
Emergence of MUC1 in Mammals for Adaptation of Barrier Epithelia


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Discovery of MUC1 in Breast Cancer
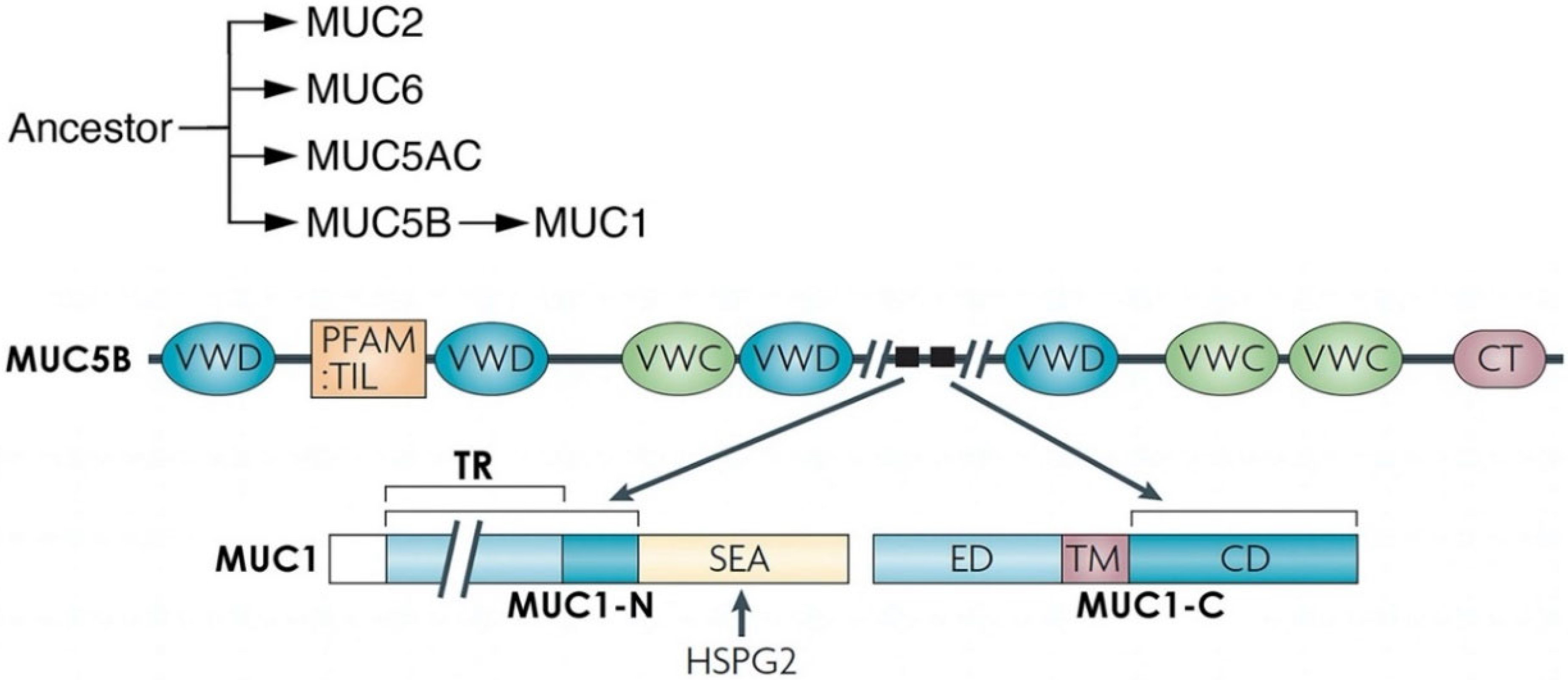
1.2. Evolution of MUC1 in Mammals
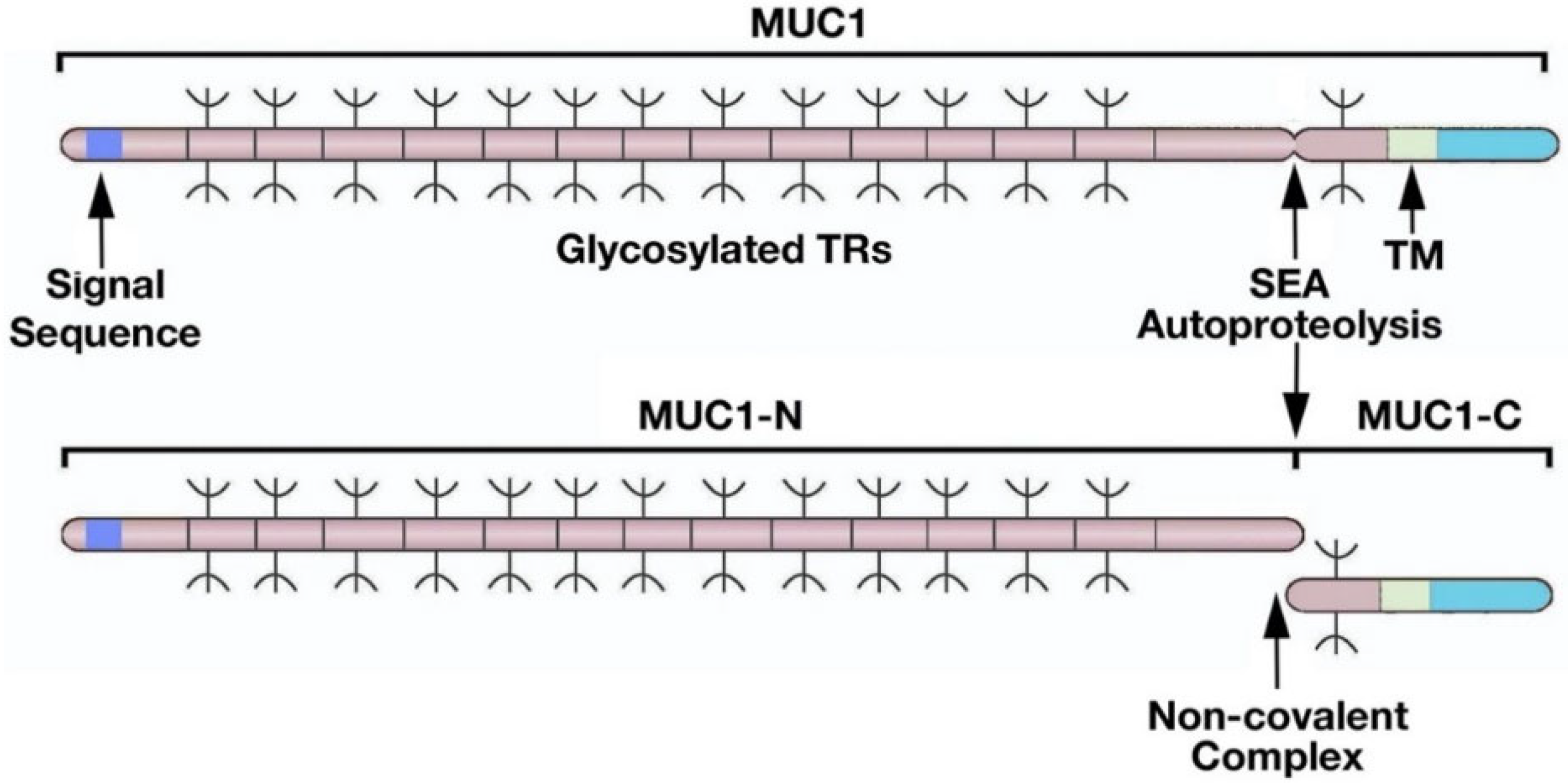
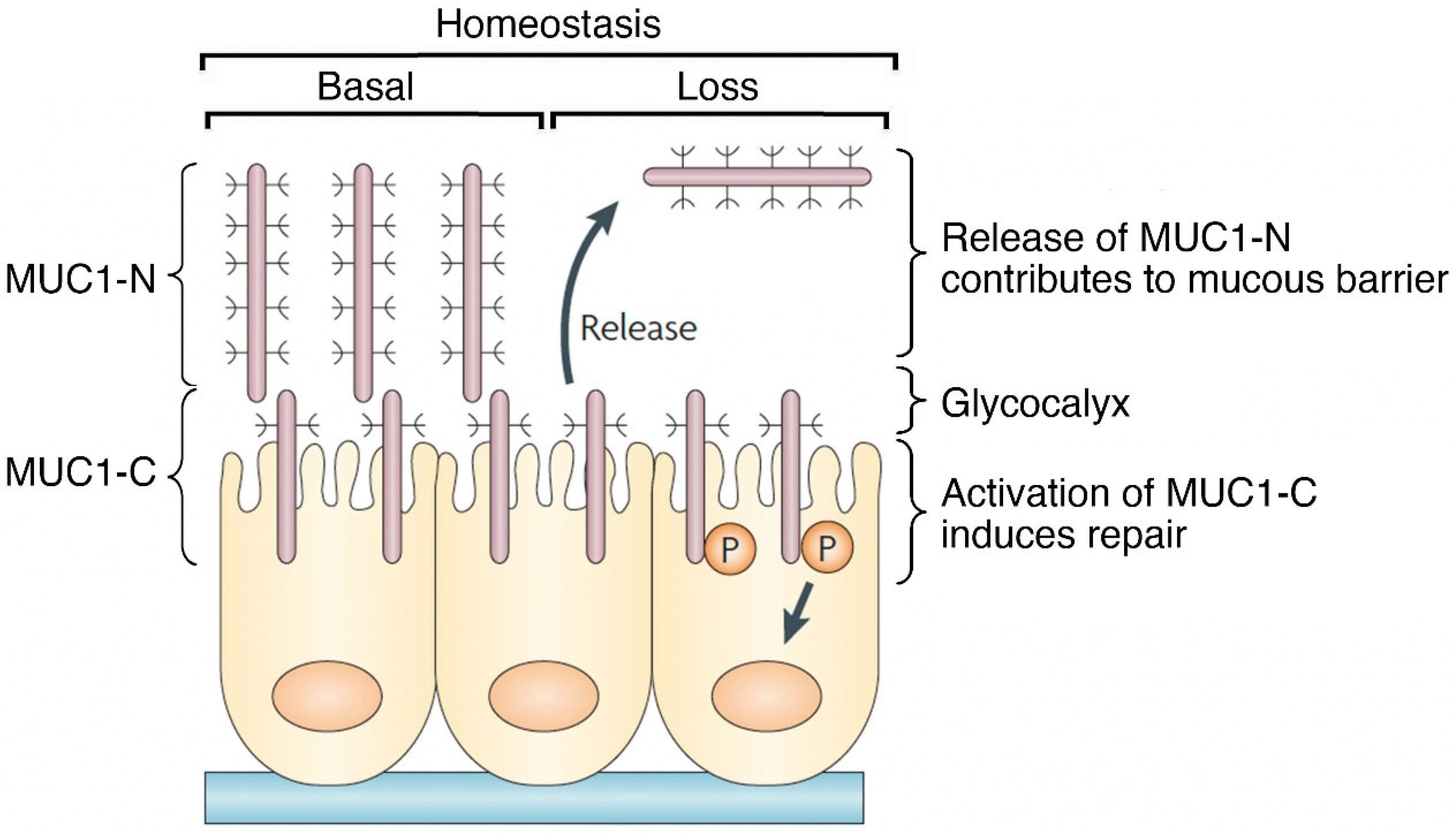
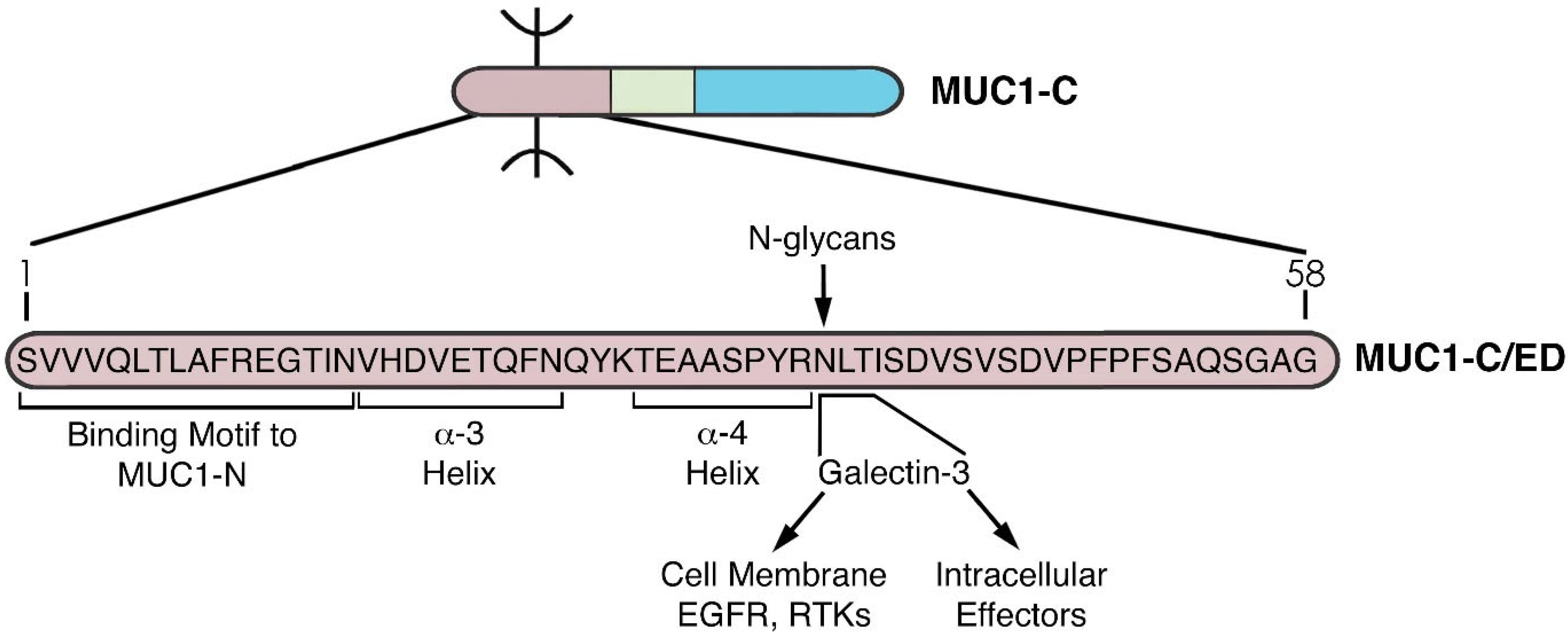
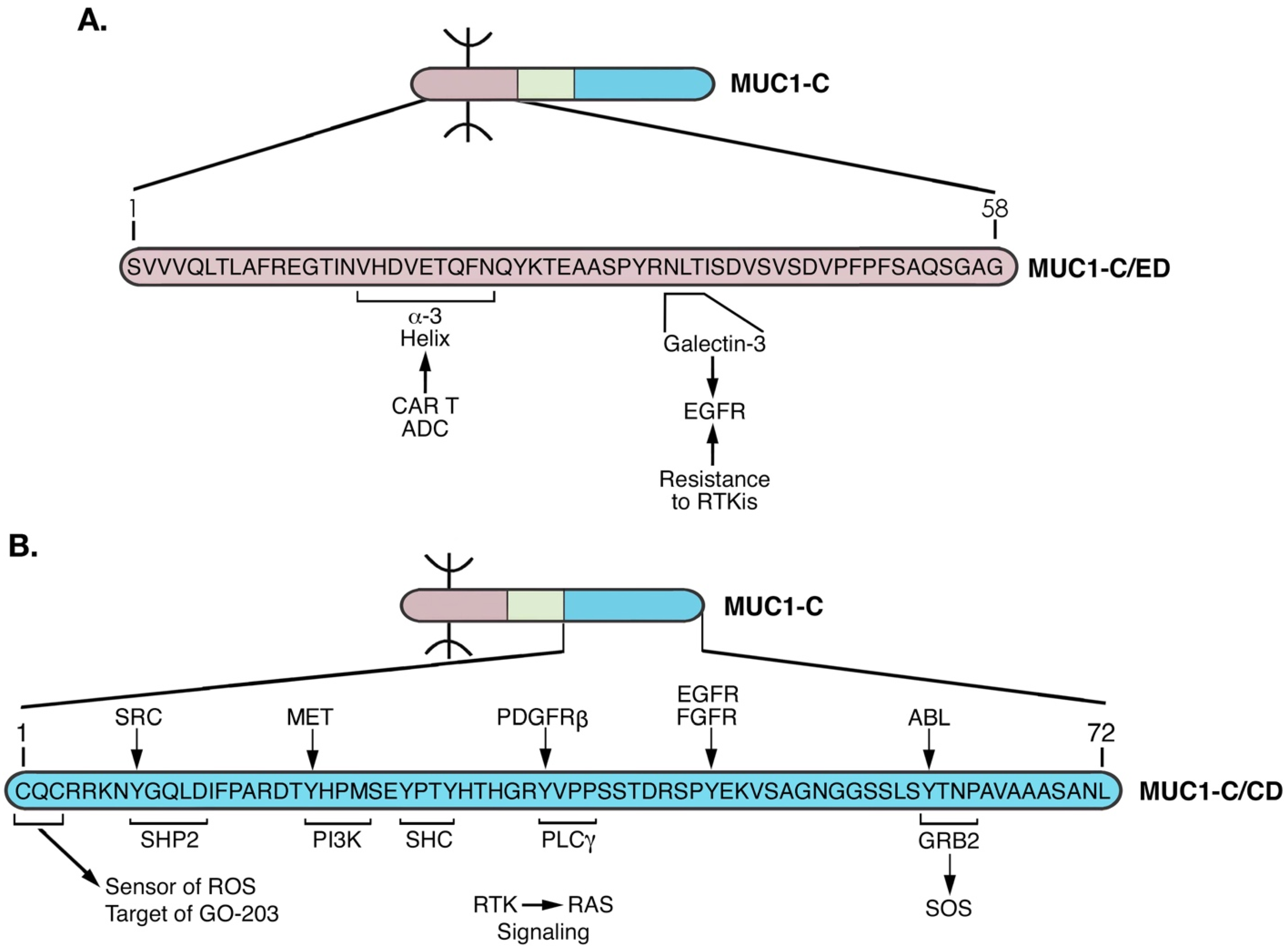
2. Importance of MUC1 Structure for Barrier Tissue Function
3. MUC1 Responds to Loss of Homeostasis by Inducing Loss of Polarity and EMT
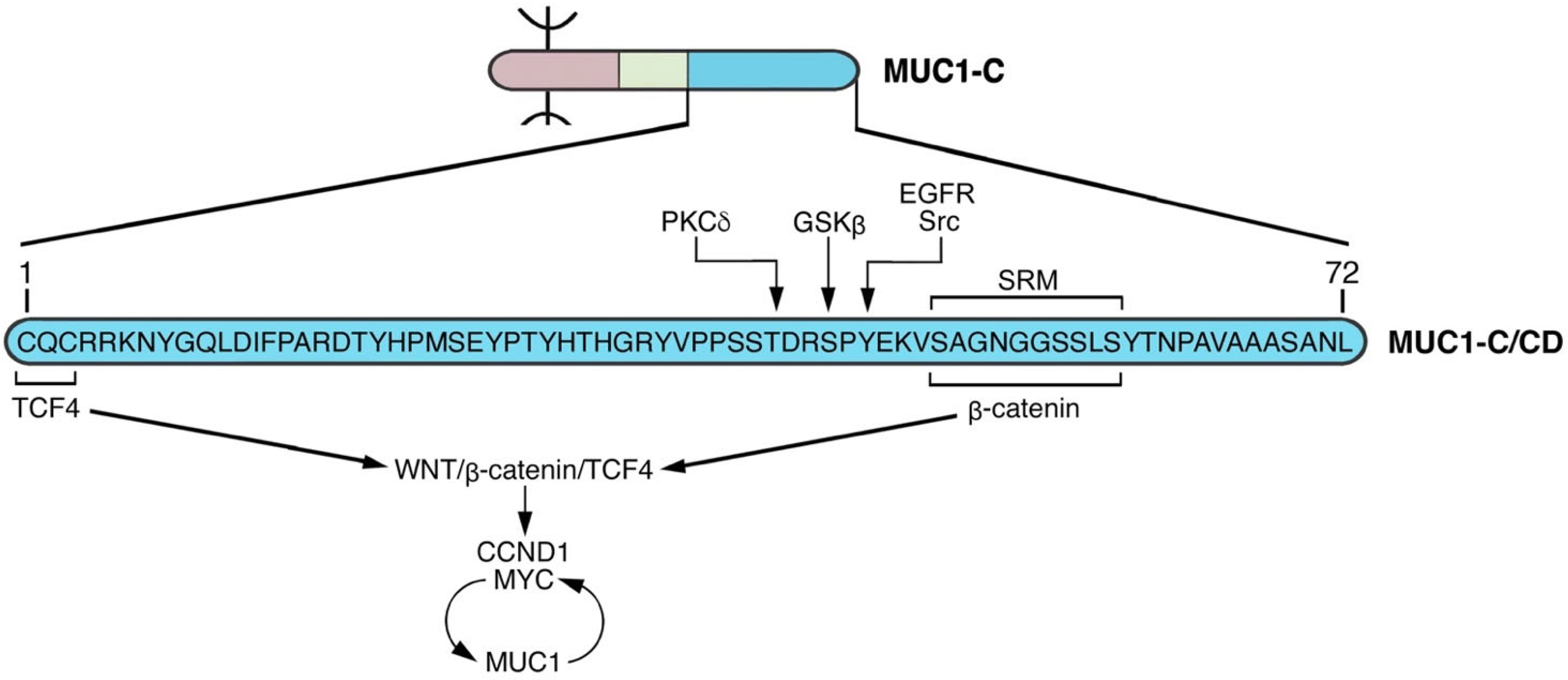
4. MUC1-C Functions as a Node for Activation of the Proliferative WNT/β-Catenin Pathway

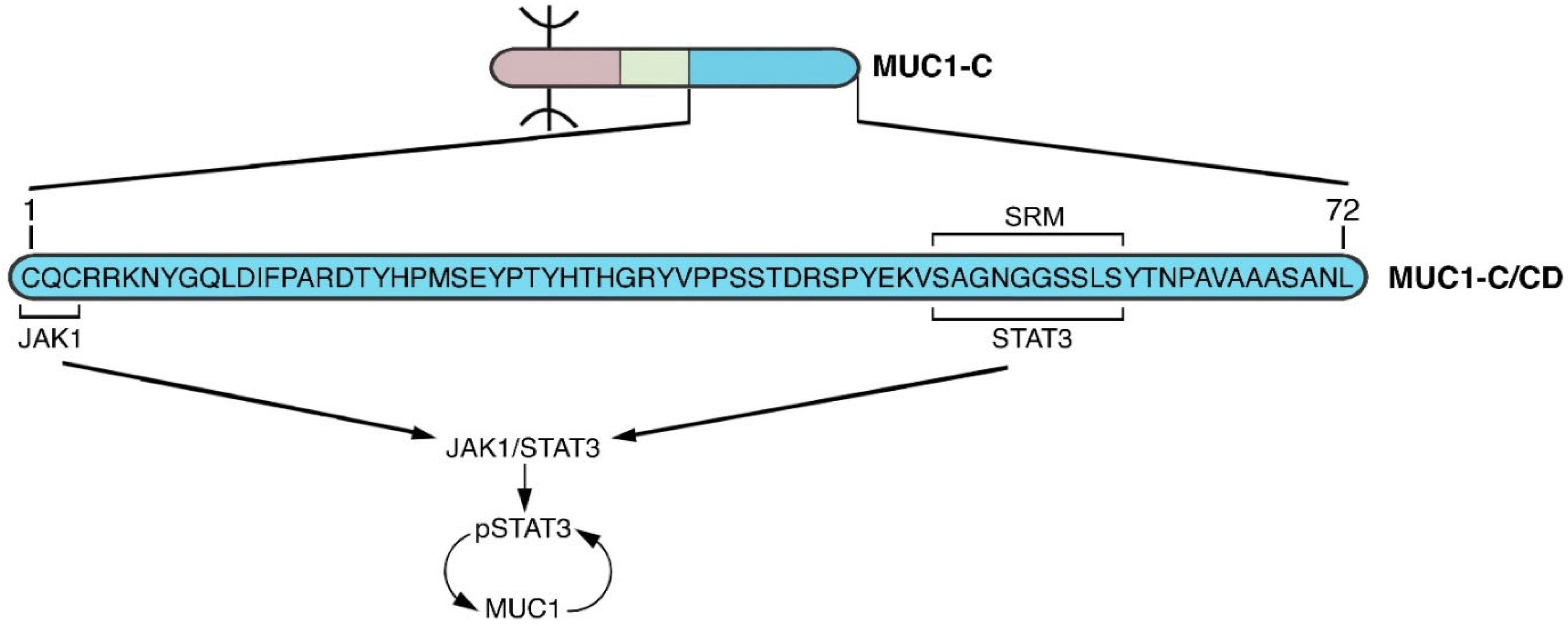
5. MUC1-C Acts as a Node for Promoting Chronic Inflammation
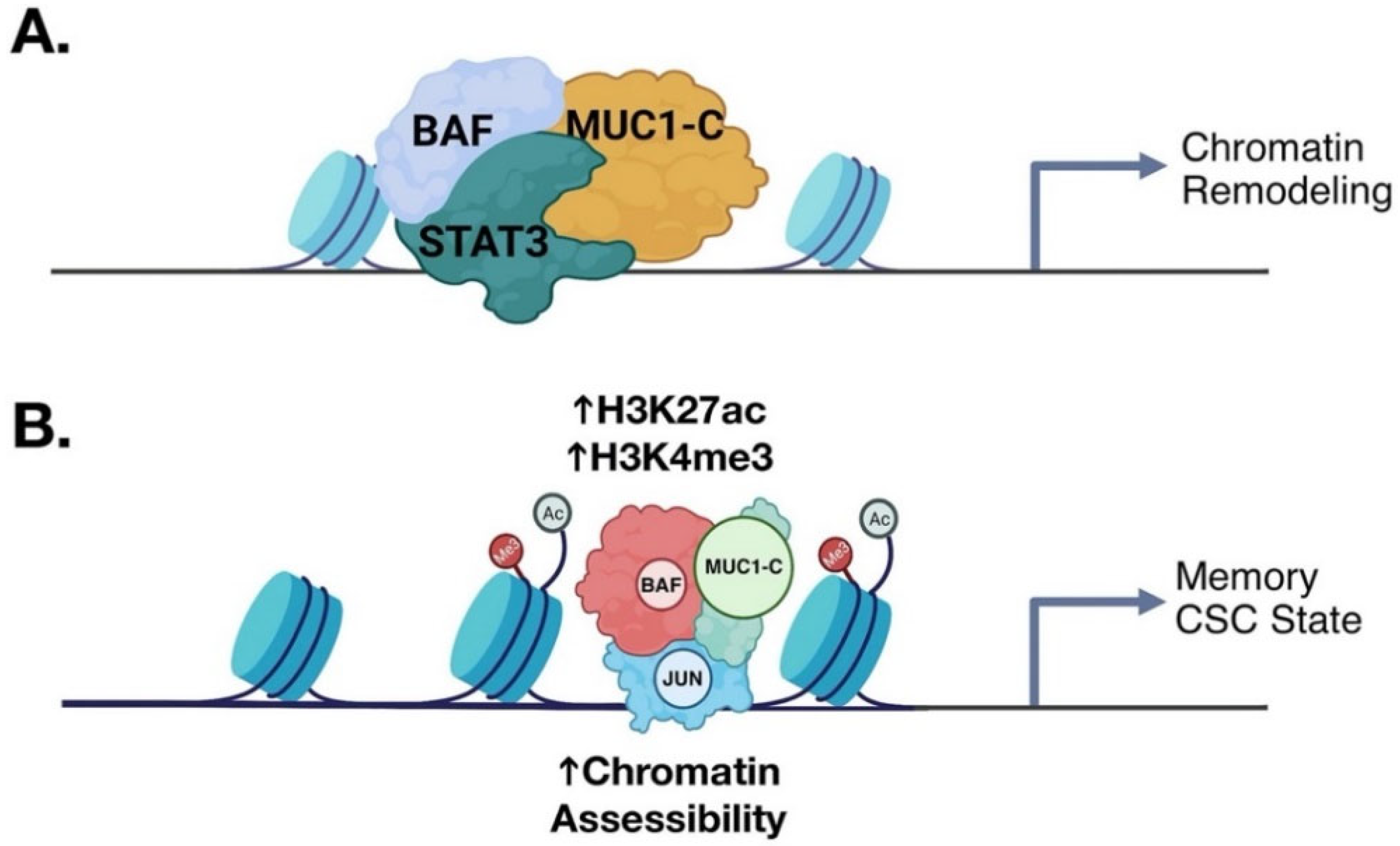
6. MUC1-C and Inflammatory Memory

7. MUC1-C Is Necessary for the CSC State
Targeting MUC1-C-Driven Auto-Inductive Nodes in CSCs for Cancer Treatment
8. Conclusions
Funding
Conflicts of Interest
References
- Kufe, D.; Inghirami, G.; Abe, M.; Hayes, D.; Justi-Wheeler, H.; Schlom, J. Differential Reactivity of a Novel Monoclonal Antibody (DF3) with Human Malignant versus Benign Breast Tumors. Hybridoma 1984, 3, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Arklie, J.; Taylor-Papadimitriou, J.; Bodmer, W.; Egan, M.; Millis, R. Differentiation antigens expressed by epithelial cells in the lactating breast are also detectable in breast cancers. Int. J. Cancer 1981, 28, 23–29. [Google Scholar] [CrossRef]
- Burchell, J.; Durbin, H.; Taylor-Papadimitriou, J. Complexity of expression of antigenic determinants, recognized by mono-clonal antibodies HMFG-1 and HMFG-2, in normal and malignant human mammary epithelial cells. J. Immunol. 1983, 131, 508–513. [Google Scholar] [PubMed]
- Ormerod, M.G.; Steele, K.; Edwards, P.A.; Taylor-Papadimitriou, J. Monoclonal antibodies that react with epithelial membrane antigen. J. Exp. Pathol. 1984, 1, 263–271. [Google Scholar] [PubMed]
- Siddiqui, J.; Abe, M.; Hayes, D.; Shani, E.; Yunis, E.; Kufe, D. Isolation and sequencing of a cDNA coding for the human DF3 breast carcinoma-associated antigen. Proc. Natl. Acad. Sci. USA 1988, 85, 2320–2323. [Google Scholar] [CrossRef] [PubMed]
- Gendler, S.; Taylor-Papadimitriou, J.; Duhig, T.; Rothbard, J.; Burchell, J.A. A highly immunogenic region of a human poly-morphic epithelial mucin expressed by carcinomas is made up of tandem repeats. J. Biol. Chem. 1988, 263, 12820–12823. [Google Scholar] [CrossRef]
- Merlo, G.R.; Siddiqui, J.; Cropp, C.S.; Liscia, D.S.; Lidereau, R.; Callahan, R.; Kufe, D.W. Frequent alteration of the DF3 tumor-associated antigen gene in primary human breast carcinomas. Cancer Res. 1989, 49, 6966–6971. [Google Scholar]
- Gendler, S.; Lancaster, C.; Taylor-Papadimitriou, J.; Duhig, T.; Peat, N.; Burchell, J.; Pemberton, L.; Lalani, E.; Wilson, D. Molecular cloning and expression of human tumor-associated polymorphic epithelial mucin. J. Biol. Chem. 1990, 265, 15286–15293. [Google Scholar] [CrossRef]
- Kufe, D. Mucins in cancer: Function, prognosis and therapy. Nat. Rev. Cancer 2009, 9, 874–885. [Google Scholar] [CrossRef]
