

Nerve Density and Neuronal Biomarkers in Cancer

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
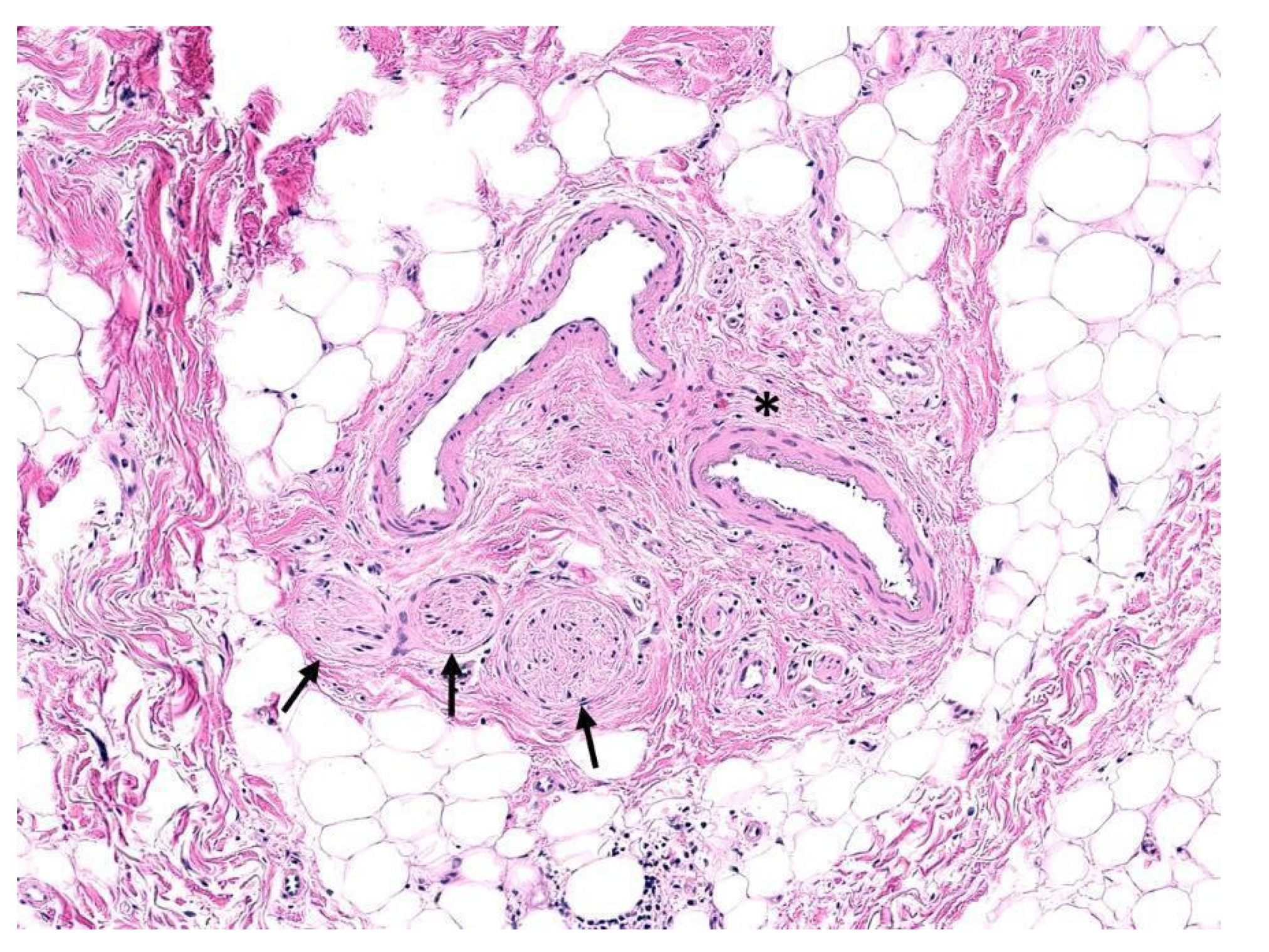
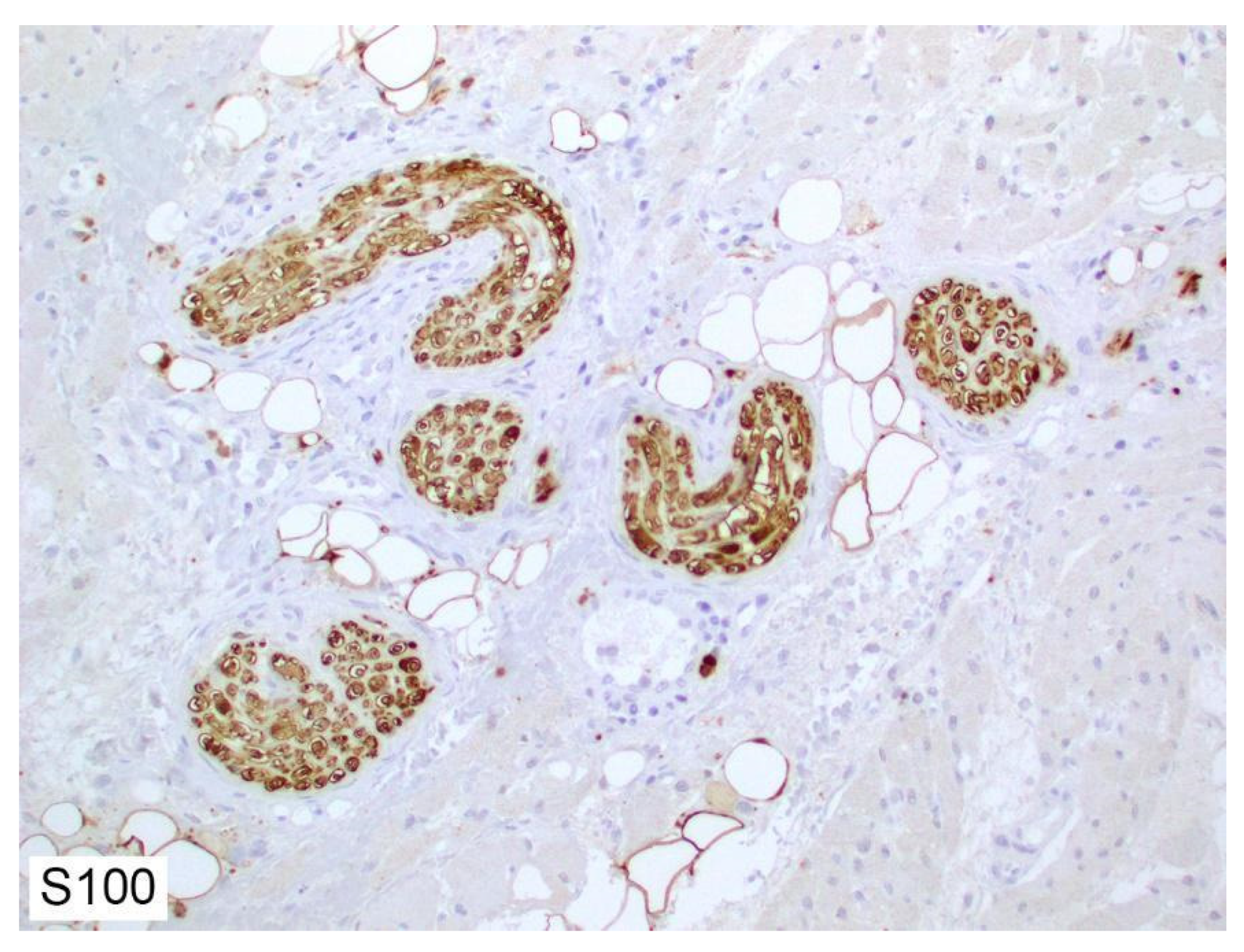
2. Histology of Peripheral Nerves
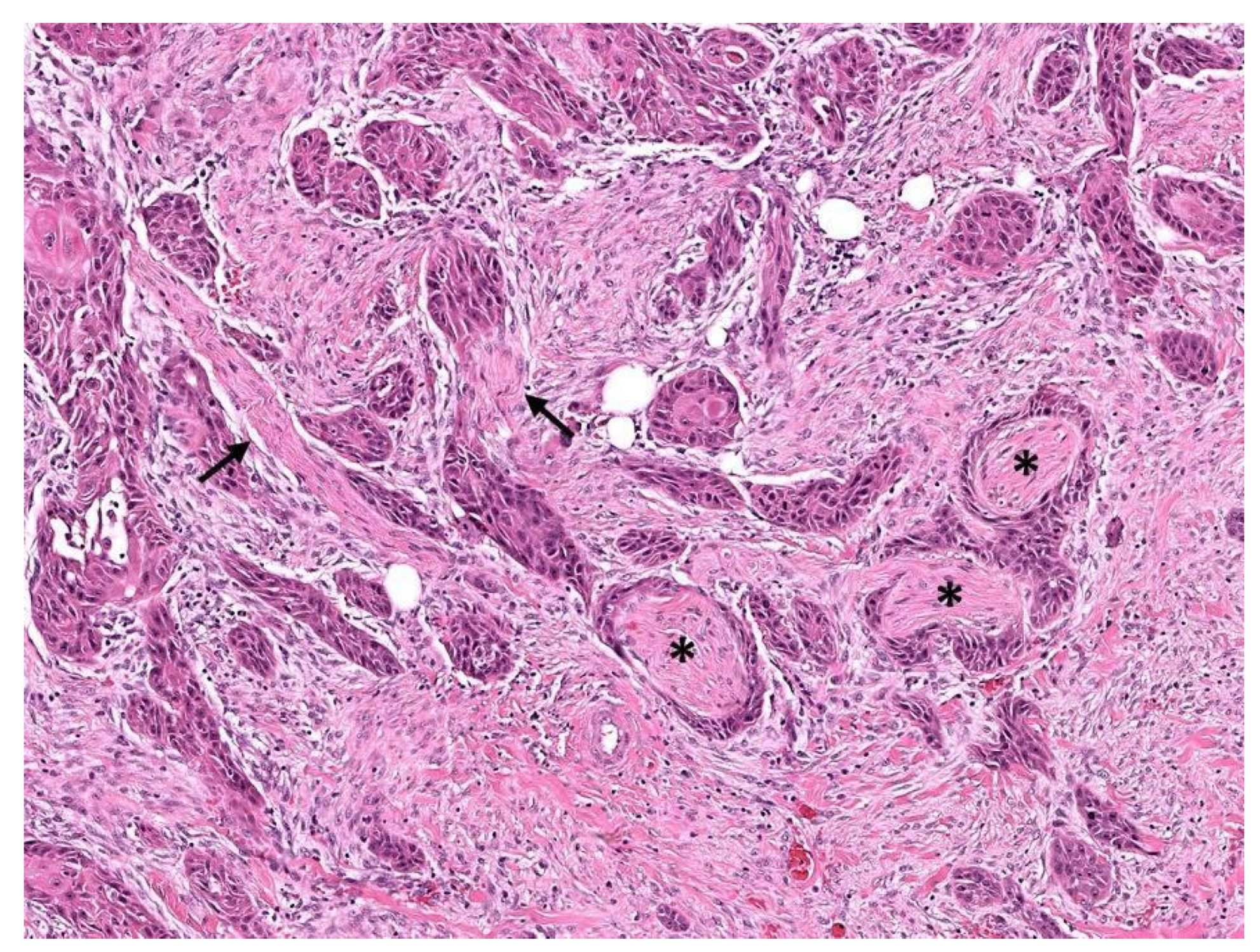
3. Perineurial Invasion
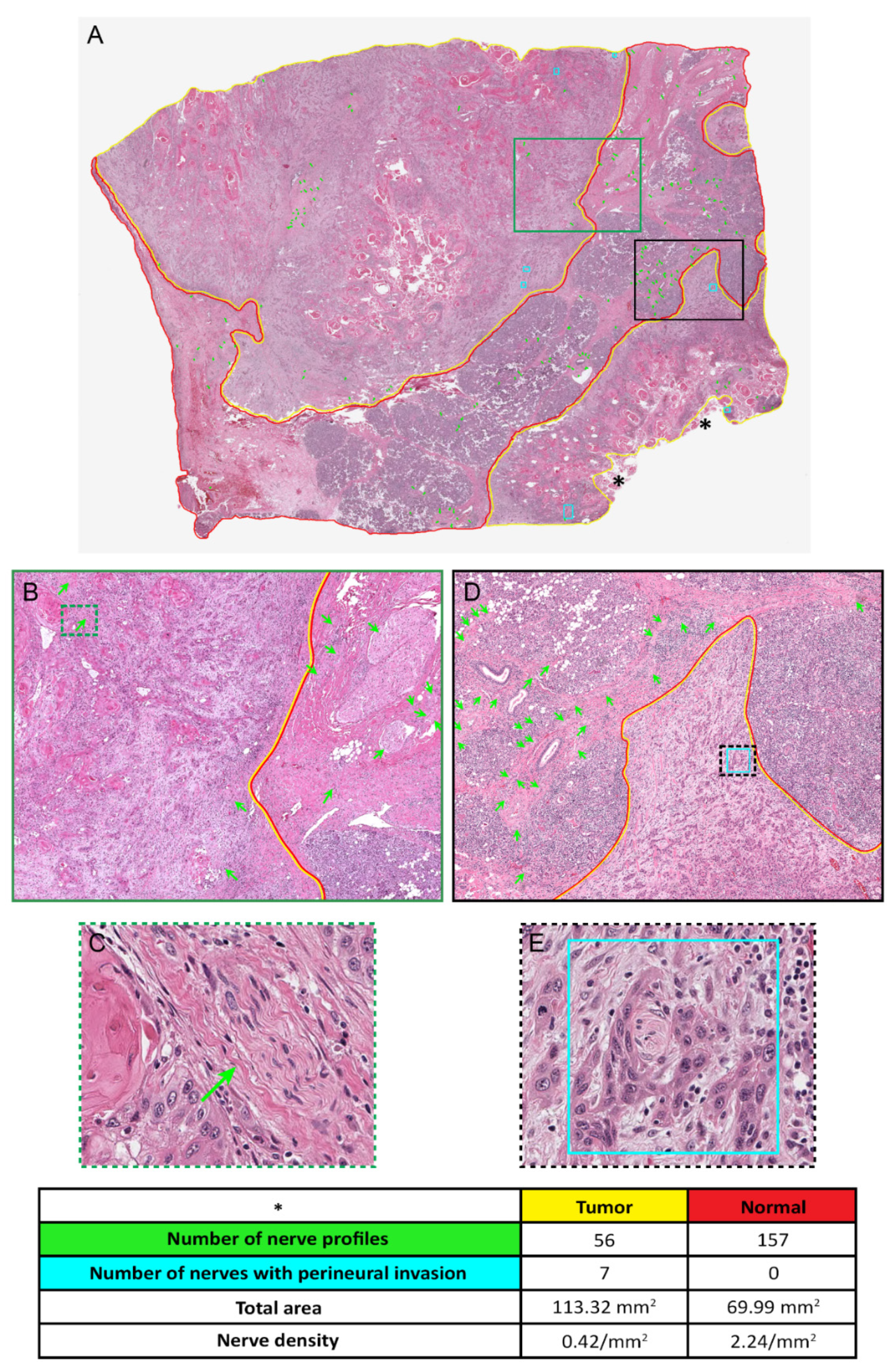
4. Determination of Nerve Density
5. Mechanism of Interaction between Nerves and Cancer
5.1. Effect of Tumor Cells on Nerves
5.2. Indirect Effects of the Nerves on Tumors
6. Neural Biomarkers and Their Role in Cancer
6.1. Biomarkers in Immature Neurons
6.1.1. NeuroD1
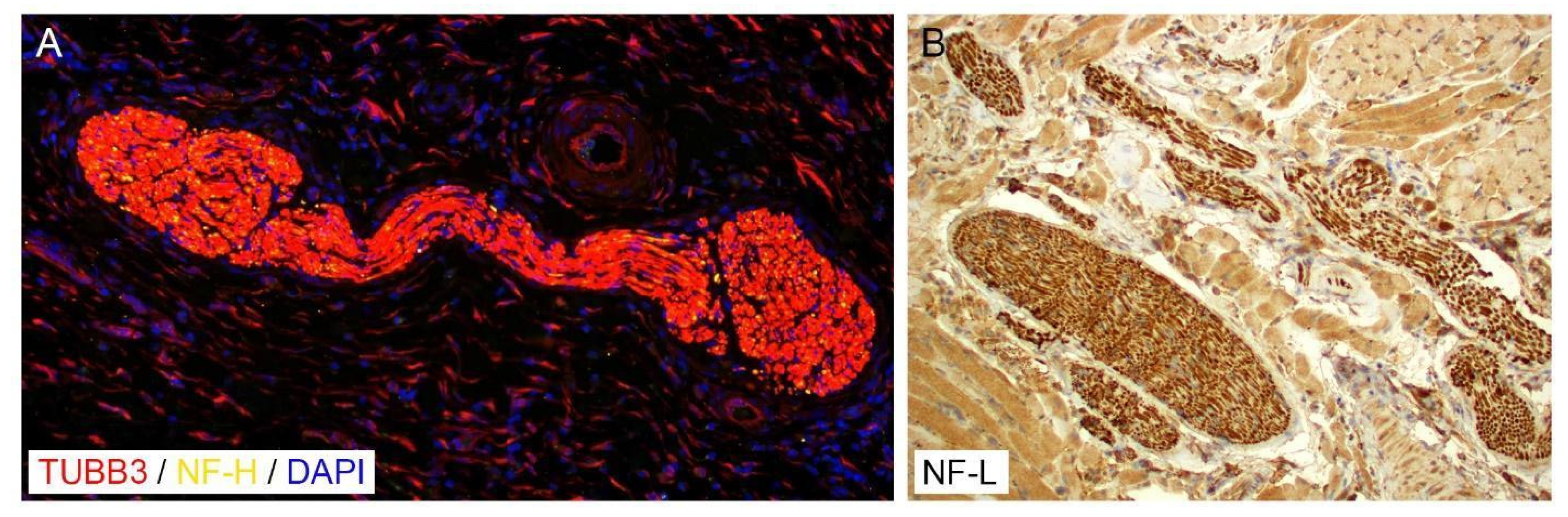
6.1.2. Tubulin Beta-3 Chain
6.1.3. Stathmin 1
6.2. Biomarkers in Mature Neurons
6.2.1. Microtubule-Associated Protein 2
6.2.2. Synaptophysin
7. Clinical Evidence of Nerve Density in Cancer
8. Discussion
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Baghban, R.; Roshangar, L.; Jahanban-Esfahlan, R.; Seidi, K.; Ebrahimi-Kalan, A.; Jaymand, M.; Kolahian, S.; Javaheri, T.; Zare, P. Tumor Microenvironment Complexity and Therapeutic Implications at a Glance. Cell Commun. Signal. 