

DGKα, Bridging Membrane Shape Changes with Specific Molecular Species of DAG/PA: Implications in Cancer and Immunosurveillance


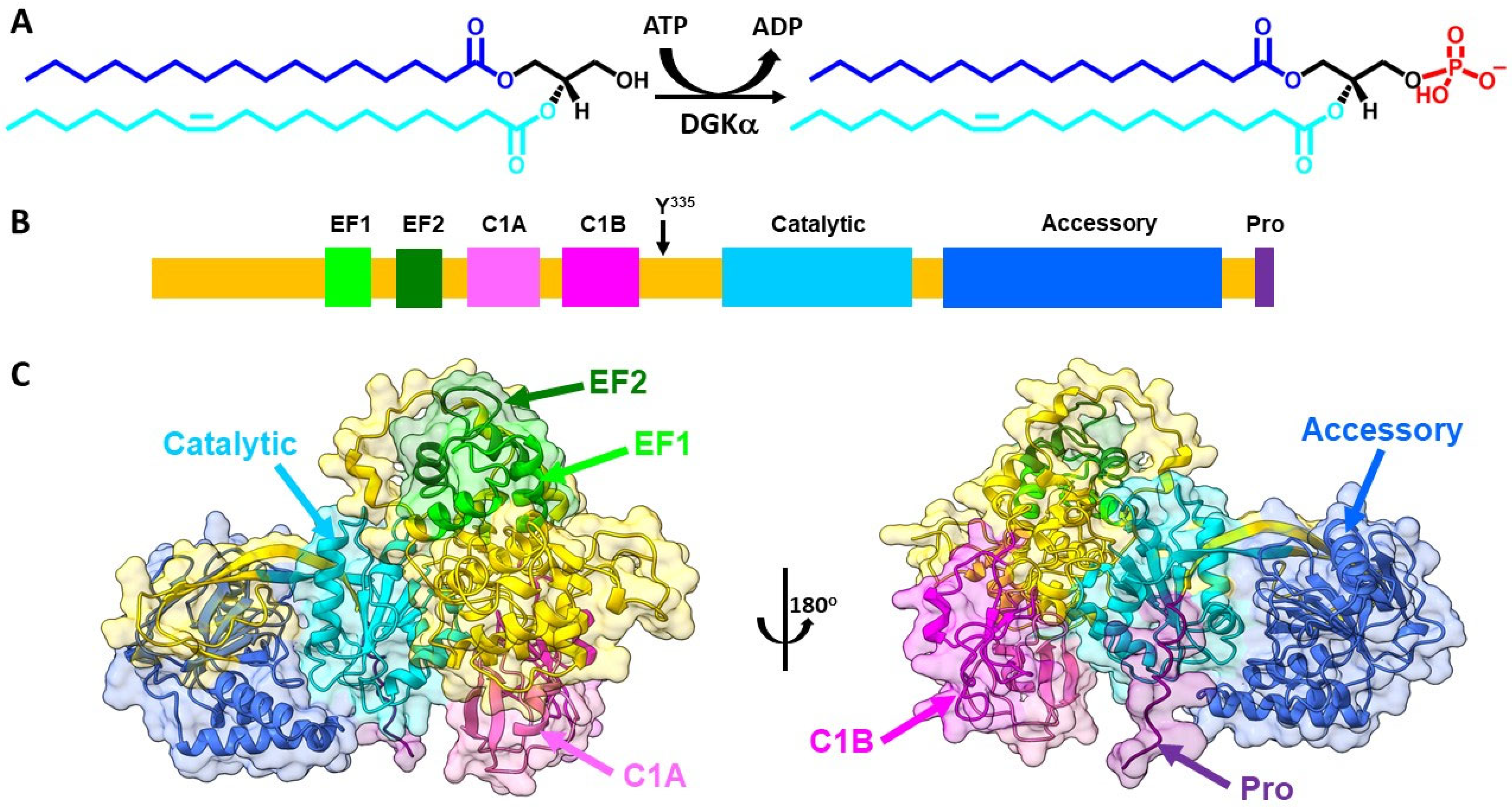{kind=link}
{kind=link}
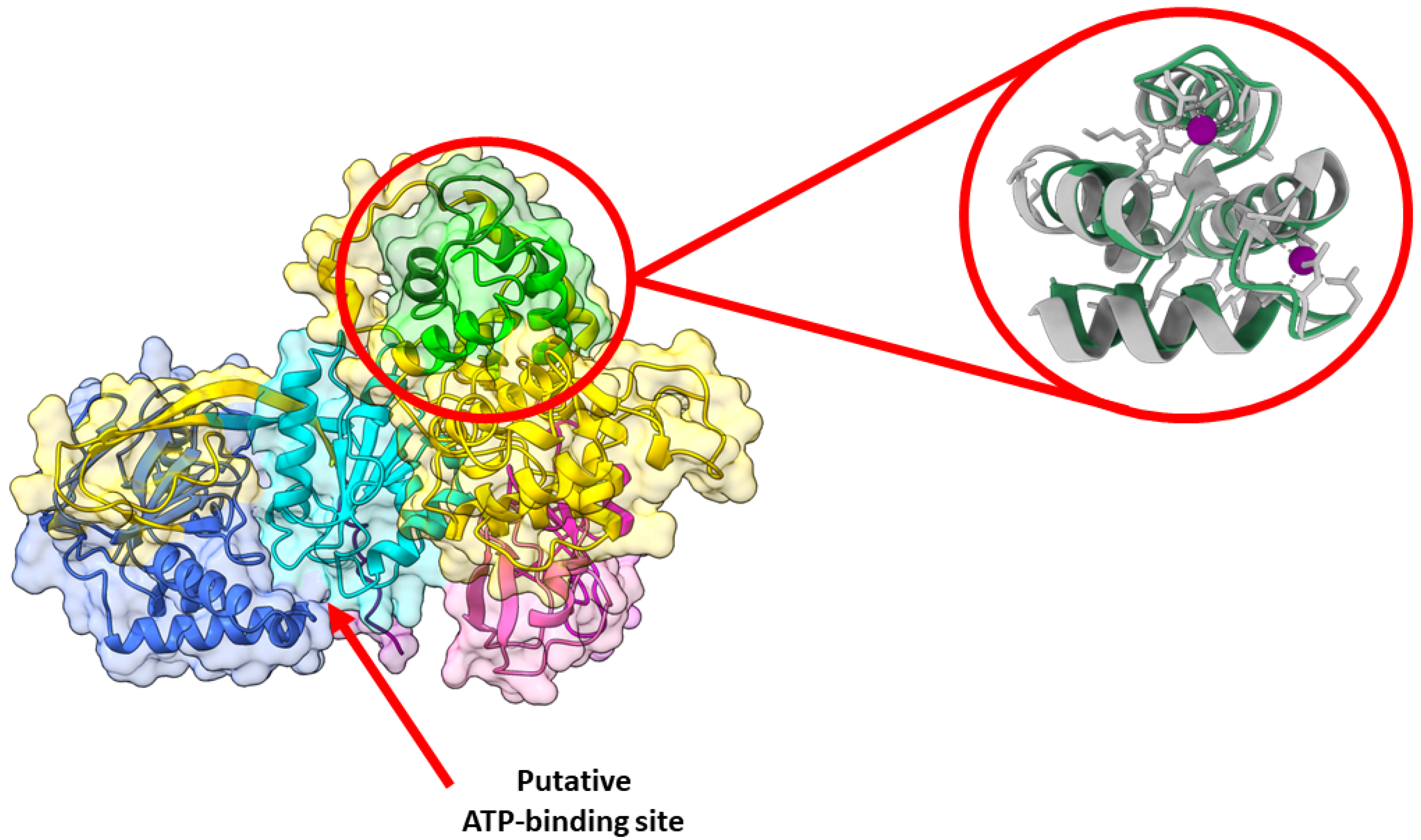{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. DGKα
2.1. Enzymatic Reaction and Substrate Acyl Chain Specificity
2.2. Structural Properties
2.3. Subcellular Localization
2.4. Modulation of DGKα by Membrane Physicochemical Properties
3. DAG and PA: Intermediates of Lipid Metabolism, Membrane Curvature Generators and Signaling Molecules
4. DGKα in Cell Biology
4.1. T Cells
4.2. Cancer Cells
5. Membrane Morphology Changes during Cancer and T Cell Biological Processes: Implications for DGKα Enzymatic Properties
5.1. TCR Activation
5.2. T Cell Anergy
5.3. Cancer Cell Migration
5.4. Nuclear Envelope
6. Specificity of DGKα Effectors Dependent on DAG/PA Molecular Species
7. Therapeutic Approaches Targeting DGKα in Cancer and Immunosurveillance
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AICD | activation-induced cell death |
| APC | antigen-presenting cell |
| Arf1 | ADP-ribosylation factor 1 |
| CU-3 | 5-[(2E)-3-(2-furyl)prop-2-enylidene]-3-[(phenylsulfonyl)amino]2-thioxo-1,3-thiazolidin-4-one) |
| DAG | diacylglycerol |
| DGK | diacylglycerol kinases |
| DGKα | the α paralog of human DGK |
| DGKε | the ε paralog of human DGK |
| ECM | extracellular matrix |
| ERK | extracellular signal-regulated kinase |
| FasL | Fas ligand |
| HIF1α | hypoxia-inducible factor 1-α |
| IL2 | interleukin 2 |
| IS | immunological synapse |
| Lck | lymphocyte-specific protein tyrosine kinase |
| LFA-1 | lymphocyte function-associated antigen-1 |
| MAPK | mitogen-activated protein kinase |
| MHC | major histocompatibility complex |
| MTOC | microtubule organizing center |
| mTOR | mammalian target of rapamycin |
| MVB | multivesicular bodies |
| NF-kB | nuclear factor kappa-light chain enhancer of activated B cells |
| NPC | nuclear pore complex |
| PA | phosphatidic acid |
| PC | phosphatidylcholine |
| PE | phosphatidylethanolamine |
| PI | phosphatidylinositol |
| PI-3,4-P2 | phosphatidylinositol-3,4-bisphosphate |
| PI-3,4,5-P3 | phosphatidylinositol-3,4,5-triphosphate |
| PI3K | phosphatidylinositol-3-kinase |
| PIP2 | phosphatidylinositol-4,5-biphosphate |
| PIPn | phosphorylated forms of phosphatidylinositol |
| PKC | protein kinase C |
| PKD | protein kinase D |
| PLC | phospholipase C |
| PM | plasma membrane |
| PS | phosphatidylserine |
| SMAC | supramolecular activation cluster |
| Rab11-FIP1 | Rab11 family of interacting protein 1 |
| Rac1 | Ras-related C3 botulinum toxin substrate 1 |
| RasGAP | Ras GTPase activating protein |
| RasGRP1 | a Ras guanyl nucleotide-releasing protein |
| RCP | Rab-coupling protein |
| Src | proto-oncogene tyrosine protein kinase |
| TCR | T-cell receptor |
| TGN | trans-Golgi network |
| TNFα | tumor necrosis factor α |
References
- Goldberg, M.S. Improving cancer immunotherapy through nanotechnology. Nat. Cancer 2019, 19, 587–602. [Google Scholar] [CrossRef]
- Waldman, A.D.; Fritz, J.M.; Lenardo, M.J. A guide to cancer immunotherapy: From T cell basic science to clinical practice. Nat. Rev. Immunol. 2020, 20, 651–668. [Google Scholar] [CrossRef]
- Johnson, D.B.; Nebhan, C.A.; Moslehi, J.J.; Balko, J.M. Immune-checkpoint inhibitors: Long-term implications of toxicity. Nat. Rev. Clin. Oncol. 2022, 19, 254–267. [Google Scholar] [CrossRef]
- Kubli, S.P.; Berger, T.; Araujo, D.V.; Siu, L.L.; Mak, T.W. Beyond immune checkpoint blockade: Emerging immunological strategies. Nat. Rev. Drug Discov. 2021, 20, 899–919. [Google Scholar] [CrossRef]
- Korman, A.J.; Garrett-Thomson, S.C.; Lonberg, N. The foundations of immune checkpoint blockade and the ipilimumab approval decennial. Nat. Rev. Drug Discov. 2021, 21, 509–528. [Google Scholar] [CrossRef]
- Buchbinder, E.I.; Hodi, F.S. Immune-checkpoint blockade—Durable cancer control. Nat. Rev. Clin. Oncol. 2016, 13, 77–78. [Google Scholar] [CrossRef]
- Noessner, E. DGK-α: A Checkpoint in Cancer-Mediated Immuno-Inhibition and Target for Immunotherapy. Front. Cell Dev. Biol. 2017, 5, 16. [Google Scholar] [CrossRef] [Green Version]
- Purow, B. Molecular Pathways: Targeting Diacylglycerol Kinase Alpha in Cancer. Clin. Cancer Res. 2015, 21, 5008–5012. [Google Scholar] [CrossRef] [Green Version]
