S.; Sng, I.; Vose, J.; Armitage, J.O.; Liang, R.; International Peripheral, T.C.L.P. Clinical differences between nasal and extranasal natural killer/T-cell lymphoma: A study of 136 cases from the International Peripheral T-Cell Lymphoma Project. Blood 2009, 113, 3931–3937. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Zhou, X.; Song, A.; Huo, Z.; Wang, Y.; Liu, Y. Nasal-type extranodal natural killer/T-cell lymphoma presenting with a mass on the buttock: A case report. Medicine 2019, 98, e18260. [Google Scholar] [CrossRef]
- Ng, S.B.; Lai, K.W.; Murugaya, S.; Lee, K.M.; Loong, S.L.; Fook-Chong, S.; Tao, M.; Sng, I. Nasal-type extranodal natural killer/T-cell lymphomas: A clinicopathologic and genotypic study of 42 cases in Singapore. Mod. Pathol. 2004, 17, 1097–1107. [Google Scholar] [CrossRef]
- Harabuchi, Y.; Imai, S.; Wakashima, J.; Hirao, M.; Kataura, A.; Osato, T.; Kon, S. Nasal T-cell lymphoma causally associated with Epstein-Barr virus: Clinicopathologic, phenotypic, and genotypic studies. Cancer 1996, 77, 2137–2149. [Google Scholar] [CrossRef]
- Nagata, H.; Konno, A.; Kimura, N.; Zhang, Y.; Kimura, M.; Demachi, A.; Sekine, T.; Yamamoto, K.; Shimizu, N. Characterization of novel natural killer (NK)-cell and gammadelta T-cell lines established from primary lesions of nasal T/NK-cell lymphomas associated with the Epstein-Barr virus. Blood 2001, 97, 708–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pongpruttipan, T.; Sukpanichnant, S.; Assanasen, T.; Wannakrairot, P.; Boonsakan, P.; Kanoksil, W.; Kayasut, K.; Mitarnun, W.; Khuhapinant, A.; Bunworasate, U.; et al. Extranodal NK/T-cell lymphoma, nasal type, includes cases of natural killer cell and alphabeta, gammadelta, and alphabeta/gammadelta T-cell origin: A comprehensive clinicopathologic and phenotypic study. Am. J. Surg. Pathol. 2012, 36, 481–499. [Google Scholar] [CrossRef] [PubMed]
- Au, W.Y.; Ma, S.Y.; Chim, C.S.; Choy, C.; Loong, F.; Lie, A.K.; Lam, C.C.; Leung, A.Y.; Tse, E.; Yau, C.C.; et al. Clinicopathologic features and treatment outcome of mature T-cell and natural killer-cell lymphomas diagnosed according to the World Health Organization classification scheme: A single center experience of 10 years. Ann. Oncol. 2005, 16, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Gaal, K.; Sun, N.C.; Hernandez, A.M.; Arber, D.A. Sinonasal NK/T-cell lymphomas in the United States. Am. J. Surg. Pathol. 2000, 24, 1511–1517. [Google Scholar] [CrossRef]
- Schwartz, E.J.; Molina-Kirsch, H.; Zhao, S.; Marinelli, R.J.; Warnke, R.A.; Natkunam, Y. Immunohistochemical characterization of nasal-type extranodal NK/T-cell lymphoma using a tissue microarray: An analysis of 84 cases. Am. J. Clin. Pathol. 2008, 130, 343–351. [Google Scholar] [CrossRef]
- Heller, N.B.-B.R.; Naler, L.; Sen, J.M. Natural Killer T (NKT) Cells in Mice and Men. In Signaling Mechanisms Regulating T Cell Diversity and Function; Soboloff, J.K.D., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2018; pp. 119–146. [Google Scholar]
- Krovi, S.H.; Gapin, L. Invariant Natural Killer T Cell Subsets—More Than Just Developmental Intermediates. Front. Immunol. 2018, 9, 1393. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Mitsui, T.; Wei, M.; Mao, H.; Butchar, J.P.; Shah, M.V.; Zhang, J.; Mishra, A.; Alvarez-Breckenridge, C.; Liu, X.; et al. NKp46 identifies an NKT cell subset susceptible to leukemic transformation in mouse and human. J. Clin. Investig. 2011, 121, 1456–1470. [Google Scholar] [CrossRef] [Green Version]
- Cohavy, O.; Targan, S.R. CD56.6 Marks an Effector T Cell Subset in the Human Intestine. J. Immunol. 2007, 178, 5524–5532. [Google Scholar] [CrossRef] [Green Version]
- de Mel, S.; Hue, S.S.; Jeyasekharan, A.D.; Chng, W.J.; Ng, S.B. Molecular pathogenic pathways in extranodal NK/T cell lymphoma. J. Hematol. Oncol. 2019, 12, 33. [Google Scholar] [CrossRef]
- Ng, S.B.; Selvarajan, V.; Huang, G.; Zhou, J.; Feldman, A.L.; Law, M.; Kwong, Y.L.; Shimizu, N.; Kagami, Y.; Aozasa, K.; et al. Activated oncogenic pathways and therapeutic targets in extranodal nasal-type NK/T cell lymphoma revealed by gene expression profiling. J. Pathol. 2011, 223, 496–510. [Google Scholar] [CrossRef]
- Saleem, A.; Natkunam, Y. Extranodal NK/T-Cell Lymphomas: The Role of Natural Killer Cells and EBV in Lymphomagenesis. Int. J. Mol. Sci. 2020, 21, 1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchekioua, A.; Scourzic, L.; de Wever, O.; Zhang, Y.; Cervera, P.; Aline-Fardin, A.; Mercher, T.; Gaulard, P.; Nyga, R.; Jeziorowska, D.; et al. JAK3 deregulation by activating mutations confers invasive growth advantage in extranodal nasal-type natural killer cell lymphoma. Leukemia 2014, 28, 338–348. [Google Scholar] [CrossRef] [PubMed]
- Koo, G.C.; Tan, S.Y.; Tang, T.; Poon, S.L.; Allen, G.E.; Tan, L.; Chong, S.C.; Ong, W.S.; Tay, K.; Tao, M.; et al. Janus kinase 3-activating mutations identified in natural killer/T-cell lymphoma. Cancer Discov. 2012, 2, 591–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, T.L.; Nairismagi, M.L.; Laurensia, Y.; Lim, J.Q.; Tan, J.; Li, Z.M.; Pang, W.L.; Kizhakeyil, A.; Wijaya, G.C.; Huang, D.C.; et al. Oncogenic activation of the STAT3 pathway drives PD-L1 expression in natural killer/T-cell lymphoma. Blood 2018, 132, 1146–1158. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; de Reynies, A.; de Leval, L.; Ghazi, B.; Martin-Garcia, N.; Travert, M.; Bosq, J.; Briere, J.; Petit, B.; Thomas, E.; et al. Gene expression profiling identifies emerging oncogenic pathways operating in extranodal NK/T-cell lymphoma, nasal type. Blood 2010, 115, 1226–1237. [Google Scholar] [CrossRef]
- Iqbal, J.; Weisenburger, D.D.; Chowdhury, A.; Tsai, M.Y.; Srivastava, G.; Greiner, T.C.; Kucuk, C.; Deffenbacher, K.; Vose, J.; Smith, L.; et al. Natural killer cell lymphoma shares strikingly similar molecular features with a group of non-hepatosplenic gammadelta T-cell lymphoma and is highly sensitive to a novel aurora kinase A inhibitor in vitro. Leukemia 2011, 25, 348–358. [Google Scholar] [CrossRef] [Green Version]
- Dobashi, A.; Tsuyama, N.; Asaka, R.; Togashi, Y.; Ueda, K.; Sakata, S.; Baba, S.; Sakamoto, K.; Hatake, K.; Takeuchi, K. Frequent BCOR aberrations in extranodal NK/T-Cell lymphoma, nasal type. Genes Chromosomes Cancer 2016, 55, 460–471. [Google Scholar] [CrossRef]
- Lee, S.; Park, H.Y.; Kang, S.Y.; Kim, S.J.; Hwang, J.; Lee, S.; Kwak, S.H.; Park, K.S.; Yoo, H.Y.; Kim, W.S.; et al. Genetic alterations of JAK/STAT cascade and histone modification in extranodal NK/T-cell lymphoma nasal type. Oncotarget 2015, 6, 17764–17776. [Google Scholar] [CrossRef] [Green Version]
- Hongyo, T.; Li, T.; Syaifudin, M.; Baskar, R.; Ikeda, H.; Kanakura, Y.; Aozasa, K.; Nomura, T. Specific c-kit mutations in sinonasal natural killer/T-cell lymphoma in China and Japan. Cancer Res. 2000, 60, 2345–2347. [Google Scholar]
- de Mel, S.; Soon, G.S.; Mok, Y.; Chung, T.H.; Jeyasekharan, A.D.; Chng, W.J.; Ng, S.B. The Genomics and Molecular Biology of Natural Killer/T-Cell Lymphoma: Opportunities for Translation. Int. J. Mol. Sci. 2018, 19, 1931. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Gu, Z.H.; Yan, Z.X.; Zhao, X.; Xie, Y.Y.; Zhang, Z.G.; Pan, C.M.; Hu, Y.; Cai, C.P.; Dong, Y.; et al. Exome sequencing identifies somatic mutations of DDX3X in natural killer/T-cell lymphoma. Nat. Genet. 2015, 47, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Quintanilla-Martinez, L.; Kremer, M.; Keller, G.; Nathrath, M.; Gamboa-Dominguez, A.; Meneses, A.; Luna-Contreras, L.; Cabras, A.; Hoefler, H.; Mohar, A.; et al. p53 Mutations in nasal natural killer/T-cell lymphoma from Mexico: Association with large cell morphology and advanced disease. Am. J. Pathol. 2001, 159, 2095–2105. [Google Scholar] [CrossRef]
