

Small Tweaks, Major Changes: Post-Translational Modifications That Occur within M2 Macrophages in the Tumor Microenvironment


{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. The Origin of M2 Macrophages
3. Differences between M2 Macrophages and TAMs
4. M2 Macrophages in the Tumor Microenvironment
5. Prognostic Significance of M2 Macrophages in the Tumor Microenvironment
6. Performance of M2 Macrophages in the TME
6.1. Propelling Metastasis
6.2. Immunosuppressive Roles
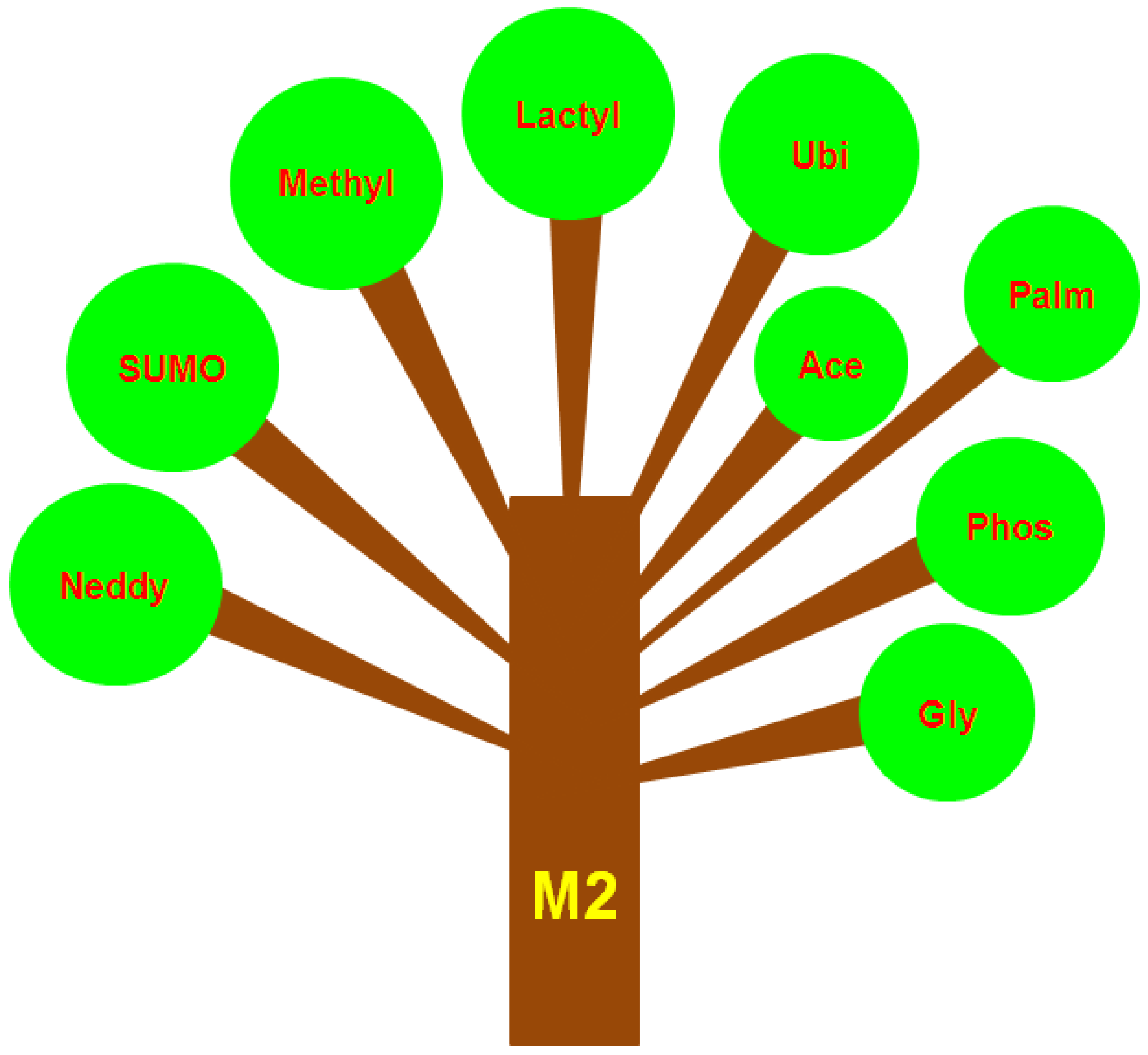
7. Different Types of PTMs That Occur in M2 Macrophages or TAMs
7.1. SUMOylation
7.2. Methylation
7.3. Lactylation
7.4. Ubiquitylation
7.5. Acetylation
7.6. Phosphorylation
7.7. Glycosylation
7.8. Neddylation
7.9. Palmitoylation
8. Technical Considerations when Studying M2 Macrophages in Tumors
9. Inhibitors or Drugs That Target PTMs
10. Artificial Intelligence (AI) and TAMs
11. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Locati, M.; Curtale, G.; Mantovani, A. Diversity, Mechanisms, and Significance of Macrophage Plasticity. Annu. Rev. Pathol. 2020, 15, 123–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, M.T.; Nagaria, B.D.; Britton, W.J.; Saunders, B.M. Macrophages of different tissue origin exhibit distinct inflammatory responses to mycobacterial infection. Immunol. Cell Biol. 2021, 99, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Naito, M.; Hasegawa, G.; Takahashi, K. Development, differentiation, and maturation of Kupffer cells. Microsc. Res. Tech. 1997, 39, 350–364. [Google Scholar] [CrossRef]
- Evren, E.; Ringqvist, E.; Willinger, T. Origin and ontogeny of lung macrophages: From mice to humans. Immunology 2020, 160, 126–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prinz, M.; Jung, S.; Priller, J. Microglia Biology: One Century of Evolving Concepts. Cell 2019, 179, 292–311. [Google Scholar] [CrossRef]
- Deane, H.W. Some Electron Microscopic Observations on the Lamina Propria of the Gut, with Comments on the Close Association of Macrophages, Plasma Cells, and Eosinophils. Anat. Rec. 1964, 149, 453–473. [Google Scholar] [CrossRef] [PubMed]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Yunna, C.; Mengru, H.; Lei, W.; Weidong, C. Macrophage M1/M2 polarization. Eur. J. Pharmacol. 2020, 877, 173090. [Google Scholar] [CrossRef]
- Zhang, Y.; Zuo, C.; Liu, L.; Hu, Y.; Yang, B.; Qiu, S.; Li, Y.; Cao, D.; Ju, Z.; Ge, J.; et al. Single-cell RNA-sequencing atlas reveals an MDK-dependent immunosuppressive environment in ErbB pathway-mutated gallbladder cancer. J. Hepatol. 2021, 75, 1128–1141. [Google Scholar] [CrossRef]