- Mérida, I.; Torres-Ayuso, P.; Ávila-Flores, A.; Arranz-Nicolás, J.; Andrada, E.; Tello-Lafoz, M.; Liébana, R.; Arcos, R. Diacylglycerol kinases in cancer. Adv. Biol. Regul. 2017, 63, 22–31. [Google Scholar] [CrossRef]
- Yanagisawa, K.; Yasuda, S.; Kai, M.; Imai, S.-I.; Yamada, K.; Yamashita, T.; Jimbow, K.; Kanoh, H.; Sakane, F. Diacylglycerol kinase α suppresses tumor necrosis factor-α-induced apoptosis of human melanoma cells through NF-κB activation. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2007, 1771, 462–474. [Google Scholar] [CrossRef]
- Torres-Ayuso, P.; Daza-Martín, M.; Martín-Pérez, J.; Ávila-Flores, A.; Mérida, I. Diacylglycerol kinase α promotes 3D cancer cell growth and limits drug sensitivity through functional interaction with Src. Oncotarget 2014, 5, 9710–9726. [Google Scholar] [CrossRef] [Green Version]
- Bacchiocchi, R.; Baldanzi, G.; Carbonari, D.; Capomagi, C.; Colombo, E.; Van Blitterswijk, W.J.; Graziani, A.; Fazioli, F. Activation of -diacylglycerol kinase is critical for the mitogenic properties of anaplastic lymphoma kinase. Blood 2005, 106, 2175–2182. [Google Scholar] [CrossRef] [Green Version]
- Takeishi, K.; Taketomi, A.; Shirabe, K.; Toshima, T.; Motomura, T.; Ikegami, T.; Yoshizumi, T.; Sakane, F.; Maehara, Y. Diacylglycerol kinase alpha enhances hepatocellular carcinoma progression by activation of Ras–Raf–MEK–ERK pathway. J. Hepatol. 2012, 57, 77–83. [Google Scholar] [CrossRef]
- Dominguez, C.; Floyd, D.; Xiao, A.; Mullins, G.; Kefas, B.; Xin, W.; Yacur, M.; Abounader, R.; Lee, J.; Wilson, G.; et al. Diacylglycerol Kinase α Is a Critical Signaling Node and Novel Therapeutic Target in Glioblastoma and Other Cancers. Cancer Discov. 2013, 3, 782–797. [Google Scholar] [CrossRef] [Green Version]
- Zha, Y.; Marks, R.; Ho, A.W.; Peterson, A.C.; Janardhan, S.; Brown, I.; Praveen, K.; Stang, S.; Stone, J.C.; Gajewski, T.F. T cell anergy is reversed by active Ras and is regulated by diacylglycerol kinase-α. Nat. Immunol. 2006, 7, 1166–1173. [Google Scholar] [CrossRef]
- Olenchock, B.A.; Guo, R.; Carpenter, J.H.; Jordan, M.; Topham, M.K.; Koretzky, G.A.; Zhong, X.P. Disruption of Diacylglycerol Metabolism Impairs the Induction of T Cell Anergy. Nat. Immunol. 2006, 7, 1174–1181. [Google Scholar] [CrossRef]
- Sanjuán, M.A.; Jones, D.R.; Izquierdo, M.; Mérida, I. Role of Diacylglycerol Kinase Alpha in the Attenuation of Receptor Signaling. J. Cell Biol. 2001, 153, 207–219. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.R.; Sanjuán, M.A.; Stone, J.C.; Mérida, I. Expression of a Catalytically Inactive Form of Diacylglycerol Kinase Alpha Induces Sustained Signaling through RasGRP. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2002, 16, 595–597. [Google Scholar] [CrossRef]
- Murakami, Y.; Murakami, C.; Hoshino, F.; Lu, Q.; Akiyama, R.; Yamaki, A.; Takahashi, D.; Sakane, F. Palmitic Acid- and/or Palmitoleic Acid-Containing Phosphatidic Acids Are Generated by Diacylglycerol Kinase α in Starved Jurkat T Cells. Biochem. Biophys. Res. Commun. 2020, 525, 1054–1060. [Google Scholar] [CrossRef]
- Sakane, F.; Hoshino, F.; Murakami, C. New Era of Diacylglycerol Kinase, Phosphatidic Acid and Phosphatidic Acid-Binding Protein. Int. J. Mol. Sci. 2020, 21, 6794. [Google Scholar] [CrossRef]
- Yamaki, A.; Akiyama, R.; Murakami, C.; Takao, S.; Murakami, Y.; Mizuno, S.; Takahashi, D.; Kado, S.; Taketomi, A.; Shirai, Y.; et al. Diacylglycerol Kinase α-Selective Inhibitors Induce Apoptosis and Reduce Viability of Melanoma and Several Other Cancer Cell Lines. J. Cell. Biochem. 2019, 120, 10043–10056. [Google Scholar] [CrossRef] [PubMed]
- Ware, T.B.; Franks, C.E.; Granade, M.E.; Zhang, M.; Kim, K.-B.; Park, K.-S.; Gahlmann, A.; Harris, T.E.; Hsu, K.-L. Reprogramming Fatty Acyl Specificity of Lipid Kinases via C1 Domain Engineering. Nat. Chem. Biol. 2020, 16, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Bozelli, J.C.; Yune, J.; Takahashi, D.; Sakane, F.; Epand, R.M. Membrane Morphology Determines Diacylglycerol Kinase α Substrate Acyl Chain Specificity. FASEB J. 2021, 35, e21602. [Google Scholar] [CrossRef] [PubMed]
