
Exploratory Circular RNA Profiling in Adrenocortical Tumors
 , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Collection and Ethics Approval
2.2. Sample Processing and RNA Isolation
2.3. RNase R Treatment, Polyadenylation, and Poly(A)+ RNA Depletion (RPAD)
2.4. Next-Generation Sequencing (NGS)
2.5. Bioinformatics Analysis of circRNA
2.6. CircRNA Validation by RT-qPCR
2.7. In Silico Prediction of Potential circRNA-miRNA Interactions
2.8. miRNA Analysis by RT-qPCR
3. Results
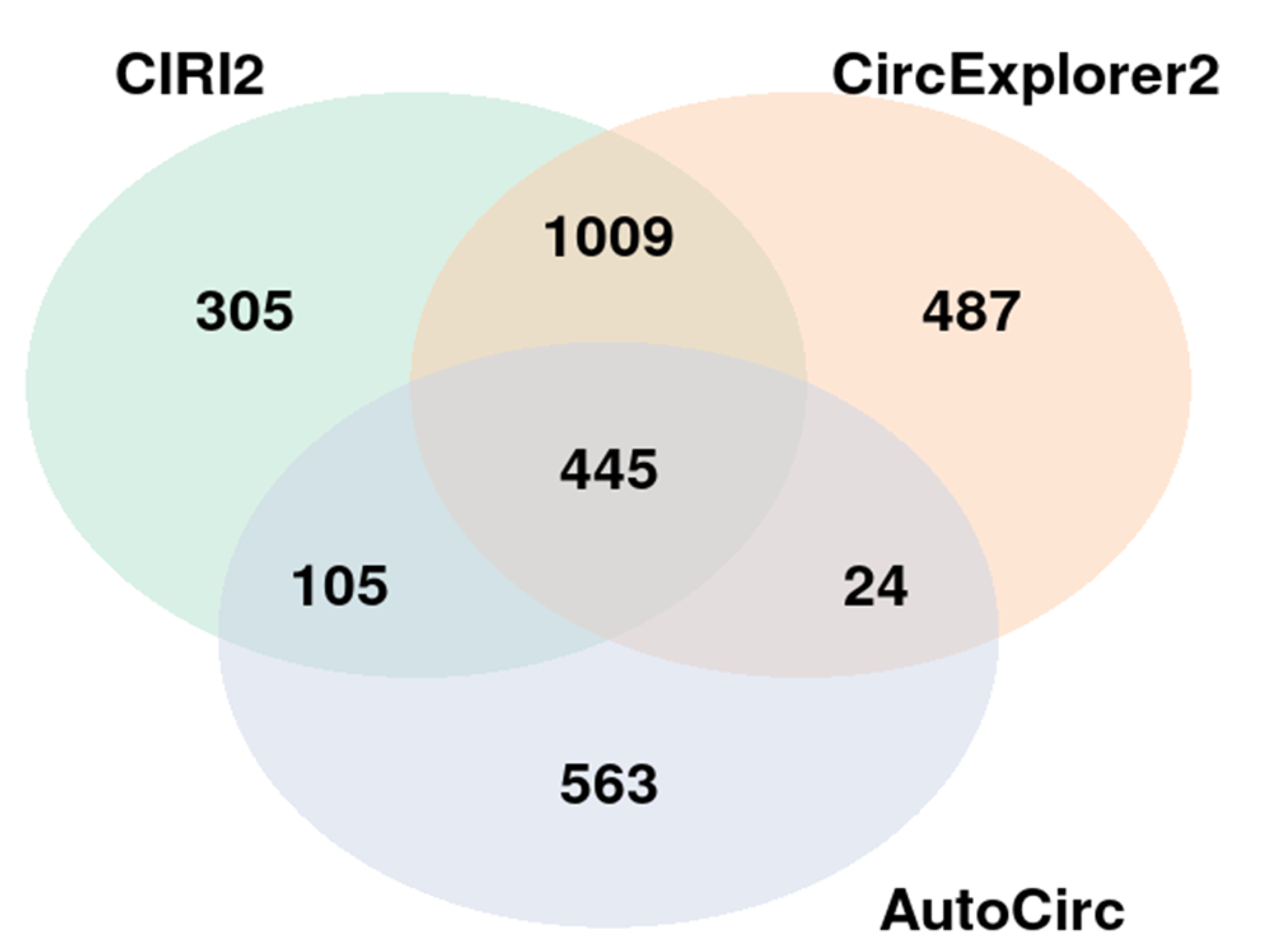
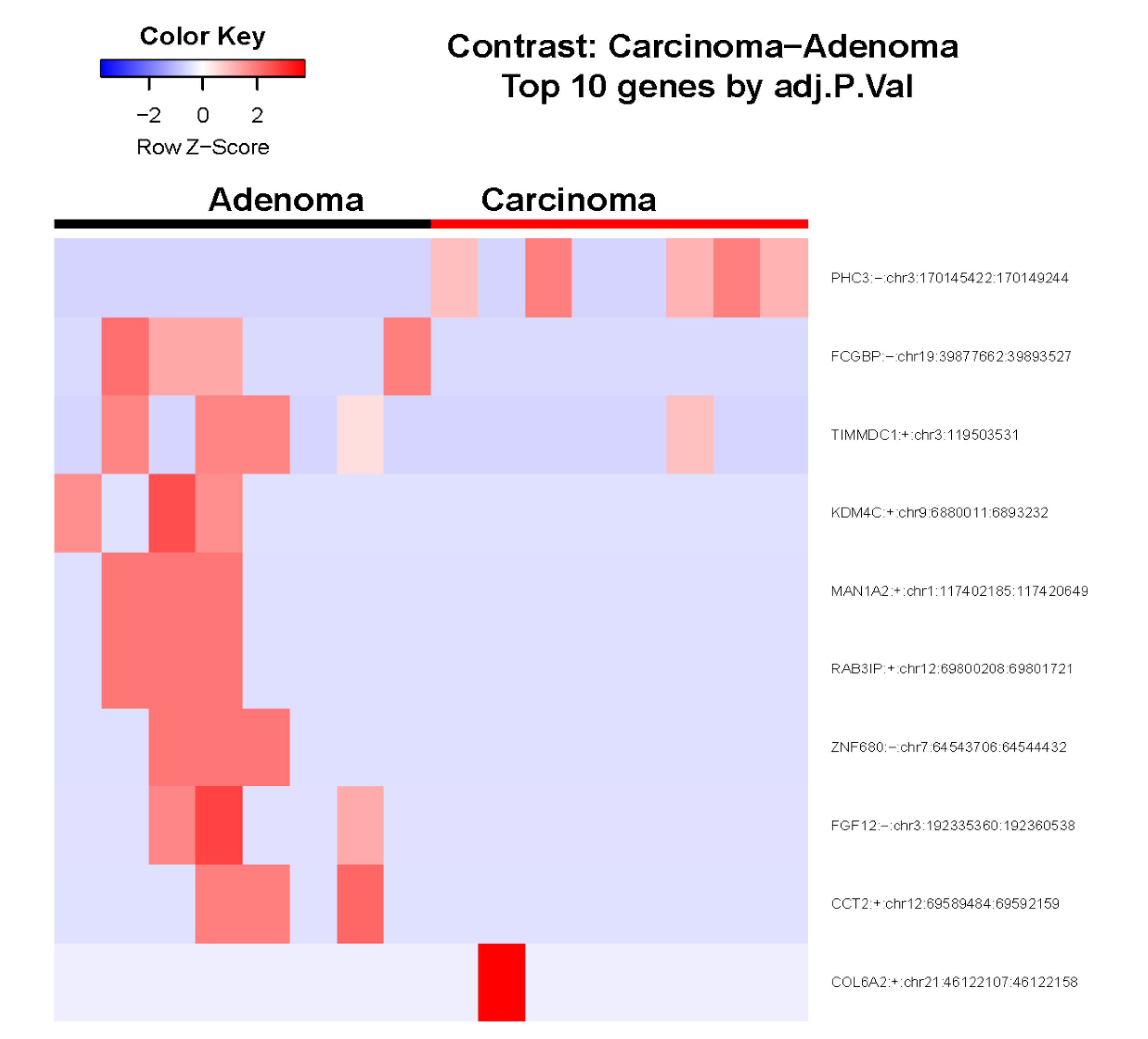
3.1. Identification of Differentially Expressed circRNAs
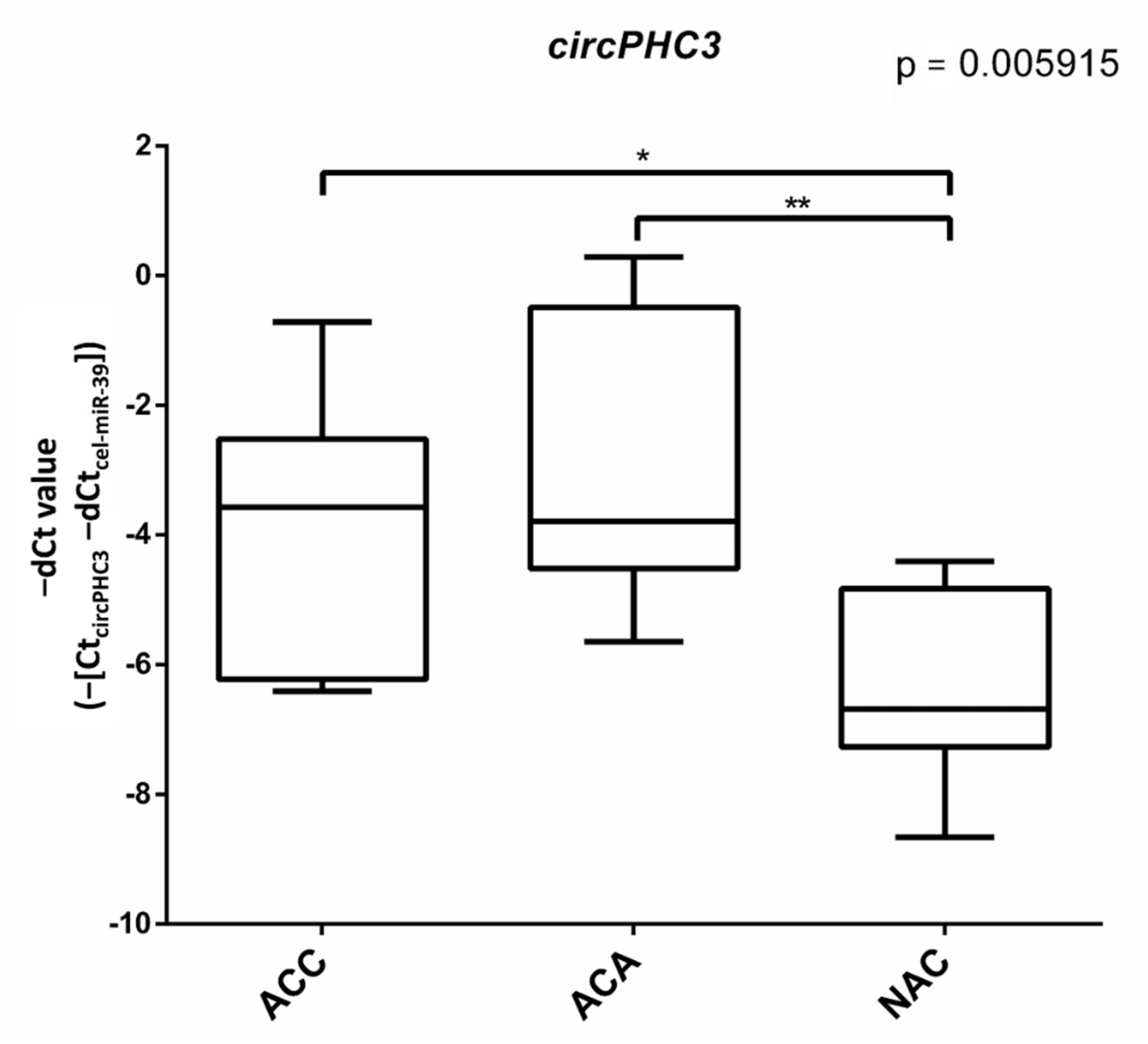
3.2. Validation of Differentially Expressed circRNAs
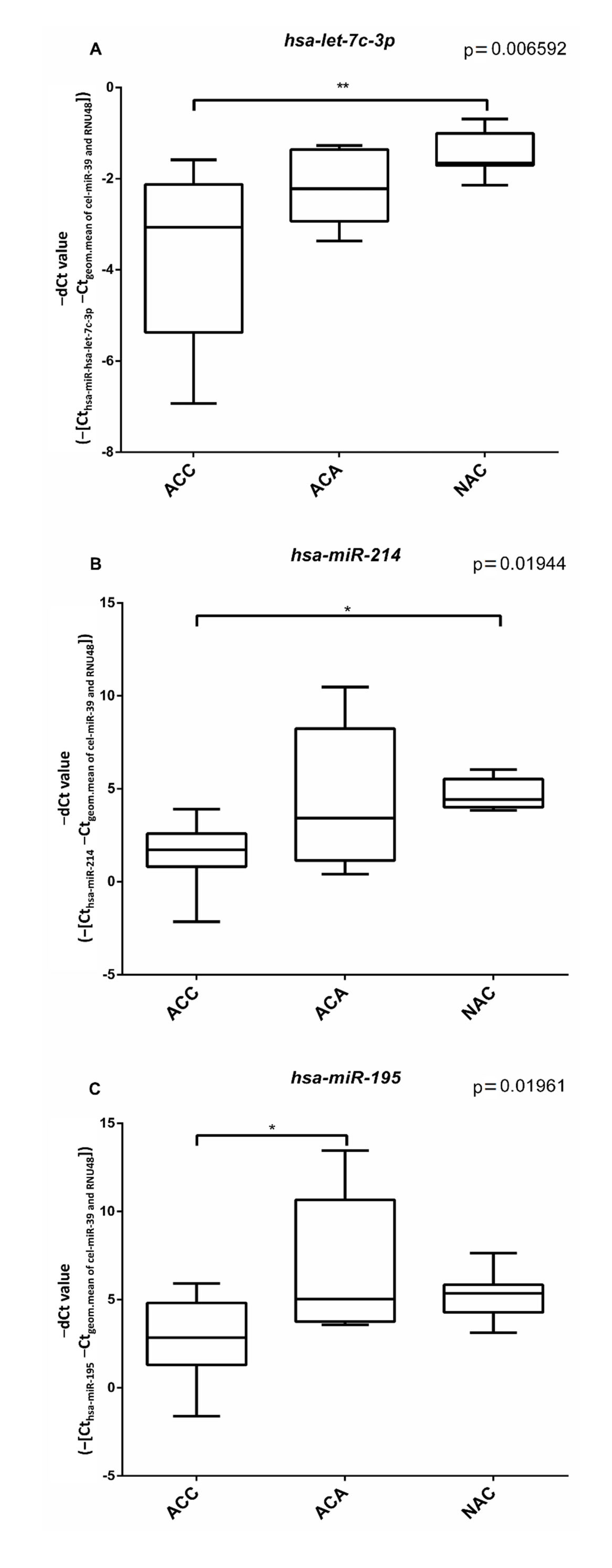
3.3. In Silico Prediction of Potential circRNA-microRNA Interactions (Sponging)
3.4. Putative miRNA Interaction with circPHC3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bovio, S.; Cataldi, A.; Reimondo, G.; Sperone, P.; Novello, S.; Berruti, A.; Borasio, P.; Fava, C.; Dogliotti, L.; Scagliotti, G.V.; et al. Prevalence of Adrenal Incidentaloma in a Contemporary Computerized Tomography Series. J. Endocrinol. Investig. 2006, 29, 298–302. [Google Scholar] [CrossRef]
- Libé, R.; Borget, I.; Ronchi, C.L.; Zaggia, B.; Kroiss, M.; Kerkhofs, T.; Bertherat, J.; Volante, M.; Quinkler, M.; Chabre, O.; et al. Prognostic Factors in Stage III–IV Adrenocortical Carcinomas (ACC): An European Network for the Study of Adrenal Tumor (ENSAT) Study. Ann. Oncol. 2015, 26, 2119–2125. [Google Scholar] [CrossRef] [PubMed]
- Fassnacht, M.; Dekkers, O.M.; Else, T.; Baudin, E.; Berruti, A.; De Krijger, R.R.; Haak, H.R.; Mihai, R.; Assie, G.; Terzolo, M. European Society of Endocrinology Clinical Practice Guidelines on the Management of Adrenocortical Carcinoma in Adults, in Collaboration with the European Network for the Study of Adrenal Tumors. Eur. J. Endocrinol. 2018, 179, G1–G46. [Google Scholar] [CrossRef] [PubMed]