2020, 18, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batalla-Covello, J.; Ali, S.; Xie, T.; Amit, M. β-Adrenergic Signaling in Skin Cancer. FASEB BioAdvances 2022, 4, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Amit, M.; Takahashi, H.; Dragomir, M.P.; Lindemann, A.; Gleber-Netto, F.O.; Pickering, C.R.; Anfossi, S.; Osman, A.A.; Cai, Y.; Wang, R.; et al. Loss of p53 Drives Neuron Reprogramming in Head and Neck Cancer. Nature 2020, 578, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Smits, M. MRI Biomarkers in Neuro-Oncology. Nat. Rev. Neurol. 2021, 17, 486–500. [Google Scholar] [CrossRef]
- Shield, L.K.; King, R.H.; Thomas, P.K. A Morphometric Study of Human Fetal Sural Nerve. Acta Neuropathol. 1986, 70, 60–70. [Google Scholar] [CrossRef]
- Geuna, S.; Raimondo, S.; Ronchi, G.; Di Scipio, F.; Tos, P.; Czaja, K.; Fornaro, M. Chapter 3 Histology of the Peripheral Nerve and Changes Occurring During Nerve Regeneration. In International Review of Neurobiology; Academic Press: Cambridge, MA, USA, 2009; Volume 87, pp. 27–46. [Google Scholar]