- Chen, Y.W.; Guo, T.; Shen, L.; Wong, K.Y.; Tao, Q.; Choi, W.W.; Au-Yeung, R.K.; Chan, Y.P.; Wong, M.L.; Tang, J.C.; et al. Receptor-type tyrosine-protein phosphatase kappa directly targets STAT3 activation for tumor suppression in nasal NK/T-cell lymphoma. Blood 2015, 125, 1589–1600. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Kucuk, C.; Deleeuw, R.J.; Srivastava, G.; Tam, W.; Geng, H.; Klinkebiel, D.; Christman, J.K.; Patel, K.; Cao, K.; et al. Genomic analyses reveal global functional alterations that promote tumor growth and novel tumor suppressor genes in natural killer-cell malignancies. Leukemia 2009, 23, 1139–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karube, K.; Nakagawa, M.; Tsuzuki, S.; Takeuchi, I.; Honma, K.; Nakashima, Y.; Shimizu, N.; Ko, Y.H.; Morishima, Y.; Ohshima, K.; et al. Identification of FOXO3 and PRDM1 as tumor-suppressor gene candidates in NK-cell neoplasms by genomic and functional analyses. Blood 2011, 118, 3195–3204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Liang, L.; Li, D.; Nong, L.; Liu, J.; Qu, L.; Zheng, Y.; Zhang, B.; Li, T. Hypermethylation of PRDM1/Blimp-1 promoter in extranodal NK/T-cell lymphoma, nasal type: An evidence of predominant role in its downregulation. Hematol. Oncol. 2017, 35, 645–654. [Google Scholar] [CrossRef]
- Xiong, J.; Cui, B.W.; Wang, N.; Dai, Y.T.; Zhang, H.; Wang, C.F.; Zhong, H.J.; Cheng, S.; Ou-Yang, B.S.; Hu, Y.; et al. Genomic and Transcriptomic Characterization of Natural Killer T Cell Lymphoma. Cancer Cell 2020, 37, 403–419.e6. [Google Scholar] [CrossRef]
- Sanguedolce, F.; Zanelli, M.; Zizzo, M.; Luminari, S.; Martino, G.; Soriano, A.; Ricci, L.; Caprera, C.; Ascani, S. Indolent T-Cell Lymphoproliferative Disorders of the Gastrointestinal Tract (iTLPD-GI): A Review. Cancers 2021, 13, 2790. [Google Scholar] [CrossRef]
- Carbonnel, F.; d’Almagne, H.; Lavergne, A.; Matuchansky, C.; Brouet, J.C.; Sigaux, F.; Beaugerie, L.; Nemeth, J.; Coffin, B.; Cosnes, J.; et al. The clinicopathological features of extensive small intestinal CD4 T cell infiltration. Gut 1999, 45, 662–667. [Google Scholar] [CrossRef]
- Egawa, N.; Fukayama, M.; Kawaguchi, K.; Hishima, T.; Hayashi, Y.; Funata, N.; Ibuka, T.; Koike, M.; Miyashita, H.; Tajima, T. Relapsing oral and colonic ulcers with monoclonal T-cell infiltration. A low grade mucosal T-lymphoproliferative disease of the digestive tract. Cancer 1995, 75, 1728–1733. [Google Scholar] [CrossRef]
- Margolskee, E.; Jobanputra, V.; Lewis, S.K.; Alobeid, B.; Green, P.H.; Bhagat, G. Indolent small intestinal CD4+ T-cell lymphoma is a distinct entity with unique biologic and clinical features. PLoS ONE 2013, 8, e68343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, A.M.; Warnke, R.A.; Hu, Q.; Gaulard, P.; Copie-Bergman, C.; Alkan, S.; Wang, H.Y.; Cheng, J.X.; Bacon, C.M.; Delabie, J.; et al. Indolent T-cell lymphoproliferative disease of the gastrointestinal tract. Blood 2013, 122, 3599–3606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbonnel, F.; Lavergne, A.; Messing, B.; Tsapis, A.; Berger, R.; Galian, A.; Nemeth, J.; Brouet, J.C.; Rambaud, J.C. Extensive small intestinal T-cell lymphoma of low-grade malignancy associated with a new chromosomal translocation. Cancer 1994, 73, 1286–1291. [Google Scholar] [CrossRef]
- Hirakawa, K.; Fuchigami, T.; Nakamura, S.; Daimaru, Y.; Ohshima, K.; Sakai, Y.; Ichimaru, T. Primary gastrointestinal T-cell lymphoma resembling multiple lymphomatous polyposis. Gastroenterology 1996, 111, 778–782. [Google Scholar] [CrossRef] [PubMed]
- Sena Teixeira Mendes, L.; Attygalle, A.D.; Cunningham, D.; Benson, M.; Andreyev, J.; Gonzales-de-Castro, D.; Wotherspoon, A. CD4-positive small T-cell lymphoma of the intestine presenting with severe bile-acid malabsorption: A supportive symptom control approach. Br. J. Haematol. 2014, 167, 265–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svrcek, M.; Garderet, L.; Sebbagh, V.; Rosenzwajg, M.; Parc, Y.; Lagrange, M.; Bennis, M.; Lavergne-Slove, A.; Flejou, J.F.; Fabiani, B. Small intestinal CD4+ T-cell lymphoma: A rare distinctive clinicopathological entity associated with prolonged survival. Virchows Arch. 2007, 451, 1091–1093. [Google Scholar] [CrossRef]
- Tsutsumi, Y.; Inada, K.; Morita, K.; Suzuki, T. T-cell lymphomas diffusely involving the intestine: Report of two rare cases. Jpn. J. Clin. Oncol. 1996, 26, 264–272. [Google Scholar] [CrossRef] [Green Version]
- Zivny, J.; Banner, B.F.; Agrawal, S.; Pihan, G.; Barnard, G.F. CD4+ T-cell lymphoproliferative disorder of the gut clinically mimicking celiac sprue. Dig. Dis. Sci. 2004, 49, 551–555. [Google Scholar] [CrossRef]
- Soderquist, C.R.; Patel, N.; Murty, V.V.; Betman, S.; Aggarwal, N.; Young, K.H.; Xerri, L.; Leeman-Neill, R.; Lewis, S.K.; Green, P.H.; et al. Genetic and phenotypic characterization of indolent T-cell lymphoproliferative disorders of the gastrointestinal tract. Haematologica 2020, 105, 1895–1906. [Google Scholar] [CrossRef] [Green Version]
- Leventaki, V.; Manning, J.T., Jr.; Luthra, R.; Mehta, P.; Oki, Y.; Romaguera, J.E.; Medeiros, L.J.; Vega, F. Indolent peripheral T-cell lymphoma involving the gastrointestinal tract. Hum. Pathol. 2014, 45, 421–426. [Google Scholar] [CrossRef]
- Malamut, G.; Meresse, B.; Kaltenbach, S.; Derrieux, C.; Verkarre, V.; Macintyre, E.; Ruskone-Fourmestraux, A.; Fabiani, B.; Radford-Weiss, I.; Brousse, N.; et al. Small intestinal CD4+ T-cell lymphoma is a heterogenous entity with common pathology features. Clin. Gastroenterol. Hepatol. 2014, 12, 599–608.e1. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Oishi, N.; Boddicker, R.L.; Hu, G.; Benson, H.K.; Ketterling, R.P.; Greipp, P.T.; Knutson, D.L.; Kloft-Nelson, S.M.; He, R.; et al. Recurrent STAT3-JAK2 fusions in indolent T-cell lymphoproliferative disorder of the gastrointestinal tract. Blood 2018, 131, 2262–2266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranheim, E.A.; Jones, C.; Zehnder, J.L.; Warnke, R.; Yuen, A. Spontaneously relapsing clonal, mucosal cytotoxic T-cell lymphoproliferative disorder: Case report and review of the literature. Am. J. Surg. Pathol. 2000, 24, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Egan, L.J.; Walsh, S.V.; Stevens, F.M.; Connolly, C.E.; Egan, E.L.; McCarthy, C.F. Celiac-associated lymphoma. A single institution experience of 30 cases in the combination chemotherapy era. J. Clin. Gastroenterol. 1995, 21, 123–129. [Google Scholar] [CrossRef]
- Wang, X.; Ng, C.S.; Chen, C.; Yu, G.; Yin, W. An unusual case report of indolent T-cell lymphoproliferative disorder with aberrant CD20 expression involving the gastrointestinal tract and bone marrow. Diagn. Pathol. 2018, 13, 82. [Google Scholar] [CrossRef]
- Cheminant, M.; Bruneau, J.; Malamut, G.; Sibon, D.; Guegan, N.; van Gils, T.; Cording, S.; Trinquand, A.; Verkarre, V.; Lhermitte, L.; et al. NKp46 is a diagnostic biomarker and may be a therapeutic target in gastrointestinal T-cell lymphoproliferative diseases: A CELAC study. Gut 2019, 68, 1396–1405. [Google Scholar] [CrossRef]
- Matnani, R.; Ganapathi, K.A.; Lewis, S.K.; Green, P.H.; Alobeid, B.; Bhagat, G. Indolent T- and NK-cell lymphoproliferative disorders of the gastrointestinal tract: A review and update. Hematol. Oncol. 2017, 35, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Laabi, Y.; Gras, M.P.; Carbonnel, F.; Brouet, J.C.; Berger, R.; Larsen, C.J.; Tsapis, A. A new gene, BCM, on chromosome 16 is fused to the interleukin 2 gene by a t(4;16)(q26;p13) translocation in a malignant T cell lymphoma. EMBO J. 1992, 11, 3897–3904. [Google Scholar] [CrossRef]
- Singh, P.; Arora, A.; Strand, T.A.; Leffler, D.A.; Catassi, C.; Green, P.H.; Kelly, C.P.; Ahuja, V.; Makharia, G.K. Global Prevalence of Celiac Disease: Systematic Review and Meta-analysis. Clin. Gastroenterol. Hepatol 2018, 16, 823–836.e2. [Google Scholar] [CrossRef] [Green Version]
- Barker, J.M.; Liu, E. Celiac disease: Pathophysiology, clinical manifestations, and associated autoimmune conditions. Adv. Pediatr. 2008, 55, 349–365. [Google Scholar] [CrossRef] [Green Version]
- Ianiro, G.; Gasbarrini, A.; Cammarota, G. Endoscopic tools for the diagnosis and evaluation of celiac disease. World J. Gastroenterol. 2013, 19, 8562–8570. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Zhang, Q.; Olofson, A.M.; Jhala, N.; Liu, X. Celiac Disease: Updates on Pathology and Differential Diagnosis. Adv. Anat. Pathol. 2019, 26, 292–312. [Google Scholar] [CrossRef] [PubMed]
