
Expression of SARS-CoV-2-Related Surface Proteins in Non-Small-Cell Lung Cancer Patients and the Influence of Standard of Care Therapy

 , , , ,
, , , ,  ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. FFPE Tissue Samples
2.2. Serum Samples
2.3. Immunohistochemical Staining
2.4. ACE2 ELISA
2.5. TCGA and Statistical Analysis
3. Results
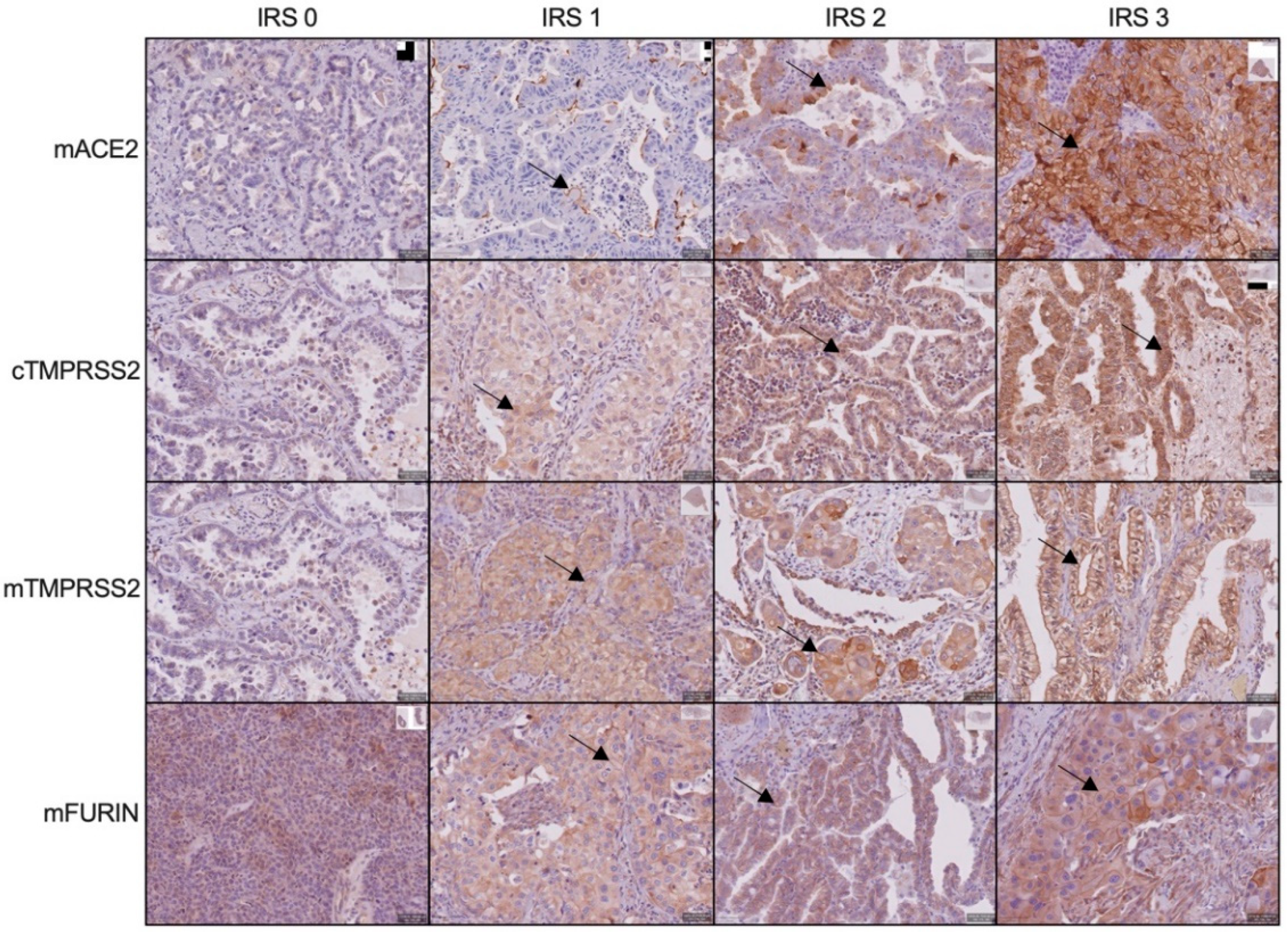
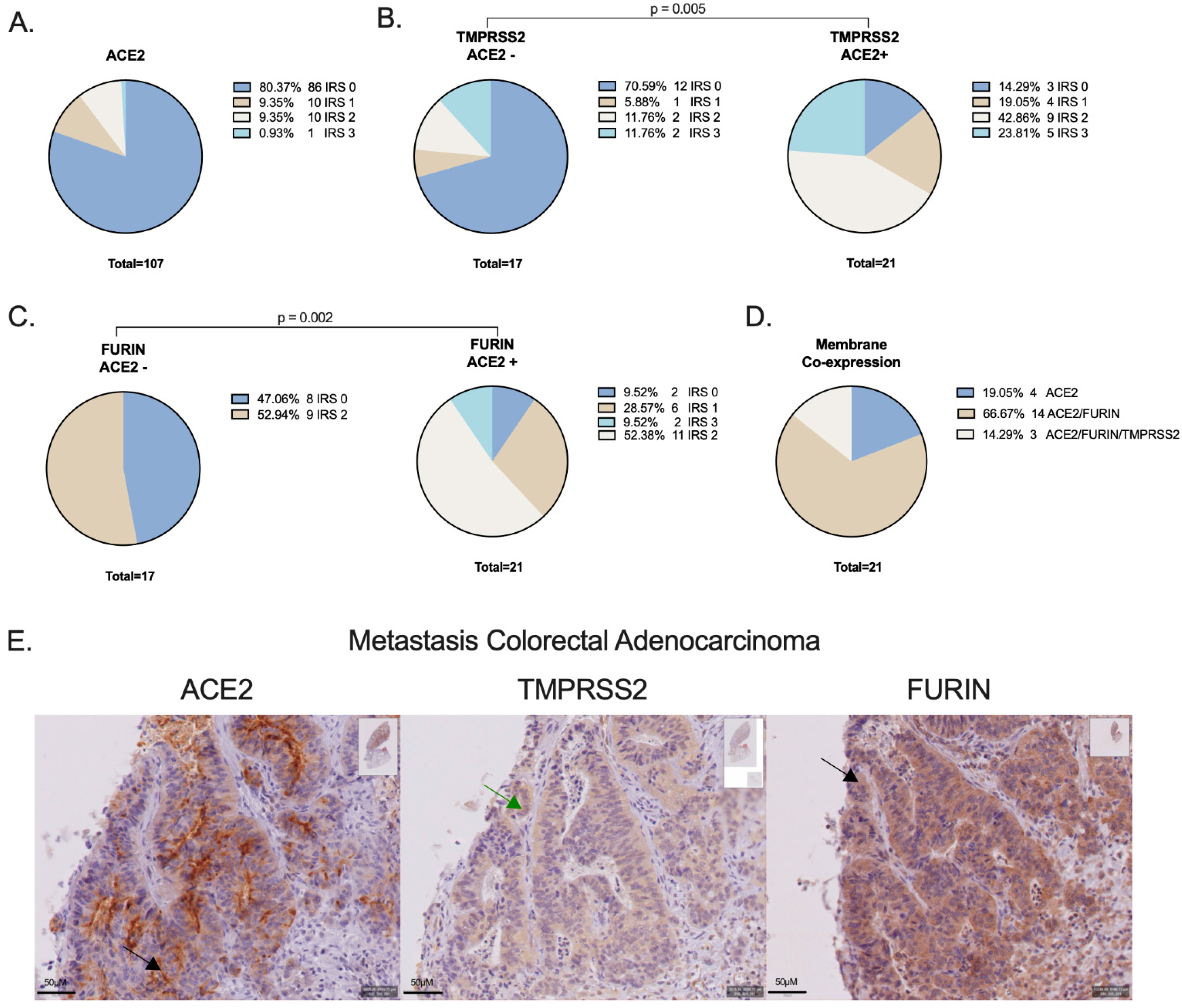
3.1. ACE2, TMPRSS2 and FURIN Protein Expression on NSCLC Cells
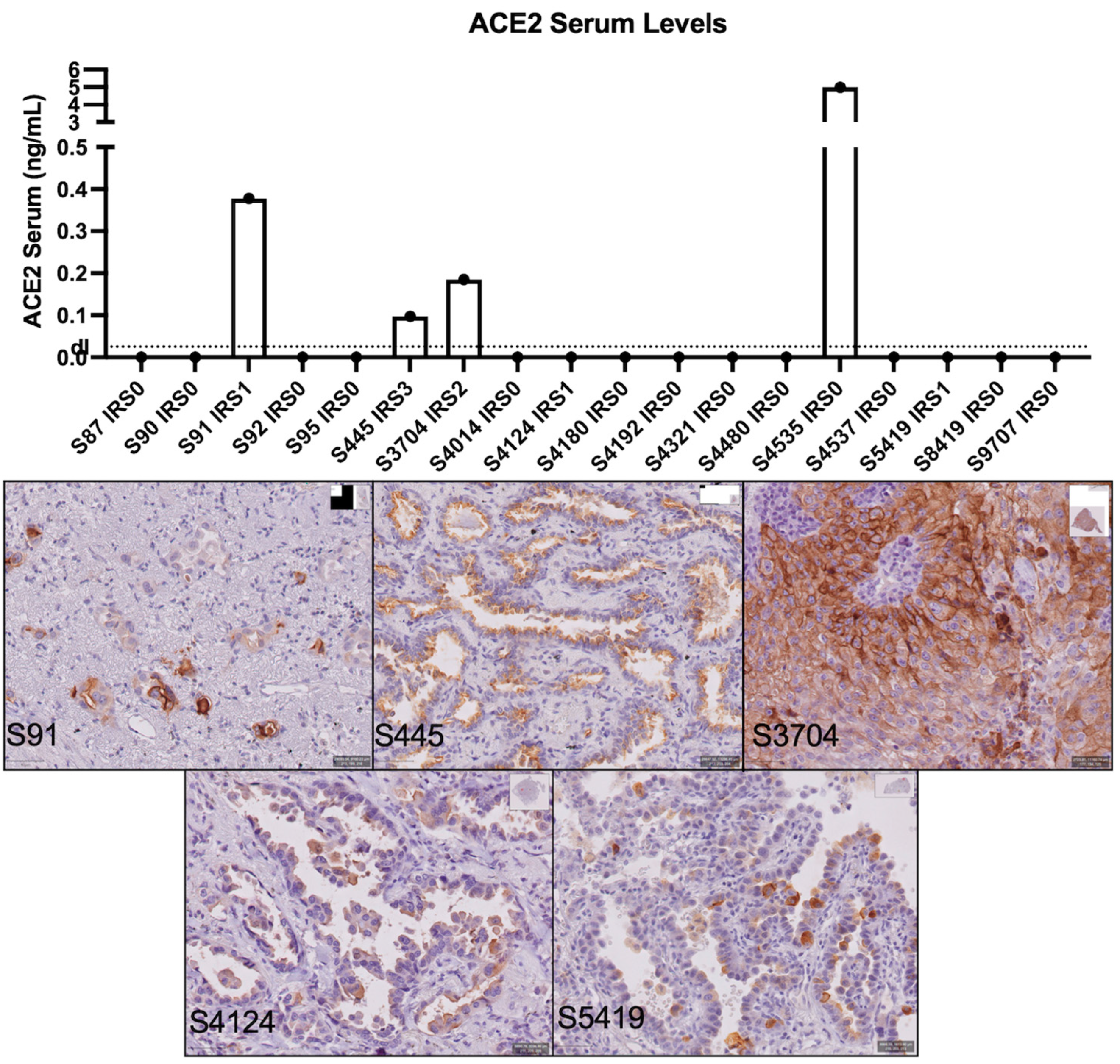
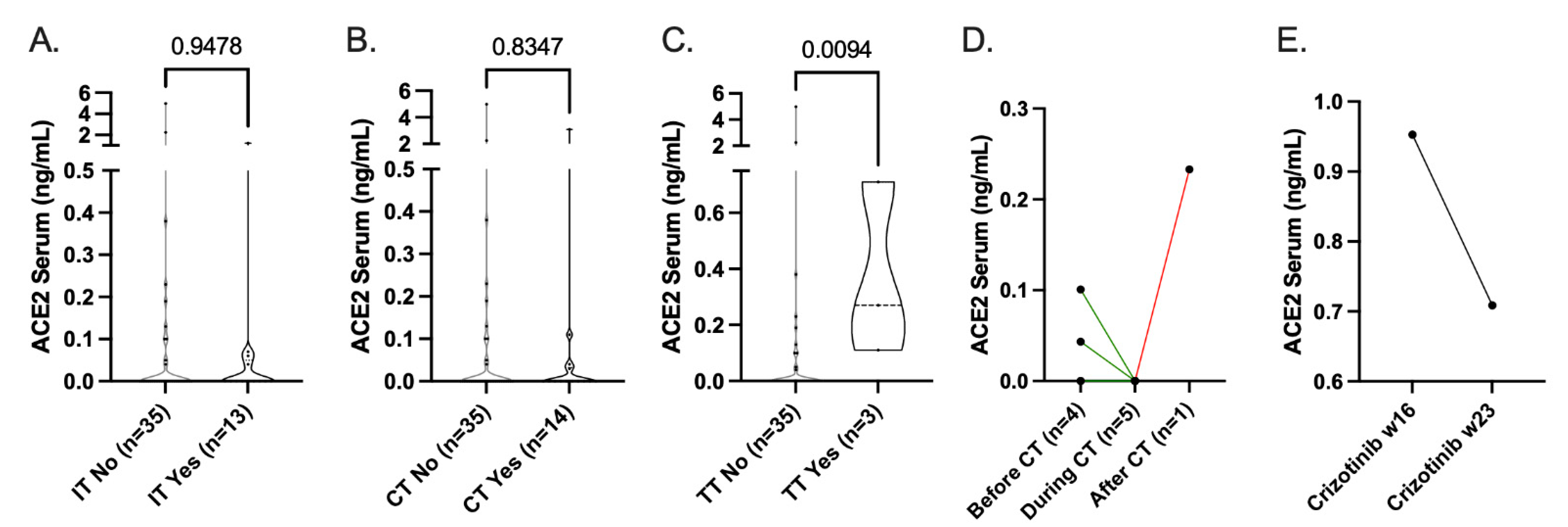
3.2. sACE2 Levels
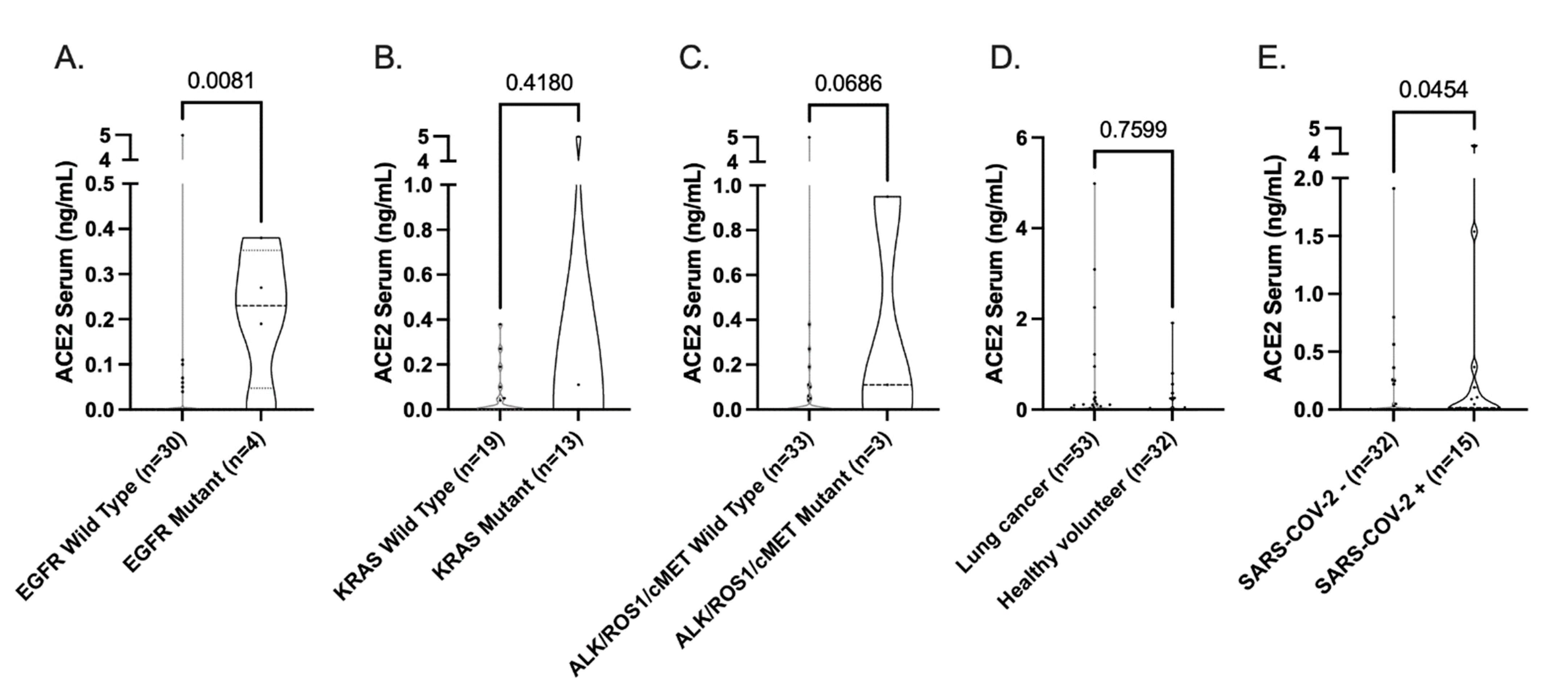
3.3. Influence of Standard-of-Care Systemic Therapy on ACE2 Expression
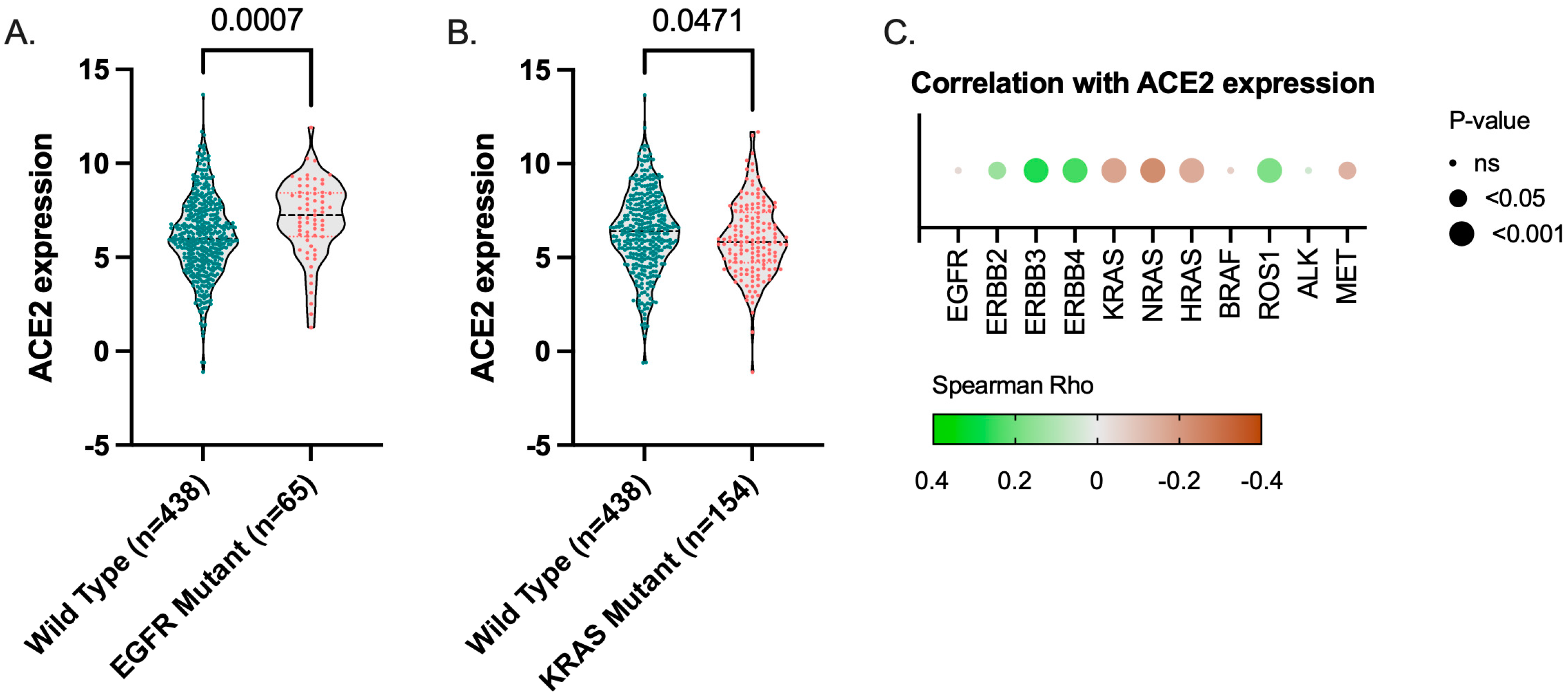
3.4. ACE2 Expression TCGA LUAD Cohort
