
Potential Prognostic Markers for Relapsed/Refractory vs. Responsive Acute Myeloid Leukemia


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Gene Expression Analysis by RT-qPCR
2.3. Statistical Analysis
3. Results
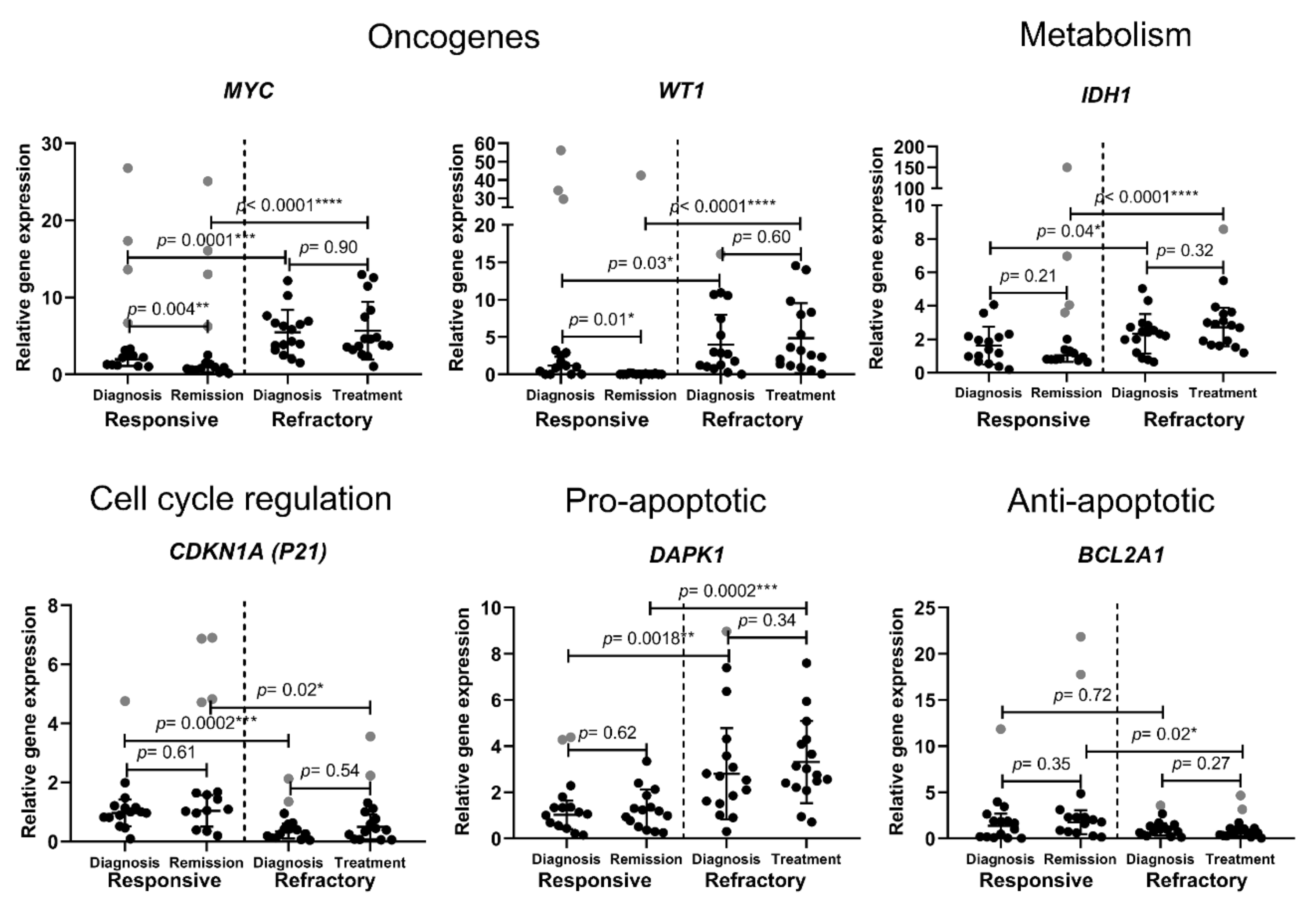
3.1. Cell-Fate-Important Gene Expression in AML Patients
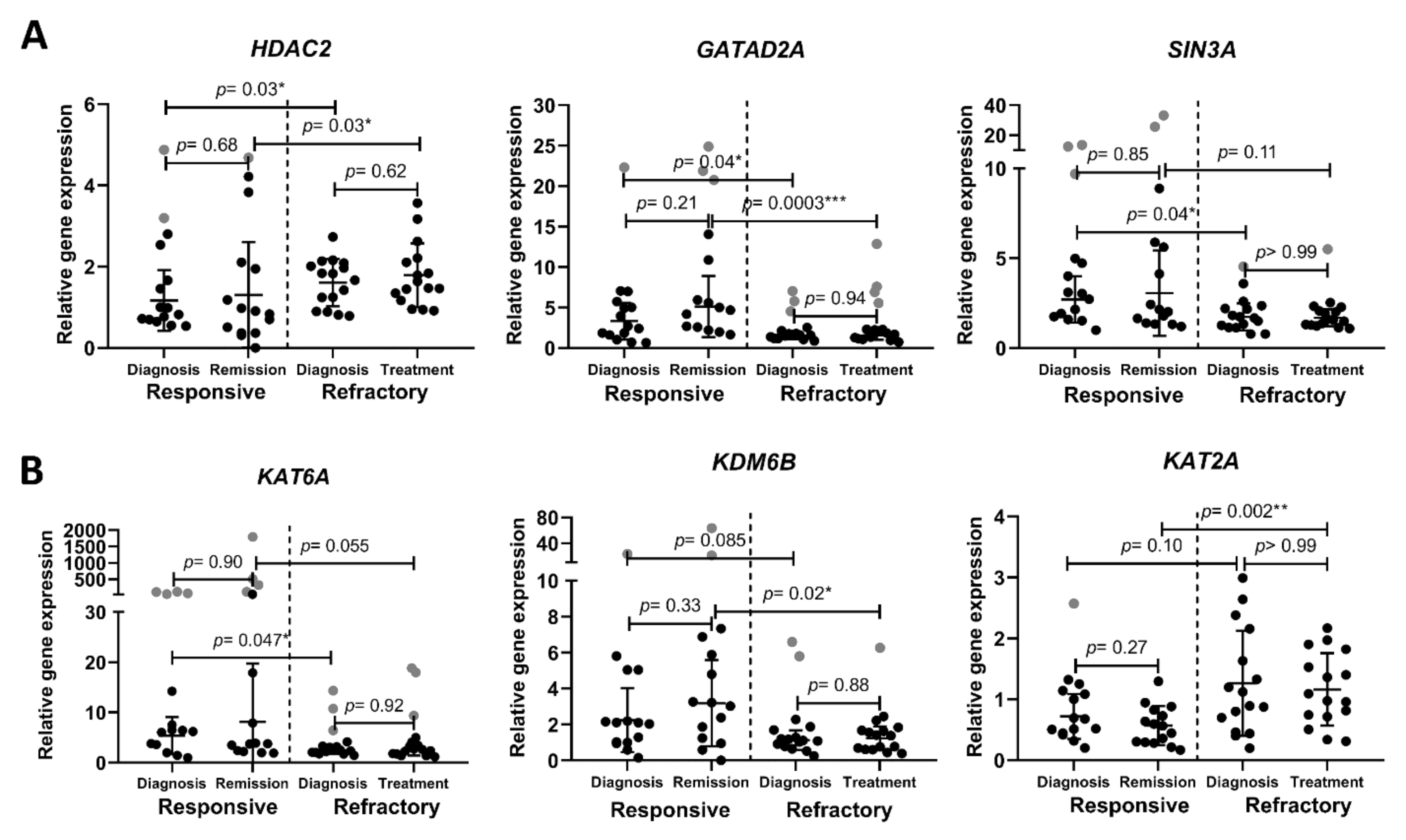
3.2. Chromatin-Remodeling-Related Gene Expression in AML Patients
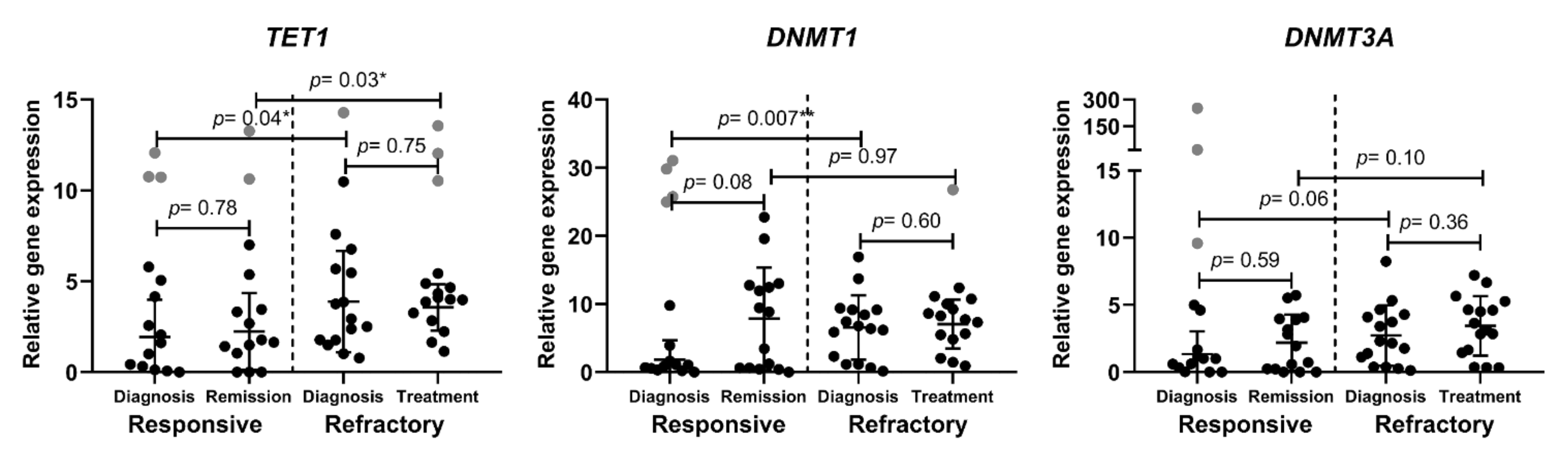
3.3. Epigenetic DNA Modification-Related Gene Expression in AML Patients
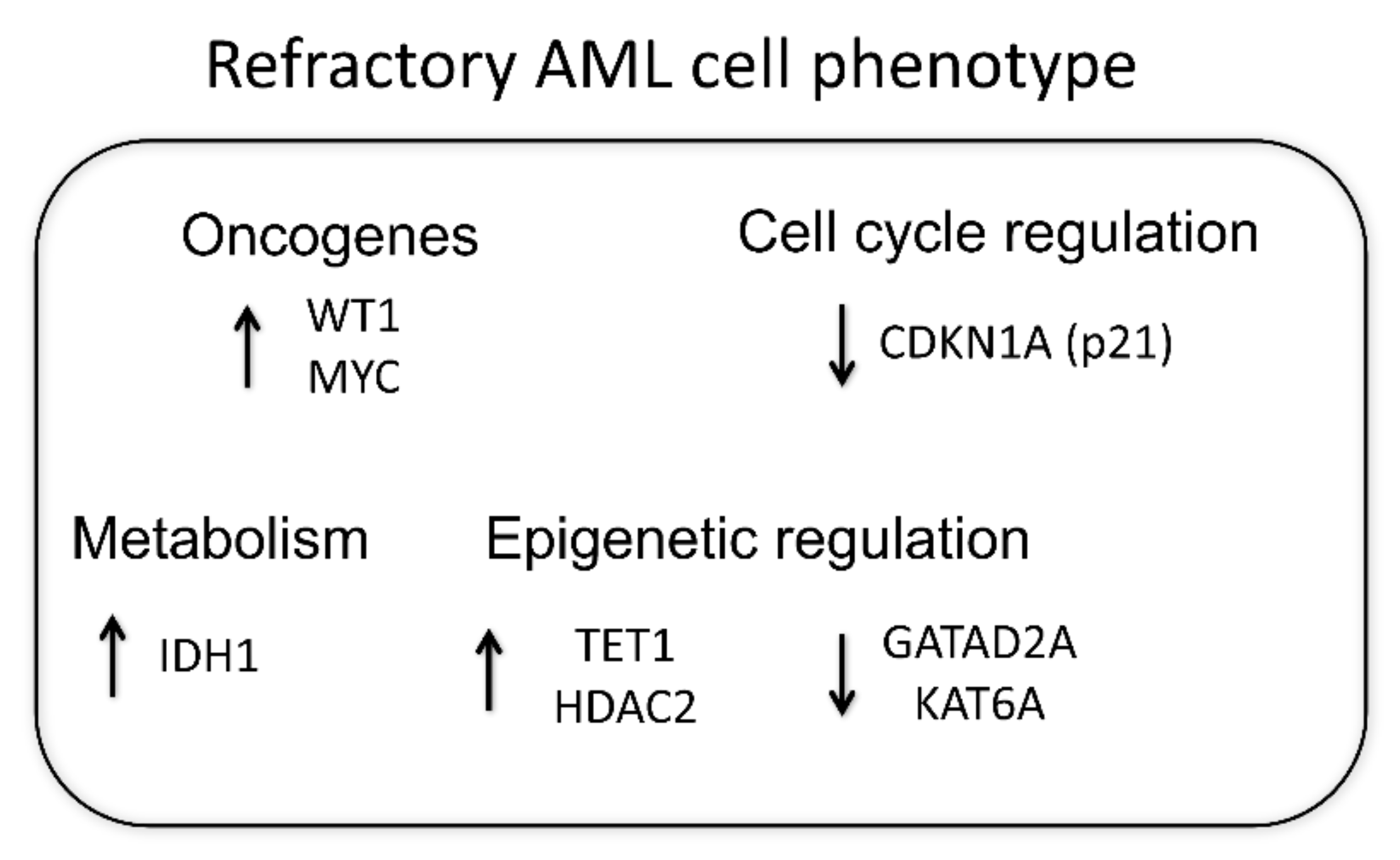
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, X.-L.; Liu, H.-Q.; Li, J.; Mao, C.-Y.; He, J.-T.; Zhao, X. Role of Epigenetic in Leukemia: From Mechanism to Therapy. Chem. Biol. Interact. 2020, 317, 108963. [Google Scholar] [CrossRef] [PubMed]
- Döhner, H.; Weisdorf, D.J.; Bloomfield, C.D. Acute Myeloid Leukemia. N. Engl. J. Med. 2015, 373, 1136–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagunas-Rangel, F.A.; Chávez-Valencia, V. FLT3-ITD and Its Current Role in Acute Myeloid Leukaemia. Med. Oncol. 2017, 34, 114. [Google Scholar] [CrossRef]
- Cai, S.F.; Levine, R.L. Genetic and Epigenetic Determinants of AML Pathogenesis. Semin. Hematol. 2019, 56, 84–89. [Google Scholar] [CrossRef]
- Schlenk, R.F.; Müller-Tidow, C.; Benner, A.; Kieser, M. Relapsed/Refractory Acute Myeloid Leukemia: Any Progress? Curr. Opin. Oncol. 2017, 29, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Bose, P.; Vachhani, P.; Cortes, J.E. Treatment of Relapsed/Refractory Acute Myeloid Leukemia. Curr. Treat. Options Oncol. 2017, 18, 17. [Google Scholar] [CrossRef] [PubMed]