- Dickson, B.C.; Streutker, C.J.; Chetty, R. Coeliac disease: An update for pathologists. J. Clin. Pathol. 2006, 59, 1008–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Castañon, M.; Castro, B.G.; Toca, M.; Santacruz, C.; Arias-Loste, M.; Iruzubieta, P.; Crespo, J.; López-Hoyos, M. Intraepithelial lymphocytes subsets in different forms of celiac disease. Autoimmun. Highlights 2016, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steenholt, J.V.; Nielsen, C.; Baudewijn, L.; Staal, A.; Rasmussen, K.S.; Sabir, H.J.; Barington, T.; Husby, S.; Toft-Hansen, H. The composition of T cell subtypes in duodenal biopsies are altered in coeliac disease patients. PLoS ONE 2017, 12, e0170270. [Google Scholar] [CrossRef] [Green Version]
- Hüe, S.; Mention, J.J.; Monteiro, R.C.; Zhang, S.; Cellier, C.; Schmitz, J.; Verkarre, V.; Fodil, N.; Bahram, S.; Cerf-Bensussan, N.; et al. A direct role for NKG2D/MICA interaction in villous atrophy during celiac disease. Immunity 2004, 21, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Jabri, B.; De Serre, N.P.M.; Cellier, C.; Evans, K.; Gache, C.; Carvalho, C.; Mougenot, J.F.; Allez, M.; Jian, R.; Desreumaux, P.; et al. Selective expansion of intraepithelial lymphocytes expressing the HLA-E–specific natural killer receptor CD94 in celiac disease. Gastroenterology 2000, 118, 867–879. [Google Scholar] [CrossRef]
- Hudacko, R.; Kathy Zhou, X.; Yantiss, R.K. Immunohistochemical stains for CD3 and CD8 do not improve detection of gluten-sensitive enteropathy in duodenal biopsies. Mod. Pathol. 2013, 26, 1241–1245. [Google Scholar] [CrossRef] [Green Version]
- Biagi, F.; Corazza, G.R. Defining gluten refractory enteropathy. Eur. J. Gastroenterol. Hepatol. 2001, 13, 561–565. [Google Scholar] [CrossRef]
- Al-Toma, A.; Verbeek, W.H.; Hadithi, M.; von Blomberg, B.M.; Mulder, C.J. Survival in refractory coeliac disease and enteropathy-associated T-cell lymphoma: Retrospective evaluation of single-centre experience. Gut 2007, 56, 1373–1378. [Google Scholar] [CrossRef] [Green Version]
- Malamut, G.; Afchain, P.; Verkarre, V.; Lecomte, T.; Amiot, A.; Damotte, D.; Bouhnik, Y.; Colombel, J.F.; Delchier, J.C.; Allez, M.; et al. Presentation and long-term follow-up of refractory celiac disease: Comparison of type I with type II. Gastroenterology 2009, 136, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Tapia, A.; Kelly, D.G.; Lahr, B.D.; Dogan, A.; Wu, T.T.; Murray, J.A. Clinical staging and survival in refractory celiac disease: A single center experience. Gastroenterology 2009, 136, 99–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daum, S.; Wahnschaffe, U.; Glasenapp, R.; Borchert, M.; Ullrich, R.; Zeitz, M.; Faiss, S. Capsule endoscopy in refractory celiac disease. Endoscopy 2007, 39, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Daum, S.; Weiss, D.; Hummel, M.; Ullrich, R.; Heise, W.; Stein, H.; Riecken, E.O.; Foss, H.D.; Intestinal Lymphoma Study Group. Frequency of clonal intraepithelial T lymphocyte proliferations in enteropathy-type intestinal T cell lymphoma, coeliac disease, and refractory sprue. Gut 2001, 49, 804–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cellier, C.; Delabesse, E.; Helmer, C.; Patey, N.; Matuchansky, C.; Jabri, B.; Macintyre, E.; Cerf-Bensussan, N.; Brousse, N. Refractory sprue, coeliac disease, and enteropathy-associated T-cell lymphoma. French Coeliac Disease Study Group. Lancet 2000, 356, 203–208. [Google Scholar] [CrossRef]
- Cellier, C.; Patey, N.; Mauvieux, L.; Jabri, B.; Delabesse, E.; Cervoni, J.P.; Burtin, M.L.; Guy-Grand, D.; Bouhnik, Y.; Modigliani, R.; et al. Abnormal intestinal intraepithelial lymphocytes in refractory sprue. Gastroenterology 1998, 114, 471–481. [Google Scholar] [CrossRef]
- Tjon, J.M.; Verbeek, W.H.; Kooy-Winkelaar, Y.M.; Nguyen, B.H.; van der Slik, A.R.; Thompson, A.; Heemskerk, M.H.; Schreurs, M.W.; Dekking, L.H.; Mulder, C.J.; et al. Defective synthesis or association of T-cell receptor chains underlies loss of surface T-cell receptor-CD3 expression in enteropathy-associated T-cell lymphoma. Blood 2008, 112, 5103–5110. [Google Scholar] [CrossRef] [Green Version]
- Verbeek, W.H.; Goerres, M.S.; von Blomberg, B.M.; Oudejans, J.J.; Scholten, P.E.; Hadithi, M.; Al-Toma, A.; Schreurs, M.W.; Mulder, C.J. Flow cytometric determination of aberrant intra-epithelial lymphocytes predicts T-cell lymphoma development more accurately than T-cell clonality analysis in Refractory Celiac Disease. Clin. Immunol. 2008, 126, 48–56. [Google Scholar] [CrossRef]
- Malamut, G.; Meresse, B.; Cellier, C.; Cerf-Bensussan, N. Refractory celiac disease: From bench to bedside. Semin Immunopathol. 2012, 34, 601–613. [Google Scholar] [CrossRef]
- van Wanrooij, R.L.; Muller, D.M.; Neefjes-Borst, E.A.; Meijer, J.; Koudstaal, L.G.; Heideman, D.A.; Bontkes, H.J.; von Blomberg, B.M.; Bouma, G.; Mulder, C.J. Optimal strategies to identify aberrant intra-epithelial lymphocytes in refractory coeliac disease. J. Clin. Immunol. 2014, 34, 828–835. [Google Scholar] [CrossRef]
- Hussein, S.; Gindin, T.; Lagana, S.M.; Arguelles-Grande, C.; Krishnareddy, S.; Alobeid, B.; Lewis, S.K.; Mansukhani, M.M.; Green, P.H.R.; Bhagat, G. Clonal T cell receptor gene rearrangements in coeliac disease: Implications for diagnosing refractory coeliac disease. J. Clin. Pathol. 2018, 71, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Tack, G.J.; van Wanrooij, R.L.; Langerak, A.W.; Tjon, J.M.; von Blomberg, B.M.; Heideman, D.A.; van Bergen, J.; Koning, F.; Bouma, G.; Mulder, C.J.; et al. Origin and immunophenotype of aberrant IEL in RCDII patients. Mol. Immunol. 2012, 50, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Foss, F.M.; Horwitz, S.M.; Civallero, M.; Bellei, M.; Marcheselli, L.; Kim, W.S.; Cabrera, M.E.; Dlouhy, I.; Nagler, A.; Advani, R.H.; et al. Incidence and outcomes of rare T cell lymphomas from the T Cell Project: Hepatosplenic, enteropathy associated and peripheral gamma delta T cell lymphomas. Am. J. Hematol. 2020, 95, 151–155. [Google Scholar] [CrossRef]
- Verbeek, W.; van de water, J.; Al-Toma, A.; Oudejans, J.; Mulder, C.; Coupe, V. Incidence of enteropathy—Associated T-cell lymphoma: A nation-wide study of a population-based registry in The Netherlands. Scand. J. Gastroenterol. 2008, 43, 1322–1328. [Google Scholar] [CrossRef] [PubMed]
- Sharaiha, R.Z.; Lebwohl, B.; Reimers, L.; Bhagat, G.; Green, P.H.; Neugut, A.I. Increasing incidence of enteropathy-associated T-cell lymphoma in the United States, 1973–2008. Cancer 2012, 118, 3786–3792. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.E.; Song, Y.; Kim, S.J.; Yoon, D.H.; Chen, T.Y.; Koh, Y.; Kang, K.W.; Lee, H.S.; Tay, K.K.W.; Lim, S.T.; et al. Comprehensive analysis of peripheral T-cell and natural killer/T-cell lymphoma in Asian patients: A multinational, multicenter, prospective registry study in Asia. Lancet Reg. Health West. Pac. 2021, 10, 100126. [Google Scholar] [CrossRef]
- Delabie, J.; Holte, H.; Vose, J.M.; Ullrich, F.; Jaffe, E.S.; Savage, K.J.; Connors, J.M.; Rimsza, L.; Harris, N.L.; Muller-Hermelink, K.; et al. Enteropathy-associated T-cell lymphoma: Clinical and histological findings from the international peripheral T-cell lymphoma project. Blood 2011, 118, 148–155. [Google Scholar] [CrossRef] [Green Version]
- Bhagat, G.; Chott, A. Enteropathy-associated T-cell lymphoma. In Digestive System Tumours; WHO Classification of Tumours Editorial Board, Ed.; International Agency for Research on Cancer: Lyon, France, 2019; pp. 386–389. [Google Scholar]
- Jaffe, E.S.; Chott, A.; Ott, G.; Chan, J.K.C.; Bhagat, G.; Tan, S.Y.; Stein, H.; Isaacson, P.G. Intestinal T-cell lymphoma. In WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues; Swerdlow, S.H., Campo, E., Harris, N.L., Jaffe, E.S., Pileri, S.A., Stein, H., Thiele, J., Eds.; International Agency for Research on Cancer: Lyon, France, 2017; pp. 372–380. [Google Scholar]