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-COV-2 cell entry depends on ace2 and tmprss2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Lukassen, S.; Lorenz Chua, R.; Trefzer, T.; Kahn, N.C.; Schneider, M.A.; Muley, T.; Winter, H.; Meister, M.; Veith, C.; Boots, A.W.; et al. SARS-COV-2 receptor ace2 and tmprss2 are primarily expressed in bronchial transient secretory cells. EMBO J. 2020, 39, e105114. [Google Scholar] [CrossRef] [PubMed]
- Abassi, Z.A.; Skorecki, K.; Heyman, S.N.; Kinaneh, S.; Armaly, Z. COVID-19 infection and mortality-a physiologist’s perspective enlightening clinical features and plausible interventional strategies. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2020, 318, L1020–L1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papa, G.; Mallery, D.L.; Albecka, A.; Welch, L.G.; Cattin-Ortolá, J.; Luptak, J.; Paul, D.; McMahon, H.T.; Goodfellow, I.G.; Carter, A.; et al. Furin cleavage of SARS-COV-2 spike promotes but is not essential for infection and cell-cell fusion. PLoS Pathog. 2021, 17, e1009246. [Google Scholar] [CrossRef]
- Pinto, B.G.G.; Oliveira, A.E.R.; Singh, Y.; Jimenez, L.; Gonçalves, A.N.A.; Ogava, R.L.T.; Creighton, R.; Schatzmann Peron, J.P.; Nakaya, H.I. Ace2 expression is increased in the lungs of patients with comorbidities associated with severe COVID-19. J. Infect. Dis. 2020, 222, 556–563. [Google Scholar] [CrossRef]
- Zhang, L.; Han, X.; Shi, Y. Comparative analysis of SARS-COV-2 receptor ace2 expression in multiple solid tumors and matched non-diseased tissues. Infect. Genet. Evol. 2020, 85, 104428. [Google Scholar] [CrossRef] [PubMed]
- van Dam, P.A.; Huizing, M.; Mestach, G.; Dierckxsens, S.; Tjalma, W.; Trinh, X.B.; Papadimitriou, K.; Altintas, S.; Vermorken, J.; Vulsteke, C.; et al. SARS-COV-2 and cancer: Are they really partners in crime? Cancer Treat. Rev. 2020, 89, 102068. [Google Scholar] [CrossRef]