- Kim, N.; Kim, H.K.; Lee, K.; Hong, Y.; Cho, J.H.; Choi, J.W.; Lee, J.I.; Suh, Y.L.; Ku, B.M.; Eum, H.H.; et al. Single-cell RNA sequencing demonstrates the molecular and cellular reprogramming of metastatic lung adenocarcinoma. Nat. Commun. 2020, 11, 2285. [Google Scholar] [CrossRef]
- Qian, B.Z.; Pollard, J.W. Macrophage diversity enhances tumor progression and metastasis. Cell 2010, 141, 39–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laviron, M.; Boissonnas, A. Ontogeny of Tumor-Associated Macrophages. Front. Immunol. 2019, 10, 1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Herscowitz, H.B.; Pennline, K.J.; Conrad, R.E.; Ullrich, S.E.; Gerber, H.R. Macrophage-lymphocyte interactions mediated by soluble factors. Ann. N. Y. Acad. Sci. 1979, 332, 464–481. [Google Scholar] [CrossRef] [PubMed]
- Evans, R.L.; Breard, J.M.; Lazarus, H.; Schlossman, S.F.; Chess, L. Detection, isolation, and functional characterization of two human T-cell subclasses bearing unique differentiation antigens. J. Exp. Med. 1977, 145, 221–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinherz, E.L.; Schlossman, S.F. Con A-inducible suppression of MLC: Evidence for mediation by the TH2+ T cell subset in man. J. Immunol. 1979, 122, 1335–1341. [Google Scholar]
- Stein, M.; Keshav, S.; Harris, N.; Gordon, S. Interleukin 4 potently enhances murine macrophage mannose receptor activity: A marker of alternative immunologic macrophage activation. J. Exp. Med. 1992, 176, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Mills, C.D.; Kincaid, K.; Alt, J.M.; Heilman, M.J.; Hill, A.M. M-1/M-2 macrophages and the Th1/Th2 paradigm. J. Immunol. 2000, 164, 6166–6173. [Google Scholar] [CrossRef] [Green Version]
- Munder, M.; Eichmann, K.; Moran, J.M.; Centeno, F.; Soler, G.; Modolell, M. Th1/Th2-regulated expression of arginase isoforms in murine macrophages and dendritic cells. J. Immunol. 1999, 163, 3771–3777. [Google Scholar]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage polarization: Tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002, 23, 549–555. [Google Scholar] [CrossRef]
- Siveen, K.S.; Kuttan, G. Role of macrophages in tumour progression. Immunol. Lett. 2009, 123, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.X.; Joshi, S. “Re-Educating” Tumor Associated Macrophages as a Novel Immunotherapy Strategy for Neuroblastoma. Front. Immunol. 2020, 11, 1947. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.; Klink, M. The Role of Tumor-Associated Macrophages in the Progression and Chemoresistance of Ovarian Cancer. Cells 2020, 9, 1299. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Makino, T.; Sato, E.; Noma, T.; Urakawa, S.; Takeoka, T.; Yamashita, K.; Saito, T.; Tanaka, K.; Takahashi, T.; et al. Tumor-infiltrating M2 macrophage in pretreatment biopsy sample predicts response to chemotherapy and survival in esophageal cancer. Cancer Sci. 2020, 111, 1103–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarosz-Biej, M.; Smolarczyk, R.; Cichon, T.; Kulach, N. Tumor Microenvironment as A “Game Changer” in Cancer Radiotherapy. Int. J. Mol. Sci. 2019, 20, 3212. [Google Scholar] [CrossRef] [Green Version]