- Shulga, Y.; Topham, M.; Epand, R. Regulation and Functions of Diacylglycerol Kinases. Chem. Rev. 2011, 111, 6186–6208. [Google Scholar] [CrossRef] [PubMed]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively Expanding the Structural Coverage of Protein-Sequence Space with High-Accuracy Models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.; Goddard, T.; Huang, C.; Couch, G.; Greenblatt, D.; Meng, E.; Ferrin, T. UCSF Chimera-a Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-Based Map of the Human Proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Sakane, F.; Yamada, K.; Imai, S.I.; Kanoh, H. Porcine 80-KDa Diacylglycerol Kinase Is a Calcium-Binding and Calcium/Phospholipid-Dependent Enzyme and Undergoes Calcium-Dependent Translocation. J. Biol. Chem. 1991, 266, 7096–7100. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Aulakh, S.S.; Bozelli, J.C., Jr.; Epand, R.M. Exploring the AlphaFold Predicted Conformational Properties of Human Diacylglycerol Kinases. J. Phys. Chem. B 2022, 126, 7172–7183. [Google Scholar] [CrossRef]
- Yamada, K.; Sakane, F.; Matsushima, N.; Kanoh, H. EF-Hand Motifs of Alpha, Beta and Gamma Isoforms of Diacylglycerol Kinase Bind Calcium with Different Affinities and Conformational Changes. Biochem. J. 1997, 321, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, D.; Suzuki, K.; Sakamoto, T.; Iwamoto, T.; Murata, T.; Sakane, F. Crystal Structure and Calcium-Induced Conformational Changes of Diacylglycerol Kinase α EF-Hand Domains. Protein Sci. 2019, 28, 694–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, Scalable Generation of High-Quality Protein Multiple Sequence Alignments Using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Jurrus, E.; Engel, D.; Star, K.; Monson, K.; Brandi, J.; Felberg, L.E.; Brookes, D.H.; Wilson, L.; Chen, J.; Liles, K.; et al. Improvements to the APBS Biomolecular Solvation Software Suite. Protein Sci. 2018, 27, 112–128. [Google Scholar] [CrossRef] [Green Version]
- Kochnev, Y.; Hellemann, E.; Cassidy, K.C.; Durrant, J.D. Webina: An Open-Source Library and Web App That Runs AutoDock Vina Entirely in the Web Browser. Bioinformatics 2020, 36, 4513–4515. [Google Scholar] [CrossRef] [PubMed]
- Flores, I.; Casaseca, T.; Martinez-A, C.; Kanoh, H.; Merida, I. Phosphatidic Acid Generation through Interleukin 2 (IL-2)-Induced Alpha-Diacylglycerol Kinase Activation Is an Essential Step in IL-2-Mediated Lymphocyte Proliferation. J. Biol. Chem. 1996, 271, 10334–10340. [Google Scholar] [CrossRef] [Green Version]
- Sanjuán, M.A.; Pradet-Balade, B.; Jones, D.R.; Martínez-A, C.; Stone, J.C.; Garcia-Sanz, J.A.; Mérida, I. T Cell Activation in Vivo Targets Diacylglycerol Kinase Alpha to the Membrane: A Novel Mechanism for Ras Attenuation. J. Immunol. 2003, 170, 2877–2883. [Google Scholar] [CrossRef] [Green Version]
- Schaap, D.; van der Wal, J.; van Blitterswijk, W.J.; van der Bend, R.L.; Ploegh, H.L. Diacylglycerol Kinase Is Phosphorylated in Vivo upon Stimulation of the Epidermal Growth Factor Receptor and Serine/Threonine Kinases, Including Protein Kinase C-Epsilon. Biochem. J. 1993, 289, 875–881. [Google Scholar] [CrossRef]
- Cutrupi, S.; Baldanzi, G.; Gramaglia, D.; Maffe, A.; Schaap, D.; Giraudo, E.; Van Blitterswijk, W.J.; Bussolino, F.; Comoglio, P.M.; Graziani, A. Src-mediated activation of α-diacylglycerol kinase is required for hepatocyte growth factor-induced cell motility. EMBO J. 2000, 19, 4614–4622. [Google Scholar] [CrossRef] [Green Version]
- Merino, E.; Sanjuán, M.A.; Moraga, I.; Ciprés, A.; Mérida, I. Role of the Diacylglycerol Kinase Alpha-Conserved Domains in Membrane Targeting in Intact T Cells. J. Biol. Chem. 2007, 282, 35396–35404. [Google Scholar] [CrossRef]
- Moraga, I.; Saito, N.; Mérida, I.; Merino, E.; Ávila-Flores, A.; Shirai, Y. Membrane Association in T Cells Regulates Its α Diacylglycerol Kinase Lck-Dependent Tyrosine Phosphorylation Of. J. Immunol. Ref. 2008, 180, 5805–5815. [Google Scholar] [CrossRef] [Green Version]
- Ciprés, A.; Carrasco, S.; Merino, E.; Díaz, E.; Krishna, U.M.; Falck, J.R.; Martínez-A, C.; Mérida, I. Regulation of Diacylglycerol Kinase α by Phosphoinositide 3-Kinase Lipid Products. J. Biol. Chem. 2003, 278, 35629–35635. [Google Scholar] [CrossRef] [Green Version]
- Baldanzi, G.; Mitola, S.; Cutrupi, S.; Filigheddu, N.; van Blitterswijk, W.J.; Sinigaglia, F.; Bussolino, F.; Graziani, A. Activation of Diacylglycerol Kinase α Is Required for VEGF-Induced Angiogenic Signaling in Vitro. Oncogene 2004, 23, 4828–4838. [Google Scholar] [CrossRef] [Green Version]
- Alonso, R.; Mazzeo, C.; Rodriguez, M.C.; Marsh, M.; Fraile-Ramos, A.; Calvo, V.; Avila-Flores, A.; Merida, I.; Izquierdo, M. Diacylglycerol Kinase α Regulates the Formation and Polarisation of Mature Multivesicular Bodies Involved in the Secretion of Fas Ligand-Containing Exosomes in T Lymphocytes. Cell Death Differ. 2011, 18, 1161–1173. [Google Scholar] [CrossRef] [Green Version]
