
Exosomes from EGFR-Mutated Adenocarcinoma Induce a Hybrid EMT and MMP9-Dependant Tumor Invasion

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Serum Samples
2.2. Cell Lines
2.3. Isolation of Exosomes from Serum and Cell Supernatant by Differential Centrifugation
2.4. Western Blot Analysis
2.5. Phospho-Kinase Array
2.6. Transmission Electron Microscopy of Exosomes
2.7. Nanotracking Analysis
2.8. Immunofluorescence
2.9. Migration and Invasion Assays
2.10. 3D-Organotypic Growth Assays
2.11. Quantitative Real-Time PCR
2.12. EMT Scoring Method
2.13. Statistical Analysis
2.14. Study Approval
3. Results
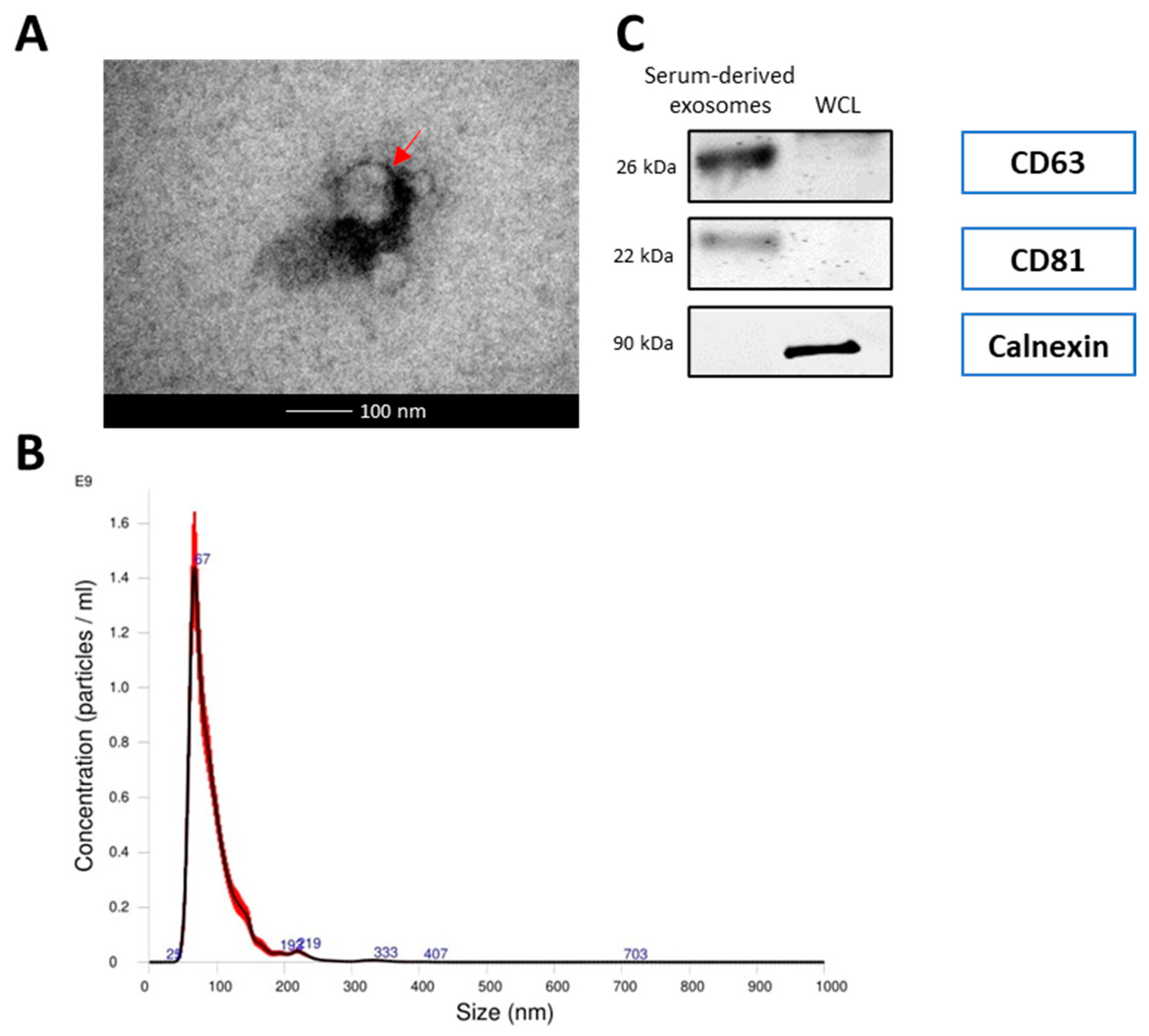
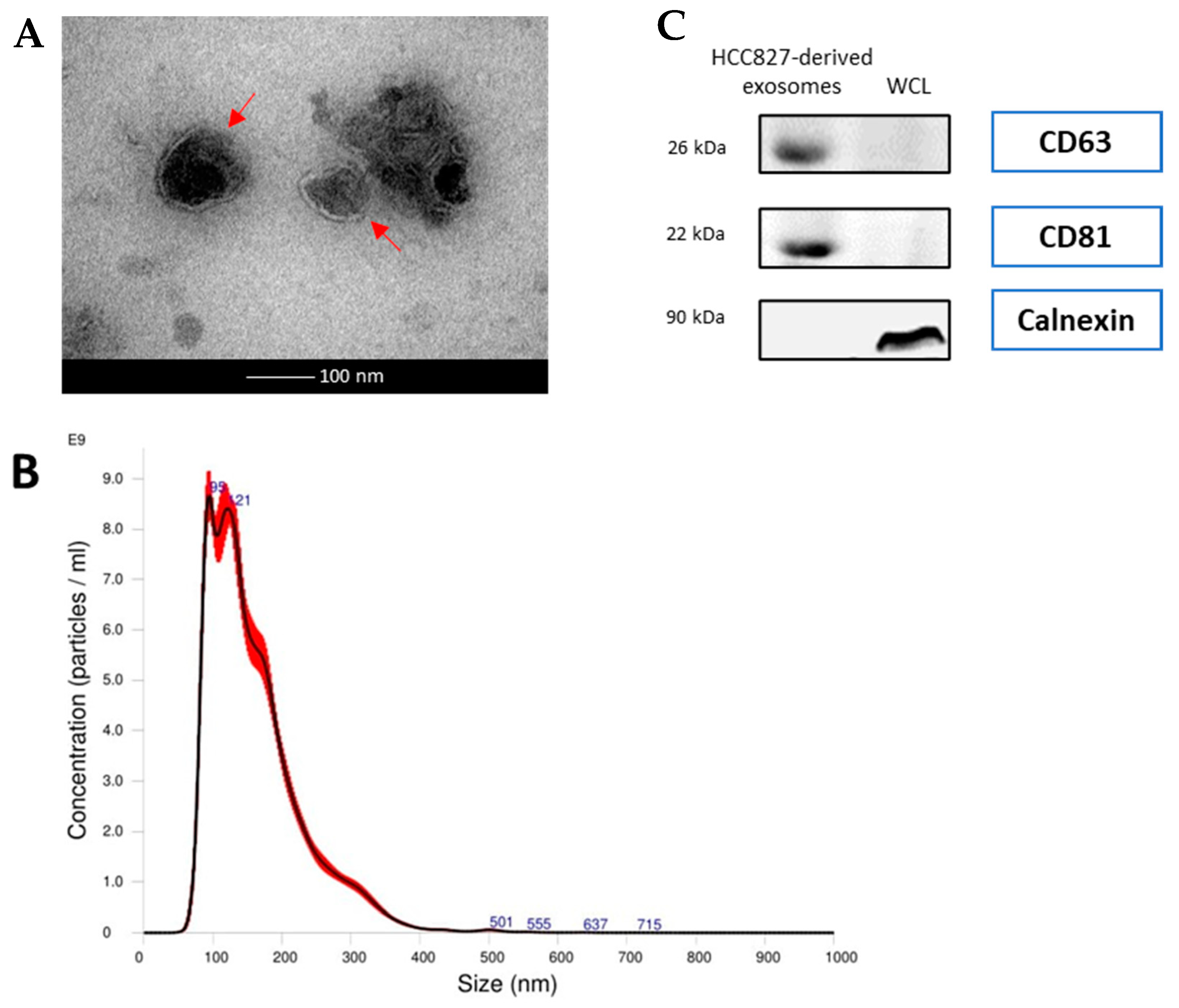
3.1. Exosomes Derived from Serum of Lung Cancer Patients and HCC827 Cells Were Isolated and Confirmed
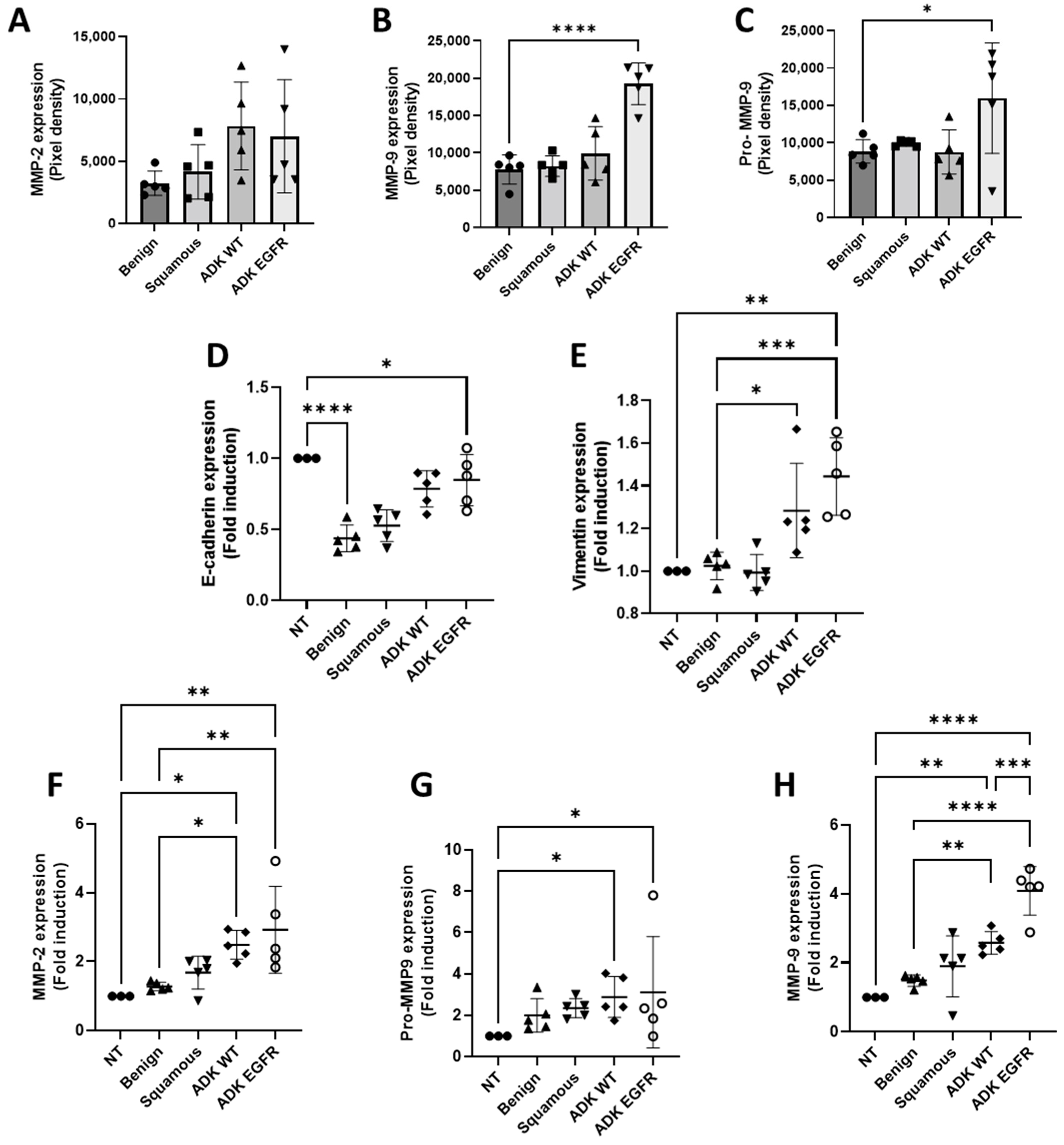
3.2. Exosomes Derived from Serum of EGFR-Mutated Patients and from EGFR-Mutated Cell Line Promote Extracellular Matrix Degradation and EMT
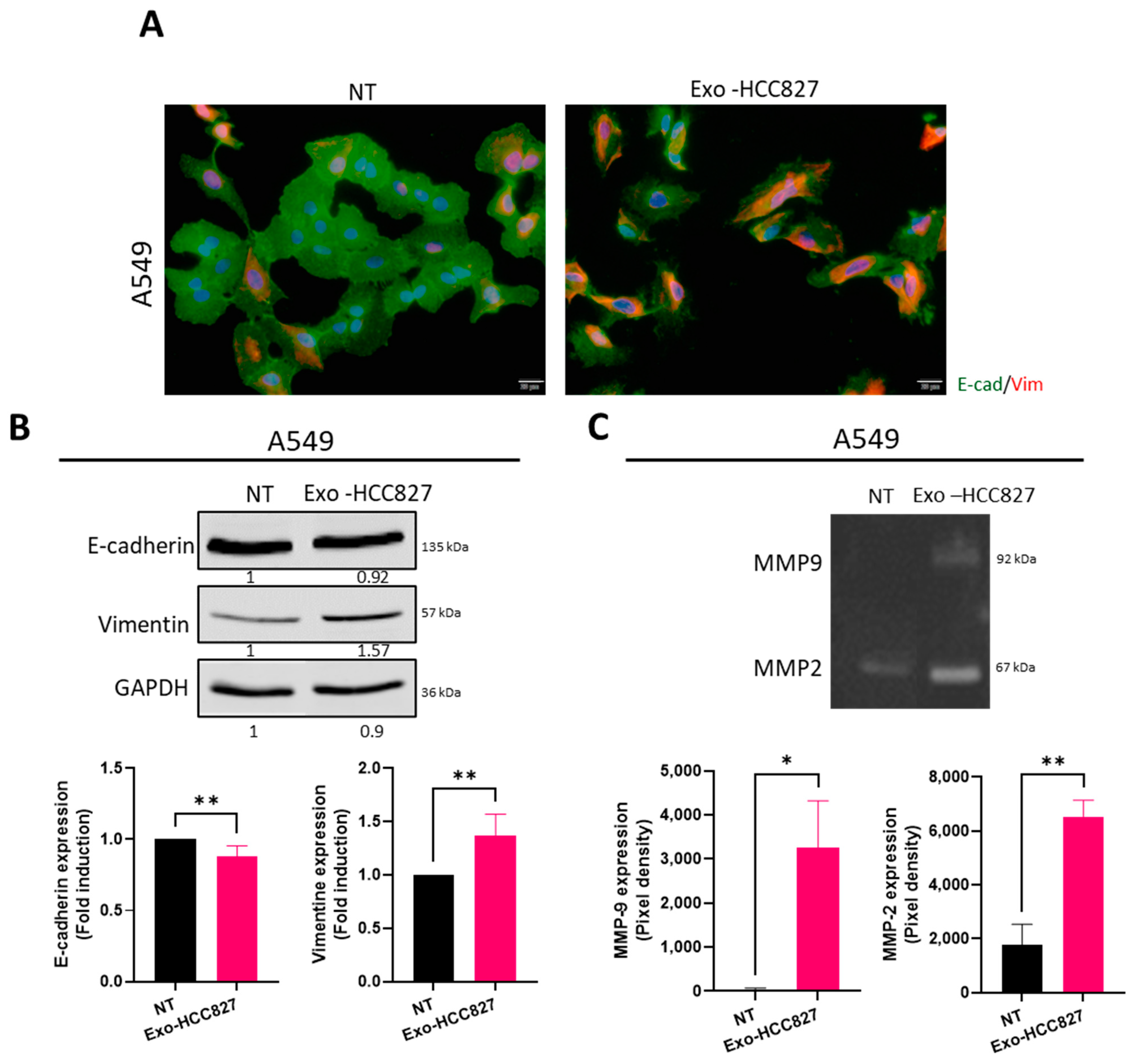
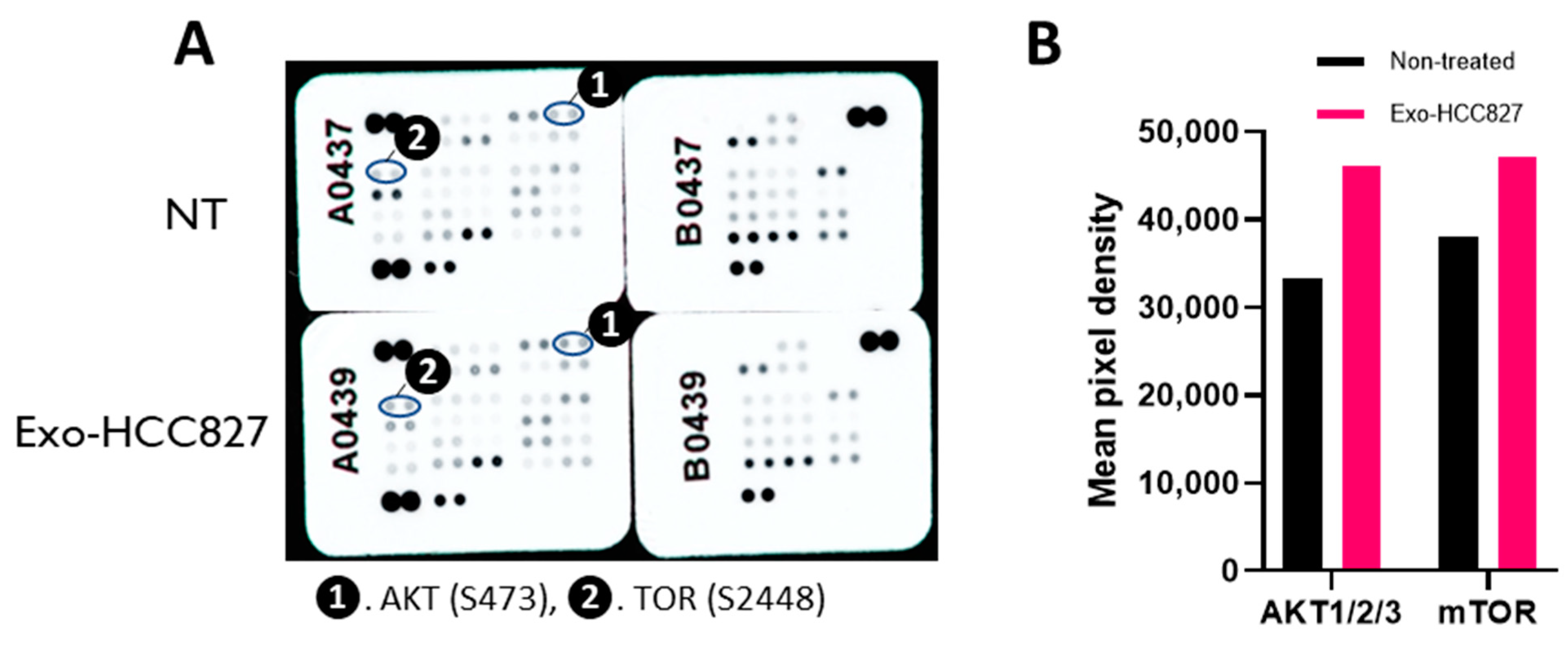
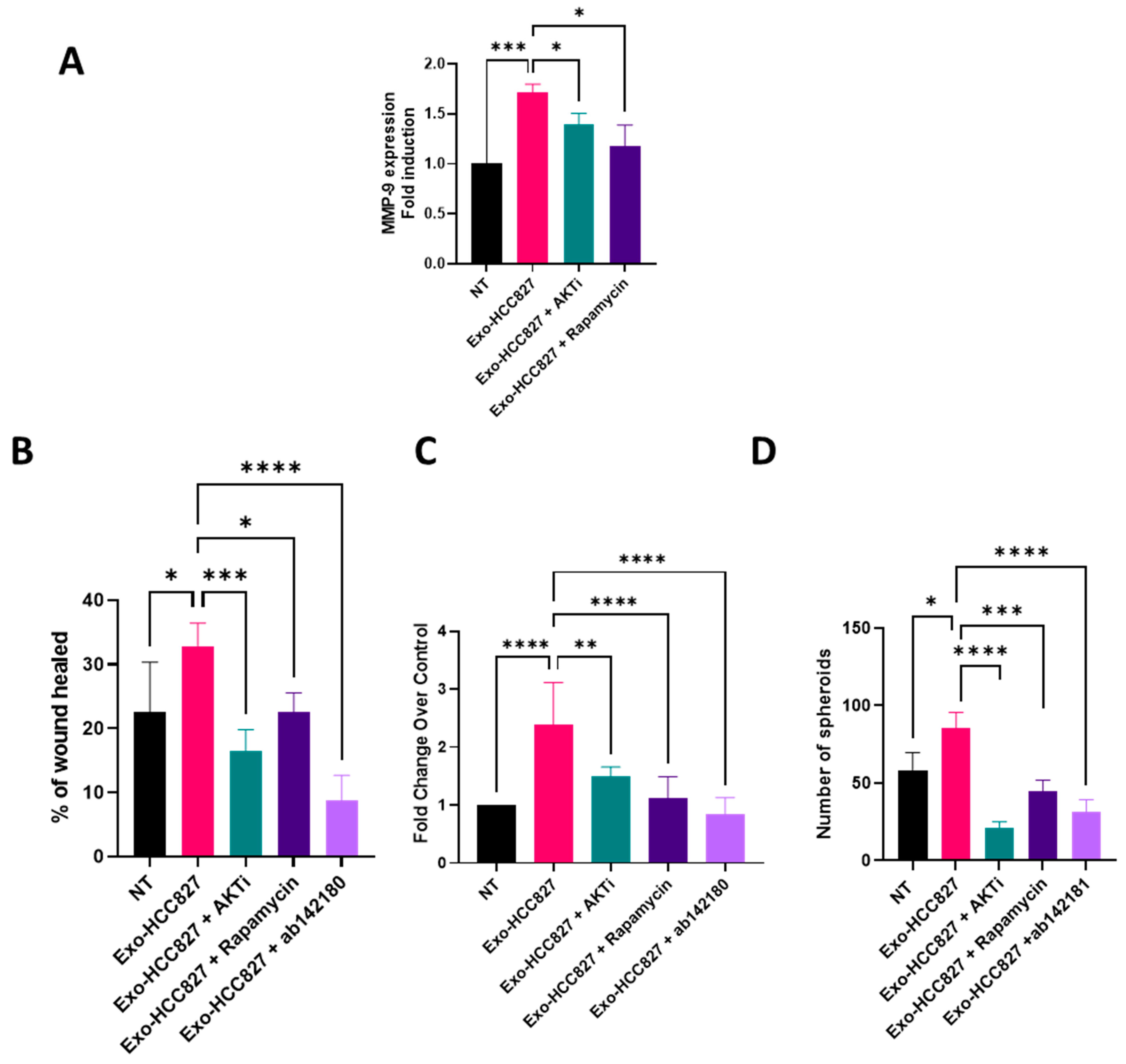
3.3. Exosomes Derived from EGFR-Mutated Cell Line Induce Invasion That Is MMP-9 Dependent
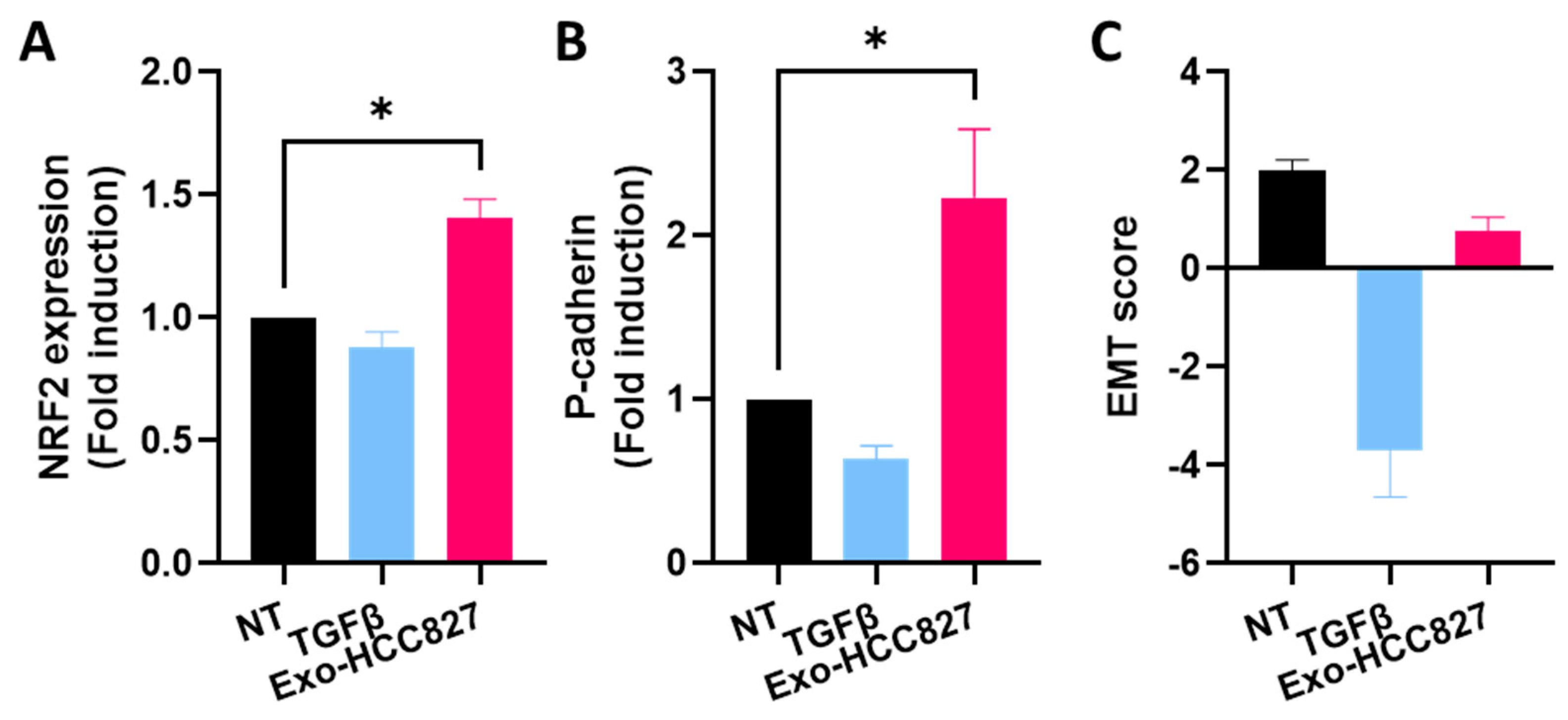
3.4. Exosomes Derived from EGFR-Mutated Cell Line Induce Partial/Hybrid EMT
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R. The Biology and Function of Exosomes in Cancer. J. Clin. Investig. 2016, 126, 1208–1215. [Google Scholar] [CrossRef] [PubMed]