- Samad, A.; Jafar, T.; Rafi, J.H. Identification of angiotensin-converting enzyme 2 (ace2) protein as the potential biomarker in SARS-COV-2 infection-related lung cancer using computational analyses. Genomics 2020, 112, 4912–4923. [Google Scholar] [CrossRef]
- Feng, Y.; Wan, H.; Liu, J.; Zhang, R.; Ma, Q.; Han, B.; Xiang, Y.; Che, J.; Cao, H.; Fei, X.; et al. The angiotensin-converting enzyme 2 in tumor growth and tumor-associated angiogenesis in non-small cell lung cancer. Oncol. Rep. 2010, 23, 941–948. [Google Scholar]
- Yamaguchi, M.; Hirai, S.; Sumi, T.; Tanaka, Y.; Tada, M.; Nishii, Y.; Hasegawa, T.; Uchida, H.; Yamada, G.; Watanabe, A.; et al. Angiotensin-converting enzyme 2 is a potential therapeutic target for egfr-mutant lung adenocarcinoma. Biochem. Biophys. Res. Commun. 2017, 487, 613–618. [Google Scholar] [CrossRef]
- Teng, J.P.; Yang, Z.Y.; Zhu, Y.M.; Ni, D.; Zhu, Z.J.; Li, X.Q. Gemcitabine and cisplatin for treatment of lung cancer in vitro and vivo. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3819–3825. [Google Scholar] [PubMed]
- Krishnamurthy, S.; Lockey, R.F.; Kolliputi, N. Soluble ace2 as a potential therapy for COVID-19. Am. J. Physiol. Cell Physiol. 2021, 320, C279–C281. [Google Scholar] [CrossRef]
- Yeung, M.L.; Teng, J.L.L.; Jia, L.; Zhang, C.; Huang, C.; Cai, J.P.; Zhou, R.; Chan, K.H.; Zhao, H.; Zhu, L.; et al. Soluble ace2-mediated cell entry of SARS-COV-2 via interaction with proteins related to the renin-angiotensin system. Cell 2021, 184, 2212–2228. [Google Scholar] [CrossRef]
- BE71030031000; Biobank@UZA, Belgian Virtual Tumourbank Funded by the National Cancer Plan [BIORESOURCE].