- Ortiz-Hidalgo, C.W.R.O. Histology for Pathologists; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2012. [Google Scholar]
- Shea, T.B.; Chan, W.K.-H.; Kushkuley, J.; Lee, S. Organizational Dynamics, Functions, and Pathobiological Dysfunctions of Neurofilaments. Results Probl. Cell Differ. 2009, 48, 29–45. [Google Scholar]
- Al-Chalabi, A.; Miller, C.C.J. Neurofilaments and Neurological Disease. BioEssays 2003, 25, 346–355. [Google Scholar] [CrossRef]
- Julien, J.P.; Mushynski, W.E. Neurofilaments in Health and Disease. Prog. Nucleic Acid Res. Mol. Biol. 1998, 61, 1–23. [Google Scholar]
- Taveggia, C.; Feltri, M.L. Beyond Wrapping: Canonical and Noncanonical Functions of Schwann Cells. Annu. Rev. Neurosci. 2022, 45, 561–580. [Google Scholar] [CrossRef]
- Sherman, D.L.; Brophy, P.J. Mechanisms of Axon Ensheathment and Myelin Growth. Nat. Rev. Neurosci. 2005, 6, 683–690. [Google Scholar] [CrossRef]
- Mirsky, R.; Jessen, K.R. The Neurobiology of Schwann Cells. Brain Pathol. 1999, 9, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Garavito, Z.V.; Sutachán, J.J.; Muñetón, V.C.; Hurtado, H. Is S-100 Protein a Suitable Marker for Adult Schwann Cells? In Vitro Cell. Dev. Biol. Anim. 2000, 36, 281–283. [Google Scholar] [CrossRef]