- Staudt, D.; Murray, H.C.; McLachlan, T.; Alvaro, F.; Enjeti, A.K.; Verrills, N.M.; Dun, M.D. Targeting Oncogenic Signaling in Mutant FLT3 Acute Myeloid Leukemia: The Path to Least Resistance. Int. J. Mol. Sci. 2018, 19, E3198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levidou, G.; Gajdzis, P.; Cassoux, N.; Donizy, P.; Masaoutis, C.; Gajdzis, M.; Gardrat, S.; Pergaris, A.; Danas, E.; Klijanienko, J.; et al. Histone Deacetylase (HDAC)-1, -2, -4, and -6 in Uveal Melanomas: Associations with Clinicopathological Parameters and Patients’ Survival. Cancers 2021, 13, 4763. [Google Scholar] [CrossRef] [PubMed]
- Spruijt, C.G.; Luijsterburg, M.S.; Menafra, R.; Lindeboom, R.G.H.; Jansen, P.W.T.C.; Edupuganti, R.R.; Baltissen, M.P.; Wiegant, W.W.; Voelker-Albert, M.C.; Matarese, F.; et al. ZMYND8 Co-Localizes with NuRD on Target Genes and Regulates Poly(ADP-Ribose)-Dependent Recruitment of GATAD2A/NuRD to Sites of DNA Damage. Cell Rep. 2016, 17, 783–798. [Google Scholar] [CrossRef] [Green Version]
- Lai, A.Y.; Wade, P.A. Cancer Biology and NuRD: A Multifaceted Chromatin Remodelling Complex. Nat. Rev. Cancer 2011, 11, 588–596. [Google Scholar] [CrossRef]
- Lewis, M.J.; Liu, J.; Libby, E.F.; Lee, M.; Crawford, N.P.S.; Hurst, D.R. SIN3A and SIN3B Differentially Regulate Breast Cancer Metastasis. Oncotarget 2016, 7, 78713–78725. [Google Scholar] [CrossRef] [Green Version]
- Sheikh, B.N.; Yang, Y.; Schreuder, J.; Nilsson, S.K.; Bilardi, R.; Carotta, S.; McRae, H.M.; Metcalf, D.; Voss, A.K.; Thomas, T. MOZ (KAT6A) Is Essential for the Maintenance of Classically Defined Adult Hematopoietic Stem Cells. Blood 2016, 128, 2307–2318. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Abmayr, S.M.; Workman, J.L. Regulation of KAT6 Acetyltransferases and Their Roles in Cell Cycle Progression, Stem Cell Maintenance, and Human Disease. Mol. Cell. Biol. 2016, 36, 1900–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, D.; Jia, F.; Hou, Y.; Sang, Y.; Alvarez, A.A.; Zhang, W.; Gao, W.