- Fassnacht, M.; Arlt, W.; Bancos, I.; Dralle, H.; Newell-Price, J.; Sahdev, A.; Tabarin, A.; Terzolo, M.; Tsagarakis, S.; Dekkers, O.M. Management of Adrenal Incidentalomas: European Society of Endocrinology Clinical Practice Guideline in Collaboration with the European Network for the Study of Adrenal Tumors. Eur. J. Endocrinol. 2016, 175, G1–G34. [Google Scholar] [CrossRef] [PubMed]
- Terzolo, M.; Angeli, A.; Fassnacht, M.; Daffara, F.; Tauchmanova, L.; Conton, P.A.; Rossetto, R.; Buci, L.; Sperone, P.; Grossrubatscher, E.; et al. Adjuvant Mitotane Treatment for Adrenocortical Carcinoma. N. Engl. J. Med. 2007, 356, 2372–2380. [Google Scholar] [CrossRef]
- Altieri, B.; Ronchi, C.L.; Kroiss, M.; Fassnacht, M. Next-Generation Therapies for Adrenocortical Carcinoma. Best Pract. Res. Clin. Endocrinol. Metab. 2020, 34, 101434. [Google Scholar] [CrossRef]
- Abiven, G.; Coste, J.; Groussin, L.; Anract, P.; Tissier, F.; Legmann, P.; Dousset, B.; Bertagna, X.; Bertherat, J. Clinical and Biological Features in the Prognosis of Adrenocortical Cancer: Poor Outcome of Cortisol-Secreting Tumors in a Series of 202 Consecutive Patients. J. Clin. Endocrinol. Metab. 2006, 91, 2650–2655. [Google Scholar] [CrossRef] [PubMed]
- Viëtor, C.L.; Creemers, S.G.; van Kemenade, F.J.; van Ginhoven, T.M.; Hofland, L.J.; Feelders, R.A. How to Differentiate Benign from Malignant Adrenocortical Tumors? Cancers 2021, 13, 4383. [Google Scholar] [CrossRef]
- Mizdrak, M.; Kurir, T.T.; Božić, J. The Role of Biomarkers in Adrenocortical Carcinoma: A Review of Current Evidence and Future Perspectives. Biomedicines 2021, 9, 174. [Google Scholar] [CrossRef]