- Scarpini, E.; Meola, G.; Baron, P.L.; Beretta, S.; Velicogna, M.; Moggio, M.; Buscaglia, M.; Doronzo, R.; Scarlato, G. Human Schwann Cells: Cytochemical, Ultrastructural and Immunological Studies in Vivo and in Vitro. Basic Appl. Histochem. 1987, 31, 33–42. [Google Scholar] [PubMed]
- Khalifa, M.A.; Montgomery, E.A.; Ismiil, N.; Azumi, N. What Are the CD34+ Cells in Benign Peripheral Nerve Sheath Tumors? Double Immunostaining Study of CD34 and S-100 Protein. Am. J. Clin. Pathol. 2000, 114, 123–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piña-Oviedo, S.; Ortiz-Hidalgo, C. The Normal and Neoplastic Perineurium: A Review. Adv. Anat. Pathol. 2008, 15, 147. [Google Scholar] [CrossRef]
- Tassler, P.L.; Dellon, A.L.; Canoun, C. Identification of Elastic Fibres in the Peripheral Nerve. J. Hand Surg. Br. 1994, 19, 48–54. [Google Scholar] [CrossRef]
- Neurosurgery. Available online: https://journals.lww.com/neurosurgery/pages/default.aspx (accessed on 27 May 2022).