- Skog, J.; Würdinger, T.; van Rijn, S.; Meijer, D.H.; Gainche, L.; Sena-Esteves, M.; Curry, W.T.; Carter, B.S.; Krichevsky, A.M.; Breakefield, X.O. Glioblastoma Microvesicles Transport RNA and Proteins That Promote Tumour Growth and Provide Diagnostic Biomarkers. Nat. Cell Biol. 2008, 10, 1470–1476. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-Mediated Transfer of MRNAs and MicroRNAs Is a Novel Mechanism of Genetic Exchange between Cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Melo, S.A.; Sugimoto, H.; O’Connell, J.T.; Kato, N.; Villanueva, A.; Vidal, A.; Qiu, L.; Vitkin, E.; Perelman, L.T.; Melo, C.A.; et al. Cancer Exosomes Perform Cell-Independent MicroRNA Biogenesis and Promote Tumorigenesis. Cancer Cell 2014, 26, 707–721. [Google Scholar] [CrossRef] [Green Version]
- Melo, S.A.; Luecke, L.B.; Kahlert, C.; Fernandez, A.F.; Gammon, S.T.; Kaye, J.; LeBleu, V.S.; Mittendorf, E.A.; Weitz, J.; Rahbari, N.; et al. Glypican1 Identifies Cancer Exosomes and Facilitates Early Detection of Cancer. Nature 2015, 523, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Caradec, J.; Kharmate, G.; Hosseini-Beheshti, E.; Adomat, H.; Gleave, M.; Guns, E. Reproducibility and Efficiency of Serum-Derived Exosome Extraction Methods. Clin. Biochem. 2014, 47, 1286–1292. [Google Scholar] [CrossRef]
- Maisano, D.; Mimmi, S.; Dattilo, V.; Marino, F.; Gentile, M.; Vecchio, E.; Fiume, G.; Nisticò, N.; Aloisio, A.; de Santo, M.P.; et al. A Novel Phage Display Based Platform for Exosome Diversity Characterization. Nanoscale 2022, 14, 2998–3003. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Barger, J.F.; Lovat, F.; Gao, M.; Otterson, G.A.; Nana-Sinkam, P. Lung Cancer Exosomes as Drivers of Epithelial Mesenchymal Transition. Oncotarget 2016, 7, 54852–54866. [Google Scholar] [CrossRef]