- Mete, O.; Gucer, H.; Kefeli, M.; Asa, S.L. Diagnostic and Prognostic Biomarkers of Adrenal Cortical Carcinoma. Am. J. Surg. Pathol. 2018, 42, 201–213. [Google Scholar] [CrossRef]
- Decmann, A.; Perge, P.; Turai, P.I.; Patócs, A.; Igaz, P. Non-Coding RNAs in Adrenocortical Cancer: From Pathogenesis to Diagnosis. Cancers 2020, 12, 461. [Google Scholar] [CrossRef]
- Lerario, A.M.; Moraitis, A.; Hammer, G.D. Genetics and Epigenetics of Adrenocortical Tumors. Mol. Cell. Endocrinol. 2014, 386, 67. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-Coding RNAs and Their Integrated Networks. J. Integr. Bioinform. 2019, 16. [Google Scholar] [CrossRef]
- Nigro, J.M.; Cho, K.R.; Fearon, E.R.; Kern, S.E.; Ruppert, J.M.; Oliner, J.D.; Kinzler, K.W.; Vogelstein, B. Scrambled Exons. Cell 1991, 64, 607–613. [Google Scholar] [CrossRef]
- Ebbesen, K.K.; Hansen, T.B.; Kjems, J. Insights into Circular RNA Biology. RNA Biol. 2017, 14, 1035. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, L.; Chen, L.L. The Biogenesis, Functions, and Challenges of Circular RNAs. Mol. Cell 2018, 71, 428–442. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The Biogenesis, Biology and Characterization of Circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef]
- Chen, L.L. The Biogenesis and Emerging Roles of Circular RNAs. Nat. Rev. Mol. Cell Biol. 2016, 17, 205–211. [Google Scholar] [CrossRef]
- Jeck, W.R.; Sharpless, N.E. Detecting and Characterizing Circular RNAs. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Hansen, T.B.; Venø, M.T.; Kjems, J. Circular RNAs in Cancer: Opportunities and Challenges in the Field. Oncogene 2018, 37, 555–565. [Google Scholar] [CrossRef] [Green Version]