- Corfield, A.P. Mucins: A biologically relevant glycan barrier in mucosal protection. Biochim. Biophys. Acta 2015, 1850, 236–252. [Google Scholar] [CrossRef]
- Johansson, M.E.V.; Sjövall, H.; Hansson, G.C. The gastrointestinal mucus system in health and disease. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 352–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linden, S.K.; Sutton, P.; Karlsson, N.G.; Korolik, V.; McGuckin, M.A. Mucins in the mucosal barrier to infection. Mucosal Immunol. 2008, 1, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Liu, F.; Zhao, H.; An, G.; Feng, G. Prognostic significance of mucin antigen MUC1 in various human epithelial cancers, a meta-analysis. Medicine 2015, 94, e2286. [Google Scholar] [CrossRef] [PubMed]
- Kufe, D.W. MUC1-C in chronic inflammation and carcinogenesis; Emergence as a target for cancer treatment. Carcinogenesis 2020, 41, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Kufe, D.W. Chronic activation of MUC1-C in wound repair promotes progression to cancer stem cells. J. Cancer Metastasis Treat. 2022, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, N.; Kufe, D. Addiction of cancer stem cells to MUC1-C in triple-negative breast cancer progression. Int. J. Mol. Sci. 2022, 23, 8219. [Google Scholar] [CrossRef] [PubMed]
- Duraisamy, S.; Kufe, T.; Ramasamy, S.; Kufe, N. Evolution of the human MUC1 oncoprotein. Int. J. Oncol. 2007, 31, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Villanueva-Cañas, J.L.; Ruiz-Orera, J.; Agea, M.I.; Gallo, M.; Andreu, D.; Albà, M.M. New Genes and Functional Innovation in Mammals. Genome Biol. Evol. 2017, 9, 1886–1900. [Google Scholar] [CrossRef] [PubMed]
- Moens, E.; Veldhoen, M. Epithelial barrier biology: Good fences make good neighbours. Immunology 2012, 135, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Franke, F.E.; Kraus, S.; Eiermann, C.; Pauls, K.; Lalani, E.-N.; Bergmann, M. MUC1 in normal and impaired spermatogenesis. Mol. Hum. Reprod. 2001, 7, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Bork, P.; Patthy, L. The SEA module: A new extracellular domain associated with O-glycosylation. Protein Sci. 1995, 4, 1421–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duraisamy, S.; Ramasamy, S.; Kharbanda, S.; Kufe, D. Distinct evolution of the human carcinoma-associated transmembrane mucins, MUC1, MUC4 AND MUC16. Gene 2006, 373, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Grishin, N.V. Expansion of divergent SEA domains in cell surface proteins and nucleoporin 54. Protein Sci. 2017, 26, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Panchamoorthy, G.; Jin, C.; Raina, D.; Bharti, A.; Yamamoto, M.; Adeebge, D.; Zhao, Q.; Bronson, R.; Jiang, S.; Li, L.; et al. Targeting the human MUC1-C oncoprotein with an antibody-drug conjugate. JCI Insight 2018, 3, e99880. [Google Scholar] [CrossRef] [PubMed]
- Pigny, P.; Guyonnet-Duperat, V.; Hill, A.S.; Pratt, W.S.; Galiegue-Zouitina, S.; d’Hooge, M.C.; Laine, A.; Van-Seuningen, I.; Degand, P.; Gum, J.R.; et al. Human mucin genes assigned to 11p15.5: Identification and organization of a cluster of genes. Genomics 1996, 38, 340–352. [Google Scholar] [CrossRef]