- Leblond, M.M.; Peres, E.A.; Helaine, C.; Gerault, A.N.; Moulin, D.; Anfray, C.; Divoux, D.; Petit, E.; Bernaudin, M.; Valable, S. M2 macrophages are more resistant than M1 macrophages following radiation therapy in the context of glioblastoma. Oncotarget 2017, 8, 72597–72612. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, A.; Bottazzi, B.; Colotta, F.; Sozzani, S.; Ruco, L. The origin and function of tumor-associated macrophages. Immunol. Today 1992, 13, 265–270. [Google Scholar] [CrossRef]
- Oshi, M.; Tokumaru, Y.; Asaoka, M.; Yan, L.; Satyananda, V.; Matsuyama, R.; Matsuhashi, N.; Futamura, M.; Ishikawa, T.; Yoshida, K.; et al. M1 Macrophage and M1/M2 ratio defined by transcriptomic signatures resemble only part of their conventional clinical characteristics in breast cancer. Sci. Rep. 2020, 10, 16554. [Google Scholar] [CrossRef]
- You, Y.; Tian, Z.; Du, Z.; Wu, K.; Xu, G.; Dai, M.; Wang, Y.; Xiao, M. M1-like tumor-associated macrophages cascade a mesenchymal/stem-like phenotype of oral squamous cell carcinoma via the IL6/Stat3/THBS1 feedback loop. J. Exp. Clin. Cancer Res. 2022, 41, 10. [Google Scholar] [CrossRef]
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef]
- Galdiero, M.R.; Marone, G.; Mantovani, A. Cancer Inflammation and Cytokines. Cold Spring Harb. Perspect. Biol. 2018, 10, a028662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szebeni, G.J.; Vizler, C.; Kitajka, K.; Puskas, L.G. Inflammation and Cancer: Extra- and Intracellular Determinants of Tumor-Associated Macrophages as Tumor Promoters. Mediators Inflamm. 2017, 2017, 9294018. [Google Scholar] [CrossRef] [PubMed]
- Solinas, G.; Germano, G.; Mantovani, A.; Allavena, P. Tumor-associated macrophages (TAM) as major players of the cancer-related inflammation. J. Leukoc. Biol. 2009, 86, 1065–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Candido, J.; Hagemann, T. Cancer-related inflammation. J. Clin. Immunol. 2013, 33 (Suppl. 1), S79–S84. [Google Scholar] [CrossRef] [PubMed]
- Bingle, L.; Brown, N.J.; Lewis, C.E. The role of tumour-associated macrophages in tumour progression: Implications for new anticancer therapies. J. Pathol. 2002, 196, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Janeiro, A.; Padilla-Ansala, C.; de Andrea, C.E.; Hardisson, D.; Melero, I. Prognostic value of macrophage polarization markers in epithelial neoplasms and melanoma. A systematic review and meta-analysis. Mod. Pathol. 2020, 33, 1458–1465. [Google Scholar] [CrossRef]