- Greene, J.; Baird, A.M.; Casey, O.; Brady, L.; Blackshields, G.; Lim, M.; O’Brien, O.; Gray, S.G.; McDermott, R.; Finn, S.P. Circular RNAs Are Differentially Expressed in Prostate Cancer and Are Potentially Associated with Resistance to Enzalutamide. Sci. Rep. 2019, 9, 10739. [Google Scholar] [CrossRef] [PubMed]
- Greene, J.; Baird, A.M.; Lim, M.; Flynn, J.; McNevin, C.; Brady, L.; Sheils, O.; Gray, S.G.; McDermott, R.; Finn, S.P. Differential CircRNA Expression Signatures May Serve as Potential Novel Biomarkers in Prostate Cancer. Front. Cell Dev. Biol. 2021, 9, 344. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Li, X.; Xu, M.; Zhang, L.; Zuo, H.; Xin, Y.; Zhang, L.; Gong, P. Differential Expression and Bioinformatics Analysis of CircRNA in Non-Small Cell Lung Cancer. Front. Genet. 2020, 11, 586814. [Google Scholar] [CrossRef]
- Jahani, S.; Nazeri, E.; Majidzadeh-A, K.; Jahani, M.; Esmaeili, R. Circular RNA; a New Biomarker for Breast Cancer: A Systematic Review. J. Cell. Physiol. 2020, 235, 5501–5510. [Google Scholar] [CrossRef] [PubMed]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs Are Abundant, Conserved, and Associated with ALU Repeats. RNA 2013, 19, 141. [Google Scholar] [CrossRef]
- Pandey, P.R.; Rout, P.K.; Das, A.; Gorospe, M.; Panda, A.C. RPAD (RNase R Treatment, Polyadenylation, and Poly(A)+ RNA Depletion) Method to Isolate Highly Pure Circular RNA. Methods 2019, 155, 41–48. [Google Scholar] [CrossRef]
- Panda, A.C.; De, S.; Grammatikakis, I.; Munk, R.; Yang, X.; Piao, Y.; Dudekula, D.B.; Abdelmohsen, K.; Gorospe, M. High-Purity Circular RNA Isolation Method (RPAD) Reveals Vast Collection of Intronic CircRNAs. Nucleic Acids Res. 2017, 45, e116. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows-Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Chen, H.; Boutros, P.C. VennDiagram: A Package for the Generation of Highly-Customizable Venn and Euler Diagrams in R. BMC Bioinform. 2011, 12, 35. [Google Scholar] [CrossRef]