- van Dam, P.; Huizing, M.; Roelant, E.; Hotterbeekx, A.; De Winter, F.H.R.; Kumar-Singh, S.; Moons, P.; Amajoud, Z.; Vulsteke, C.; Croes, L.; et al. Immunoglobin g/total antibody testing for SARS-COV-2: A prospective cohort study of ambulatory patients and health care workers in two belgian oncology units comparing three commercial tests. Eur. J. Cancer 2021, 148, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Ponten, F.; Jirstrom, K.; Uhlen, M. The human protein atlas-A tool for pathology. J. Pathol. 2008, 216, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Hoadley, K.A.; Yau, C.; Hinoue, T.; Wolf, D.M.; Lazar, A.J.; Drill, E.; Shen, R.; Taylor, A.M.; Cherniack, A.D.; Thorsson, V.; et al. Cell-of-origin patterns dominate the molecular classification of 10,000 tumors from 33 types of cancer. Cell 2018, 173, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cbioportal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cbio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Addeo, A.; Friedlaender, A. Cancer and COVID-19: Unmasking their ties. Cancer Treat. Rev. 2020, 88, 102041. [Google Scholar] [CrossRef]
- de Azambuja, E.; Brandão, M.; Wildiers, H.; Laenen, A.; Aspeslagh, S.; Fontaine, C.; Collignon, J.; Lybaert, W.; Verheezen, J.; Rutten, A.; et al. Impact of solid cancer on in-hospital mortality overall and among different subgroups of patients with COVID-19: A nationwide, population-based analysis. ESMO Open 2020, 5, e000947. [Google Scholar] [CrossRef]
- Luo, J.; Rizvi, H.; Preeshagul, I.R.; Egger, J.V.; Hoyos, D.; Bandlamudi, C.; McCarthy, C.G.; Falcon, C.J.; Schoenfeld, A.J.; Arbour, K.C.; et al. COVID-19 in patients with lung cancer. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. ESMO 2020, 31, 1386–1396. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.A.; Gay, C.M.; Ramkumar, K.; Cargill, K.R.; Cardnell, R.J.; Nilsson, M.B.; Heeke, S.; Park, E.M.; Kundu, S.T.; Diao, L.; et al. Lung cancer models reveal SARS-COV-2-induced emt contributes to COVID-19 pathophysiology. bioRxiv 2021. [Google Scholar] [CrossRef]
- Venkataraman, T.; Frieman, M.B. The role of epidermal growth factor receptor (egfr) signaling in sars coronavirus-induced pulmonary fibrosis. Antivir. Res. 2017, 143, 142–150. [Google Scholar] [CrossRef]
- Foote, M.B.; White, J.R.; Jee, J.; Argilés, G.; Wan, J.C.M.; Rousseau, B.; Pessin, M.S.; Diaz, L.A., Jr. Association of antineoplastic therapy with decreased SARS-COV-2 infection rates in patients with cancer. JAMA Oncol. 2021, 7, 1686–1691. [Google Scholar] [CrossRef]
- Leonetti, A.; Facchinetti, F.; Zielli, T.; Brianti, E.; Tiseo, M. COVID-19 in lung cancer patients receiving alk/ros1 inhibitors. Eur. J. Cancer 2020, 132, 122–124. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Rizvi, H.; Egger, J.V.; Preeshagul, I.R.; Wolchok, J.D.; Hellmann, M.D. Impact of pd-1 blockade on severity of COVID-19 in patients with lung cancers. Cancer Discov. 2020, 10, 1121–1128. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (n = 107) | n (%) |
|---|---|
| ≤65 years | 55 (51.4) |
| >65 years | 52 (48.6) |
| Gender (n = 107) | |
| Male | 67 (62.6) |
| Female | 40 (37.4) |
| T-classification (n = 106) | |
| T1 | 38 (35.8) |
| T2 | 37 (34.9) |
| T3 | 23 (21.7) |
| T4 | 8 (7.5) |
| N-classification (n = 107) | |
| N0 | 79 (73.8) |
| N1 | 17 (15.9) |
| N2 | 11 (10.3) |