- Allavena, P.; Mantovani, A. Immunology in the clinic review series; focus on cancer: Tumour-associated macrophages: Undisputed stars of the inflammatory tumour microenvironment. Clin. Exp. Immunol. 2012, 167, 195–205. [Google Scholar] [CrossRef]
- Marchesi, F.; Cirillo, M.; Bianchi, A.; Gately, M.; Olimpieri, O.M.; Cerchiara, E.; Renzi, D.; Micera, A.; Balzamino, B.O.; Bonini, S.; et al. High density of CD68+/CD163+ tumour-associated macrophages (M2-TAM) at diagnosis is significantly correlated to unfavorable prognostic factors and to poor clinical outcomes in patients with diffuse large B-cell lymphoma. Hematol. Oncol. 2015, 33, 110–112. [Google Scholar] [CrossRef]
- Antel, K.; Chetty, D.; Oosthuizen, J.; Mohamed, Z.; Van der Vyver, L.; Verburgh, E. CD68-positive tumour associated macrophages, PD-L1 expression, and EBV latent infection in a high HIV-prevalent South African cohort of Hodgkin lymphoma patients. Pathology 2021, 53, 628–634. [Google Scholar] [CrossRef]
- Tiainen, S.; Tumelius, R.; Rilla, K.; Hamalainen, K.; Tammi, M.; Tammi, R.; Kosma, V.M.; Oikari, S.; Auvinen, P. High numbers of macrophages, especially M2-like (CD163-positive), correlate with hyaluronan accumulation and poor outcome in breast cancer. Histopathology 2015, 66, 873–883. [Google Scholar] [CrossRef]
- Cencini, E.; Fabbri, A.; Rigacci, L.; Lazzi, S.; Gini, G.; Cox, M.C.; Mancuso, S.; Abruzzese, E.; Kovalchuk, S.; Goteri, G.; et al. Evaluation of the prognostic role of tumour-associated macrophages in newly diagnosed classical Hodgkin lymphoma and correlation with early FDG-PET assessment. Hematol. Oncol. 2017, 35, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Shu, Q.H.; Ge, Y.S.; Ma, H.X.; Gao, X.Q.; Pan, J.J.; Liu, D.; Xu, G.L.; Ma, J.L.; Jia, W.D. Prognostic value of polarized macrophages in patients with hepatocellular carcinoma after curative resection. J. Cell. Mol. Med. 2016, 20, 1024–1035. [Google Scholar] [CrossRef]
- Behnes, C.L.; Bremmer, F.; Hemmerlein, B.; Strauss, A.; Strobel, P.; Radzun, H.J. Tumor-associated macrophages are involved in tumor progression in papillary renal cell carcinoma. Virchows Arch. 2014, 464, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Deng, L.; Liao, X.; Nie, L.; Qi, F.; Jin, K.; Tu, X.; Zheng, X.; Li, J.; Liu, L.; et al. Tumor-associated macrophages promote bladder tumor growth through PI3K/AKT signal induced by collagen. Cancer Sci. 2019, 110, 2110–2118. [Google Scholar] [CrossRef] [PubMed]