- Neumann, E. Secundäre Cancroidinfiltration Des Nervus Mentalis Bei Einem Fall von Lippencancroid. Arch. Pathol. Anat. Physiol. Klin. Med. 1862, 24, 201–202. [Google Scholar] [CrossRef] [Green Version]
- Dunn, M.; Morgan, M.B.; Beer, T.W. Perineural Invasion: Identification, Significance, and a Standardized Definition. Dermatol. Surg. 2009, 35, 214–221. [Google Scholar] [CrossRef]
- Batsakis, J.G. Nerves and Neurotropic Carcinomas. Ann. Otol. Rhinol. Laryngol. 1985, 94, 426–427. [Google Scholar]
- Liebig, C.; Ayala, G.; Wilks, J.A.; Berger, D.H.; Albo, D. Perineural Invasion in Cancer. Cancer 2009, 115, 3379–3391. [Google Scholar] [CrossRef]
- Gershenwald, J.E.; Scolyer, R.A.; Hess, K.R.; Sondak, V.K.; Long, G.V.; Ross, M.I.; Lazar, A.J.; Faries, M.B.; Kirkwood, J.M.; McArthur, G.A.; et al. Melanoma Staging: Evidence-Based Changes in the American Joint Committee on Cancer Eighth Edition Cancer Staging Manual. CA Cancer J. Clin. 2017, 67, 472–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanwar, M.; Branstetter, B.F., IV. Mimics of Perineural Tumor Spread in the Head and Neck. Br. J. Radiol. 2021, 94, 20210099. [Google Scholar] [CrossRef] [PubMed]
- Ronaghy, A.; Yaar, R.; Goldberg, L.J.; Mahalingam, M.; Bhawan, J. Perineural Involvement: What Does It Mean? Am. J. Dermatopathol. 2010, 32, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.-H.; Zhang, S.; Wang, H.; Xue, J.-L.; Zhang, Z.-G. Emerging Experimental Models for Assessing Perineural Invasion in Human Cancers. Cancer Lett. 2022, 535, 215610. [Google Scholar] [CrossRef] [PubMed]
- Rosa, S.L.; La Rosa, S.; Rigoli, E.; Uccella, S.; Novario, R.; Capella, C. Prognostic and Biological Significance of Cytokeratin 19 in Pancreatic Endocrine Tumours. Histopathology 2007, 50, 597–606. [Google Scholar] [CrossRef]
- Chatterjee, D.; Katz, M.H.; Rashid, A.; Wang, H.; Iuga, A.C.; Varadhachary, G.R.; Wolff, R.A.; Lee, J.E.; Pisters, P.W.; Crane, C.H.; et al. Perineural and Intraneural Invasion in Posttherapy Pancreaticoduodenectomy Specimens Predicts Poor Prognosis in Patients With Pancreatic Ductal Adenocarcinoma. Am. J. Surg. Pathol. 2012, 36, 409–417. [Google Scholar] [CrossRef] [Green Version]
- Bettschart, V.; Rahman, M.Q.; Engelken, F.J.F.; Madhavan, K.K.; Parks, R.W.; Garden, O.J. Presentation, Treatment and Outcome in Patients with Ampullary Tumours. Br. J. Surg. 2004, 91, 1600–1607. [Google Scholar] [CrossRef]
- Yamaguchi, R.; Nagino, M.; Oda, K.; Kamiya, J.; Uesaka, K.; Nimura, Y. Perineural Invasion Has a Negative Impact on Survival of Patients with Gallbladder Carcinoma. Br. J. Surg. 2002, 89, 1130–1136. [Google Scholar] [CrossRef]
- Murakami, Y.; Uemura, K.; Hayashidani, Y.; Sudo, T.; Ohge, H.; Sueda, T. Pancreatoduodenectomy for Distal Cholangiocarcinoma: Prognostic Impact of Lymph Node Metastasis. World J. Surg. 2007, 31, 337–342. [Google Scholar] [CrossRef]
- An, J.Y.; Baik, Y.H.; Choi, M.G.; Noh, J.H.; Sohn, T.S.; Kim, S. Predictive Factors for Lymph Node Metastasis in Early Gastric Cancer With Submucosal Invasion. Ann. Surg. 2007, 246, 749–753. [Google Scholar] [CrossRef]
- Ueno, H.; Shirouzu, K.; Eishi, Y.; Yamada, K.; Kusumi, T.; Kushima, R.; Ikegami, M.; Murata, A.; Okuno, K.; Sato, T.; et al. Characterization of Perineural Invasion As a Component of Colorectal Cancer Staging. Am. J. Surg. Pathol. 2013, 37, 1542–1549. [Google Scholar] [CrossRef]
- Gomez, D.; Zaitoun, A.M.; De Rosa, A.; Hossaini, S.; Beckingham, I.J.; Brooks, A.; Cameron, I.C. Critical Review of the Prognostic Significance of Pathological Variables in Patients Undergoing Resection for Colorectal Liver Metastases. HPB 2014, 16, 836–844. [Google Scholar] [CrossRef] [Green Version]
- Velazquez, E.F.; Ayala, G.; Liu, H.; Chaux, A.; Zanotti, M.; Torres, J.; Cho, S.I.; Barreto, J.E.; Soares, F.; Cubilla, A.L. Histologic Grade and Perineural Invasion Are More Important Than Tumor Thickness as Predictor of Nodal Metastasis in Penile Squamous Cell Carcinoma Invading 5 to 10 Mm. Am. J. Surg. Pathol. 2008, 32, 974–979. [Google Scholar] [CrossRef]
- Kang, M.; Oh, J.J.; Lee, S.; Hong, S.K.; Lee, S.E.; Byun, S.-S. Perineural Invasion and Lymphovascular Invasion Are Associated with Increased Risk of Biochemical Recurrence in Patients Undergoing Radical Prostatectomy. Ann. Surg. Oncol. 2016, 23, 2699–2706. [Google Scholar] [CrossRef]