-Q.; Hu, B.; Cheng, S.-Y.; Ge, J.; et al. Histone Acetyltransferase KAT6A Upregulates PI3K/AKT Signaling through TRIM24 Binding. Cancer Res. 2017, 77, 6190–6201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.-H.; Zhu, K.-Y.; Chen, J.; Liu, X.-Z.; Xu, P.-F.; Zhang, W.; Yan, L.; Guo, H.-Z.; Zhu, J. JMJD3 Facilitates C/EBPβ-Centered Transcriptional Program to Exert Oncorepressor Activity in AML. Nat. Commun. 2018, 9, 3369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, D.; Song, Y.; Yu, Y.; Wang, D.; Liu, B.; Chen, L.; Li, X.; Li, Y.; Cheng, L.; Lv, F.; et al. KAT2A-Mediated AR Translocation into Nucleus Promotes Abiraterone-Resistance in Castration-Resistant Prostate Cancer. Cell Death Dis. 2021, 12, 787. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, N.; Zhang, Y.; Yu, H.; Wu, Q. KAT2B Is an Immune Infiltration-Associated Biomarker Predicting Prognosis and Response to Immunotherapy in Non-Small Cell Lung Cancer. Investig. New Drugs 2022, 40, 43–57. [Google Scholar] [CrossRef]
- Zhang, T.; Zhao, Y.; Zhao, Y.; Zhou, J. Expression and Prognosis Analysis of TET Family in Acute Myeloid Leukemia. Aging 2020, 12, 5031–5047. [Google Scholar] [CrossRef]
- Wong, K.K.; Lawrie, C.H.; Green, T.M. Oncogenic Roles and Inhibitors of DNMT1, DNMT3A, and DNMT3B in Acute Myeloid Leukaemia. Biomark. Insights 2019, 14, 6454. [Google Scholar] [CrossRef]
- Rein, L.A.M.; Chao, N.J. WT1 Vaccination in Acute Myeloid Leukemia: New Methods of Implementing Adoptive Immunotherapy. Expert Opin. Investig. Drugs 2014, 23, 417–426. [Google Scholar] [CrossRef]