- Nieto, M.A.; Huang, R.Y.-J.; Jackson, R.A.; Thiery, J.P. EMT: 2016. Cell 2016, 166, 21–45. [Google Scholar] [CrossRef] [Green Version]
- Jolly, M.K.; Boareto, M.; Huang, B.; Jia, D.; Lu, M.; Ben-Jacob, E.; Onuchic, J.N.; Levine, H. Implications of the Hybrid Epithelial/Mesenchymal Phenotype in Metastasis. Front. Oncol. 2015, 5, 155. [Google Scholar] [CrossRef] [Green Version]
- Suster, D.I.; Mino-Kenudson, M. Molecular Pathology of Primary Non-Small Cell Lung Cancer. Arch. Med. Res. 2020, 51, 784–798. [Google Scholar] [CrossRef]
- Tfayli, A.H.; Fakhri, G.B.; Al Assaad, M.S. Prevalence of the Epidermal Growth Factor Receptor Mutations in Lung Adenocarcinoma Patients from the Middle East Region. Ann. Thorac. Med. 2019, 14, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Digumarthy, S.R.; Mendoza, D.P.; Padole, A.; Chen, T.; Peterson, P.G.; Piotrowska, Z.; Sequist, L.V. Diffuse Lung Metastases in EGFR-Mutant Non-Small Cell Lung Cancer. Cancers 2019, 11, 1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindeman, N.I.; Cagle, P.T.; Aisner, D.L.; Arcila, M.E.; Beasley, M.B.; Bernicker, E.H.; Colasacco, C.; Dacic, S.; Hirsch, F.R.; Kerr, K.; et al. Updated Molecular Testing Guideline for the Selection of Lung Cancer Patients for Treatment With Targeted Tyrosine Kinase Inhibitors: Guideline From the College of American Pathologists, the International Association for the Study of Lung Cancer, and the Association for Molecular Pathology. Arch. Pathol. Lab. Med. 2018, 142, 321–346. [Google Scholar] [CrossRef] [Green Version]