- Desseyn, J.-L.; Aubert, J.-P.; Porchet, N.; Laine, A. Evolution of the Large Secreted Gel-Forming Mucins. Mol. Biol. Evol. 2000, 17, 1175–1184. [Google Scholar] [CrossRef]
- Lang, T.; Hansson, G.C.; Samuelsson, T. Gel-forming mucins appeared early in metazoan evolution. Proc. Natl. Acad. Sci. USA 2007, 104, 16209–16214. [Google Scholar] [CrossRef]
- Roy, M.G.; Livraghi-Butrico, A.; Fletcher, A.A.; McElwee, M.M.; Evans, S.E.; Boerner, R.M.; Alexander, S.N.; Bellinghausen, L.K.; Song, A.S.; Petrova, Y.M.; et al. Muc5b is required for airway defence. Nature 2014, 505, 412–416. [Google Scholar] [CrossRef]
- Carpenter, J.; Wang, Y.; Gupta, R.; Li, Y.; Haridass, P.; Subramani, D.B.; Reidel, B.; Morton, L.; Ridley, C.; O’Neal, W.K.; et al. Assembly and organization of the N-terminal region of mucin MUC5AC: Indications for structural and functional distinction from MUC5B. Proc. Natl. Acad. Sci USA 2021, 118, e2104490118. [Google Scholar] [CrossRef]
- Cruz, L.A.; Tellman, T.V.; Farach-Carson, M.C. Flipping the Molecular Switch: Influence of Perlecan and Its Modifiers in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1245, 133–146. [Google Scholar]
- Quiroz, F.G.; Fiore, V.F.; Levorse, J.; Polak, L.; Wong, E.; Pasolli, H.A.; Fuchs, E. Liquid-liquid phase separation drives skin barrier formation. Science 2020, 367, eaax9554. [Google Scholar] [CrossRef] [PubMed]
- Niec, R.E.; Rudensky, A.Y.; Fuchs, E. Inflammatory adaptation in barrier tissues. Cell 2021, 184, 3361–3375. [Google Scholar] [CrossRef] [PubMed]
- Macao, B.; Johansson, D.G.A.; Hansson, G.C.; Härd, T. Autoproteolysis coupled to protein folding in the SEA domain of the membrane-bound MUC1 mucin. Nat. Struct. Mol. Biol. 2005, 13, 71–76. [Google Scholar] [CrossRef]
- Shurer, C.R.; Kuo, J.C.-H.; Roberts, L.M.; Gandhi, J.G.; Colville, M.J.; Enoki, T.A.; Pan, H.; Su, J.; Noble, J.M.; Hollander, M.J.; et al. Physical Principles of Membrane Shape Regulation by the Glycocalyx. Cell 2019, 177, 1757–1770.e21. [Google Scholar] [CrossRef] [PubMed]
- Johansson, D.G.; Macao, B.; Sandberg, A.; Hard, T. SEA domain autoproteolysis accelerated by conformational strain: Mecha-nistic aspects. J. Mol. Biol. 2008, 377, 1130–1143. [Google Scholar] [CrossRef] [PubMed]
- Pelaseyed, T.; Zäch, M.; Petersson, A.C.; Svensson, F.; Johansson, D.G.A.; Hansson, G.C. Unfolding dynamics of the mucin SEA domain probed by force spectroscopy suggest that it acts as a cell-protective device. FEBS J. 2013, 280, 1491–1501. [Google Scholar] [CrossRef] [PubMed]
- Thathiah, A.; Carson, D.D. MT1-MMP mediates MUC1 shedding independent of TACE/ADAM17. Biochem. J. 2004, 382, 363–373. [Google Scholar] [CrossRef]
- Julian, J.; Dharmaraj, N.; Carson, D.D. MUC1 is a substrate for gamma-secretase. J. Cell. Biochem. 2009, 108, 802–815. [Google Scholar] [CrossRef]
- Al-Bataineh, M.M.; Kinlough, C.L.; Mi, Z.; Jackson, E.K.; Mutchler, S.M.; Emlet, D.R.; Kellum, J.A.; Hughey, R.P. KIM-1-mediated anti-inflammatory activity is preserved by MUC1 induction in the proximal tubule during ischemia-reperfusion injury. Am. J. Physiol. Physiol. 2021, 321, F135–F148. [Google Scholar] [CrossRef]