- Howell, W.M.; Leung, S.T.; Jones, D.B.; Nakshabendi, I.; Hall, M.A.; Lanchbury, J.S.; Ciclitira, P.J.; Wright, D.H. HLA-DRB, -DQA, and -DQB polymorphism in celiac disease and enteropathy-associated T-cell lymphoma. Common features and additional risk factors for malignancy. Hum. Immunol. 1995, 43, 29–37. [Google Scholar] [CrossRef]
- Malamut, G.; Chandesris, O.; Verkarre, V.; Meresse, B.; Callens, C.; Macintyre, E.; Bouhnik, Y.; Gornet, J.M.; Allez, M.; Jian, R.; et al. Enteropathy associated T cell lymphoma in celiac disease: A large retrospective study. Dig. Liver Dis. 2013, 45, 377–384. [Google Scholar] [CrossRef]
- Sieniawski, M.; Angamuthu, N.; Boyd, K.; Chasty, R.; Davies, J.; Forsyth, P.; Jack, F.; Lyons, S.; Mounter, P.; Revell, P.; et al. Evaluation of enteropathy-associated T-cell lymphoma comparing standard therapies with a novel regimen including autologous stem cell transplantation. Blood 2010, 115, 3664–3670. [Google Scholar] [CrossRef] [Green Version]
- Isaacson, P.G.; Du, M.Q. Gastrointestinal lymphoma: Where morphology meets molecular biology. J. Pathol. 2005, 205, 255–274. [Google Scholar] [CrossRef] [PubMed]
- Di Sabatino, A.; Biagi, F.; Gobbi, P.G.; Corazza, G.R. How I treat enteropathy-associated T-cell lymphoma. Blood 2012, 119, 2458–2468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, M.F.; Nishimura, Y.; Nishikori, A.; Yoshino, T.; Sato, Y. Primary Gastrointestinal T-Cell Lymphoma and Indolent Lymphoproliferative Disorders: Practical Diagnostic and Treatment Approaches. Cancers 2021, 13, 5774. [Google Scholar] [CrossRef] [PubMed]
- van Wanrooij, R.L.; de Jong, D.; Langerak, A.W.; Ylstra, B.; van Essen, H.F.; Heideman, D.A.; Bontkes, H.J.; Mulder, C.J.; Bouma, G. Novel variant of EATL evolving from mucosal gammadelta-T-cells in a patient with type I RCD. BMJ Open Gastroenterol. 2015, 2, e000026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ting, Y.T.; Dahal-Koirala, S.; Kim, H.S.K.; Qiao, S.W.; Neumann, R.S.; Lundin, K.E.A.; Petersen, J.; Reid, H.H.; Sollid, L.M.; Rossjohn, J. A molecular basis for the T cell response in HLA-DQ2.2 mediated celiac disease. Proc. Natl. Acad. Sci. USA 2020, 117, 3063–3073. [Google Scholar] [CrossRef]
- Vader, W.; Stepniak, D.; Kooy, Y.; Mearin, L.; Thompson, A.; van Rood, J.J.; Spaenij, L.; Koning, F. The HLA-DQ2 gene dose effect in celiac disease is directly related to the magnitude and breadth of gluten-specific T cell responses. Proc. Natl. Acad. Sci. USA 2003, 100, 12390–12395. [Google Scholar] [CrossRef] [Green Version]
- Al-Toma, A.; Goerres, M.S.; Meijer, J.W.; Pena, A.S.; Crusius, J.B.; Mulder, C.J. Human leukocyte antigen-DQ2 homozygosity and the development of refractory celiac disease and enteropathy-associated T-cell lymphoma. Clin. Gastroenterol. Hepatol. 2006, 4, 315–319. [Google Scholar] [CrossRef]
- Costes, L.M.M.; Lindenbergh-Kortleve, D.J.; van Berkel, L.A.; Veenbergen, S.; Raatgeep, H.R.C.; Simons-Oosterhuis, Y.; van Haaften, D.H.; Karrich, J.J.; Escher, J.C.; Groeneweg, M.; et al. IL-10 signaling prevents gluten-dependent intraepithelial CD4(+) cytotoxic T lymphocyte infiltration and epithelial damage in the small intestine. Mucosal Immunol. 2019, 12, 479–490. [Google Scholar] [CrossRef]
- Maiuri, L.; Troncone, R.; Mayer, M.; Coletta, S.; Picarelli, A.; De Vincenzi, M.; Pavone, V.; Auricchio, S. In vitro activities of A-gliadin-related synthetic peptides: Damaging effect on the atrophic coeliac mucosa and activation of mucosal immune response in the treated coeliac mucosa. Scand. J. Gastroenterol. 1996, 31, 247–253. [Google Scholar] [CrossRef]
- Meresse, B.; Chen, Z.; Ciszewski, C.; Tretiakova, M.; Bhagat, G.; Krausz, T.N.; Raulet, D.H.; Lanier, L.L.; Groh, V.; Spies, T.; et al. Coordinated induction by IL15 of a TCR-independent NKG2D signaling pathway converts CTL into lymphokine-activated killer cells in celiac disease. Immunity 2004, 21, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Di Sabatino, A.; Ciccocioppo, R.; Cupelli, F.; Cinque, B.; Millimaggi, D.; Clarkson, M.M.; Paulli, M.; Cifone, M.G.; Corazza, G.R. Epithelium derived interleukin 15 regulates intraepithelial lymphocyte Th1 cytokine production, cytotoxicity, and survival in coeliac disease. Gut 2006, 55, 469–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, R.; Spolski, R.; Finkelstein, S.E.; Oh, S.; Kovanen, P.E.; Hinrichs, C.S.; Pise-Masison, C.A.; Radonovich, M.F.; Brady, J.N.; Restifo, N.P.; et al. Synergy of IL-21 and IL-15 in regulating CD8+ T cell expansion and function. J. Exp. Med. 2005, 201, 139–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mention, J.J.; Ben Ahmed, M.; Begue, B.; Barbe, U.; Verkarre, V.; Asnafi, V.; Colombel, J.F.; Cugnenc, P.H.; Ruemmele, F.M.; McIntyre, E.; et al. Interleukin 15: A key to disrupted intraepithelial lymphocyte homeostasis and lymphomagenesis in celiac disease. Gastroenterology 2003, 125, 730–745. [Google Scholar] [CrossRef]
- Sarra, M.; Cupi, M.L.; Monteleone, I.; Franze, E.; Ronchetti, G.; Di Sabatino, A.; Gentileschi, P.; Franceschilli, L.; Sileri, P.; Sica, G.; et al. IL-15 positively regulates IL-21 production in celiac disease mucosa. Mucosal Immunol. 2013, 6, 244–255. [Google Scholar] [CrossRef]
- Ebert, E.C. Interleukin 21 up-regulates perforin-mediated cytotoxic activity of human intra-epithelial lymphocytes. Immunology 2009, 127, 206–215. [Google Scholar] [CrossRef] [Green Version]
- Parrish-Novak, J.; Dillon, S.R.; Nelson, A.; Hammond, A.; Sprecher, C.; Gross, J.A.; Johnston, J.; Madden, K.; Xu, W.; West, J.; et al. Interleukin 21 and its receptor are involved in NK-cell expansion and regulation of lymphocyte function. Nature 2000, 408, 57–63. [Google Scholar] [CrossRef]
- Malamut, G.; El Machhour, R.; Montcuquet, N.; Martin-Lanneree, S.; Dusanter-Fourt, I.; Verkarre, V.; Mention, J.J.; Rahmi, G.; Kiyono, H.; Butz, E.A.; et al. IL-15 triggers an antiapoptotic pathway in human intraepithelial lymphocytes that is a potential new target in celiac disease-associated inflammation and lymphomagenesis. J. Clin. Investig. 2010, 120, 2131–2143. [Google Scholar] [CrossRef] [Green Version]
- Chander, U.; Leeman-Neill, R.J.; Bhagat, G. Pathogenesis of Enteropathy-Associated T Cell Lymphoma. Curr. Hematol. Malig. Rep. 2018, 13, 308–317. [Google Scholar] [CrossRef]
- Deleeuw, R.J.; Zettl, A.; Klinker, E.; Haralambieva, E.; Trottier, M.; Chari, R.; Ge, Y.; Gascoyne, R.D.; Chott, A.; Muller-Hermelink, H.K.; et al. Whole-genome analysis and HLA genotyping of enteropathy-type T-cell lymphoma reveals 2 distinct lymphoma subtypes. Gastroenterology 2007, 132, 1902–1911. [Google Scholar] [CrossRef]
- Ondrejka, S.L.; Moffitt, A.B.; Tse, E.; Hsi, E.D.; Goodlad, J.R.; au-Yeung, R.; Kwong, Y.-L.; Srivastava, G.; Gascoyne, R.D.; Rajagopalan, D.; et al. Whole Exome Sequencing of Type 1 and Type 2 Enteropathy-Associated T Cell Lymphoma Reveals Genetic Basis of Eatl Oncogenesis. Blood 2015, 126, 575. [Google Scholar] [CrossRef]
- Cording, S.; Lhermitte, L.; Malamut, G.; Berrabah, S.; Trinquand, A.; Guegan, N.; Villarese, P.; Kaltenbach, S.; Meresse, B.; Khater, S.; et al. Oncogenetic landscape of lymphomagenesis in coeliac disease. Gut 2022, 71, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Moffitt, A.B.; Ondrejka, S.L.; McKinney, M.; Rempel, R.E.; Goodlad, J.R.; Teh, C.H.; Leppa, S.; Mannisto, S.; Kovanen, P.E.; Tse, E.; et al. Enteropathy-associated T cell lymphoma subtypes are characterized by loss of function of SETD2. J. Exp. Med. 2017, 214, 1371–1386. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, M.; Nakamura, S.; Kikuma, K.; Nakayama, Y.; Nimura, S.; Yao, T.; Urabe, S.; Ogawara, S.; Yonemasu, H.; Matsushita, Y.; et al. Pathological and immunohistological findings and genetic aberrations of intestinal enteropathy-associated T cell lymphoma in Japan. Histopathology 2011, 58, 395–407. [Google Scholar] [CrossRef]
- Ko, Y.H.; Karnan, S.; Kim, K.M.; Park, C.K.; Kang, E.S.; Kim, Y.H.; Kang, W.K.; Kim, S.J.; Kim, W.S.; Lee, W.Y.; et al. Enteropathy-associated T-cell lymphoma--a clinicopathologic and array comparative genomic hybridization study. Hum. Pathol. 2010, 41, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.K.; Chan, A.C.; Cheuk, W.; Wan, S.K.; Lee, W.K.; Lui, Y.H.; Chan, W.K. Type II enteropathy-associated T-cell lymphoma: A distinct aggressive lymphoma with frequent gammadelta T-cell receptor expression. Am. J. Surg. Pathol. 2011, 35, 1557–1569. [Google Scholar] [CrossRef]