- Alonso, R.; Mazzeo, C.; Mérida, I.; Izquierdo, M. A New Role of Diacylglycerol Kinase α on the Secretion of Lethal Exosomes Bearing Fas Ligand during Activation-Induced Cell Death of T Lymphocytes. Biochimie 2007, 89, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Alonso, R.; Rodríguez, M.C.; Pindado, J.; Merino, E.; Mérida, I.; Izquierdo, M. Diacylglycerol Kinase α Regulates the Secretion of Lethal Exosomes Bearing Fas Ligand during Activation-induced Cell Death of T Lymphocytes. J. Biol. Chem. 2005, 280, 28439–28450. [Google Scholar] [CrossRef] [Green Version]
- Lomize, A.L.; Todd, S.C.; Pogozheva, I.D. Spatial Arrangement of Proteins in Planar and Curved Membranes by PPM 3.0. Protein Sci. 2022, 31, 209–220. [Google Scholar] [CrossRef]
- Fanani, M.L.; Topham, M.K.; Walsh, J.P.; Epand, R.M. Lipid Modulation of the Activity of Diacylglycerol Kinase α- and ζ-Isoforms: Activation by Phosphatidylethanolamine and Cholesterol. Biochemistry 2004, 43, 14767–14777. [Google Scholar] [CrossRef]
- Abe, T.; Lu, X.; Jiang, Y.; Boccone, C.E.; Qian, S.; Vattem, K.M.; Wek, R.C.; Walsh, J.P. Site-Directed Mutagenesis of the Active Site of Diacylglycerol Kinase Alpha: Calcium and Phosphatidylserine Stimulate Enzyme Activity via Distinct Mechanisms. Biochem. J. 2003, 375, 673–680. [Google Scholar] [CrossRef] [Green Version]
- Sakane, F.; Yamada, K.; Kanoh, H. Different Effects of Sphingosine, R59022 and Anionic Amphiphiles on Two Diacylglycerol Kinase Isozymes Purified from Porcine Thymus Cytosol. FEBS Lett. 1989, 255, 409–413. [Google Scholar] [CrossRef]
- Keiko, Y.; Fumio, S. The Different Effects of Sphingosine on Diacylglycerol Kinase Isozymes in Jurkat Cells, a Human T-Cell Line. Biochim. Biophys. Acta BBA Lipids Lipid Metab. 1993, 1169, 211–216. [Google Scholar] [CrossRef]
- Keiko, Y.; Fumio, S.; Shin-ichi, I.; Haruo, T. Sphingosine Activates Cellular Diacylglycerol Kinase in Intact Jurkat Cells, a Human T-Cell Line. Biochim. Biophys. Acta BBA Lipids Lipid Metab. 1993, 1169, 217–224. [Google Scholar] [CrossRef]
- Epand, R.M.; Kam, A.; Bridgelal, N.; Saiga, A.; Topham, M.K. The α Isoform of Diacylglycerol Kinase Exhibits Arachidonoyl Specificity with Alkylacylglycerol. Biochemistry 2004, 43, 14778–14783. [Google Scholar] [CrossRef]
- Komenoi, S.; Takemura, F.; Sakai, H.; Sakane, F. Diacylglycerol Kinase H1 Is a High Affinity Isozyme for Diacylglycerol. FEBS Lett. 2015, 589, 1272–1277. [Google Scholar] [CrossRef] [Green Version]
- Bozelli, J.; Jennings, W.; Black, S.; Hou, Y.; Lameire, D.; Chatha, P.; Kimura, T.; Berno, B.; Khondker, A.; Rheinstädter, M.; et al. Membrane Curvature Allosterically Regulates the Phosphatidylinositol Cycle, Controlling Its Rate and Acyl-Chain Composition of Its Lipid Intermediates. J. Biol. Chem. 2018, 293, 17780–17791. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Servio, P.; Rey, A.D. Rate of Entropy Production in Evolving Interfaces and Membranes under Astigmatic Kinematics: Shape Evolution in Geometric-Dissipation Landscapes. Entropy 2020, 22, 909. [Google Scholar] [CrossRef]
- Kooijman, E.E.; Burger, K.N.J. Biophysics and Function of Phosphatidic Acid: A Molecular Perspective. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2009, 1791, 881–888. [Google Scholar] [CrossRef]
- Carrasco, S.; Mérida, I. Diacylglycerol, When Simplicity Becomes Complex. Trends Biochem. Sci. 2007, 32, 27–36. [Google Scholar] [CrossRef]
- Mérida, I.; Ávila-Flores, A.; Merino, E. Diacylglycerol Kinases: At the Hub of Cell Signalling. Biochem. J. 2008, 409, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Almena, M.; Mérida, I. Shaping up the Membrane: Diacylglycerol Coordinates Spatial Orientation of Signaling. Trends Biochem. Sci. 2011, 36, 593–603. [Google Scholar] [CrossRef]
- Sprong, H.; van der Sluijs, P.; van Meer, G. How Proteins Move Lipids and Lipids Move Proteins. Nat. Rev. Mol. Cell Biol. 2001, 2, 504–513. [Google Scholar] [CrossRef]
- Zimmerberg, J.; Kozlov, M.M. How Proteins Produce Cellular Membrane Curvature. Nat. Rev. Mol. Cell Biol. 2005, 7, 9–19. [Google Scholar] [CrossRef]
- Xie, S.; Naslavsky, N.; Caplan, S. Diacylglycerol Kinases in Membrane Trafficking. Cell. Logist. 2015, 5, e1078431. [Google Scholar] [CrossRef] [Green Version]
- Toker, A. The Biology and Biochemistry of Diacylglycerol Signalling. Meeting on Molecular Advances in Diacylglycerol Signalling. EMBO Rep. 2005, 6, 310–314. [Google Scholar] [CrossRef]