- Bernabé, R.; Hickson, N.; Wallace, A.; Blackhall, F.H. What Do We Need to Make Circulating Tumour DNA (CtDNA) a Routine Diagnostic Test in Lung Cancer? Eur. J. Cancer 2017, 81, 66–73. [Google Scholar] [CrossRef]
- O’Callaghan, M.; Ryan, M.E.; Cotter, M.; Kelly, A.; O’Mahony, A.; Hanrahan, E.; Higgins, M.; Keane, M.; Fabre, A. 51—Plasma Cell-Free DNA EGFR Mutation Detection in Patients with Non-Small Cell Lung Carcinoma Using Two Commercially Available Platforms the COBAS® and IdyllaTM. Lung Cancer 2021, 156, S21. [Google Scholar] [CrossRef]
- Jouida, A.; McCarthy, C.; Fabre, A.; Keane, M.P. Exosomes: A New Perspective in EGFR-Mutated Lung Cancer. Cancer Metastasis Rev. 2021, 40, 589–601. [Google Scholar] [CrossRef]
- Chae, Y.K.; Chang, S.; Ko, T.; Anker, J.; Agte, S.; Iams, W.; Choi, W.M.; Lee, K.; Cruz, M. Epithelial-Mesenchymal Transition (EMT) Signature Is Inversely Associated with T-Cell Infiltration in Non-Small Cell Lung Cancer (NSCLC). Sci. Rep. 2018, 8, 2918. [Google Scholar] [CrossRef] [Green Version]
- Chirshev, E.; Hojo, N.; Bertucci, A.; Sanderman, L.; Nguyen, A.; Wang, H.; Suzuki, T.; Brito, E.; Martinez, S.R.; Castañón, C.; et al. Epithelial/Mesenchymal Heterogeneity of High-grade Serous Ovarian Carcinoma Samples Correlates with MiRNA Let-7 Levels and Predicts Tumor Growth and Metastasis. Mol. Oncol. 2020, 14, 2796–2813. [Google Scholar] [CrossRef]