- Bhagat, G.; Tan, S.Y. Intestinal T-cell lymphoma NOS. In Digestive System Tumours; WHO Classification of Tumours Editorial Board, Ed.; International Agency for Research on Cancer: Lyon, France, 2019; pp. 393–394. [Google Scholar]
- Tan, S.Y.; de Leval, L. Monomorphic epitheliotropic intestinal T-cell lymphoma. In Digestive System Tumours; WHO Classification of Tumours Editorial Board, Ed.; International Agency for Research on Cancer: Lyon, France, 2019; pp. 390–392. [Google Scholar]
- Cavalieri, D.; Tournilhac, O.; Missiglia, E.; Bonnet, C.; Ledoux-Pilon, A.; Bisig, B.; Cairoli, A.; Poullot, E.; Fataccioli, V.; Parrens, M.; et al. Momomorphic epitheliotropic intestinal T-cell lymphoma (MEITL): Clinico-pathological analysis of a multicenter European cohort. Hematol. Oncol. 2021, 39. [Google Scholar] [CrossRef]
- Haddad, P.A.; Dadi, N. Clinicopathologic Determinants of Survival in Monomorphic Epitheliotropic Intestinal T-Cell Lymphoma (MEITL): Analysis of a Pooled Database. Blood 2020, 136, 28. [Google Scholar] [CrossRef]
- Tan, S.Y.; Chuang, S.S.; Tang, T.; Tan, L.; Ko, Y.H.; Chuah, K.L.; Ng, S.B.; Chng, W.J.; Gatter, K.; Loong, F.; et al. Type II EATL (epitheliotropic intestinal T-cell lymphoma): A neoplasm of intra-epithelial T-cells with predominant CD8αα phenotype. Leukemia 2013, 27, 1688–1696. [Google Scholar] [CrossRef] [Green Version]
- Tse, E.; Gill, H.; Loong, F.; Kim, S.J.; Ng, S.B.; Tang, T.; Ko, Y.H.; Chng, W.J.; Lim, S.T.; Kim, W.S.; et al. Type II enteropathy-associated T-cell lymphoma: A multicenter analysis from the Asia Lymphoma Study Group. Am. J. Hematol. 2012, 87, 663–668. [Google Scholar] [CrossRef]
- Garcia-Herrera, A.; Song, J.Y.; Chuang, S.S.; Villamor, N.; Colomo, L.; Pittaluga, S.; Alvaro, T.; Rozman, M.; de Anda Gonzalez, J.; Arrunategui, A.M.; et al. Nonhepatosplenic gammadelta T-cell lymphomas represent a spectrum of aggressive cytotoxic T-cell lymphomas with a mainly extranodal presentation. Am. J. Surg. Pathol. 2011, 35, 1214–1225. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, H.; Nimura, S.; Kayashima, Y.; Takamatsu, Y.; Aoyagi, K.; Harada, N.; Kadowaki, M.; Kamio, T.; Sakisaka, S.; Takeshita, M. Multiple lesions of gastrointestinal tract invasion by monomorphic epitheliotropic intestinal T-cell lymphoma, accompanied by duodenal and intestinal enteropathy-like lesions and microscopic lymphocytic proctocolitis: A case series. Diagn. Pathol. 2016, 11, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.; Zhou, G.; Chen, M.; Liu, W.; Zhao, S. Monomorphic Epitheliotropic Intestinal T-cell Lymphoma of the Stomach: Two Case Reports and a Literature Review. Int. J. Surg. Pathol. 2021, 29, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Antoniadou, F.; Dimitrakopoulou, A.; Voutsinas, P.M.; Vrettou, K.; Vlahadami, I.; Voulgarelis, M.; Korkolopoulou, P.; Kafasi, N.; Mikou, P. Monomorphic epitheliotropic intestinal T-cell lymphoma in pleural effusion: A case report. Diagn. Cytopathol. 2017, 45, 1050–1054. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.S.Y.; Lee, E.; Khong, P.-L.; Tse, E.W.C.; Kwong, Y.-L. Positron emission tomography computed tomography features of monomorphic epitheliotropic intestinal T-cell lymphoma. Hematology 2018, 23, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Tan, S.-Y.; Petersson, B.F.; Khor, Y.M.; Gopalakrishnan, S.K.; Tan, D. Occult recurrence of monomorphic epitheliotropic intestinal T-cell lymphoma and the role of MATK gene expression in diagnosis. Hematol. Oncol. 2017, 35, 852–855. [Google Scholar] [CrossRef]
- Morimoto, A.; Fujioka, Y.; Ushiku, T.; Kurokawa, M. Monomorphic Epitheliotropic Intestinal T-cell Lymphoma Invades Brain. Intern. Med. 2021, 60, 815–816. [Google Scholar] [CrossRef]
- Ritter, J.P.; Flores, R.; Nazarullah, A. Monomorphic Epitheliotropic Intestinal T-Cell Lymphoma (MEITL) Presenting As An Obstructive Pancreaticoduodenal Mass: A Case Report. Am. J. Clin. Pathol. 2020, 154, S104. [Google Scholar] [CrossRef]
- Suzuki, Y.; Minemura, H.; Tomita, H.; Saito, M.; Watanabe, N.; Umeda, T.; Kawamata, T.; Rikimaru, M.; Morimoto, J.; Koizumi, T.; et al. Monomorphic Epitheliotropic Intestinal T-cell Lymphoma Involving the Lung and Brain: A Rare Case Study. Intern. Med. 2020, 59, 2559–2563. [Google Scholar] [CrossRef]
- Aoki, Y.; Sujino, T.; Takabayashi, K.; Mutakuchi, M.; Emoto, K.; Hosoe, N.; Ogata, H.; Kanai, T. Various Endoscopic Features in Monomorphic Epitheliotropic Intestinal T-Cell Lymphoma. Case Rep. Gastroenterol. 2021, 15, 312–322. [Google Scholar] [CrossRef]
- Hong, Y.S.; Woo, Y.S.; Park, G.; Lee, K.; Kang, S.H.; Lee, H.W.; Kim, E.R.; Hong, S.N.; Chang, D.K.; Kim, Y.H.; et al. Endoscopic Findings of Enteropathy-Associated T-Cell Lymphoma Type II: A Case Series. Gut Liver 2016, 10, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, H.; Nimura, S.; Hirai, F.; Harada, N.; Iwasaki, H.; Kawauchi, S.; Oshiro, Y.; Matsuyama, A.; Nakamura, S.; Takamatsu, Y.; et al. Endoscopic and clinicopathological characteristics of colorectal T/NK-cell lymphoma. Diagn. Pathol. 2020, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Xiao, S.Y.; Chen, Q.; Liu, H.; Ping, J. Monomorphic epitheliotropic intestinal T-cell lymphoma may mimic intestinal inflammatory disorders. Int. J. Immunopathol. Pharm. 2019, 33, 2058738419829387. [Google Scholar] [CrossRef]
- Kitano, Y.; Oura, S.; Mushiake, Y.; Makimoto, S. Monomorphic Epitheliotropic Intestinal T-Cell Lymphoma in the Ileum with Successful Preoperative Endoscopic Evaluation. Case Rep. Oncol. 2021, 14, 1283–1288. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.L.; Swerdlow, S.H.; Przybylski, G.K.; Surti, U.; Choi, J.K.; Campo, E.; Trucco, M.M.; Van Oss, S.B.; Felgar, R.E. Intestinal γδ T-cell lymphomas are most frequently of type II enteropathy-associated T-cell type. Hum. Pathol. 2013, 44, 1131–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutzbauer, G.; Maurus, K.; Buszello, C.; Pischimarov, J.; Roth, S.; Rosenwald, A.; Chott, A.; Geissinger, E. SYK expression in monomorphic epitheliotropic intestinal T-cell lymphoma. Mod. Pathol. 2018, 31, 505–516. [Google Scholar] [CrossRef]
- Tomita, S.; Kikuti, Y.Y.; Carreras, J.; Kojima, M.; Ando, K.; Takasaki, H.; Sakai, R.; Takata, K.; Yoshino, T.; Bea, S.; et al. Genomic and immunohistochemical profiles of enteropathy-associated T-cell lymphoma in Japan. Mod. Pathol. 2015, 28, 1286–1296. [Google Scholar] [CrossRef] [Green Version]
- Tomita, S.; Kikuti, Y.Y.; Carreras, J.; Nakamura, N. Monomorphic epitheliotropic intestinal T-cell lymphoma with T-cell receptor (TCR) of silent phenotype shows rearrangement of TCRβ or TCRγ gene. Pathol. Int. 2019, 69, 117–118. [Google Scholar] [CrossRef]
- Chott, A.; Haedicke, W.; Mosberger, I.; Fodinger, M.; Winkler, K.; Mannhalter, C.; Muller-Hermelink, H.K. Most CD56+ intestinal lymphomas are CD8+CD5-T-cell lymphomas of monomorphic small to medium size histology. Am. J. Pathol. 1998, 153, 1483–1490. [Google Scholar] [CrossRef]
- Tan, S.Y.; Ooi, A.S.; Ang, M.K.; Koh, M.; Wong, J.C.; Dykema, K.; Ngeow, J.; Loong, S.; Gatter, K.; Tan, L.; et al. Nuclear expression of MATK is a novel marker of type II enteropathy-associated T-cell lymphoma. Leukemia 2011, 25, 555–557. [Google Scholar] [CrossRef]
- Nairismagi, M.L.; Tan, J.; Lim, J.Q.; Nagarajan, S.; Ng, C.C.; Rajasegaran, V.; Huang, D.; Lim, W.K.; Laurensia, Y.; Wijaya, G.C.; et al. JAK-STAT and G-protein-coupled receptor signaling pathways are frequently altered in epitheliotropic intestinal T-cell lymphoma. Leukemia 2016, 30, 1311–1319. [Google Scholar] [CrossRef] [Green Version]
- Okumura, K.; Ikebe, M.; Shimokama, T.; Takeshita, M.; Kinjo, N.; Sugimachi, K.; Higashi, H. An unusual enteropathy-associated T-cell lymphoma with MYC translocation arising in a Japanese patient: A case report. World J. Gastroenterol. 2012, 18, 2434–2437. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Gong, Y.; Yang, Y.; Xia, Q.; Rao, Q.; Shao, Y.; Zhu, L.; Zhang, J.; Li, X.; Ji, P.; et al. Clinicopathological and molecular genomic features of monomorphic epitheliotropic intestinal T-cell lymphoma in the Chinese population: A study of 20 cases. Diagn. Pathol. 2021, 16, 114. [Google Scholar] [CrossRef] [PubMed]