- Schuhmacher, M.; Grasskamp, A.; Barahtjan, P.; Wagner, N.; Lombardot, B.; Schuhmacher, J.; Sala, P.; Lohmann, A.; Henry, I.; Shevchenko, A.; et al. Live-Cell Lipid Biochemistry Reveals a Role of Diacylglycerol Side-Chain Composition for Cellular Lipid Dynamics and Protein Affinities. Proc. Natl. Acad. Sci. USA 2020, 117, 7729–7738. [Google Scholar] [CrossRef] [Green Version]
- Mérida, I.; Andrada, E.; Gharbi, S.; Ávila-Flores, A. Redundant and Specialized Roles for Diacylglycerol Kinases α and ζ in the Control of T Cell Functions. Sci. Signal. 2015, 8, re6. [Google Scholar] [CrossRef]
- Huppa, J.B.; Davis, M.M. The Interdisciplinary Science of T-Cell Recognition. Adv. Immunol. 2013, 119, 1–50. [Google Scholar] [CrossRef]
- Chauveau, A.; le Floc’h, A.; Bantilan, N.S.; Koretzky, G.A.; Huse, M. Diacylglycerol Kinase α Establishes T Cell Polarity by Shaping Diacylglycerol Accumulation at the Immunological Synapse. Sci. Signal. 2014, 7, ra82. [Google Scholar] [CrossRef] [Green Version]
- Macián, F.; García-Cózar, F.; Im, S.H.; Horton, H.F.; Byrne, M.C.; Rao, A. Transcriptional Mechanisms Underlying Lymphocyte Tolerance. Cell 2002, 109, 719–731. [Google Scholar] [CrossRef] [Green Version]
- Quann, E.J.; Merino, E.; Furuta, T.; Huse, M. Localized Diacylglycerol Drives the Polarization of the Microtubule-Organizing Center in T Cells. Nat. Immunol. 2009, 10, 627–635. [Google Scholar] [CrossRef]
- Andrada, E.; Almena, M.; de Guinoa, J.S.; Merino-Cortes, S.V.; Liébana, R.; Arcos, R.; Carrasco, S.; Carrasco, Y.R.; Merida, I. Diacylglycerol Kinase ζ Limits the Polarized Recruitment of Diacylglycerol-Enriched Organelles to the Immune Synapse in T Cells. Sci. Signal. 2016, 9, ra127. [Google Scholar] [CrossRef] [PubMed]
- Andrada, E.; Liébana, R.; Merida, I. Diacylglycerol Kinase ζ Limits Cytokine-Dependent Expansion of CD8+ T Cells with Broad Antitumor Capacity. EBioMedicine 2017, 19, 39–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Mancha, N.; Mérida, I. Interplay Between SNX27 and DAG Metabolism in the Control of Trafficking and Signaling at the IS. Int. J. Mol. Sci. 2020, 21, 4254. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.S.; Hu, Z.; Zhong, X.P. Diacylglycerol Kinases in T Cell Tolerance and Effector Function. Front. Cell Dev. Biol. 2016, 4, 130. [Google Scholar] [CrossRef] [Green Version]
- Kefas, B.; Floyd, D.H.; Comeau, L.; Frisbee, A.; Dominguez, C.; DiPierro, C.G.; Guessous, F.; Abounader, R.; Purow, B. A MiR-297/Hypoxia/DGK-α Axis Regulating Glioblastoma Survival. Neuro Oncol. 2013, 15, 1652–1663. [Google Scholar] [CrossRef]
- Kai, M.; Yasuda, S.; Imai, S.-I.; Toyota, M.; Kanoh, H.; Sakane, F. Diacylglycerol kinase α enhances protein kinase Cζ-dependent phosphorylation at Ser311 of p65/RelA subunit of nuclear factor-κB. FEBS Lett. 2009, 583, 3265–3268. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Pan, C.; Boese, A.C.; Kang, J.; Umano, A.D.; Magliocca, K.R.; Yang, W.; Zhang, Y.; Lonial, S.; Jin, L.; et al. DGKA Provides Platinum Resistance in Ovarian Cancer Through Activation of c-JUN–WEE1 Signaling. Clin. Cancer Res. 2020, 26, 3843–3855. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, W.; Wang, Y.; Zhao, D.; Wu, M.; Fan, J.; Li, J.; Gong, Y.; Dan, N.; Yang, D.; et al. The Diacylglycerol Kinase α (DGKα)/Akt/NF-ΚB Feedforward Loop Promotes Esophageal Squamous Cell Carcinoma (ESCC) Progression via FAK-Dependent and FAK-Independent Manner. Oncogene 2018, 38, 2533–2550. [Google Scholar] [CrossRef]
- Rainero, E.; Caswell, P.T.; Muller, P.A.J.; Grindlay, J.; Mccaffrey, M.W.; Zhang, Q.; Wakelam, M.J.O.; Vousden, K.H.; Graziani, A.; Norman, J.C. Diacylglycerol Kinase α Controls RCP-Dependent Integrin Trafficking to Promote Invasive Migration. J. Cell Biol. 2012, 196, 277–295. [Google Scholar] [CrossRef]
- Rainero, E.; Cianflone, C.; Porporato, P.E.; Chianale, F.; Malacarne, V.; Bettio, V.; Ruffo, E.; Ferrara, M.; Benecchia, F.; Capello, D.; et al. The Diacylglycerol Kinase α/Atypical PKC/Β1 Integrin Pathway in SDF-1α Mammary Carcinoma Invasiveness. PLoS ONE 2014, 9, e97144. [Google Scholar] [CrossRef]
- Grakoui, A.; Bromley, S.K.; Sumen, C.; Davis, M.M.; Shaw, A.S.; Allen, P.M.; Dustin, M.L. The Immunological Synapse: A Molecular Machine Controlling T Cell Activation. Science 1999, 285, 221–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, S.Y.; Groves, J.T.; Chakraborty, A.K. Synaptic Pattern Formation during Cellular Recognition. Proc. Natl. Acad. Sci. USA 2001, 98, 6548–6553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campi, G.; Varma, R.; Dustin, M.L. Actin and Agonist MHC-Peptide Complex-Dependent T Cell Receptor Microclusters as Scaffolds for Signaling. J. Exp. Med. 2005, 202, 1031–1036. [Google Scholar] [CrossRef] [PubMed]