- Moreno-Bueno, G.; Peinado, H.; Molina, P.; Olmeda, D.; Cubillo, E.; Santos, V.; Palacios, J.; Portillo, F.; Cano, A. The Morphological and Molecular Features of the Epithelial-to-Mesenchymal Transition. Nat. Protoc. 2009, 4, 1591–1613. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Lei, L.; Wang, W.; Lin, L.; Zhu, Y.; Wang, H.; Miao, L.; Wang, L.; Zhuang, W.; Fang, M.; et al. Molecular Characteristics and Clinical Outcomes of EGFR Exon 19 C-Helix Deletion in Non–Small Cell Lung Cancer and Response to EGFR TKIs. Transl. Oncol. 2020, 13, 100791. [Google Scholar] [CrossRef] [PubMed]
- Bocci, F.; Tripathi, S.C.; Vilchez Mercedes, S.A.; George, J.T.; Casabar, J.P.; Wong, P.K.; Hanash, S.M.; Levine, H.; Onuchic, J.N.; Jolly, M.K. NRF2 Activates a Partial Epithelial-Mesenchymal Transition and Is Maximally Present in a Hybrid Epithelial/Mesenchymal Phenotype. Integr. Biol. Quant. Biosci. Nano Macro 2019, 11, 251–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, A.S.; Paredes, J. P-Cadherin Linking Breast Cancer Stem Cells and Invasion: A Promising Marker to Identify an “Intermediate/Metastable” EMT State. Front. Oncol. 2014, 4, 371. [Google Scholar] [CrossRef]
- Kim, B.N.; Ahn, D.H.; Kang, N.; Yeo, C.D.; Kim, Y.K.; Lee, K.Y.; Kim, T.-J.; Lee, S.H.; Park, M.S.; Yim, H.W.; et al. TGF-β Induced EMT and Stemness Characteristics Are Associated with Epigenetic Regulation in Lung Cancer. Sci. Rep. 2020, 10, 10597. [Google Scholar] [CrossRef] [PubMed]
- Merchant, N.; Nagaraju, G.P.; Rajitha, B.; Lammata, S.; Jella, K.K.; Buchwald, Z.S.; Lakka, S.S.; Ali, A.N. Matrix Metalloproteinases: Their Functional Role in Lung Cancer. Carcinogenesis 2017, 38, 766–780. [Google Scholar] [CrossRef] [Green Version]