- Zuo, Z.; Gandhi, N.S.; Arndt, K.M.; Mancera, R.L. Free energy calculations of the interactions of c-Jun-based synthetic peptides with the c-Fos protein. Biopolymers 2012, 97, 899–909. [Google Scholar] [CrossRef]
- Alam, M.; Bouillez, A.; Tagde, A.; Ahmad, R.; Rajabi, H.; Maeda, T.; Hiraki, M.; Suzuki, Y.; Kufe, D. MUC1-C Represses the Crumbs Complex Polarity Factor CRB3 and Downregulates the Hippo Pathway. Mol. Cancer Res. 2016, 14, 1266–1276. [Google Scholar] [CrossRef] [PubMed]
- Rajabi, H.; Alam, M.; Takahashi, H.; Kharbanda, A.; Guha, M.; Ahmad, R.; Kufe, D. MUC1-C oncoprotein activates the ZEB1/miR-200c regulatory loop and epithelial-mesenchymal transition. Oncogene 2014, 33, 1680–1689. [Google Scholar] [CrossRef] [Green Version]
- Hata, T.; Rajabi, H.; Yamamoto, M.; Jin, C.; Ahmad, R.; Zhang, Y.; Kui, L.; Li, W.; Yasumizu, Y.; Hong, D.; et al. Targeting MUC1-C Inhibits TWIST1 Signaling in Triple-Negative Breast Cancer. Mol. Cancer Ther. 2019, 18, 1744–1754. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, S.; Duraisamy, S.; Barbashov, S.; Kawano, T.; Kharbanda, S.; Kufe, D. The MUC1 and galectin-3 oncoproteins function in a microRNA-dependent regulatory loop. Mol. Cell 2007, 27, 992–1004. [Google Scholar] [CrossRef] [PubMed]
- Fritsch, K.; Mernberger, M.; Nist, A.; Stiewe, T.; Brehm, A.; Jacob, R. Galectin-3 interacts with components of the nuclear ribo-nucleoprotein complex. BMC Cancer 2016, 16, 502. [Google Scholar] [CrossRef] [PubMed]
- Nangia-Makker, P.; Hogan, V.; Raz, A. Galectin-3 and cancer stemness. Glycobiology 2018, 28, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Nie, M.; Bal, M.S.; Yang, Z.; Liu, J.; Rivera, C.; Wenzel, A.; Beck, B.B.; Sakhaee, K.; Marciano, D.K.; Wolf, M.T. Mucin-1 Increases Renal TRPV5 Activity In Vitro, and Urinary Level Associates with Calcium Nephrolithiasis in Patients. J. Am. Soc. Nephrol. 2016, 27, 3447–3458. [Google Scholar] [CrossRef]
- Leng, Y.; Cao, C.; Ren, J.; Huang, L.; Chen, D.; Ito, M.; Kufe, D. Nuclear import of the MUC1-C oncoprotein is mediated by nu-cleoporin Nup62. J. Biol. Chem. 2007, 282, 19321–19330. [Google Scholar] [CrossRef]
- Raina, D.; Ahmad, R.; Rajabi, H.; Panchamoorthy, G.; Kharbanda, S.; Kufe, D. Targeting cysteine-mediated dimerization of the MUC1-C oncoprotein in human cancer cells. Int. J. Oncol. 2012, 40, 1643–1649. [Google Scholar] [CrossRef]
- Kinlough, C.L.; McMahan, R.J.; Poland, P.A.; Bruns, J.B.; Harkleroad, K.L.; Stremple, R.J.; Kashlan, O.B.; Weixel, K.M.; Weisz, O.A.; Hughey, R.P. Recycling of MUC1 Is Dependent on Its Palmitoylation. J. Biol. Chem. 2006, 281, 12112–12122. [Google Scholar] [CrossRef]
- Hanisch, F.-G.; Kinlough, C.L.; Staubach, S.; Hughey, R.P. MUC1 Membrane Trafficking: Protocols for Assessing Biosynthetic Delivery, Endocytosis, Recycling, and Release Through Exosomes. Methods Mol. Biol. 2011, 842, 123–140. [Google Scholar] [CrossRef]
- Ren, J.; Agata, N.; Chen, D.; Li, Y.; Yu, W.; Huang, L.; Raina, D.; Chen, W.; Kharbanda, S.; Kufe, D. Human MUC1 carcino-ma-associated protein confers resistance to genotoxic anti-cancer agents. Cancer Cell 2004, 5, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Bharti, A.; Raina, D.; Chen, W.; Ahmad, R.; Kufe, D. MUC1 oncoprotein is targeted to mitochondria by heregulin-induced activation of c-Src and the molecular chaperone HSP90. Oncogene 2006, 25, 20–31. [Google Scholar] [CrossRef]