- Zhou, C.; Molinie, B.; Daneshvar, K.; Pondick, J.V.; Wang, J.; Wittenberghe, N.O.V.; Xing, Y.; Giallourakis, C.C.; Mullen, A.C. Genome-Wide Maps of M6A CircRNAs Identify Widespread and Cell-Type-Specific Methylation Patterns That Are Distinct from MRNAs. Cell Rep. 2017, 20, 2262–2276. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Zhang, J.; Zhao, F. Circular RNA Identification Based on Multiple Seed Matching. Brief. Bioinform. 2018, 19, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.O.; Dong, R.; Zhang, Y.; Zhang, J.L.; Luo, Z.; Zhang, J.; Chen, L.L.; Yang, L. Diverse Alternative Back-Splicing and Alternative Splicing Landscape of Circular RNAs. Genome Res. 2016, 26, 1277–1287. [Google Scholar] [CrossRef] [PubMed]
- Aufiero, S.; Reckman, Y.J.; Tijsen, A.J.; Pinto, Y.M.; Creemers, E.E. CircRNAprofiler: An R-Based Computational Framework for the Downstream Analysis of Circular RNAs. BMC Bioinform. 2020, 21, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Law, C.W.; Chen, Y.; Shi, W.; Smyth, G.K. Voom: Precision Weights Unlock Linear Model Analysis Tools for RNA-Seq Read Counts. Genome Biol. 2014, 15, R29. [Google Scholar] [CrossRef] [PubMed]
- CircTools: Identify and Annotate CircRNAs and Their Interactions with MicroRNAs (MiRNAs) from Large-Scale CLIP-Seq (HITS-CLIP, PAR-CLIP, ICLIP, CLASH) and RNA-Seq Data. Available online: https://starbase.sysu.edu.cn/starbase2/circTools.php (accessed on 20 June 2022).
- Tömböl, Z.; Szabó, P.M.; Molnár, V.; Wiener, Z.; Tölgyesi, G.; Horányi, J.; Riesz, P.; Reismann, P.; Patócs, A.; Likó, I.; et al. Integrative Molecular Bioinformatics Study of Human Adrenocortical Tumors: MicroRNA, Tissue-Specific Target Prediction, and Pathway Analysis. Endocr. Relat. Cancer 2009, 16, 895–906. [Google Scholar] [CrossRef]