- Shimoda, M.; Khokha, R. Metalloproteinases in Extracellular Vesicles. Biochim. Biophys. Acta BBA-Mol. Cell Res. 2017, 1864, 1989–2000. [Google Scholar] [CrossRef]
- Nawaz, M.; Shah, N.; Zanetti, B.R.; Maugeri, M.; Silvestre, R.N.; Fatima, F.; Neder, L.; Valadi, H. Extracellular Vesicles and Matrix Remodeling Enzymes: The Emerging Roles in Extracellular Matrix Remodeling, Progression of Diseases and Tissue Repair. Cells 2018, 7, 167. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Xie, Y.; Xu, L.; Zhan, S.; Xiao, Y.; Gao, Y.; Wu, B.; Ge, W. Protein Content and Functional Characteristics of Serum-Purified Exosomes from Patients with Colorectal Cancer Revealed by Quantitative Proteomics. Int. J. Cancer 2017, 140, 900–913. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Adhikari, N.; Banerjee, S.; Amin, S.A.; Jha, T. Matrix Metalloproteinase-9 (MMP-9) and Its Inhibitors in Cancer: A Minireview. Eur. J. Med. Chem. 2020, 194, 112260. [Google Scholar] [CrossRef]
- Peng, Y.; Wang, Y.; Zhou, C.; Mei, W.; Zeng, C. PI3K/Akt/MTOR Pathway and Its Role in Cancer Therapeutics: Are We Making Headway? Front. Oncol. 2022, 12, 819128. [Google Scholar] [CrossRef]
- Jolly, M.K.; Somarelli, J.A.; Sheth, M.; Biddle, A.; Tripathi, S.C.; Armstrong, A.J.; Hanash, S.M.; Bapat, S.A.; Rangarajan, A.; Levine, H. Hybrid Epithelial/Mesenchymal Phenotypes Promote Metastasis and Therapy Resistance across Carcinomas. Pharmacol. Ther. 2019, 194, 161–184. [Google Scholar] [CrossRef] [PubMed]