- Ren, J.; Raina, D.; Chen, W.; Li, G.; Huang, L.; Kufe, D. MUC1 Oncoprotein Functions in Activation of Fibroblast Growth Factor Receptor Signaling. Mol. Cancer Res. 2006, 4, 873–883. [Google Scholar] [CrossRef]
- Yamamoto, M.; Bharti, A.; Li, Y.; Kufe, D. Interaction of the DF3/MUC1 breast carcinoma-associated antigen and β-catenin in cell adhesion. J. Biol. Chem. 1997, 272, 12492–12494. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Bharti, A.; Chen, D.; Gong, J.; Kufe, D. Interaction of glycogen synthase kinase 3β with the DF3/MUC1 carcinoma-associated antigen and β-catenin. Mol. Cell. Biol. 1998, 18, 7216–7224. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kuwahara, H.; Ren, J.; Wen, G.; Kufe, D. The c-Src tyrosine kinase regulates signaling of the human DF3/MUC1 carci-noma-associated antigen with GSK3β and β-catenin. J. Biol. Chem. 2001, 276, 6061–6064. [Google Scholar] [CrossRef]
- Li, Y.; Ren, J.; Yu, W.; Li, G.; Kuwahara, H.; Yin, L.; Carraway, K.L.; Kufe, D. The EGF receptor regulates interaction of the human DF3/MUC1 carcinoma antigen with c-Src and β-catenin. J. Biol. Chem. 2001, 276, 35239–35242. [Google Scholar] [CrossRef]
- Ren, J.; Li, Y.; Kufe, D. Protein kinase C δ regulates function of the DF3/MUC1 carcinoma antigen in β-catenin signaling. J. Biol. Chem. 2002, 277, 17616–17622. [Google Scholar] [CrossRef]
- Huang, L.; Chen, D.; Liu, D.; Yin, L.; Kharbanda, S.; Kufe, D. MUC1 oncoprotein blocks GSK3β-mediated phosphorylation and degradation of β-catenin. Cancer Res. 2005, 65, 10413–10422. [Google Scholar] [CrossRef]
- Rajabi, H.; Ahmad, R.; Jin, C.; Kosugi, M.; Alam, M.; Joshi, M.D.; Kufe, D. MUC1-C Oncoprotein Induces TCF7L2 Transcription Factor Activation and Promotes Cyclin D1 Expression in Human Breast Cancer Cells. J. Biol. Chem. 2012, 287, 10703–10713. [Google Scholar] [CrossRef] [PubMed]
- Berlow, R.; Dyson, H.J.; Wright, P.E. Expanding the Paradigm: Intrinsically Disordered Proteins and Allosteric Regulation. J. Mol. Biol. 2018, 430, 2309–2320. [Google Scholar] [CrossRef]
- Bouillez, A.; Rajabi, H.; Pitroda, S.; Jin, C.; Alam, M.; Kharbanda, A.; Tagde, A.; Wong, K.-K.; Kufe, D. Inhibition of MUC1-C Suppresses MYC Expression and Attenuates Malignant Growth in KRAS Mutant Lung Adenocarcinomas. Cancer Res. 2016, 76, 1538–1548. [Google Scholar] [CrossRef]
- Hata, T.; Rajabi, H.; Takahashi, H.; Yasumizu, Y.; Li, W.; Jin, C.; Long, M.D.; Hu, Q.; Liu, S.; Fushimi, A.; et al. MUC1-C Activates the NuRD Complex to Drive Dedifferentiation of Triple-Negative Breast Cancer Cells. Cancer Res. 2019, 79, 5711–5722. [Google Scholar] [CrossRef]
- Fushimi, A.; Morimoto, Y.; Ishikawa, S.; Yamashita, N.; Bhattacharya, A.; Daimon, T.; Rajabi, H.; Jin, C.; Hagiwara, M.; Yasumizu, Y.; et al. Dependence on the MUC1-C oncoprotein in classic, variant and non-neuroendocrine small cell lung cancer. Mol. Cancer Res. 2022, 20, 1379–1390. [Google Scholar] [CrossRef]