- Roberti, A.; Dobay, M.P.; Bisig, B.; Vallois, D.; Boechat, C.; Lanitis, E.; Bouchindhomme, B.; Parrens, M.C.; Bossard, C.; Quintanilla-Martinez, L.; et al. Type II enteropathy-associated T-cell lymphoma features a unique genomic profile with highly recurrent SETD2 alterations. Nat. Commun. 2016, 7, 12602. [Google Scholar] [CrossRef] [PubMed]
- Tomita, S.; Kikuti, Y.Y.; Carreras, J.; Sakai, R.; Takata, K.; Yoshino, T.; Bea, S.; Campo, E.; Missiaglia, E.; Bouilly, J.; et al. Monomorphic Epitheliotropic Intestinal T-Cell Lymphoma in Asia Frequently Shows SETD2 Alterations. Cancers 2020, 12, 3539. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.; Adduri, R.S.; Smyth, P.; Quinn, F.; Jeffers, M.; Dunne, B.; O’Leary, J.; McKiernan, S.; Vandenberghe, E.; Pyne, S.; et al. Potentially important miRNAs in enteropathy-associated T-cell lymphoma pathogenesis: A pilot study. Leuk. Res. Rep. 2018, 10, 52–54. [Google Scholar] [CrossRef]
- Park, S.; Ko, Y.H. Peripheral T cell lymphoma in Asia. Int. J. Hematol. 2014, 99, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Ciccocioppo, R.; Croci, G.A.; Biagi, F.; Vanoli, A.; Alvisi, C.; Cavenaghi, G.; Riboni, R.; Arra, M.; Gobbi, P.G.; Paulli, M.; et al. Intestinal T-cell lymphoma with enteropathy-associated T-cell lymphoma-like features arising in the setting of adult autoimmune enteropathy. Hematol. Oncol. 2018, 36, 481–488. [Google Scholar] [CrossRef]
- Attygalle, A.D.; Cabecadas, J.; Gaulard, P.; Jaffe, E.S.; de Jong, D.; Ko, Y.H.; Said, J.; Klapper, W. Peripheral T-cell and NK-cell lymphomas and their mimics; taking a step forward—report on the lymphoma workshop of the XVIth meeting of the European Association for Haematopathology and the Society for Hematopathology. Histopathology 2014, 64, 171–199. [Google Scholar] [CrossRef]
- Kawamoto, K.; Nakamura, S.; Iwashita, A.; Watanabe, J.; Oshiro, Y.; Nakayama, Y.; Nimura, S.; Kimura, N.; Aoyagi, K.; Yao, T.; et al. Clinicopathological characteristics of primary gastric T-cell lymphoma. Histopathology 2009, 55, 641–653. [Google Scholar] [CrossRef]
- Sun, J.; Lu, Z.; Yang, D.; Chen, J. Primary intestinal T-cell and NK-cell lymphomas: A clinicopathological and molecular study from China focused on type II enteropathy-associated T-cell lymphoma and primary intestinal NK-cell lymphoma. Mod. Pathol. 2011, 24, 983–992. [Google Scholar] [CrossRef] [Green Version]
- Nicolae, A.; Xi, L.; Pham, T.H.; Pham, T.A.; Navarro, W.; Meeker, H.G.; Pittaluga, S.; Jaffe, E.S.; Raffeld, M. Mutations in the JAK/STAT and RAS signaling pathways are common in intestinal T-cell lymphomas. Leukemia 2016, 30, 2245–2247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwong, Y.-L.; Kim, W.S.; Lim, S.T.; Kim, S.J.; Tang, T.; Tse, E.; Leung, A.Y.H.; Chim, C.-S. SMILE for natural killer/T-cell lymphoma: Analysis of safety and efficacy from the Asia Lymphoma Study Group. Blood 2012, 120, 2973–2980. [Google Scholar] [CrossRef] [PubMed]
- Allen, P.B.; Lechowicz, M.J. Management of NK/T-Cell Lymphoma, Nasal Type. J. Oncol. Pract. 2019, 15, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Tse, E.; Kwong, Y.-L. How I treat NK/T-cell lymphomas. Blood 2013, 121, 4997–5005. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Kim, W.S.; Park, Y.H.; Park, S.H.; Park, K.W.; Kang, J.H.; Lee, S.S.; Lee, S.I.; Lee, S.H.; Kim, K.; et al. Nasal-type NK/T cell lymphoma: Clinical features and treatment outcome. Br. J. Cancer 2005, 92, 1226–1230. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, M.; Kwong, Y.-L.; Kim, W.S.; Maeda, Y.; Hashimoto, C.; Suh, C.; Izutsu, K.; Ishida, F.; Isobe, Y.; Sueoka, E.; et al. Phase II Study of SMILE Chemotherapy for Newly Diagnosed Stage IV, Relapsed, or Refractory Extranodal Natural Killer (NK)/T-Cell Lymphoma, Nasal Type: The NK-Cell Tumor Study Group Study. J. Clin. Oncol. 2011, 29, 4410–4416. [Google Scholar] [CrossRef]
- Lee, J.; Au, W.Y.; Park, M.J.; Suzumiya, J.; Nakamura, S.; Kameoka, J.; Sakai, C.; Oshimi, K.; Kwong, Y.L.; Liang, R.; et al. Autologous hematopoietic stem cell transplantation in extranodal natural killer/T cell lymphoma: A multinational, multicenter, matched controlled study. Biol. Blood Marrow Transplant. 2008, 14, 1356–1364. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Cui, Y.; Sun, Z.; Zhang, L.; Li, L.; Wang, X.; Wu, J.; Fu, X.; Ma, W.; Zhang, X.; et al. DDGP versus SMILE in Newly Diagnosed Advanced Natural Killer/T-Cell Lymphoma: A Randomized Controlled, Multicenter, Open-label Study in China. Clin. Cancer Res. 2016, 22, 5223–5228. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.H.; Wang, L.; Liu, C.C.; Xia, Z.J.; Huang, H.Q.; Lin, T.Y.; Jiang, W.Q.; Lu, Y. Efficacy of combined gemcitabine, oxaliplatin and pegaspargase (P-gemox regimen) in patients with newly diagnosed advanced-stage or relapsed/refractory extranodal NK/T-cell lymphoma. Oncotarget 2016, 7, 29092–29101. [Google Scholar] [CrossRef]
- Zhang, L.; Jia, S.; Ma, Y.; Li, L.; Li, X.; Wang, X.; Fu, X.; Ma, W.; Qin, Y.; Li, W.; et al. Efficacy and safety of cisplatin, dexamethasone, gemcitabine and pegaspargase (DDGP) regimen in newly diagnosed, advanced-stage extranodal natural killer/T-cell lymphoma: Interim analysis of a phase 4 study NCT01501149. Oncotarget 2016, 7, 55721–55731. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Fan, S.; Chang, Y.; Liu, X.; Li, W.; Ma, Q.; Li, Y.; Wang, Y.; Zhang, L.; Zhang, M. Clinical efficacy of cisplatin, dexamethasone, gemcitabine and pegaspargase (DDGP) in the initial treatment of advanced stage (stage III-IV) extranodal NK/T-cell lymphoma, and its correlation with Epstein-Barr virus. Cancer Manag. Res. 2019, 11, 3555–3564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Oki, Y. Novel Immunotherapy Options for Extranodal NK/T-Cell Lymphoma. Front. Oncol. 2018, 8, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, K.; Li, X.; Yu, H.; Chen, X.; Zhang, M.; Wu, X. Selection of new immunotherapy targets for NK/T cell lymphoma. Am. J. Transl Res. 2020, 12, 7034–7047. [Google Scholar]
- Aeppli, S.; Driessen, C.; Graf, L.; Hitz, F. Systemic treatment of a patient with relapsed and refractory extranodal NK/T-cell lymphoma (ENKL) and meningeosis leukemica with daratumumab. Hematol. Oncol. 2018, 36, 713–714. [Google Scholar] [CrossRef] [PubMed]
- Hari, P.; Raj, R.V.; Olteanu, H. Targeting CD38 in Refractory Extranodal Natural Killer Cell-T-Cell Lymphoma. N. Engl. J. Med. 2016, 375, 1501–1502. [Google Scholar] [CrossRef]
- Kim, H.K.; Moon, S.M.; Moon, J.H.; Park, J.E.; Byeon, S.; Kim, W.S. Complete remission in CD30-positive refractory extranodal NK/T-cell lymphoma with brentuximab vedotin. Blood Res. 2015, 50, 254–256. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Kim, S.J.; Hong, J.Y.; Yoon, D.H.; Kim, J.S.; Kang, H.J.; Eom, H.-S.; Lee, M.H.; Suh, C.; Kim, W.S. A Phase II Study of Brentuximab Vedotin for Relapsed or Refractory CD30-Positive Non-Hodgkin Lymphomas Other Than Anaplastic Large Cell Lymphoma. Blood 2017, 130, 4077. [Google Scholar]
- Poon, L.M.; Kwong, Y.L. Complete remission of refractory disseminated NK/T cell lymphoma with brentuximab vedotin and bendamustine. Ann. Hematol. 2016, 95, 847–849. [Google Scholar] [CrossRef]
- Huang, H.; Zhu, J.; Yao, M.; Kim, T.M.; Yoon, D.H.; Cho, S.G.; Eom, H.S.; Lim, S.T.; Yeh, S.P.; Song, Y.; et al. Daratumumab monotherapy for patients with relapsed or refractory natural killer/T-cell lymphoma, nasal type: An open-label, single-arm, multicenter, phase 2 study. J. Hematol. Oncol. 2021, 14, 25. [Google Scholar] [CrossRef]
- Chan, T.S.Y.; Li, J.; Loong, F.; Khong, P.L.; Tse, E.; Kwong, Y.L. PD1 blockade with low-dose nivolumab in NK/T cell lymphoma failing L-asparaginase: Efficacy and safety. Ann. Hematol. 2018, 97, 193–196. [Google Scholar] [CrossRef]