- Yokosuka, T.; Sakata-Sogawa, K.; Kobayashi, W.; Hiroshima, M.; Hashimoto-Tane, A.; Tokunaga, M.; Dustin, M.L.; Saito, T. Newly Generated T Cell Receptor Microclusters Initiate and Sustain T Cell Activation by Recruitment of Zap70 and SLP-76. Nat. Immunol. 2005, 6, 1253–1262. [Google Scholar] [CrossRef]
- Bunnell, S.C.; Hong, D.I.; Kardon, J.R.; Yamazaki, T.; McGlade, C.J.; Barr, V.A.; Samelson, L.E. T Cell Receptor Ligation Induces the Formation of Dynamically Regulated Signaling Assemblies. J. Cell Biol. 2002, 158, 1263–1275. [Google Scholar] [CrossRef] [Green Version]
- Monks, C.R.F.; Freiberg, B.A.; Kupfer, H.; Sciaky, N.; Kupfer, A. Three-Dimensional Segregation of Supramolecular Activation Clusters in T Cells. Nature 1998, 395, 82–86. [Google Scholar] [CrossRef]
- Dustin, M.; Chakraborty, A.K.; Shaw, A.S. Understanding the Structure and Function of the Immunological Synapse. Cold Spring Harb. Perspect. Biol. 2010, 2, a002311. [Google Scholar] [CrossRef]
- Baldanzi, G.; Bettio, V.; Malacarne, V.; Graziani, A. Diacylglycerol Kinases: Shaping Diacylglycerol and Phosphatidic Acid Gradients to Control Cell Polarity. Front. Cell Dev. Biol. 2016, 4, 140. [Google Scholar] [CrossRef] [Green Version]
- Al-Aghbar, M.A.; Jainarayanan, A.K.; Dustin, M.L.; Roffler, S.R. The Interplay between Membrane Topology and Mechanical Forces in Regulating T Cell Receptor Activity. Commun. Biol. 2022, 5, 1–16. [Google Scholar] [CrossRef]
- Onnis, A.; Baldari, C.T. Orchestration of Immunological Synapse Assembly by Vesicular Trafficking. Front. Cell Dev. Biol. 2019, 7, 110. [Google Scholar] [CrossRef]
- Onnis, A.; Finetti, F.; Baldari, C.T. Vesicular Trafficking to the Immune Synapse: How to Assemble Receptor-Tailored Pathways from a Basic Building Set. Front. Immunol. 2016, 7, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niedergang, F.; Dautry-Varsat, A.; Alcover, A. Peptide Antigen or Superantigen-Induced down-Regulation of TCRs Involves Both Stimulated and Unstimulated Receptors. J. Immunol. 1997, 159, 1703–1710. [Google Scholar] [PubMed]
- Monjas, A.; Alcover, A.; Alarcón, B. Engaged and Bystander T Cell Receptors Are Down-Modulated by Different Endocytotic Pathways. J. Biol. Chem. 2004, 279, 55376–55384. [Google Scholar] [CrossRef] [Green Version]
- Bonefeld, C.M.; Rasmussen, A.B.; Lauritsen, J.P.H.; von Essen, M.; Ødum, N.; Andersen, P.S.; Geisler, C. TCR Comodulation of Nonengaged TCR Takes Place by a Protein Kinase C and CD3 Gamma Di-Leucine-Based Motif-Dependent Mechanism. J. Immunol. 2003, 171, 3003–3009. [Google Scholar] [CrossRef] [Green Version]
- Das, V.; Nal, B.; Dujeancourt, A.; Thoulouze, M.I.; Galli, T.; Roux, P.; Dautry-Varsat, A.; Alcover, A. Activation-Induced Polarized Recycling Targets T Cell Antigen Receptors to the Immunological Synapse: Involvement of SNARE Complexes. Immunity 2004, 20, 577–588. [Google Scholar] [CrossRef] [Green Version]
- Mittelbrunn, M.; Gutiérrez-Vázquez, C.; Villarroya-Beltri, C.; González, S.; Sánchez-Cabo, F.; González, M.Á.; Bernad, A.; Sánchez-Madrid, F. Unidirectional Transfer of MicroRNA-Loaded Exosomes from T Cells to Antigen-Presenting Cells. Nat. Commun. 2011, 2, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huse, M.; Quann, E.J.; Davis, M.M. Shouts, Whispers and the Kiss of Death: Directional Secretion in T Cells. Nat. Immunol. 2008, 9, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Huse, M.; Lillemeier, B.F.; Kuhns, M.S.; Chen, D.S.; Davis, M.M. T Cells Use Two Directionally Distinct Pathways for Cytokine Secretion. Nat. Immunol. 2006, 7, 247–255. [Google Scholar] [CrossRef]
- Reichert, P.; Reinhardt, R.L.; Ingulli, E.; Jenkins, M.K. Cutting Edge: In Vivo Identification of TCR Redistribution and Polarized IL-2 Production by Naive CD4 T Cells. J. Immunol. 2001, 166, 4278–4281. [Google Scholar] [CrossRef] [Green Version]
- Kupfer, A.; Mosmann, T.R.; Kupfer, H. Polarized Expression of Cytokines in Cell Conjugates of Helper T Cells and Splenic B Cells. Proc. Natl. Acad. Sci. USA 1991, 88, 775–779. [Google Scholar] [CrossRef]
- Strasser, A. Death of a T Cell. Nature 1995, 373, 385–386. [Google Scholar] [CrossRef]