- Gabay, M.; Li, Y.; Felsher, D.W. MYC Activation Is a Hallmark of Cancer Initiation and Maintenance. Cold Spring Harb. Perspect. Med 2014, 4, a014241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trop-Steinberg, S.; Azar, Y. Is Myc an Important Biomarker? Myc Expression in Immune Disorders and Cancer. Am. J. Med. Sci. 2018, 355, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Megías-Vericat, J.E.; Ballesta-López, O.; Barragán, E.; Montesinos, P. IDH1-Mutated Relapsed or Refractory AML: Current Challenges and Future Prospects. Blood Lymphat. Cancer 2019, 9, 19–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.-L.; Wang, J.-H.; Wang, Y.-G.; Hu, C.; Mu, Q.-T.; Yu, M.-X.; Wang, L.; Wang, D.-M.; Yang, M.; Yin, X.-F.; et al. High IDH1 Expression Is Associated with a Poor Prognosis in Cytogenetically Normal Acute Myeloid Leukemia. Int. J. Cancer 2015, 137, 1058–1065. [Google Scholar] [CrossRef] [PubMed]
- Gu, R.; Yang, X.; Wei, H. Molecular Landscape and Targeted Therapy of Acute Myeloid Leukemia. Biomark. Res. 2018, 6, 32. [Google Scholar] [CrossRef]
- El-Deiry, W.S. P21(WAF1) Mediates Cell-Cycle Inhibition, Relevant to Cancer Suppression and Therapy. Cancer Res. 2016, 76, 5189–5191. [Google Scholar] [CrossRef] [Green Version]
- Morris, V.A.; Cummings, C.L.; Korb, B.; Boaglio, S.; Oehler, V.G. Deregulated KLF4 Expression in Myeloid Leukemias Alters Cell Proliferation and Differentiation through MicroRNA and Gene Targets. Mol. Cell. Biol. 2016, 36, 559–573. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.H.; Bogenberger, J.M.; Tibes, R. Targeting Apoptosis in Acute Myeloid Leukemia: Current Status and Future Directions of BCL-2 Inhibition with Venetoclax and Beyond. Target. Oncol. 2020, 15, 147–162. [Google Scholar] [CrossRef]