- Barrera-Rodríguez, R. Importance of the Keap1-Nrf2 Pathway in NSCLC: Is It a Possible Biomarker? Biomed. Rep. 2018, 9, 375–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, S.; Kobayashi, M.; Takasaki, C.; Ishibashi, H.; Okubo, K. High Expression of P-Cadherin Is Significantly Associated with Poor Prognosis in Patients with Non-Small-Cell Lung Cancer. Lung Cancer Amst. Neth. 2018, 118, 13–19. [Google Scholar] [CrossRef]
- Vilchez Mercedes, S.A.; Bocci, F.; Ahmed, M.; Eder, I.; Zhu, N.; Levine, H.; Onuchic, J.N.; Jolly, M.K.; Wong, P.K. Nrf2 Modulates the Hybrid Epithelial/Mesenchymal Phenotype and Notch Signaling During Collective Cancer Migration. Front. Mol. Biosci. 2022, 9, 807324. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, M. Involvement of Partial EMT in Cancer Progression. J. Biochem. (Tokyo) 2018, 164, 257–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galvez, C.; Jacob, S.; Finkelman, B.S.; Zhao, J.; Tegtmeyer, K.; Chae, Y.K.; Mohindra, N.; Salgia, R.; Jovanovic, B.; Behdad, A.; et al. The Role of EGFR Mutations in Predicting Recurrence in Early and Locally Advanced Lung Adenocarcinoma Following Definitive Therapy. Oncotarget 2020, 11, 1953–1960. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence |
|---|---|
| Vimentin | Forward primer: 5′-TGTCCAAATCGATGTGGATGTTTC-3′ Reverse primer: 5′-TTGTACCATTCTTCTGCCTCCTG-3′ |
| E-cadherin | Forward primer: 5′-GCTGAGCTGGACAGGGAGGA-3′ Reverse primer: 5′-ATGGGGGCGTTGTCATTCAC-3′ |
| MMP-9 (matrix metalloproteinase-9) | Forward primer: 5′-GGCGCTCATGTACCCTATGT-3′ Reverse primer: 5′-TCAAAGACCGAGTCCAGCTT-3′ |
| MMP-2 (matrix metalloproteinase-2) | Forward primer: 5′-GGCCCTGTCACTCCTGAGAT-3′ Reverse primer: 5′-GGCATCCAGGTTATCGGGGA-3′ |