- Soon, P.S.H.; Tacon, L.J.; Gill, A.J.; Bambach, C.P.; Sywak, M.S.; Campbell, P.R.; Yeh, M.W.; Wong, S.G.; Clifton-Bligh, R.J.; Robinson, B.G.; et al. MiR-195 and MiR-483-5p Identified as Predictors of Poor Prognosis in Adrenocortical Cancer. Clin. Cancer Res. 2009, 15, 7684–7692. [Google Scholar] [CrossRef]
- Patterson, E.E.; Holloway, A.K.; Weng, J.; Fojo, T.; Kebebew, E. MicroRNA Profiling of Adrenocortical Tumors Reveals MiR-483 as a Marker of Malignancy. Cancer 2011, 117, 1630. [Google Scholar] [CrossRef]
- Özata, D.M.; Caramuta, S.; Velázquez-Fernández, D.; Akçakaya, P.; Xie, H.; Höög, A.; Zedenius, J.; Bäckdahl, M.; Larsson, C.; Lui, W.O. The Role of MicroRNA Deregulation in the Pathogenesis of Adrenocortical Carcinoma. Endocr. Relat. Cancer 2011, 18, 643–655. [Google Scholar] [CrossRef]
- Duregon, E.; Rapa, I.; Votta, A.; Giorcelli, J.; Daffara, F.; Terzolo, M.; Scagliotti, G.V.; Volante, M.; Papotti, M. MicroRNA Expression Patterns in Adrenocortical Carcinoma Variants and Clinical Pathologic Correlations. Hum. Pathol. 2014, 45, 1555–1562. [Google Scholar] [CrossRef]
- Feinmesser, M.; Benbassat, C.; Meiri, E.; Benjamin, H.; Lebanony, D.; Lebenthal, Y.; Vries, L.D.; Drozd, T.; Spector, Y. Specific MicroRNAs Differentiate Adrenocortical Adenomas from Carcinomas and Correlate with Weiss Histopathologic System. Appl. Immunohistochem. Mol. Morphol. 2015, 23, 522–531. [Google Scholar] [CrossRef]
- Zheng, S.; Cherniack, A.D.; Dewal, N.; Moffitt, R.A.; Danilova, L.; Murray, B.A.; Lerario, A.M.; Else, T.; Knijnenburg, T.A.; Ciriello, G.; et al. Comprehensive Pan-Genomic Characterization of Adrenocortical Carcinoma. Cancer Cell 2016, 29, 723. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- CircAtlas. Available online: http://159.226.67.237:8080/new/index.php (accessed on 20 June 2022).