| M-classification (n = 107) | |
| M0 | 100 (93.5) |
| M1 | 7 (6.5) |
| Tumor Stage (n = 106) | |
| I | 49 (46.2) |
| II | 29 (27.4) |
| III | 21 (19.8) |
| IV | 7 (6.6) |
| Subtype (n = 107) | |
| AC | 100 (93.5) |
| SQ | 7 (6.5) |
| Differentiation (n = 97) | |
| Poor | 30 (30.9) |
| Moderate | 37 (38.1) |
| Strong | 30 (30.9) |
| Smoker (n = 89) | |
| No | 8 (9) |
| Yes | 81 (91.0) |
| Neoadjuvant Therapy (n = 107) | |
| No | 85 (79.4) |
| Yes | 22 (20.6) |
| EGFR Mutation (n = 56) | |
| No | 47 (83.9) |
| Yes | 9 (16.1) |
| KRAS Mutation (n = 22) | |
| No | 13 (59.1) |
| Yes | 9 (40.9) |
| PD-L1 (n = 23) | |
| ≤1% | 13 (56.5) |
| >1% | 10 (43.5) |
| ACE2 (n = 107) | |
| IRS 0 | 86 (80.4) |
| IRS 1 | 10 (9.3) |
| IRS 2 | 10 (9.3) |
| IRS 3 | 1 (0.9) |
| TMPRSS2 (n = 38) | |
| IRS 0 | 15 (39.5) |
| IRS 1 | 5 (13.2) |
| IRS 2 | 11 (28.9) |
| IRS 3 | 7 (18.4) |
| FURIN (n = 38) | |
| IRS 0 | 10 (26.3) |
| IRS 1 | 6 (15.8) |
| IRS 2 | 20 (52.6) |
| IRS 3 | 2 (5.3) |
| Membranous ACE2 | Membranous TMPRSS2 | Membranous Furin | |||||||
|---|---|---|---|---|---|---|---|---|---|
| IRS 0 (%) | IRS 1–3 (%) | p-Value | IRS 0 (%) | IRS 1–3 (%) | p-Value | IRS 0 (%) | IRS 1–3 (%) | p-Value | |
| Age (n = 107, n = 38) | |||||||||
| ≤65 | 45 (52.3) | 10 (47.6) | 0.443 | 17 (51.5) | 2 (40) | 0.631 | 7 (53.8) | 12 (48.0) | 0.732 |
| >65 | 65 (47.7) | 11 (52.4) | 16 (48.5) | 3 (60) | 6 (46.2) | 13 (52.0) | |||
| Gender (n = 107, n = 38) | |||||||||
| Male | 56 (65.1) | 11 (52.4) | 0.320 | 17 (51.5) | 3 (60) | 0.723 | 7 (53.8) | 13 (52.0) | 0.914 |
| Female | 30 (34.9) | 10 (47.6) | 16 (48.5) | 2 (40) | 6 (46.2) | 12 (48.0) | |||
| T-classification (n = 106, n = 37) | |||||||||
| T1 | 32 (37.6) | 6 (28.6) | 0.774 | 13 (40.6) | 1 (20.0) | 0.227 | 6 (50.0) | 8 (32.0) | 0.441 |
| T2 | 30 (35.3) | 7 (33.3) | 8 (25.0) | 1 (20.0) | 1 (8.3) | 8 (32.0) | |||
| T3 | 17 (20) | 6 (28.6) | 6 (18.8) | 3 (60.0) | 3 (25.0) | 6 (24.0) | |||
| T4 | 6 (7.1) | 2 (9.5) | 5 (15.6) | 0 (0.0) | 2 (16.7) | 3 (12.0) | |||
| N-classification (n = 107, n = 38) | |||||||||
| N0 | 62 (72.1) | 17 (81) | 0.608 | 27 (81.8) | 5 (100.0) | 0.583 | 11 (84.6) | 21 (84.0) | 0.830 |
| N1 | 14 (16.3) | 3 (14.3) | 4 (12.1) | 0 (0.0) | 1 (7.7) | 3 (12.0) | |||
| N2 | 10 (11.6) | 1 (4.8) | 2 (6.1) | 0 (0.0) | 1 (7.7) | 1(4.0) | |||
| M-classfication (n = 107, n = 38) | |||||||||
| M0 | 80 (93.0) | 20 (95.2) | 1.000 | 31 (93.9) | 5 (100.0) | 0.572 | 12 (92.3) | 24 (96.0) | 0.629 |
| M1 | 6 (7.0) | 1 (4.8) | 2 (6.1) | 0 (0.0) | 1 (7.7) | 1 (4.0) | |||
| Tumor Stage (n = 106, n = 37)) | |||||||||
| I | 41 (48.2) | 8 (38.1) | 0.120 | 12 (37.5) | 2 (40) | 0.688 | 4 (33.3) | 10 (40.0) | 0.821 |
| II | 19 (22.4) | 10 (47.6) | 13 (40.6) | 3 (60) | 6 (50.0) | 10 (40.0) | |||
| III | 19 (22.4) | 2 (9.5) | 5 (15.6) | 0 (0.0) | 1 (8.3) | 4 (16.0) | |||
| IV | 6 (7.1) | 1 (4.8) | 2 (6.3) | 0 (0.0 | 1 (8.3) | 1 (4.0) | |||
| Subtype (n = 107, n = 38)) | |||||||||
| AC | 79 (91.9) | 21 (100) | 0.341 | 30 (90.9) | 3 (60.0) | 0.057 | 9 (69.2) | 24 (96.0) | 0.021 |
| SQ | 7 (8.1) | 0 (0.0) | 3 (9.1) | 2 (40.0) | 4 (30.8) | 1 (4.0) | |||
| Differentiation (n = 97, n = 30)) | |||||||||
| Poor | 25 (31.6) | 5 (27.8) | 0.490 | 7 (28.0) | 2 (40.0) | 0.603 | 4 (36.4) | 5 (26.3) | 0.786 |
| Moderate | 28 (35.4) | 9 (50.0) | 14 (56.0) | 3 (60.0) | 6 (54.4) | 11 (57.9) | |||