| P-cadherin/CDH3 | Forward primer: 5′-AAATGCTCAACCCTGTGTCC-3′ Reverse primer: 5′-ATAGCAACGCAACAGGGAAA-3′ |
| EPCAM (Epithelial cell adhesion molecule) | Forward primer: 5′-GAAGGCTGAGATAAAGGAGATGGG-3′ Reverse primer:5′-TTAACGATGGAGTCCAA GTTCTGG-3′ |
| NRF2 (nuclear factor erythroid 2-related factor 2) | Forward primer: 5′-CAGCGACGGAAAGAGTATGA-3′ Reverse primer:5′-TGGGCAACCTGGGAGTAG-3′ |
| GAPDH (Glyceraldehyde 3-phosphate dehydrogenase) | Forward primer: 5′-CCATGTTCGTCATGGGTGTG-3′ Reverse primer: 5′-CAGGGGTGCTAAGCAGTTGG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jouida, A.; O’Callaghan, M.; Mc Carthy, C.; Fabre, A.; Nadarajan, P.; Keane, M.P. Exosomes from EGFR-Mutated Adenocarcinoma Induce a Hybrid EMT and MMP9-Dependant Tumor Invasion. Cancers 2022, 14, 3776. https://doi.org/10.3390/cancers14153776
Jouida A, O’Callaghan M, Mc Carthy C, Fabre A, Nadarajan P, Keane MP. Exosomes from EGFR-Mutated Adenocarcinoma Induce a Hybrid EMT and MMP9-Dependant Tumor Invasion. Cancers. 2022; 14(15):3776. https://doi.org/10.3390/cancers14153776
Chicago/Turabian StyleJouida, Amina, Marissa O’Callaghan, Cormac Mc Carthy, Aurelie Fabre, Parthiban Nadarajan, and Michael P. Keane. 2022. "Exosomes from EGFR-Mutated Adenocarcinoma Induce a Hybrid EMT and MMP9-Dependant Tumor Invasion" Cancers 14, no. 15: 3776. https://doi.org/10.3390/cancers14153776
APA StyleJouida, A., O’Callaghan, M., Mc Carthy, C., Fabre, A., Nadarajan, P., & Keane, M. P. (2022). Exosomes from EGFR-Mutated Adenocarcinoma Induce a Hybrid EMT and MMP9-Dependant Tumor Invasion. Cancers, 14(15), 3776. https://doi.org/10.3390/cancers14153776