- Lei, B.; Tian, Z.; Fan, W.; Ni, B. Circular RNA: A Novel Biomarker and Therapeutic Target for Human Cancers. Int. J. Med. Sci. 2019, 16, 292. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yang, T.; Xiao, J. Circular RNAs: Promising Biomarkers for Human Diseases. EBioMedicine 2018, 34, 267–274. [Google Scholar] [CrossRef]
- Verduci, L.; Tarcitano, E.; Strano, S.; Yarden, Y.; Blandino, G. CircRNAs: Role in Human Diseases and Potential Use as Biomarkers. Cell Death Dis. 2021, 12, 468. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, K.; Tan, S.; Xin, J.; Yuan, Q.; Xu, H.; Xu, X.; Liang, Q.; Christiani, D.C.; Wang, M.; et al. Circular RNAs in Body Fluids as Cancer Biomarkers: The New Frontier of Liquid Biopsies. Mol. Cancer 2021, 20, 13. [Google Scholar] [CrossRef]
- Li, W.; Liu, R.; Wei, D.; Zhang, W.; Zhang, H.; Huang, W.; Hao, L. Circular RNA Circ-CCAC1 Facilitates Adrenocortical Carcinoma Cell Proliferation, Migration, and Invasion through Regulating the MiR-514a-5p/C22orf46 Axis. Biomed Res. Int. 2020, 2020, 3501451. [Google Scholar] [CrossRef]
- Belmihoub, I.; Silvera, S.; Sibony, M.; Dousset, B.; Legmann, P.; Bertagna, X.; Bertherat, J.; Assié, G. From Benign Adrenal Incidentaloma to Adrenocortical Carcinoma: An Exceptional Random Event. Eur. J. Endocrinol. 2017, 176, K15–K19. [Google Scholar] [CrossRef]
- Kohli, H.S.; Manthri, S.; Jain, S.; Kashyap, R.; Chen, S.; Koritala, T.; Tekin, A.; Adhikari, R.; Tirupathi, R.; Barbaryan, A.; et al. An Adrenocortical Carcinoma Evolving After Nine Years of Latency From a Small Adrenal Incidentaloma. Cureus 2021, 13, e16851. [Google Scholar] [CrossRef]
- Rebielak, M.E.; Wolf, M.R.; Jordan, R.; Oxenberg, J.C. Adrenocortical Carcinoma Arising from an Adrenal Adenoma in a Young Adult Female. J. Surg. Case Rep. 2019, 2019, rjz200. [Google Scholar] [CrossRef]
- Lerario, A.M.; Mohan, D.R.; Hammer, G.D.; Hammer, D. Update on Biology and Genomics of Adrenocortical Carcinomas: Rationale for Emerging Therapies. Endocr. Rev. 2022, 109, bnac012. [Google Scholar] [CrossRef] [PubMed]
- Levine, S.S.; Weiss, A.; Erdjument-Bromage, H.; Shao, Z.; Tempst, P.; Kingston, R.E. The Core of the Polycomb Repressive Complex Is Compositionally and Functionally Conserved in Flies and Humans. Mol. Cell. Biol. 2002, 22, 6070. [Google Scholar] [CrossRef]
- Iwata, S.; Takenobu, H.; Kageyama, H.; Koseki, H.; Ishii, T.; Nakazawa, A.; Tatezaki, S.; Nakagawara, A.; Kamijo, T. Polycomb Group Molecule PHC3 Regulates Polycomb Complex Composition and Prognosis of Osteosarcoma. Cancer Sci. 2010, 101, 1646–1652. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Feng, J.; Lei, L.; Hu, J.; Xia, L.; Wang, J.; Xiang, Y.; Liu, L.; Zhong, S.; Han, L.; et al. Comprehensive Characterization of Tissue-Specific Circular RNAs in the Human and Mouse Genomes. Brief. Bioinform. 2017, 18, 984–992. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Li, N.; Liang, S.; Huang, Q.; Coukos, G.; Zhang, L. Identification of MicroRNAs Regulating Reprogramming Factor LIN28 in Embryonic Stem Cells and Cancer Cells. J. Biol. Chem. 2010, 285, 41961–41971. [Google Scholar] [CrossRef]