| Strong | 26 (32.9) | 4 (22.2) | 4 (16.0) | 0 (0.0) | 1 (9.1) | 3 (15.8) | |||
| Smoker (n = 89, n = 32) | |||||||||
| No | 6 (8.7) | 2 (10) | 1.000 | 5 (17.2) | 0 (0.0) | 0.434 | 1 (10.0) | 4 (18.2) | 0.555 |
| Yes | 63 (91.3) | 18 (90) | 24 (82.8) | 3 (100) | 9 (90.0) | 18 (81.8) | |||
| Neoadjuvant Therapy (n = 107, n = 38) | |||||||||
| No | 66 (76.7) | 19 (90.5) | 0.232 | 27 (81.8) | 4 (80.0) | 0.922 | 10 (76.9) | 21 (84.0) | 0.593 |
| Yes | 20 (23.3) | 2 (9.5) | 6 (18.2) | 1 (20.0) | 3 (23.1) | 4 (16.0) | |||
| EGFR Mutation (n = 56, n = 26) | |||||||||
| No | 40 (88.9) | 7 (63.6) | 0.063 | 18 (75) | 2 (100.0) | 0.420 | 9 (81.8) | 11 (73.3) | 0.612 |
| Yes | 5 (11.1) | 4 (36.4) | 6 (25) | 0 (0.0) | 2 (18.2) | 4 (26.7) | |||
| KRAS Mutation (n = 22, n = 19)) | |||||||||
| No | 9 (50.0) | 4 (100) | 0.115 | 10 (58.8) | 2 (100.0) | 0.253 | 6 (66.7) | 6 (60.0) | 0.764 |
| Yes | 9 (50.0) | 0 (0.0) | 7 (41.2) | 0 (0.0) | 3 (33.3) | 4 (40.0) | |||
| PD-L1 (n = 23, n = 20) | |||||||||
| ≤1% | 10 (52.6) | 3 (75.0) | 0.604 | 11 (61.1) | 1 (50.0) | 0.761 | 6 (60.0) | 6 (60.0 | 1.000 |
| >1% | 9 (47.4) | 1 (25.0) | 7 (38.9) | 1 (50.0) | 4 (40.0) | 4 (60.0) | |||
| TMPRSS2 (n = 38) | |||||||||
| IRS 0 | 12 (70.6) | 3 (14.3) | 0.005 | N/A | N/A | 6 (46.2) | 9 (36.0) | 0.618 | |
| IRS 1 | 1 (5.9) | 4 (19.0) | N/A | N/A | 2 (15.4) | 3 (12.0) | |||
| IRS 2 | 2 (11.8) | 9 (42.9) | N/A | N/A | 2 (15.4) | 9 (36.0) | |||
| IRS 3 | 2 (11.8) | 5 (23.8) | 3 (23.1) | 4 (16.0) | |||||
| FURIN (n = 38) | |||||||||
| IRS 0 | 8 (47.1) | 1 (4.8) | 0.002 | 8 (24.2) | 2 (40.0) | 0.641 | N/A | N/A | |
| IRS 1 | 0 (0.0) | 7 (33.3) | 6 (18.2) | 0 (0.0) | N/A | N/A | |||
| IRS 2 | 9 (52.9) | 11 (52.4) | 17 (51.5) | 3 (60.0) | N/A | N/A | |||
| IRS 3 | 0 (0.0) | 2 (9.5) | 2 (6.1) | 0 (0.0) | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deben, C.; Le Compte, M.; Siozopoulou, V.; Lambrechts, H.; Hermans, C.; Lau, H.W.; Huizing, M.; Lamote, K.; Hendriks, J.M.H.; Van Dam, P.; et al. Expression of SARS-CoV-2-Related Surface Proteins in Non-Small-Cell Lung Cancer Patients and the Influence of Standard of Care Therapy. Cancers 2022, 14, 4074. https://doi.org/10.3390/cancers14174074
Deben C, Le Compte M, Siozopoulou V, Lambrechts H, Hermans C, Lau HW, Huizing M, Lamote K, Hendriks JMH, Van Dam P, et al. Expression of SARS-CoV-2-Related Surface Proteins in Non-Small-Cell Lung Cancer Patients and the Influence of Standard of Care Therapy. Cancers. 2022; 14(17):4074. https://doi.org/10.3390/cancers14174074
Chicago/Turabian StyleDeben, Christophe, Maxim Le Compte, Vasiliki Siozopoulou, Hilde Lambrechts, Christophe Hermans, Ho Wa Lau, Manon Huizing, Kevin Lamote, Jeroen M. H. Hendriks, Peter Van Dam, and et al. 2022. "Expression of SARS-CoV-2-Related Surface Proteins in Non-Small-Cell Lung Cancer Patients and the Influence of Standard of Care Therapy" Cancers 14, no. 17: 4074. https://doi.org/10.3390/cancers14174074
APA StyleDeben, C., Le Compte, M., Siozopoulou, V., Lambrechts, H., Hermans, C., Lau, H. W., Huizing, M., Lamote, K., Hendriks, J. M. H., Van Dam, P., Pauwels, P., Smits, E. L. J., Peeters, M., & Lardon, F. (2022). Expression of SARS-CoV-2-Related Surface Proteins in Non-Small-Cell Lung Cancer Patients and the Influence of Standard of Care Therapy. Cancers, 14(17), 4074. https://doi.org/10.3390/cancers14174074