- Kwong, Y.L.; Chan, T.S.Y.; Tan, D.; Kim, S.J.; Poon, L.M.; Mow, B.; Khong, P.L.; Loong, F.; Au-Yeung, R.; Iqbal, J.; et al. PD1 blockade with pembrolizumab is highly effective in relapsed or refractory NK/T-cell lymphoma failing l-asparaginase. Blood 2017, 129, 2437–2442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Cheng, Y.; Zhang, M.; Yan, J.; Li, L.; Fu, X.; Zhang, X.; Chang, Y.; Sun, Z.; Yu, H.; et al. Activity of pembrolizumab in relapsed/refractory NK/T-cell lymphoma. J. Hematol. Oncol. 2018, 11, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.J.; Hyeon, J.; Cho, I.; Ko, Y.H.; Kim, W.S. Comparison of Efficacy of Pembrolizumab between Epstein-Barr Virus-Positive and -Negative Relapsed or Refractory Non-Hodgkin Lymphomas. Cancer Res. Treat. 2019, 51, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, J.; Liu, P.; Huang, H.; Li, Y.; Ma, S.; Zhou, H.; Tian, X.; Zhang, Y.; Gao, Y.; Xia, Y.; et al. Combination of anti-PD-1 antibody with P-GEMOX as a potentially effective immunochemotherapy for advanced natural killer/T cell lymphoma. Signal Transduct. Target. 2020, 5, 289. [Google Scholar] [CrossRef] [PubMed]
- Farid, M.; Yau, Y.W.; Tay, K.; Quek, R.; Tao, M.; Koo, G.C.; Loong, S.; Lim, S.T. A promising new regimen for the treatment of advanced extranodal NK/T cell lymphoma. Acta Oncol. 2011, 50, 589–590. [Google Scholar] [CrossRef]
- Chen, C.; He, H. Treatment of relapsed extranodal natural killer/T-cell lymphoma with bortezomib plus fludarabine. Mol. Clin. Oncol. 2017, 7, 525–528. [Google Scholar] [CrossRef] [Green Version]
- Moskowitz, A.J.; Ghione, P.; Jacobsen, E.; Ruan, J.; Schatz, J.H.; Noor, S.; Myskowski, P.; Vardhana, S.; Ganesan, N.; Hancock, H.; et al. A phase 2 biomarker-driven study of ruxolitinib demonstrates effectiveness of JAK/STAT targeting in T-cell lymphomas. Blood 2021, 138, 2828–2837. [Google Scholar] [CrossRef]
- Sieniawski, M.K.; Lennard, A.L. Enteropathy-associated T-cell lymphoma: Epidemiology, clinical features, and current treatment strategies. Curr. Hematol. Malig. Rep. 2011, 6, 231–240. [Google Scholar] [CrossRef]
- Sibon, D.; Khater, S.; Bruneau, J.; Brouzes, C.; Lhermitte, L.; Molina, T.J.; Cartron, G.; Morel, V.; Malamut, G.; Chauchet, A.; et al. The Eatl-001 Trial: Results of a Phase 2 Study of Brentuximab Vedotin and CHP Followed By Consolidation with High-Dose Therapy—Autologous Stem-Cell Transplantation (HDT-ASCT) in the Frontline Treatment of Patients with Enteropathy-Associated T-Cell Lymphoma. Blood 2021, 138, 136. [Google Scholar] [CrossRef]
- Gentille, C.; Qin, Q.; Barbieri, A.; Ravi, P.S.; Iyer, S. Use of PEG-asparaginase in monomorphic epitheliotropic intestinal T-cell lymphoma, a disease with diagnostic and therapeutic challenges. Ecancermedicalscience 2017, 11, 771. [Google Scholar] [CrossRef]
- Vose, J.; Armitage, J.; Weisenburger, D.; International, T.C.L.P. International peripheral T-cell and natural killer/T-cell lymphoma study: Pathology findings and clinical outcomes. J. Clin. Oncol. 2008, 26, 4124–4130. [Google Scholar] [CrossRef] [PubMed]
- Ellin, F.; Landström, J.; Jerkeman, M.; Relander, T. Real-world data on prognostic factors and treatment in peripheral T-cell lymphomas: A study from the Swedish Lymphoma Registry. Blood 2014, 124, 1570–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, N.; Trümper, L.; Ziepert, M.; Nickelsen, M.; Ho, A.D.; Metzner, B.; Peter, N.; Loeffler, M.; Rosenwald, A.; Pfreundschuh, M. Treatment and prognosis of mature T-cell and NK-cell lymphoma: An analysis of patients with T-cell lymphoma treated in studies of the German High-Grade Non-Hodgkin Lymphoma Study Group. Blood 2010, 116, 3418–3425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altmann, B.; Wulf, G.; Truemper, L.; d’Amore, F.; Relander, T.; Toldbod, H.; Delabie, J.M.A.; Rosenwald, A.; Ziepert, M.; Loeffler, M. Alemtuzumab Added to CHOP for Treatment of Peripheral T-Cell Lymphoma (PTCL) in Previously Untreated Young and Elderly Patients: Pooled Analysis of the International ACT-1/2 Phase III Trials. Blood 2018, 132, 1622. [Google Scholar] [CrossRef]
- Gallamini, A.; Zaja, F.; Patti, C.; Billio, A.; Specchia, M.R.; Tucci, A.; Levis, A.; Manna, A.; Secondo, V.; Rigacci, L.; et al. Alemtuzumab (Campath-1H) and CHOP chemotherapy as first-line treatment of peripheral T-cell lymphoma: Results of a GITIL (Gruppo Italiano Terapie Innovative nei Linfomi) prospective multicenter trial. Blood 2007, 110, 2316–2323. [Google Scholar] [CrossRef] [Green Version]
- Wulf, G.G.; Altmann, B.; Ziepert, M.; D’Amore, F.; Held, G.; Greil, R.; Tournilhac, O.; Relander, T.; Viardot, A.; Wilhelm, M.; et al. Alemtuzumab plus CHOP versus CHOP in elderly patients with peripheral T-cell lymphoma: The DSHNHL2006-1B/ACT-2 trial. Leukemia 2021, 35, 143–155. [Google Scholar] [CrossRef]
- Foss, F.M.; Sjak-Shie, N.N.; Goy, A.; Advani, R.; Jacobsen, E.D. Phase II study of denileukin diftitox with CHOP chemotherapy in newly-diagnosed PTCL: CONCEPT trial. J. Clin. Oncol. 2010, 28, 8045. [Google Scholar] [CrossRef]
- Kim, S.J.; Yoon, D.H.; Kang, H.J.; Kim, J.S.; Park, S.K.; Kim, H.J.; Lee, J.; Ryoo, B.Y.; Ko, Y.H.; Huh, J.; et al. Bortezomib in combination with CHOP as first-line treatment for patients with stage III/IV peripheral T-cell lymphomas: A multicentre, single-arm, phase 2 trial. Eur. J. Cancer 2012, 48, 3223–3231. [Google Scholar] [CrossRef]
- Yang, H.; Li, C. The Combination of Bortezomib with Cyclophosphamide, Epirubicin, Etoposide and Prednisone (BCHEP) Regimen As First-Line Treatment for Untreated Patients with Peripheral T Cell Lymphoma: A Prospective, Single Arm, Phase II Study. Blood 2021, 138, 2482. [Google Scholar] [CrossRef]
- Escalón, M.P.; Liu, N.S.; Yang, Y.; Hess, M.; Walker, P.L.; Smith, T.L.; Dang, N.H. Prognostic factors and treatment of patients with T-cell non-Hodgkin lymphoma: The M. D. Anderson Cancer Center experience. Cancer 2005, 103, 2091–2098. [Google Scholar] [CrossRef]
- Gisselbrecht, C.; Gaulard, P.; Lepage, E.; Coiffier, B.; Brière, J.; Haioun, C.; Cazals-Hatem, D.; Bosly, A.; Xerri, L.; Tilly, H.; et al. Prognostic significance of T-cell phenotype in aggressive non-Hodgkin’s lymphomas. Groupe d’Etudes des Lymphomes de l’Adulte (GELA). Blood 1998, 92, 76–82. [Google Scholar] [PubMed]
- Xu, Y.; Wu, X.J.; Wang, Y.; Jin, Z.M.; Sun, A.N.; Wu, D.P. Hyper-CVAD chemotherapy or autologous stem cell transplantation in patients with peripheral T cell lymphomas: A single centre report. Chin. Med. J. 2012, 125, 4134–4137. [Google Scholar] [CrossRef] [PubMed]
- Corazzelli, G.; Frigeri, F.; Marcacci, G.; Becchimanzi, C.; Capobianco, G.; Arcamone, M.; Morelli, E.; Volzone, F.; Russo, F.; Pinto, A. Gemcitabine, Ifosfamide, Oxaliplatin (GIFOX) as First-Line Treatment In High-Risk Peripheral T-Cell/NK Lymphomas: A Phase II Trial. Blood 2010, 116, 2829. [Google Scholar] [CrossRef]
- Dong, M.; He, X.H.; Liu, P.; Qin, Y.; Yang, J.L.; Zhou, S.Y.; Yang, S.; Zhang, C.G.; Gui, L.; Zhou, L.Q.; et al. Gemcitabine-based combination regimen in patients with peripheral T-cell lymphoma. Med. Oncol. 2013, 30, 351. [Google Scholar] [CrossRef] [PubMed]
- Evens, A.M.; Rosen, S.T.; Helenowski, I.; Kline, J.; Larsen, A.; Colvin, J.; Winter, J.N.; van Besien, K.M.; Gordon, L.I.; Smith, S.M. A phase I/II trial of bortezomib combined concurrently with gemcitabine for relapsed or refractory DLBCL and peripheral T-cell lymphomas. Br. J. Haematol. 2013, 163, 55–61. [Google Scholar] [CrossRef]
- Yhim, H.Y.; Kim, T.; Kim, S.J.; Shin, H.J.; Koh, Y.; Kim, J.S.; Park, J.; Park, G.S.; Kim, W.S.; Moon, J.H.; et al. Combination treatment of copanlisib and gemcitabine in relapsed/refractory PTCL (COSMOS): An open-label phase I/II trial. Ann. Oncol. 2021, 32, 552–559. [Google Scholar] [CrossRef]
- Zinzani, P.L.; Venturini, F.; Stefoni, V.; Fina, M.; Pellegrini, C.; Derenzini, E.; Gandolfi, L.; Broccoli, A.; Argnani, L.; Quirini, F.; et al. Gemcitabine as single agent in pretreated T-cell lymphoma patients: Evaluation of the long-term outcome. Ann. Oncol. 2010, 21, 860–863. [Google Scholar] [CrossRef]