- Hagiwara, M.; Yasumizu, Y.; Yamashita, N.; Rajabi, H.; Fushimi, A.; Long, M.D.; Li, W.; Bhattacharya, A.; Ahmad, R.; Oya, M.; et al. MUC1-C Activates the BAF (mSWI/SNF) Complex in Prostate Cancer Stem Cells. Cancer Res. 2021, 81, 1111–1122. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, M.; Fushimi, A.; Yamashita, N.; Bhattacharya, A.; Rajabi, H.; Long, M.D.; Yasumizu, Y.; Oya, M.; Liu, S.; Kufe, D. MUC1-C activates the PBAF chromatin remodeling complex in integrating redox balance with progression of human prostate cancer stem cells. Oncogene 2021, 40, 4930–4940. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Fushimi, A.; Yamashita, N.; Hagiwara, M.; Morimoto, Y.; Rajabi, H.; Long, M.D.; Abdulla, M.; Ahmad, R.; Street, K.; et al. MUC1-C Dictates JUN and BAF-Mediated Chromatin Remodeling at Enhancer Signatures in Cancer Stem Cells. Mol. Cancer Res. 2022, 20, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Jin, C.; Rajabi, H.; Pitroda, S.P.; Alam, M.; Ahmad, R.; Raina, D.; Hasegawa, M.; Suzuki, Y.; Tagde, A.; et al. MUC1-C activates the TAK1 inflammatory pathway in colon cancer. Oncogene 2015, 34, 5187–5197. [Google Scholar] [CrossRef] [PubMed]
- Rajabi, H.; Hiraki, M.; Tagde, A.; Alam, M.; Bouillez, A.; Christensen, C.L.; Samur, M.; Wong, K.-K.; Kufe, D. MUC1-C activates EZH2 expression and function in human cancer cells. Sci. Rep. 2017, 7, 7481. [Google Scholar] [CrossRef]
- Hiraki, M.; Maeda, T.; Bouillez, A.; Alam, M.; Tagde, A.; Hinohara, K.; Suzuki, Y.; Markert, T.; Miyo, M.; Komura, K.; et al. MUC1-C activates BMI1 in human cancer cells. Oncogene 2017, 36, 2791–2801. [Google Scholar] [CrossRef] [PubMed]
- Burgy, O.; Königshoff, M. The WNT signaling pathways in wound healing and fibrosis. Matrix Biol. 2018, 68–69, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Bastakoty, D.; Young, P.P. Wnt/beta-catenin pathway in tissue injury: Roles in pathology and therapeutic opportunities for regeneration. FASEB J. 2016, 30, 3271–3284. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, M.; Werner, S. Cancer as an overhealing wound: An old hypothesis revisited. Nat. Rev. Mol. Cell Biol. 2008, 9, 628–638. [Google Scholar] [CrossRef] [PubMed]
- Arwert, E.N.; Hoste, E.; Watt, F.M. Epithelial stem cells, wound healing and cancer. Nat. Rev. Cancer 2012, 12, 170–180. [Google Scholar] [CrossRef]
- Ge, Y.; Fuchs, E. Stretching the limits, from homeostasis to stem cell plasticity in wound healing and cancer. Nat. Rev. Genet. 2018, 19, 311–325. [Google Scholar] [CrossRef]
- Ahmad, R.; Raina, D.; Joshi, M.D.; Kawano, T.; Kharbanda, S.; Kufe, D. MUC1-C oncoprotein functions as a direct activator of the NF-kappaB p65 transcription factor. Cancer Res 2009, 69, 7013–7021. [Google Scholar] [CrossRef] [PubMed]
- Rajabi, H.; Kufe, D. MUC1-C Oncoprotein Integrates a Program of EMT, Epigenetic Reprogramming and Immune Evasion in Human Carcinomas. Biochim. Biophys. Acta Rev. Cancer 2017, 1868, 117–122. [Google Scholar] [CrossRef]
- Rajabi, H.; Tagde, A.; Alam, M.; Bouillez, A.; Pitroda, S.; Suzuki, Y.; Kufe, D. DNA methylation by DNMT1 and DNMT3b me-thyltransferases is driven by the MUC1-C oncoprotein in human carcinoma cells. Oncogene 2016, 35, 6439–6445. [Google Scholar] [CrossRef]