- Martínez-Lorenzo, M.J.; Anel, A.; Alava, M.A.; Piñeiro, A.; Naval, J.; Lasierra, P.; Larrad, L. The Human Melanoma Cell Line MelJuSo Secretes Bioactive FasL and APO2L/TRAIL on the Surface of Microvesicles. Possible Contribution to Tumor Counterattack. Exp. Cell Res. 2004, 295, 315–329. [Google Scholar] [CrossRef] [PubMed]
- Stoorvogel, W.; Kleijmeer, M.J.; Geuze, H.J.; Raposo, G. The Biogenesis and Functions of Exosomes. Traffic 2002, 3, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Prinz, P.U.; Mendler, A.N.; Masouris, I.; Durner, L.; Oberneder, R.; Noessner, E. High DGK-α and Disabled MAPK Pathways Cause Dysfunction of Human Tumor-Infiltrating CD8+ T Cells That Is Reversible by Pharmacologic Intervention. J. Immunol. 2012, 188, 5990–6000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohrbough, J.; Broadie, K. Lipid Regulation of the Synaptic Vesicle Cycle. Nat. Rev. Neurosci. 2005, 6, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Friedl, P.; Wolf, K. Tumour-Cell Invasion and Migration: Diversity and Escape Mechanisms. Nat. Rev. Cancer 2003, 3, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Lauffenburger, D.A.; Horwitz, A.F. Cell Migration: A Physically Integrated Molecular Process. Cell 1996, 84, 359–369. [Google Scholar] [CrossRef] [Green Version]
- Chianale, F.; Rainero, E.; Cianflone, C.; Bettio, V.; Pighini, A.; Porporato, P.E.; Filigheddu, N.; Serini, G.; Sinigaglia, F.; Baldanzi, G.; et al. Diacylglycerol Kinase Alpha Mediates HGF-Induced Rac Activation and Membrane Ruffling by Regulating Atypical PKC and RhoGDI. Proc. Natl. Acad. Sci. USA 2010, 107, 4182–4187. [Google Scholar] [CrossRef] [Green Version]
- Chianale, F.; Cutrupi, S.; Rainero, E.; Baldanzi, G.; Porporato, P.E.; Traini, S.; Filigheddu, N.; Gnocchi, V.F.; Santoro, M.M.; Parolini, O.; et al. Diacylglycerol Kinase-Alpha Mediates Hepatocyte Growth Factor-Induced Epithelial Cell Scatter by Regulating Rac Activation and Membrane Ruffling. Mol. Biol. Cell 2007, 18, 4859–4871. [Google Scholar] [CrossRef] [Green Version]
- Novo, D.; Heath, N.; Mitchell, L.; Caligiuri, G.; MacFarlane, A.; Reijmer, D.; Charlton, L.; Knight, J.; Calka, M.; McGhee, E.; et al. Mutant P53s Generate Pro-Invasive Niches by Influencing Exosome Podocalyxin Levels. Nat. Commun. 2018, 9, 1–17. [Google Scholar] [CrossRef]
- Tyson, R.A.; Zatulovskiy, E.; Kay, R.R.; Bretschneider, T. How blebs and pseudopods cooperate during chemotaxis. Proc. Natl. Acad. Sci. USA 2014, 111, 11703–11708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piccolo, E.; Innominato, P.F.; Mariggio, M.A.; Maffucci, T.; Iacobelli, S.; Falasca, M. The Mechanism Involved in the Regulation of Phospholipase Cgamma1 Activity in Cell Migration. Oncogene 2002, 21, 6520–6529. [Google Scholar] [CrossRef] [Green Version]
- Bozelli, J.C.; Epand, R.M. Specificity of Acyl Chain Composition of Phosphatidylinositols. Proteomics 2019, 19, e1900138. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, T.; Ikeda, M.; Kiso, Y.; Sakuma, M.; Yoshino, K.I.; Sakane, F.; Merida, I.; Saito, N.; Shirai, Y. C-Abl Tyrosine Kinase Regulates Serum-Induced Nuclear Export of Diacylglycerol Kinase α by Phosphorylation at Tyr-218. J. Biol. Chem. 2012, 287, 5507–5517. [Google Scholar] [CrossRef] [Green Version]
- Thaller, D.J.; Tong, D.; Marklew, C.J.; Ader, N.R.; Mannino, P.J.; Borah, S.; King, M.C.; Ciani, B.; Lusk, C.P. Direct Binding of ESCRT Protein Chm7 to Phosphatidic Acid-Rich Membranes at Nuclear Envelope Herniations. J. Cell Biol. 2021, 220, e202004222. [Google Scholar] [CrossRef]
- Bley, C.J.; Nie, S.; Mobbs, G.W.; Petrovic, S.; Gres, A.T.; Liu, X.; Mukherjee, S.; Harvey, S.; Huber, F.M.; Lin, D.H.; et al. Architecture of the Cytoplasmic Face of the Nuclear Pore. Science 2022, 376, eabm9129. [Google Scholar] [CrossRef]
- Petrovic, S.; Samanta, D.; Perriches, T.; Bley, C.J.; Thierbach, K.; Brown, B.; Nie, S.; Mobbs, G.W.; Stevens, T.A.; Liu, X.; et al. Architecture of the linker-scaffold in the nuclear pore. Science 2022, 376, eabm9798. [Google Scholar] [CrossRef]
- Carrasco, S.; Merida, I. Diacylglycerol-dependent Binding Recruits PKCθ and RasGRP1 C1 Domains to Specific Subcellular Localizations in Living T Lymphocytes. Mol. Biol. Cell 2004, 15, 2932–2942. [Google Scholar] [CrossRef]
- Rubio, I.; Grund, S.; Song, S.-P.; Biskup, C.; Bandemer, S.; Fricke, M.; Förster, M.; Graziani, A.; Wittig, U.; Kliche, S. TCR-Induced Activation of Ras Proceeds at the Plasma Membrane and Requires Palmitoylation of N-Ras. J. Immunol. 2010, 185, 3536–3543. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.E.; Goulding, R.E.; Ding, Z.; Partovi, A.; Anthony, K.V.; Beaulieu, N.; Tazmini, G.; Cornell, R.B.; Kay, R.J. Differential Membrane Binding and Diacylglycerol Recognition by C1 Domains of RasGRPs. Biochem. J. 2007, 406, 223–236. [Google Scholar] [CrossRef]