- Hong, J.Y.; Yoon, D.H.; Yoon, S.E.; Kim, S.J.; Lee, H.S.; Eom, H.-S.; Lee, H.W.; Shin, D.-Y.; Koh, Y.; Yoon, S.-S.; et al. Pralatrexate in patients with recurrent or refractory peripheral T-cell lymphomas: A multicenter retrospective analysis. Sci. Rep. 2019, 9, 20302. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, O.A.; Pro, B.; Pinter-Brown, L.; Bartlett, N.; Popplewell, L.; Coiffier, B.; Lechowicz, M.J.; Savage, K.J.; Shustov, A.R.; Gisselbrecht, C.; et al. Pralatrexate in patients with relapsed or refractory peripheral T-cell lymphoma: Results from the pivotal PROPEL study. J. Clin. Oncol. 2011, 29, 1182–1189. [Google Scholar] [CrossRef]
- Coiffier, B.; Pro, B.; Prince, H.M.; Foss, F.; Sokol, L.; Greenwood, M.; Caballero, D.; Borchmann, P.; Morschhauser, F.; Wilhelm, M.; et al. Results from a pivotal, open-label, phase II study of romidepsin in relapsed or refractory peripheral T-cell lymphoma after prior systemic therapy. J. Clin. Oncol. 2012, 30, 631–636. [Google Scholar] [CrossRef]
- Coiffier, B.; Pro, B.; Prince, H.M.; Foss, F.; Sokol, L.; Greenwood, M.; Caballero, D.; Morschhauser, F.; Wilhelm, M.; Pinter-Brown, L.; et al. Romidepsin for the treatment of relapsed/refractory peripheral T-cell lymphoma: Pivotal study update demonstrates durable responses. J. Hematol. Oncol. 2014, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Piekarz, R.L.; Frye, R.; Prince, H.M.; Kirschbaum, M.H.; Zain, J.; Allen, S.L.; Jaffe, E.S.; Ling, A.; Turner, M.; Peer, C.J.; et al. Phase 2 trial of romidepsin in patients with peripheral T-cell lymphoma. Blood 2011, 117, 5827–5834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morschhauser, F.; Fitoussi, O.; Haioun, C.; Thieblemont, C.; Quach, H.; Delarue, R.; Glaisner, S.; Gabarre, J.; Bosly, A.; Lister, J.; et al. A phase 2, multicentre, single-arm, open-label study to evaluate the safety and efficacy of single-agent lenalidomide (Revlimid) in subjects with relapsed or refractory peripheral T-cell non-Hodgkin lymphoma: The EXPECT trial. Eur. J. Cancer 2013, 49, 2869–2876. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Zain, J.M.; Palmer, B.; Jovanovic, B.; Mi, X.; Swaroop, A.; Winter, J.; Gordon, L.I.; Karmali, R.; Pro, B. Multicenter phase II study of romidepsin plus lenalidomide for patients with previously untreated peripheral T-cell lymphoma (PTCL). J. Clin. Oncol. 2021, 39, 7514. [Google Scholar] [CrossRef]
- Toumishey, E.; Prasad, A.; Dueck, G.; Chua, N.; Finch, D.; Johnston, J.; van der Jagt, R.; Stewart, D.; White, D.; Belch, A.; et al. Final report of a phase 2 clinical trial of lenalidomide monotherapy for patients with T-cell lymphoma. Cancer 2015, 121, 716–723. [Google Scholar] [CrossRef]
- Umakanthan, J.M.; Iqbal, J.; Batlevi, C.L.; Bouska, A.; Smith, L.M.; Shostrom, V.; Nutsch, H.; William, B.M.; Gregory Bociek, R.; Lunning, M.; et al. Phase I/II study of dasatinib and exploratory genomic analysis in relapsed or refractory non-Hodgkin lymphoma. Br. J. Haematol. 2019, 184, 744–752. [Google Scholar] [CrossRef]
- William, B.M.; Hohenstein, M.; Loberiza, F.R., Jr.; Caponetti, G.C.; Bociek, R.G.; Bierman, P.; Armitage, J.O.; Chan, W.-C.; Vose, J.M. Phase I/II Study of Dasatinib In Relapsed or Refractory Non-Hodgkin’s Lymphoma (NHL). Blood 2010, 116, 288. [Google Scholar] [CrossRef]
- Barr, P.M.; Li, H.; Spier, C.; Mahadevan, D.; LeBlanc, M.; Ul Haq, M.; Huber, B.D.; Flowers, C.R.; Wagner-Johnston, N.D.; Horwitz, S.M.; et al. Phase II Intergroup Trial of Alisertib in Relapsed and Refractory Peripheral T-Cell Lymphoma and Transformed Mycosis Fungoides: SWOG 1108. J. Clin. Oncol. 2015, 33, 2399–2404. [Google Scholar] [CrossRef]
- Friedberg, J.W.; Mahadevan, D.; Cebula, E.; Persky, D.; Lossos, I.; Agarwal, A.B.; Jung, J.; Burack, R.; Zhou, X.; Leonard, E.J.; et al. Phase II study of alisertib, a selective Aurora A kinase inhibitor, in relapsed and refractory aggressive B- and T-cell non-Hodgkin lymphomas. J. Clin. Oncol. 2014, 32, 44–50. [Google Scholar] [CrossRef]
- O’Connor, O.A.; Özcan, M.; Jacobsen, E.D.; Roncero, J.M.; Trotman, J.; Demeter, J.; Masszi, T.; Pereira, J.; Ramchandren, R.; Beaven, A.; et al. Randomized Phase III Study of Alisertib or Investigator’s Choice (Selected Single Agent) in Patients With Relapsed or Refractory Peripheral T-Cell Lymphoma. J. Clin. Oncol. 2019, 37, 613–623. [Google Scholar] [CrossRef]
- Quéméner, A.; Maillasson, M.; Arzel, L.; Sicard, B.; Vomiandry, R.; Mortier, E.; Dubreuil, D.; Jacques, Y.; Lebreton, J.; Mathé-Allainmat, M. Discovery of a Small-Molecule Inhibitor of Interleukin 15: Pharmacophore-Based Virtual Screening and Hit Optimization. J. Med. Chem. 2017, 60, 6249–6272. [Google Scholar] [CrossRef] [PubMed]







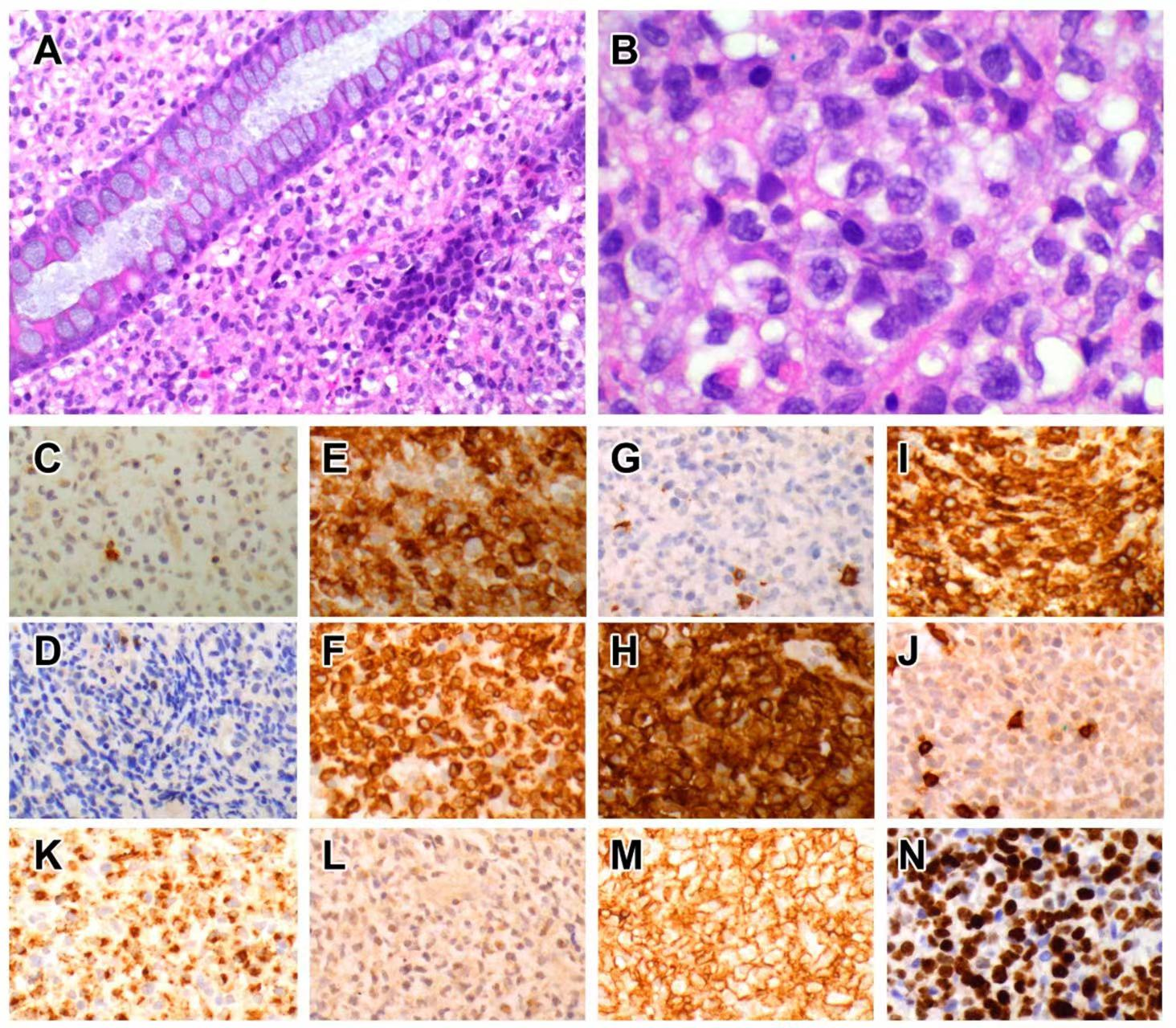{kind=link}
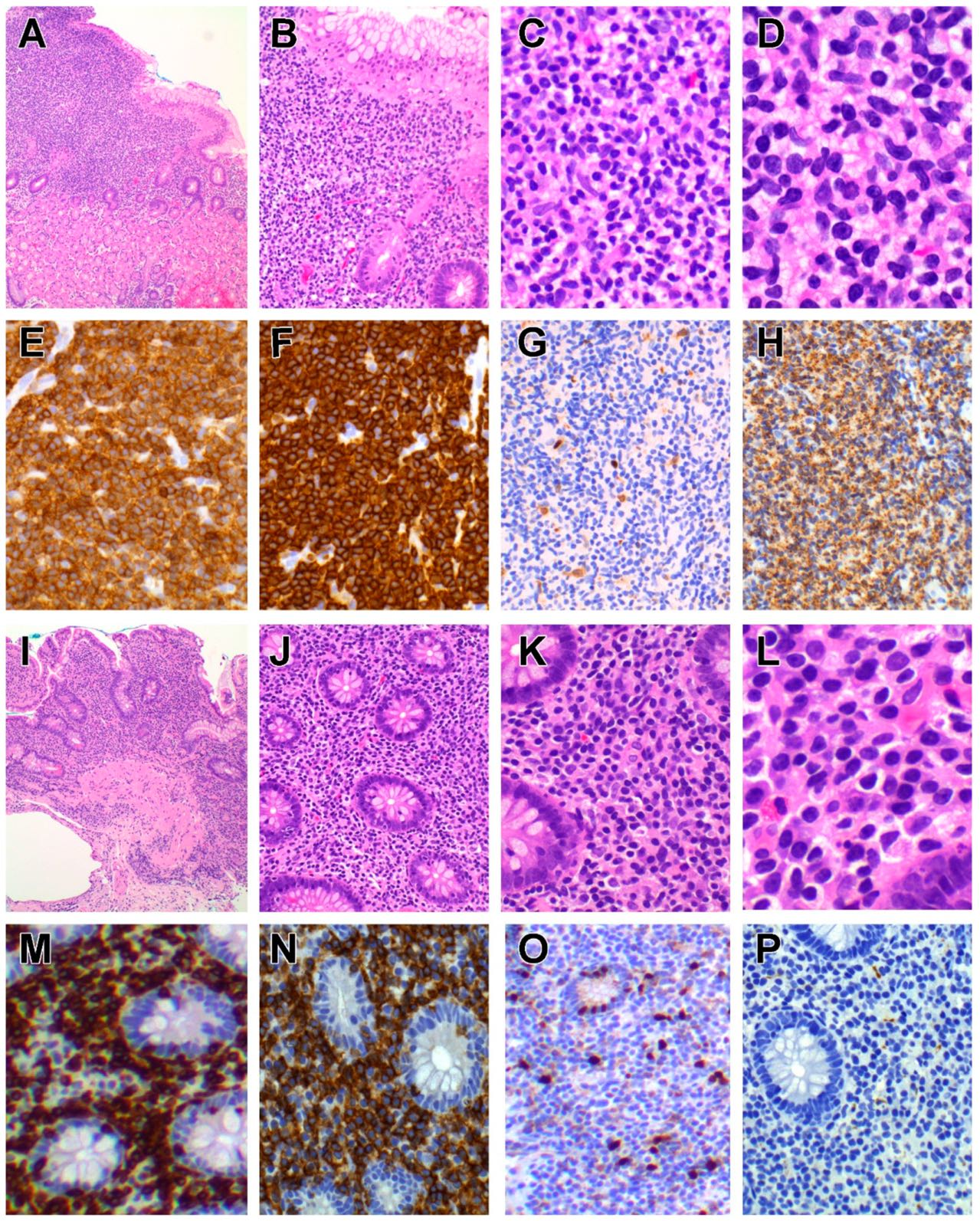{kind=link}
{kind=link}
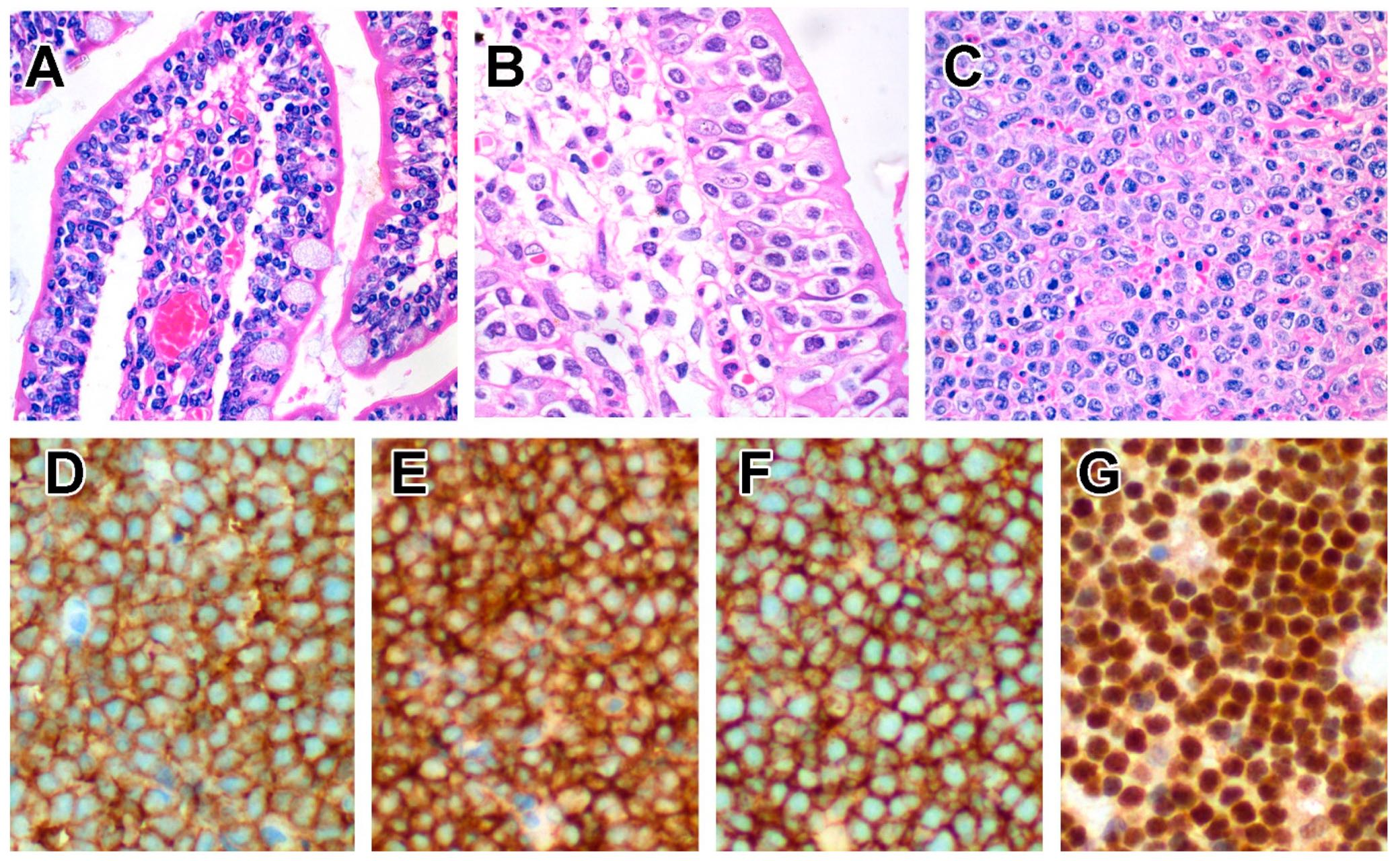{kind=link}
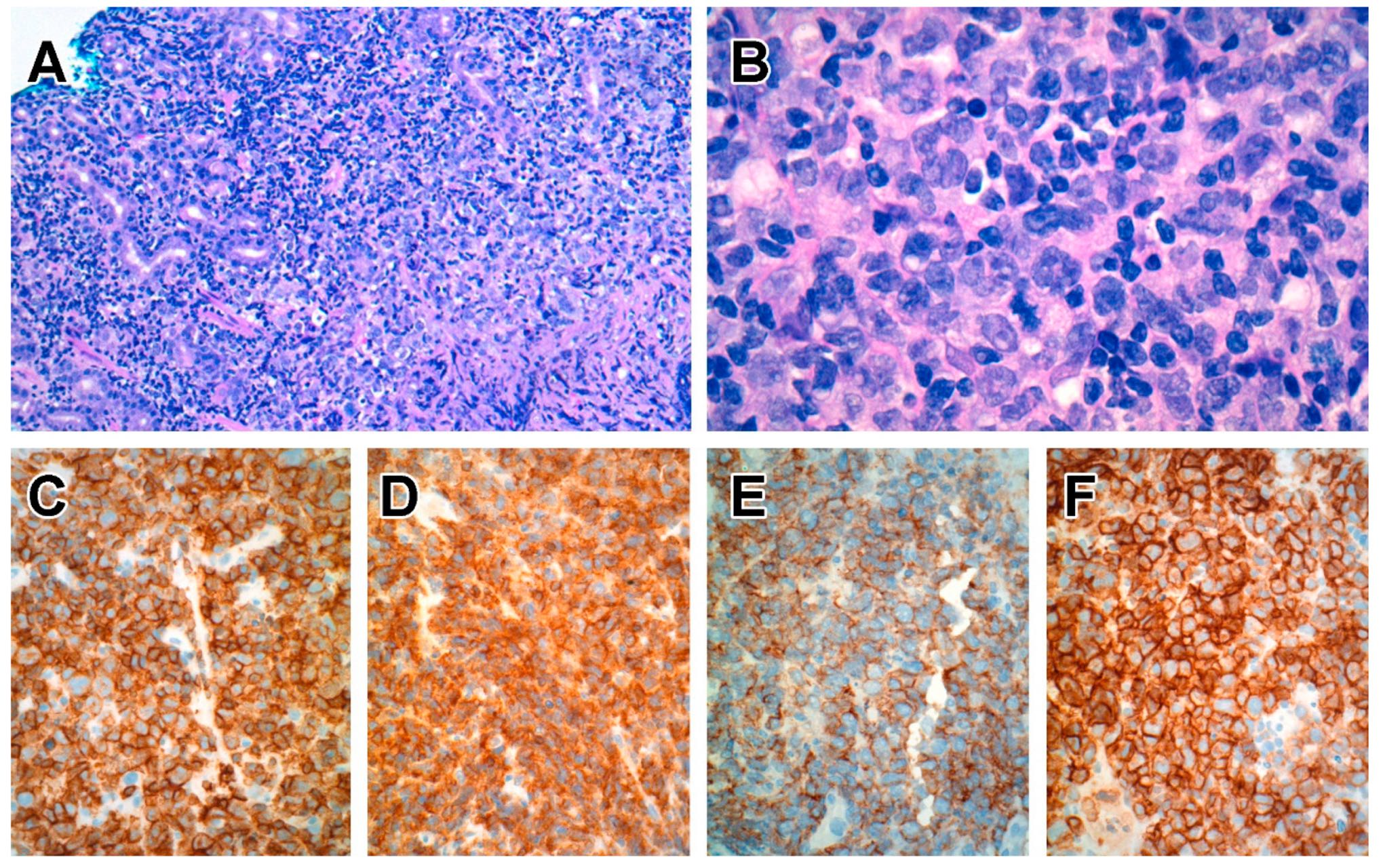{kind=link}
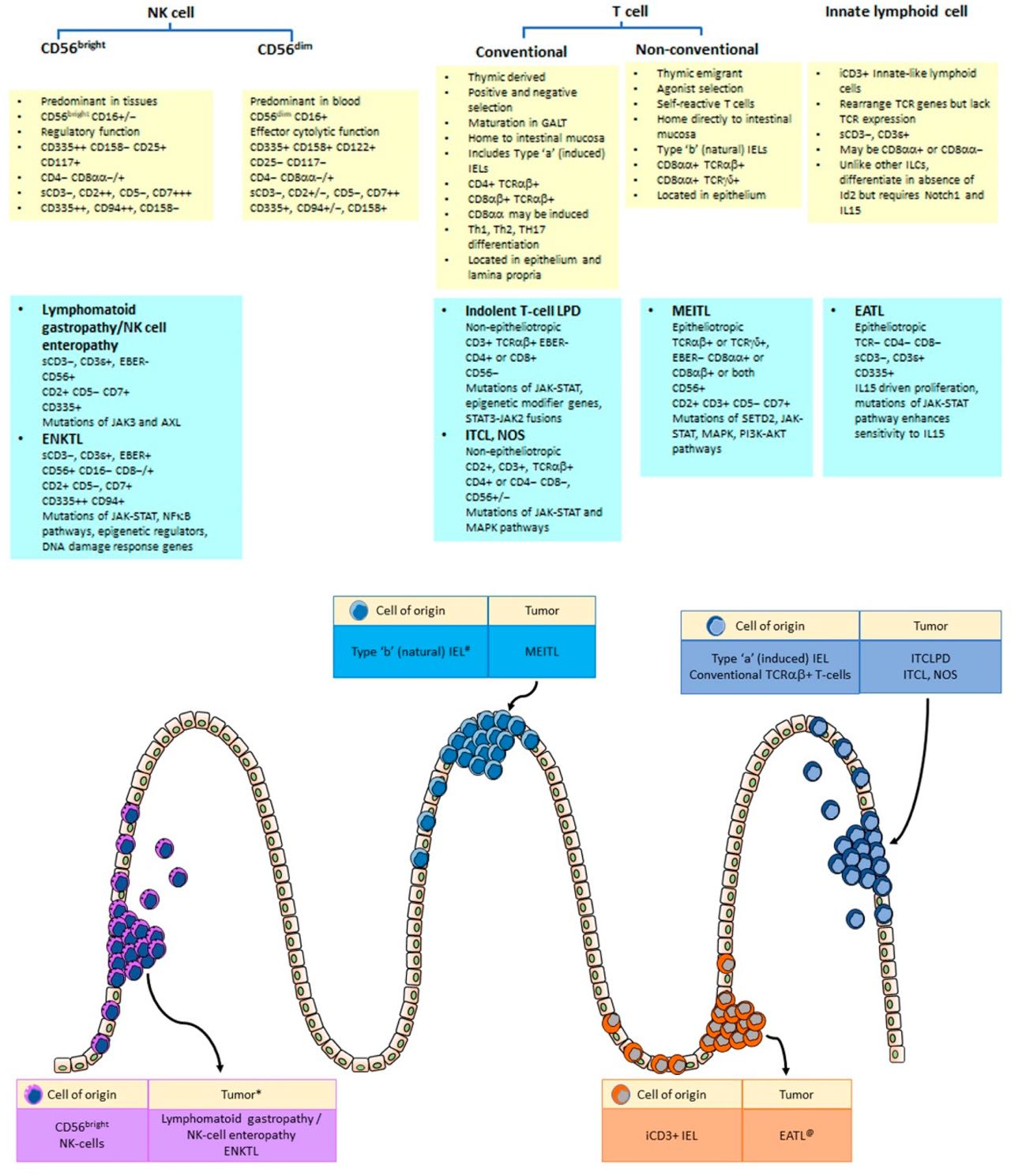{kind=link}
| ILC Subtypes | NK-cell | ILC1 | ILC2 | ILC3 | LTi (Lymphoid Tissue Inducer) |
|---|---|---|---|---|---|
| Cytokine production | TH1 cell-associated cytokines: IFNγ | TH2 cell-associated cytokines: IL5, IL13, IL6, IL9 | TH17 cell-associated cytokines: IL17, IL22 g | TH17 cell-associated cytokines: IL17, IL22: mainly IL17A+ | |
| Phenotype | CD25−/+ a, CD56+, CD117−, CD127−/+, CD161−/+, NKp44−/+ b, NKp46+, ICOS+/− CRTH2−, IL1R−, IL23R−, IL12Rβ2+, ST2−, IL17RB− | CD25+/−, CD56−, CD117−, CD127+ d, CD161+/−, NKp44−, NKp46−, ICOS+, CRTH2−, IL1R+, IL23R−, IL12Rβ2+, ST2−, IL17RB− | CD25+, CD56−, CD117+/−, CD127+, CD161+, NKp44−, NKp46−, ICOS+, CRTH2+, IL1R+, IL23R ND f, IL12Rβ2−, ST2+, IL17RB+ | CD25−/+, CD56+/−, CD117+, CD127+, CD161+ c, NKp44+ c, NKp44− e, NKp46+, ICOS+ c, CRTH2 c, IL1R+ c, IL23R+ c, IL12Rβ2− c, ST2− c, IL17RB− c | CD25+/−, CD56−, CD117+, CD127+, CD161+/−, NKp44−, NKp46−, ICOS ND f, CRTH2−, IL1R+, IL23R+, IL12Rβ2−, STR2, IL17RB− |
| Differentiation of ILCs | T-bet+, Eomes+, RORγt−, GATA3−, AhR− | T-bet+, Eomes−/+, RORγt−, GATA3−, AhR− | T-bet−, Eomes−, RORγt−, RORα+, GATA3+, AhR+ | T-bet−, Eomes−, RORγt+, GATA3−, AhR+ | T-bet−, Eomes−, RORγt+, GATA3−, AhR+ |
| Function | Pro-inflammatory against bacteria and intracellular viruses | Defence against helmintic infections | Maintain homeostasis, immune tolerance, defence against fungal infections | LTi role in formation of secondary lymphoid organs during embryogenesis | |
| CD56bright * | CD56dim * | |
|---|---|---|
| Functions | Regulatory | Effector cytolytic |
| Location | Predominantly tissue | Primary in blood |
| Maturity | Precursor | Mature |
| NK-cell markers | ||
| CD56 | ++ | + |
| CD57 | − | + |
| CD16 | +/− | ++ |
| T cell markers | ||
| CD2 | ++ | +/− |
| CD5 | − | −/+ |
| CD7 | +++ | ++ |
| CD8 | −/+ * | −/+ @ |
| Leucocyte integrins | ||
| CD11c | ++ | +/− |
| Selectins | ||
| CD62L | ++ | +/− |
| Cytokine receptors | ||
| CD122 | −/+ | + |
| CD25 | + | − |
| CD4 | −/+ | − |
| Growth factor receptor | ||
| CD117 | +/− | − |
| Inhibitory and activating receptors | ||
| KIR/CD158 | − | +/− |
| CD94/NKG2A | ++ | +/− |
| ILT2 | − | + |
| Natural cytotoxic receptors | ||
| CD335/NKp46 | ++ | + |
| Activation markers | ||
| HLA-DR | +/− | − |
| CD38 | + | + |
| CD26 | + | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hue, S.S.-S.; Ng, S.-B.; Wang, S.; Tan, S.-Y. Cellular Origins and Pathogenesis of Gastrointestinal NK- and T-Cell Lymphoproliferative Disorders. Cancers 2022, 14, 2483. https://doi.org/10.3390/cancers14102483
Hue SS-S, Ng S-B, Wang S, Tan S-Y. Cellular Origins and Pathogenesis of Gastrointestinal NK- and T-Cell Lymphoproliferative Disorders. Cancers. 2022; 14(10):2483. https://doi.org/10.3390/cancers14102483
Chicago/Turabian StyleHue, Susan Swee-Shan, Siok-Bian Ng, Shi Wang, and Soo-Yong Tan. 2022. "Cellular Origins and Pathogenesis of Gastrointestinal NK- and T-Cell Lymphoproliferative Disorders" Cancers 14, no. 10: 2483. https://doi.org/10.3390/cancers14102483
APA StyleHue, S. S. -S., Ng, S. -B., Wang, S., & Tan, S. -Y. (2022). Cellular Origins and Pathogenesis of Gastrointestinal NK- and T-Cell Lymphoproliferative Disorders. Cancers, 14(10), 2483. https://doi.org/10.3390/cancers14102483