- Nagarajan, P.; Piao, J.; Ning, J.; Noordenbos, L.E.; Curry, J.L.; Torres-Cabala, C.A.; Hafeez Diwan, A.; Ivan, D.; Aung, P.P.; Ross, M.I.; et al. Prognostic Model for Patient Survival in Primary Anorectal Mucosal Melanoma: Stage at Presentation Determines Relevance of Histopathologic Features. Mod. Pathol. 2020, 33, 496–513. [Google Scholar] [CrossRef]
- Nagarajan, P.; Curry, J.L.; Ning, J.; Piao, J.; Torres-Cabala, C.A.; Aung, P.P.; Ivan, D.; Ross, M.I.; Levenback, C.F.; Frumovitz, M.; et al. Tumor Thickness and Mitotic Rate Robustly Predict Melanoma-Specific Survival in Patients with Primary Vulvar Melanoma: A Retrospective Review of 100 Cases. Clin. Cancer Res. 2017, 23, 2093–2104. [Google Scholar] [CrossRef] [Green Version]
- Baer, S.C.; Schultz, D.; Synnestvedt, M.; Elder, D.E. Desmoplasia and Neurotropism. Prognostic Variables in Patients with Stage I Melanoma. Cancer 1995, 76, 2242–2247. [Google Scholar] [CrossRef]
- Stafford, N.D. Head and Neck Cancer: A Multidisciplinary Approach; Harrison, L.B., Sessions, R.B., Hong, W.K., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 1999; ISBN 0-397-51777-7. [Google Scholar]
- Ali, S. A Predictive Nomogram for Recurrence of Carcinoma of the Major Salivary GlandsNomogram for Salivary Gland Carcinoma Recurrence. JAMAOtolaryngol.–Head Neck Surg. 2013, 139, 698–705. [Google Scholar]
- García, M.P.P.; Pérez García, M.P.; Mateu Puchades, A.; Sanmartín Jiménez, O. Perineural Invasion in Cutaneous Squamous Cell Carcinoma. Actas Dermo-Sifiliográficas (Engl.Ed.) 2019, 110, 426–433. [Google Scholar] [CrossRef]
- Schuknecht, B. Patterns of Perineural Skull Base Tumor Extension from Extracranial Tumors. Neuroimaging Clin. N. Am. 2021, 31, 473–483. [Google Scholar] [CrossRef]
- Medvedev, O.; Hedesiu, M.; Ciurea, A.; Lenghel, M.; Rotar, H.; Dinu, C.; Roman, R.; Termure, D.; Csutak, C. Perineural Spread in Head and Neck Malignancies: Imaging Findings—An Updated Literature Review. Bosn. J. Basic Med. Sci. 2021, 22, 22–38. [Google Scholar] [CrossRef]
- Schachtel, M.J.C.; Gandhi, M.; Bowman, J.J.; Porceddu, S.V.; Panizza, B.J. Facial Nerve Perineural Spread from Cutaneous Squamous Cell Carcinoma of the Head and Neck: A Single Institution Analysis of Epidemiology, Treatment, Survival Outcomes, and Prognostic Factors. Head Neck 2022, 44, 1223–1236. [Google Scholar] [CrossRef]
- Miller, M.E.; Palla, B.; Chen, Q.; Elashoff, D.A.; Abemayor, E.; St. John, M.A.; Lai, C.K. A Novel Classification System for Perineural Invasion in Noncutaneous Head and Neck Squamous Cell Carcinoma: Histologic Subcategories and Patient Outcomes. Am. J. Otolaryngol. 2012, 33, 212–215. [Google Scholar] [CrossRef]
- Carter, J.B.; Johnson, M.M.; Chua, T.L.; Karia, P.S.; Schmults, C.D. Outcomes of Primary Cutaneous Squamous Cell Carcinoma With Perineural Invasion. JAMA Dermatol. 2013, 149, 35. [Google Scholar] [CrossRef] [Green Version]
- Karia, P.S.; Jambusaria-Pahlajani, A.; Harrington, D.P.; Murphy, G.F.; Qureshi, A.A.; Schmults, C.D. Evaluation of American Joint Committee on Cancer, International Union Against Cancer, and Brigham and Women’s Hospital Tumor Staging for Cutaneous Squamous Cell Carcinoma. J. Clin. Oncol. 2014, 32, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Califano, J.A.; Lydiatt, W.M.; Nehal, K.S.; O’Sullivan, B.; Schmults, C.; Seethala, R.R.; Weber, R.S.; Shah, J.P. Cutaneous Carcinoma of the Head and Neck. In AJCC Cancer Staging Manual; Baishideng Publishing Group: Pleasanton, CA, USA, 2017; pp. 171–181. [Google Scholar]
- Amin, M.B.; Greene, F.L.; Edge, S.B.; Compton, C.C.; Gershenwald, J.E.; Brookland, R.K.; Meyer, L.; Gress, D.M.; Byrd, D.R.; Winchester, D.P. The Eighth Edition AJCC Cancer Staging Manual: Continuing to Build a Bridge from a Population-Based to a More “personalized” Approach to Cancer Staging. CA Cancer J. Clin. 2017, 67, 93–99. [Google Scholar] [CrossRef]
- Schmitd, L.B.; Perez-Pacheco, C.; D’Silva, N.J. Nerve Density in Cancer: Less Is Better. FASEB BioAdv. 2021, 3, 773–786. [Google Scholar] [CrossRef]