- De Groot, A.E.; Myers, K.V.; Krueger, T.E.G.; Kiemen, A.L.; Nagy, N.H.; Brame, A.; Torres, V.E.; Zhang, Z.; Trabzonlu, L.; Brennen, W.N.; et al. Characterization of tumor-associated macrophages in prostate cancer transgenic mouse models. Prostate 2021, 81, 629–647. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Xiao, Q.; Wang, X.; Yang, C.; Chen, C.; Tian, X.; Wang, S.; Li, H.; Qiu, S.; Shu, J.; et al. Tumor-associated macrophages modulate angiogenesis and tumor growth in a xenograft mouse model of multiple myeloma. Leuk. Res. 2021, 110, 106709. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, B.; Saha, P.; Bose, S.; Shukla, D.; Chatterjee, N.; Kumar, S.; Tripathi, P.P.; Srivastava, A.K. MicroRNAs: As Critical Regulators of Tumor-Associated Macrophages. Int. J. Mol. Sci. 2020, 21, 7117. [Google Scholar] [CrossRef]
- Chen, F.; Chen, J.; Yang, L.; Liu, J.; Zhang, X.; Zhang, Y.; Tu, Q.; Yin, D.; Lin, D.; Wong, P.P.; et al. Extracellular vesicle-packaged HIF-1alpha-stabilizing lncRNA from tumour-associated macrophages regulates aerobic glycolysis of breast cancer cells. Nat. Cell Biol. 2019, 21, 498–510. [Google Scholar] [CrossRef]
- Zhang, Y.; Meng, W.; Yue, P.; Li, X. M2 macrophage-derived extracellular vesicles promote gastric cancer progression via a microRNA-130b-3p/MLL3/GRHL2 signaling cascade. J. Exp. Clin. Cancer Res. 2020, 39, 134. [Google Scholar] [CrossRef]
- Roh-Johnson, M.; Shah, A.N.; Stonick, J.A.; Poudel, K.R.; Kargl, J.; Yang, G.H.; di Martino, J.; Hernandez, R.E.; Gast, C.E.; Zarour, L.R.; et al. Macrophage-Dependent Cytoplasmic Transfer during Melanoma Invasion In Vivo. Dev. Cell 2017, 43, 549–562.E6. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; He, Y.; Luo, N.; Patel, S.J.; Han, Y.; Gao, R.; Modak, M.; Carotta, S.; Haslinger, C.; Kind, D.; et al. Landscape and Dynamics of Single Immune Cells in Hepatocellular Carcinoma. Cell 2019, 179, 829–845.E20. [Google Scholar] [CrossRef] [PubMed]
- Friebel, E.; Kapolou, K.; Unger, S.; Nunez, N.G.; Utz, S.; Rushing, E.J.; Regli, L.; Weller, M.; Greter, M.; Tugues, S.; et al. Single-Cell Mapping of Human Brain Cancer Reveals Tumor-Specific Instruction of Tissue-Invading Leukocytes. Cell 2020, 181, 1626–1642.E20. [Google Scholar] [CrossRef] [PubMed]
- Franklin, R.A.; Liao, W.; Sarkar, A.; Kim, M.V.; Bivona, M.R.; Liu, K.; Pamer, E.G.; Li, M.O. The cellular and molecular origin of tumor-associated macrophages. Science 2014, 344, 921–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, F.O.; Sica, A.; Mantovani, A.; Locati, M. Macrophage activation and polarization. Front. Biosci. 2008, 13, 453–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spary, L.K.; Salimu, J.; Webber, J.P.; Clayton, A.; Mason, M.D.; Tabi, Z. Tumor stroma-derived factors skew monocyte to dendritic cell differentiation toward a suppressive CD14+ PD-L1+ phenotype in prostate cancer. Oncoimmunology 2014, 3, e955331. [Google Scholar] [CrossRef] [Green Version]