- Faria, A.M.; Sbiera, S.; Ribeiro, T.C.; Soares, I.C.; Mariani, B.M.P.; Freire, D.S.; De Sousa, G.R.V.; Lerario, A.M.; Ronchi, C.L.; Deutschbein, T.; et al. Expression of LIN28 and Its Regulatory MicroRNAs in Adult Adrenocortical Cancer. Clin. Endocrinol. 2015, 82, 481–488. [Google Scholar] [CrossRef]
- Cheng, A.M.; Byrom, M.W.; Shelton, J.; Ford, L.P. Antisense Inhibition of Human MiRNAs and Indications for an Involvement of MiRNA in Cell Growth and Apoptosis. Nucleic Acids Res. 2005, 33, 1290–1297. [Google Scholar] [CrossRef]
- Rochow, H.; Franz, A.; Jung, M.; Weickmann, S.; Ralla, B.; Kilic, E.; Stephan, C.; Fendler, A.; Jung, K. Instability of Circular RNAs in Clinical Tissue Samples Impairs Their Reliable Expression Analysis Using RT-QPCR: From the Myth of Their Advantage as Biomarkers to Reality. Theranostics 2020, 10, 9268. [Google Scholar] [CrossRef]
- Munro, S.A.; Lund, S.P.; Pine, P.S.; Binder, H.; Clevert, D.A.; Conesa, A.; Dopazo, J.; Fasold, M.; Hochreiter, S.; Hong, H.; et al. Assessing Technical Performance in Differential Gene Expression Experiments with External Spike-in RNA Control Ratio Mixtures. Nat. Commun. 2014, 5, 5125. [Google Scholar] [CrossRef]
- Szabo, L.; Salzman, J. Detecting Circular RNAs: Bioinformatic and Experimental Challenges. Nat. Rev. Genet. 2016, 17, 679–692. [Google Scholar] [CrossRef] [Green Version]
- Szabo, L.; Morey, R.; Palpant, N.J.; Wang, P.L.; Afari, N.; Jiang, C.; Parast, M.M.; Murry, C.E.; Laurent, L.C.; Salzman, J. Statistically Based Splicing Detection Reveals Neural Enrichment and Tissue-Specific Induction of Circular RNA during Human Fetal Development. Genome Biol. 2015, 16, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cohort/Samples | Mean Age at Sample Taking (Years) | Mean Tumor Size (mm) | Ki-67 (%) | ENSAT Stage | Hormonal Activity |
|---|---|---|---|---|---|
| Discovery ACA (7F, 1M) | 46 | 32 | - | - | 6 cortisol |
| 2 NF | |||||
| Discovery ACC (5F, 3M) | 39 | 119 | 11–18 (2–40) | II:5, III:2; IV:1 | 1 cortisol 4 NF 1 DOC 1 DOC + cortisol + estradiol 1 cortisol + DHEAS |
| Validation ACA (7F, 1M) | 53 | 37 | - | - | 3 NF |
| 5 cortisol | |||||
| Validation ACC (6F, 4M) | 56 | 119 | 18–23 (1–50) | II:1, III:5, IV:4 | 5 NF |
| 4 cortisol 1 cortisol + androgen |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turai, P.I.; Nyirő, G.; Borka, K.; Micsik, T.; Likó, I.; Patócs, A.; Igaz, P. Exploratory Circular RNA Profiling in Adrenocortical Tumors. Cancers 2022, 14, 4313. https://doi.org/10.3390/cancers14174313
Turai PI, Nyirő G, Borka K, Micsik T, Likó I, Patócs A, Igaz P. Exploratory Circular RNA Profiling in Adrenocortical Tumors. Cancers. 2022; 14(17):4313. https://doi.org/10.3390/cancers14174313
Chicago/Turabian StyleTurai, Péter István, Gábor Nyirő, Katalin Borka, Tamás Micsik, István Likó, Attila Patócs, and Peter Igaz. 2022. "Exploratory Circular RNA Profiling in Adrenocortical Tumors" Cancers 14, no. 17: 4313. https://doi.org/10.3390/cancers14174313
APA StyleTurai, P. I., Nyirő, G., Borka, K., Micsik, T., Likó, I., Patócs, A., & Igaz, P. (2022). Exploratory Circular RNA Profiling in Adrenocortical Tumors. Cancers, 14(17), 4313. https://doi.org/10.3390/cancers14174313