- Wang, H.; Zheng, Q.; Lu, Z.; Wang, L.; Ding, L.; Xia, L.; Zhang, H.; Wang, M.; Chen, Y.; Li, G. Role of the Nervous System in Cancers: A Review. Cell Death Discov. 2021, 7, 76. [Google Scholar] [CrossRef]
- Saloman, J.L.; Albers, K.M.; Rhim, A.D.; Davis, B.M. Can Stopping Nerves, Stop Cancer? Trends Neurosci. 2016, 39, 880–889. [Google Scholar] [CrossRef] [Green Version]
- Liebig, C.; Ayala, G.; Wilks, J.; Verstovsek, G.; Liu, H.; Agarwal, N.; Berger, D.H.; Albo, D. Perineural Invasion Is an Independent Predictor of Outcome in Colorectal Cancer. J. Clin. Oncol. 2009, 27, 5131–5137. [Google Scholar] [CrossRef] [Green Version]
- Chakroborty, D.; Sarkar, C.; Basu, B.; Dasgupta, P.S.; Basu, S. Catecholamines Regulate Tumor Angiogenesis. Cancer Res. 2009, 69, 3727–3730. [Google Scholar] [CrossRef] [Green Version]
- Fiszman, G.L.; Middonno, M.C.; de la Torre, E.; Farina, M.; Español, A.J.; Sales, M.E. Activation of Muscarinic Cholinergic Receptors Induces MCF-7 Cells Proliferation and Angiogenesis by Stimulating Nitric Oxide Synthase Activity. Cancer Biol. Ther. 2007, 6, 1106–1113. [Google Scholar] [CrossRef] [Green Version]
- Romon, R.; Adriaenssens, E.; Lagadec, C.; Germain, E.; Hondermarck, H.; Le Bourhis, X. Nerve Growth Factor Promotes Breast Cancer Angiogenesis by Activating Multiple Pathways. Mol. Cancer 2010, 9, 157. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-Y.; Hung, S.-Y.; Chen, H.-T.; Tsou, H.-K.; Fong, Y.-C.; Wang, S.-W.; Tang, C.-H. Brain-Derived Neurotrophic Factor Increases Vascular Endothelial Growth Factor Expression and Enhances Angiogenesis in Human Chondrosarcoma Cells. Biochem. Pharmacol. 2014, 91, 522–533. [Google Scholar] [CrossRef]
- Eichmann, A.; Brunet, I. Arterial Innervation in Development and Disease. Sci. Transl. Med. 2014, 6, 252ps9. [Google Scholar] [CrossRef]
- Rosas-Ballina, M.; Olofsson, P.S.; Ochani, M.; Valdés-Ferrer, S.I.; Levine, Y.A.; Reardon, C.; Tusche, M.W.; Pavlov, V.A.; Andersson, U.; Chavan, S.; et al. Acetylcholine-Synthesizing T Cells Relay Neural Signals in a Vagus Nerve Circuit. Science 2011, 334, 98–101. [Google Scholar] [CrossRef] [Green Version]
- Friedman, J.R.; Richbart, S.D.; Merritt, J.C.; Brown, K.C.; Nolan, N.A.; Akers, A.T.; Lau, J.K.; Robateau, Z.R.; Miles, S.L.; Dasgupta, P. Acetylcholine Signaling System in Progression of Lung Cancers. Pharmacol. Ther. 2019, 194, 222–254. [Google Scholar] [CrossRef]
- Hristova, V.A.; Chan, D.W. Cancer Biomarker Discovery and Translation: Proteomics and beyond. Expert Rev. Proteom. 2019, 16, 93–103. [Google Scholar] [CrossRef]
- Aquino-Nunez, W.; Mielko, Z.E.; Dunn, T.; Santorella, E.M.; Hosea, C.; Leitner, L.; McCalla, D.; Simms, C.; Verola, W.M.; Vijaykumar, S.; et al. Cnd-1/NeuroD1 Functions with the Homeobox Gene Ceh-5/Vax2 and Hox Gene Ceh-13/labial To Specify Aspects of RME and DD Neuron Fate in Caenorhabditis Elegans. G3 2020, 10, 3071–3085. [Google Scholar] [CrossRef]
- Huang, P.; Kishida, S.; Cao, D.; Murakami-Tonami, Y.; Mu, P.; Nakaguro, M.; Koide, N.; Takeuchi, I.; Onishi, A.; Kadomatsu, K. The Neuronal Differentiation Factor NeuroD1 Downregulates the Neuronal Repellent Factor Slit2 Expression and Promotes Cell Motility and Tumor Formation of Neuroblastoma. Cancer Res. 2011, 71, 2938–2948. [Google Scholar] [CrossRef] [Green Version]
- Kersigo, J.; Gu, L.; Xu, L.; Pan, N.; Vijayakuma, S.; Jones, T.; Shibata, S.B.; Fritzsch, B.; Hansen, M.R. Effects of Neurod1 Expression on Mouse and Human Schwannoma Cells. Laryngoscope 2021, 131, E259–E270. [Google Scholar] [CrossRef]
- Person, F.; Wilczak, W.; Hube-Magg, C.; Burdelski, C.; Möller-Koop, C.; Simon, R.; Noriega, M.; Sauter, G.; Steurer, S.; Burdak-Rothkamm, S.; et al. Prevalence of βIII-Tubulin (TUBB3) Expression in Human Normal Tissues and Cancers. Tumour Biol. 2017, 39, 1010428317712166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsetos, C.D.; Herman, M.M.; Mörk, S.J. Class III Beta-Tubulin in Human Development and Cancer. Cell Motil. Cytoskeleton 2003, 55, 77–96. [Google Scholar] [CrossRef] [PubMed]
- Nemunaitis, J. Stathmin 1: A Protein with Many Tasks. New Biomarker and Potential Target in Cancer. Expert Opin. Ther. Targets 2012, 16, 631–634. [Google Scholar] [CrossRef] [Green Version]