- Maeda, T.; Hiraki, M.; Jin, C.; Rajabi, H.; Tagde, A.; Alam, M.; Bouillez, A.; Hu, X.; Suzuki, Y.; Miyo, M.; et al. MUC1-C Induces PD-L1 and Immune Evasion in Triple-Negative Breast Cancer. Cancer Res. 2018, 78, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Raina, D.; Trivedi, V.; Ren, J.; Rajabi, H.; Kharbanda, S.; Kufe, D. MUC1 oncoprotein activates the IκB kinase β complex and constitutive NF-κB signaling. Nat. Cell Biol. 2007, 9, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Rajabi, H.; Kosugi, M.; Joshi, M.; Alam, M.; Vasir, B.; Kawano, T.; Kharbanda, S.; Kufe, D. MUC1-C oncoprotein promotes STAT3 activation in an auto-inductive regulatory loop. Sci. Signal. 2011, 4, ra9. [Google Scholar] [CrossRef] [Green Version]
- Naik, S.; Fuchs, E. Inflammatory memory and tissue adaptation in sickness and in health. Nature 2022, 607, 249–255. [Google Scholar] [CrossRef]
- Larsen, S.B.; Cowley, C.J.; Sajjath, S.M.; Barrows, D.; Yang, Y.; Carroll, T.S.; Fuchs, E. Establishment, maintenance, and recall of inflammatory memory. Cell Stem Cell 2021, 28, 1758–1774.e8. [Google Scholar] [CrossRef] [PubMed]
- Eferl, R.; Wagner, E.F. AP-1, a double-edged sword in tumorigenesis. Nat. Cancer 2003, 3, 859–868. [Google Scholar] [CrossRef]
- Bejjani, F.; Evanno, E.; Zibara, K.; Piechaczyk, M.; Jariel-Encontre, I. The AP-1 transcriptional complex: Local switch or remote command? Biochim. Biophys. Acta Rev. Cancer 2019, 1872, 11–23. [Google Scholar] [CrossRef]
- Wang, A.G.; Son, M.; Kenna, E.; Thom, N.; Tay, S. NF-kappaB memory coordinates transcriptional responses to dynamic in-flammatory stimuli. Cell Rep. 2022, 40, 111159. [Google Scholar] [CrossRef]
- Rajabi, H.; Hiraki, M.; Kufe, D. MUC1-C activates polycomb repressive complexes and downregulates tumor suppressor genes in human cancer cells. Oncogene 2018, 37, 2079–2088. [Google Scholar] [CrossRef]
- Yamamoto, M.; Jin, C.; Hata, T.; Yasumizu, Y.; Zhang, Y.; Hong, D.; Maeda, T.; Miyo, M.; Hiraki, M.; Suzuki, Y.; et al. MUC1-C Integrates Chromatin Remodeling and PARP1 Activity in the DNA Damage Response of Triple-Negative Breast Cancer Cells. Cancer Res. 2019, 79, 2031–2041. [Google Scholar] [CrossRef]
- Piunti, A.; Shilatifard, A. The roles of Polycomb repressive complexes in mammalian development and cancer. Nat. Rev. Mol. Cell Biol. 2021, 22, 326–345. [Google Scholar] [CrossRef] [PubMed]
- Cenik, B.K.; Shilatifard, A. COMPASS and SWI/SNF complexes in development and disease. Nat. Rev. Genet. 2021, 22, 38–58. [Google Scholar] [CrossRef] [PubMed]
- Yasumizu, Y.; Rajabi, H.; Jin, C.; Hata, T.; Pitroda, S.; Long, M.D.; Hagiwara, M.; Li, W.; Hu, Q.; Liu, S.; et al. MUC1-C regulates lineage plasticity driving progression to neuroendocrine prostate cancer. Nat. Commun. 2020, 11, 338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Zhang, N.; Jin, C.; Long, M.D.; Rajabi, H.; Yasumizu, Y.; Fushimi, A.; Yamashita, N.; Hagiwara, M.; Zheng, R.; et al. MUC1-C drives stemness in progression of colitis to colorectal cancer. JCI Insight 2020, 5, 137112. [Google Scholar] [CrossRef] [PubMed]
- Luan, Z.; Morimoto, Y.; Fushimi, A.; Yamashita, N.; Suo, W.; Bhattacharya, A.; Hagiwara, M.; Jin, C.; Kufe, D. MUC1-C dictates neuroendocrine lineage specification in pancreatic ductal adenocarcinomas. Carcinogenesis 2021, 43, 67–76. [Google Scholar] [CrossRef]