- San José-Enériz, E.; Gimenez-Camino, N.; Agirre, X.; Prosper, F. HDAC Inhibitors in Acute Myeloid Leukemia. Cancers 2019, 11, 1794. [Google Scholar] [CrossRef] [Green Version]
- Min, C.; Moore, N.; Shearstone, J.R.; Quayle, S.N.; Huang, P.; van Duzer, J.H.; Jarpe, M.B.; Jones, S.S.; Yang, M. Selective Inhibitors of Histone Deacetylases 1 and 2 Synergize with Azacitidine in Acute Myeloid Leukemia. PLoS ONE 2017, 12, e0169128. [Google Scholar] [CrossRef]
- Weichert, W.; Röske, A.; Gekeler, V.; Beckers, T.; Stephan, C.; Jung, K.; Fritzsche, F.R.; Niesporek, S.; Denkert, C.; Dietel, M.; et al. Histone Deacetylases 1, 2 and 3 Are Highly Expressed in Prostate Cancer and HDAC2 Expression Is Associated with Shorter PSA Relapse Time after Radical Prostatectomy. Br. J. Cancer 2008, 98, 604–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H.; Ouchida, M.; Yamamoto, H.; Yano, M.; Toyooka, S.; Aoe, M.; Shimizu, N.; Date, H.; Shimizu, K. Decreased Expression of the SIN3A Gene, a Candidate Tumor Suppressor Located at the Prevalent Allelic Loss Region 15q23 in Non-Small Cell Lung Cancer. Lung Cancer 2008, 59, 24–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baell, J.B.; Leaver, D.J.; Hermans, S.J.; Kelly, G.L.; Brennan, M.S.; Downer, N.L.; Nguyen, N.; Wichmann, J.; McRae, H.M.; Yang, Y.; et al. Inhibitors of Histone Acetyltransferases KAT6A/B Induce Senescence and Arrest Tumour Growth. Nature 2018, 560, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Katsumoto, T.; Aikawa, Y.; Iwama, A.; Ueda, S.; Ichikawa, H.; Ochiya, T.; Kitabayashi, I. MOZ Is Essential for Maintenance of Hematopoietic Stem Cells. Genes Dev. 2006, 20, 1321–1330. [Google Scholar] [CrossRef] [Green Version]