- Augsten, M.; Pusch, R.; Biskup, C.; Rennert, K.; Wittig, U.; Beyer, K.; Blume, A.; Wetzker, R.; Friedrich, K.; Rubio, I. Live-Cell Imaging of Endogenous Ras-GTP Illustrates Predominant Ras Activation at the Plasma Membrane. EMBO Rep. 2006, 7, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, M.A.; Shome, K.; Watkins, S.C.; Romero, G. The Recruitment of Raf-1 to Membranes Is Mediated by Direct Interaction with Phosphatidic Acid and Is Independent of Association with Ras. J. Biol. Chem. 2000, 275, 23911–23918. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Strum, J.C.; Sciorra, V.A.; Daniel, L.; Bell, R.M. Raf-1 Kinase Possesses Distinct Binding Domains for Phosphatidylserine and Phosphatidic Acid: Phosphatidic Acid Regulates The Translocation Of Raf-1 in 12-O-Tetradecanoylphorbol-13-Acetate-Stimulated Madin-Darby Canine Kidney Cells. J. Biol. Chem. 1996, 271, 8472–8480. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Moore, S.; Bell, R.M.; Dush, M. Functional Analysis of a Phosphatidic Acid Binding Domain in Human Raf-1 Kinase: Mutations in the Phosphatidate Binding Domain Lead to Tail and Trunk Abnormalities in Developing Zebrafish Embryos. J. Biol. Chem. 2003, 278, 45690–45696. [Google Scholar] [CrossRef] [Green Version]
- Foster, D.A. Phosphatidic Acid Signaling to MTOR: Signals for the Survival of Human Cancer Cells. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2009, 1791, 949–955. [Google Scholar] [CrossRef] [Green Version]
- Ávila-Flores, A.; Santos, T.; Rincón, E.; Mérida, I. Modulation of the Mammalian Target of Rapamycin Pathway by Diacylglycerol Kinase-Produced Phosphatidic Acid. J. Biol. Chem. 2005, 280, 10091–10099. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.N. MTOR-What Does It Do? Transpl. Proc. 2008, 40, S5–S8. [Google Scholar] [CrossRef]
- Zhang, C.; Wendel, A.A.; Keogh, M.R.; Harris, T.E.; Chen, J.; Coleman, R.A. Glycerolipid Signals Alter MTOR Complex 2 (MTORC2) to Diminish Insulin Signaling. Proc. Natl. Acad. Sci. USA 2012, 109, 1667–1672. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Liu, K.; Sasaki, S.; Kunii, N.; Sakai, H.; Mizuno, H.; Saga, H.; Sakane, F. Evaluations of the Selectivities of the Diacylglycerol Kinase Inhibitors R59022 and R59949 among Diacylglycerol Kinase Isozymes Using a New Non-Radioactive Assay Method. Pharmacology 2013, 92, 99–107. [Google Scholar] [CrossRef]
- Boroda, S.; Niccum, M.; Raje, V.; Purow, B.W.; Harris, T.E. Dual Activities of Ritanserin and R59022 as DGKα Inhibitors and Serotonin Receptor Antagonists. Biochem. Pharm. 2017, 123, 29–39. [Google Scholar] [CrossRef]
- Liu, K.; Kunii, N.; Sakuma, M.; Yamaki, A.; Mizuno, S.; Sato, M.; Sakai, H.; Kado, S.; Kumagai, K.; Kojima, H.; et al. A Novel Diacylglycerol Kinase α -Selective Inhibitor, CU-3, Induces Cancer Cell Apoptosis and Enhances Immune Response. J. Lipid Res. 2016, 57, 368–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, N.; Sugiyama, K.; Shichi, S.; Shirai, Y.; Goto, K.; Sakane, F.; Kitamura, H.; Taketomi, A. Combination Therapy for Hepatocellular Carcinoma with Diacylglycerol Kinase Alpha Inhibition and Anti-Programmed Cell Death-1 Ligand Blockade. Cancer Immunol. Immunother. 2022, 71, 889–903. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bozelli, J.C., Jr.; Epand, R.M. DGKα, Bridging Membrane Shape Changes with Specific Molecular Species of DAG/PA: Implications in Cancer and Immunosurveillance. Cancers 2022, 14, 5259. https://doi.org/10.3390/cancers14215259
Bozelli JC Jr., Epand RM. DGKα, Bridging Membrane Shape Changes with Specific Molecular Species of DAG/PA: Implications in Cancer and Immunosurveillance. Cancers. 2022; 14(21):5259. https://doi.org/10.3390/cancers14215259
Chicago/Turabian StyleBozelli, José Carlos, Jr., and Richard M. Epand. 2022. "DGKα, Bridging Membrane Shape Changes with Specific Molecular Species of DAG/PA: Implications in Cancer and Immunosurveillance" Cancers 14, no. 21: 5259. https://doi.org/10.3390/cancers14215259
APA StyleBozelli, J. C., Jr., & Epand, R. M. (2022). DGKα, Bridging Membrane Shape Changes with Specific Molecular Species of DAG/PA: Implications in Cancer and Immunosurveillance. Cancers, 14(21), 5259. https://doi.org/10.3390/cancers14215259