- Watanabe, A.; Yamada, Y.; Yamanaka, S. Epigenetic regulation in pluripotent stem cells, a key to breaking the epigenetic barrier. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120292. [Google Scholar] [CrossRef]
- Morimoto, Y.; Fushimi, A.; Yamashita, N.; Hagiwara, M.; Bhattacharya, A.; Cheng, J.; Frost, T.C.; Ahmad, R.; Daimon, T.; Huang, L.; et al. Addiction of Merkel cell carcinoma to MUC1-C identifies a potential new target for treatment. Oncogene 2022, 41, 3511–3523. [Google Scholar] [CrossRef]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer, Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Stroopinsky, D.; Rosenblatt, J.; Ito, K.; Mills, H.; Yin, L.; Rajabi, H.; Vasir, B.; Kufe, T.; Luptakova, K.; Arnason, J.; et al. MUC1 Is a Potential Target for the Treatment of Acute Myeloid Leukemia Stem Cells. Cancer Res. 2013, 73, 5569–5579. [Google Scholar] [CrossRef]
- Jain, S.; Stroopinsky, D.; Yin, L.; Rosenblatt, J.; Alam, M.; Bhargava, P.; Clark, R.A.; Kupper, T.S.; Palmer, K.; Coll, M.D.; et al. Mucin 1 is a potential therapeutic target in cutaneous T-cell lymphoma. Blood 2015, 126, 354–362. [Google Scholar] [CrossRef]
- Tagde, A.; Rajabi, H.; Bouillez, A.; Alam, M.; Gali, R.; Bailey, S.; Tai, Y.-T.; Hideshima, T.; Anderson, K.; Avigan, D.; et al. MUC1-C drives MYC in multiple myeloma. Blood 2016, 127, 2587–2597. [Google Scholar] [CrossRef] [PubMed]
- Kharbanda, A.; Rajabi, H.; Jin, C.; Tchaicha, J.; Kikuchi, E.; Wong, K.-K.; Kufe, D. Targeting the Oncogenic MUC1-C Protein Inhibits Mutant EGFR-Mediated Signaling and Survival in Non–Small Cell Lung Cancer Cells. Clin. Cancer Res. 2014, 20, 5423–5434. [Google Scholar] [CrossRef]
- Raina, D.; Kharbanda, S.; Kufe, D. The MUC1 oncoprotein activates the anti-apoptotic PI3K/Akt and Bcl-xL pathways in rat 3Y1 fibroblasts. J. Biol. Chem. 2004, 279, 20607–20612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, P.; Kharbanda, S.; Kufe, D. Association of the DF3/MUC1 breast cancer antigen with Grb2 and the Sos/Ras exchange protein. Cancer Res. 1995, 55, 4000–4003. [Google Scholar] [PubMed]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic inflammation in the etiology of disease across the life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Ugai, T.; Sasamoto, N.; Lee, H.-Y.; Ando, M.; Song, M.; Tamimi, R.M.; Kawachi, I.; Campbell, P.T.; Giovannucci, E.L.; Weiderpass, E.; et al. Is early-onset cancer an emerging global epidemic? Current evidence and future implications. Nat. Rev. Clin. Oncol. 2022, 19, 1–18. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kufe, D.W. Emergence of MUC1 in Mammals for Adaptation of Barrier Epithelia. Cancers 2022, 14, 4805. https://doi.org/10.3390/cancers14194805
Kufe DW. Emergence of MUC1 in Mammals for Adaptation of Barrier Epithelia. Cancers. 2022; 14(19):4805. https://doi.org/10.3390/cancers14194805
Chicago/Turabian StyleKufe, Donald W. 2022. "Emergence of MUC1 in Mammals for Adaptation of Barrier Epithelia" Cancers 14, no. 19: 4805. https://doi.org/10.3390/cancers14194805
APA StyleKufe, D. W. (2022). Emergence of MUC1 in Mammals for Adaptation of Barrier Epithelia. Cancers, 14(19), 4805. https://doi.org/10.3390/cancers14194805