- Thomas, T.; Corcoran, L.M.; Gugasyan, R.; Dixon, M.P.; Brodnicki, T.; Nutt, S.L.; Metcalf, D.; Voss, A.K. Monocytic Leukemia Zinc Finger Protein Is Essential for the Development of Long-Term Reconstituting Hematopoietic Stem Cells. Genes Dev. 2006, 20, 1175–1186. [Google Scholar] [CrossRef] [Green Version]
- Mallaney, C.; Ostrander, E.L.; Celik, H.; Kramer, A.C.; Martens, A.; Kothari, A.; Koh, W.K.; Haussler, E.; Iwamori, N.; Gontarz, P.; et al. Kdm6b Regulates Context-Dependent Hematopoietic Stem Cell Self-Renewal and Leukemogenesis. Leukemia 2019, 33, 2506–2521. [Google Scholar] [CrossRef]
- Ohguchi, H.; Harada, T.; Sagawa, M.; Kikuchi, S.; Tai, Y.-T.; Richardson, P.G.; Hideshima, T.; Anderson, K.C. KDM6B Modulates MAPK Pathway Mediating Multiple Myeloma Cell Growth and Survival. Leukemia 2017, 31, 2661–2669. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Zheng, H.; Bao, N.; Jiang, S.; Bueso-Ramos, C.E.; Khoury, J.; Class, C.; Lu, Y.; Lin, K.; Yang, H.; et al. KDM6B Overexpression Activates Innate Immune Signaling and Impairs Hematopoiesis in Mice. Blood Adv. 2018, 2, 2491–2504. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, M.; Sheng, M.; Zhang, P.; Chen, Z.; Xing, W.; Bai, J.; Cheng, T.; Yang, F.-C.; Zhou, Y. Therapeutic Potential of GSK-J4, a Histone Demethylase KDM6B/JMJD3 Inhibitor, for Acute Myeloid Leukemia. J. Cancer Res. Clin. Oncol. 2018, 144, 1065–1077. [Google Scholar] [CrossRef] [Green Version]
- Arede, L.; Pina, C. Buffering Noise: KAT2A Modular Contributions to Stabilization of Transcription and Cell Identity in Cancer and Development. Exp. Hematol. 2021, 93, 25–37. [Google Scholar] [CrossRef]