- Fife, C.M.; Sagnella, S.M.; Teo, W.S.; Po’uha, S.T.; Byrne, F.L.; Yeap, Y.Y.C.; Ng, D.C.H.; Davis, T.P.; McCarroll, J.A.; Kavallaris, M. Stathmin Mediates Neuroblastoma Metastasis in a Tubulin-Independent Manner via RhoA/ROCK Signaling and Enhanced Transendothelial Migration. Oncogene 2017, 36, 501–511. [Google Scholar] [CrossRef]
- Buddle, M.; Eberhardt, E.; Ciminello, L.H.; Levin, T.; Wing, R.; DiPasquale, K.; Raley-Susman, K.M. Microtubule-Associated Protein 2 (MAP2) Associates with the NMDA Receptor and Is Spatially Redistributed within Rat Hippocampal Neurons after Oxygen-Glucose Deprivation. Brain Res. 2003, 978, 38–50. [Google Scholar] [CrossRef]
- Liu, Y.; Sturgis, C.D.; Grzybicki, D.M.; Jasnosz, K.M.; Olson, P.R.; Tong, M.; Dabbs, D.D.; Raab, S.S.; Silverman, J.F. Microtubule-Associated Protein-2: A New Sensitive and Specific Marker for Pulmonary Carcinoid Tumor and Small Cell Carcinoma. Mod. Pathol. 2001, 14, 880–885. [Google Scholar] [CrossRef] [Green Version]
- Molenaar, W.M.; Baker, D.L.; Pleasure, D.; Lee, V.M.; Trojanowski, J.Q. The Neuroendocrine and Neural Profiles of Neuroblastomas, Ganglioneuroblastomas, and Ganglioneuromas. Am. J. Pathol. 1990, 136, 375–382. [Google Scholar]
- Krishnan, C.; Higgins, J.P.; West, R.B.; Natkunam, Y.; Heerema-McKenney, A.; Arber, D.A. Microtubule-Associated Protein-2 Is a Sensitive Marker of Primary and Metastatic Neuroblastoma. Am. J. Surg. Pathol. 2009, 33, 1695–1704. [Google Scholar] [CrossRef]
- Miettinen, M.; Rapola, J. Synaptophysin--an Immuno-Histochemical Marker for Childhood Neuroblastoma. Acta Pathol. Microbiol. Immunol. Scand. A 1987, 95, 167–170. [Google Scholar] [CrossRef]
- Magnon, C.; Hall, S.J.; Lin, J.; Xue, X.; Gerber, L.; Freedland, S.J.; Frenette, P.S. Autonomic Nerve Development Contributes to Prostate Cancer Progression. Science 2013, 341, 1236361. [Google Scholar]
- Ayala, G.E.; Dai, H.; Powell, M.; Li, R.; Ding, Y.; Wheeler, T.M.; Shine, D.; Kadmon, D.; Thompson, T.; Miles, B.J.; et al. Cancer-Related Axonogenesis and Neurogenesis in Prostate Cancer. Clin. Cancer Res. 2008, 14, 7593–7603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowe, C.W.; Dill, T.; Griffin, N.; Jobling, P.; Faulkner, S.; Paul, J.W.; King, S.; Smith, R.; Hondermarck, H. Innervation of Papillary Thyroid Cancer and Its Association with Extra-Thyroidal Invasion. Sci. Rep. 2020, 10, 1539. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Yang, Y.; Liang, X.; Du, G.; Liu, L.; Lu, L.; Dong, J.; Han, H.; Zhang, G. The Clinicopathological Significance of Neurogenesis in Breast Cancer. BMC Cancer 2014, 14, 484. [Google Scholar]
- Farschtschi, S.C.; Kluwe, L.; Schön, G.; Friedrich, R.E.; Matschke, J.; Glatzel, M.; Weis, J.; Hagel, C.; Mautner, V.-F. Distinctive Low Epidermal Nerve Fiber Density in Schwannomatosis Patients Provides a Major Parameter for Diagnosis and Differential Diagnosis. Brain Pathol. 2020, 30, 386–391. [Google Scholar] [CrossRef] [Green Version]
- Tagliafico, A.; Tagliafico, G.; Martinoli, C. Nerve Density: A New Parameter to Evaluate Peripheral Nerve Pathology on Ultrasound. Preliminary Study. Ultrasound Med. Biol. 2010, 36, 1588–1593. [Google Scholar] [CrossRef]
- Melo, Z.; Castillo, X.; Moreno-Carranza, B.; Ledesma-Colunga, M.G.; Arnold, E.; López-Casillas, F.; Ruíz-Herrera, X.; Clapp, C.; Martínez de la Escalera, G. Vasoinhibin Suppresses Nerve Growth Factor-Induced Differentiation and Survival of PC12 Pheochromocytoma Cells. Neuroendocrinology 2019, 109, 152–164. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, S.R.; Jordan, M.; Nagarajan, P.; Amit, M. Nerve Density and Neuronal Biomarkers in Cancer. Cancers 2022, 14, 4817. https://doi.org/10.3390/cancers14194817
Ali SR, Jordan M, Nagarajan P, Amit M. Nerve Density and Neuronal Biomarkers in Cancer. Cancers. 2022; 14(19):4817. https://doi.org/10.3390/cancers14194817
Chicago/Turabian StyleAli, Shahrukh R., Madeleine Jordan, Priyadharsini Nagarajan, and Moran Amit. 2022. "Nerve Density and Neuronal Biomarkers in Cancer" Cancers 14, no. 19: 4817. https://doi.org/10.3390/cancers14194817
APA StyleAli, S. R., Jordan, M., Nagarajan, P., & Amit, M. (2022). Nerve Density and Neuronal Biomarkers in Cancer. Cancers, 14(19), 4817. https://doi.org/10.3390/cancers14194817