- Wang, J.; Li, F.; Ma, Z.; Yu, M.; Guo, Q.; Huang, J.; Yu, W.; Wang, Y.; Jin, J. High Expression of TET1 Predicts Poor Survival in Cytogenetically Normal Acute Myeloid Leukemia from Two Cohorts. EBioMedicine 2018, 28, 90–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuno, S.; Chijiwa, T.; Okamura, T.; Akashi, K.; Fukumaki, Y.; Niho, Y.; Sasaki, H. Expression of DNA Methyltransferases DNMT1, 3A, and 3B in Normal Hematopoiesis and in Acute and Chronic Myelogenous Leukemia. Blood 2001, 97, 1172–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Jin, X.; Wu, H.; Wang, Y.; Li, X.; Guo, Y.; Liang, S. HA117 Endows HL60 Cells with a Stem-like Signature by Inhibiting the Degradation of DNMT1 via Its Ability to down-Regulate Expression of the GGL Domain of RGS6. PLoS ONE 2017, 12, e0180142. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vitkevičienė, A.; Skliutė, G.; Žučenka, A.; Borutinskaitė, V.; Navakauskienė, R. Potential Prognostic Markers for Relapsed/Refractory vs. Responsive Acute Myeloid Leukemia. Cancers 2022, 14, 2752. https://doi.org/10.3390/cancers14112752
Vitkevičienė A, Skliutė G, Žučenka A, Borutinskaitė V, Navakauskienė R. Potential Prognostic Markers for Relapsed/Refractory vs. Responsive Acute Myeloid Leukemia. Cancers. 2022; 14(11):2752. https://doi.org/10.3390/cancers14112752
Chicago/Turabian StyleVitkevičienė, Aida, Giedrė Skliutė, Andrius Žučenka, Veronika Borutinskaitė, and Rūta Navakauskienė. 2022. "Potential Prognostic Markers for Relapsed/Refractory vs. Responsive Acute Myeloid Leukemia" Cancers 14, no. 11: 2752. https://doi.org/10.3390/cancers14112752
APA StyleVitkevičienė, A., Skliutė, G., Žučenka, A., Borutinskaitė, V., & Navakauskienė, R. (2022). Potential Prognostic Markers for Relapsed/Refractory vs. Responsive Acute Myeloid Leukemia. Cancers, 14(11), 2752. https://doi.org/10.3390/cancers14112752