- Pradel, L.P.; Franke, A.; Ries, C.H. Effects of IL-10 and Th 2 cytokines on human Mphi phenotype and response to CSF1R inhibitor. J. Leukoc. Biol. 2018, 103, 545–558. [Google Scholar] [CrossRef]
- Kim, R.; Emi, M.; Tanabe, K.; Arihiro, K. Tumor-driven evolution of immunosuppressive networks during malignant progression. Cancer Res. 2006, 66, 5527–5536. [Google Scholar] [CrossRef] [Green Version]
- Schmieder, A.; Michel, J.; Schonhaar, K.; Goerdt, S.; Schledzewski, K. Differentiation and gene expression profile of tumor-associated macrophages. Semin. Cancer Biol. 2012, 22, 289–297. [Google Scholar] [CrossRef]
- Pechkovsky, D.V.; Prasse, A.; Kollert, F.; Engel, K.M.; Dentler, J.; Luttmann, W.; Friedrich, K.; Muller-Quernheim, J.; Zissel, G. Alternatively activated alveolar macrophages in pulmonary fibrosis-mediator production and intracellular signal transduction. Clin. Immunol. 2010, 137, 89–101. [Google Scholar] [CrossRef]
- Wang, K.; Zhou, W.; Cai, Q.; Cheng, J.; Cai, R.; Xing, R. SUMOylation of KLF4 promotes IL-4 induced macrophage M2 polarization. Cell Cycle 2017, 16, 374–381. [Google Scholar] [CrossRef] [Green Version]
- Xiao, M.; Bian, Q.; Lao, Y.; Yi, J.; Sun, X.; Sun, X.; Yang, J. SENP3 loss promotes M2 macrophage polarization and breast cancer progression. Mol. Oncol. 2022, 16, 1026–1044. [Google Scholar] [CrossRef]
- Tikhanovich, I.; Zhao, J.; Olson, J.; Adams, A.; Taylor, R.; Bridges, B.; Marshall, L.; Roberts, B.; Weinman, S.A. Protein arginine methyltransferase 1 modulates innate immune responses through regulation of peroxisome proliferator-activated receptor gamma-dependent macrophage differentiation. J. Biol. Chem. 2017, 292, 6882–6894. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Tang, Z.; Huang, H.; Zhou, G.; Cui, C.; Weng, Y.; Liu, W.; Kim, S.; Lee, S.; Perez-Neut, M.; et al. Metabolic regulation of gene expression by histone lactylation. Nature 2019, 574, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Dichtl, S.; Lindenthal, L.; Zeitler, L.; Behnke, K.; Schlosser, D.; Strobl, B.; Scheller, J.; El Kasmi, K.C.; Murray, P.J. Lactate and IL6 define separable paths of inflammatory metabolic adaptation. Sci. Adv. 2021, 7, eabg3505. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Hartlova, A.; Gierlinski, M.; Prescott, A.; Castellvi, J.; Losa, J.H.; Petersen, S.K.; Wenzel, U.A.; Dill, B.D.; Emmerich, C.H.; et al. Triggering MSR1 promotes JNK-mediated inflammation in IL-4-activated macrophages. EMBO J. 2019, 38, e100299. [Google Scholar] [CrossRef]
- Kolliniati, O.; Ieronymaki, E.; Vergadi, E.; Tsatsanis, C. Metabolic Regulation of Macrophage Activation. J. Innate Immun. 2022, 14, 51–67. [Google Scholar] [CrossRef]
- Covarrubias, A.J.; Aksoylar, H.I.; Yu, J.; Snyder, N.W.; Worth, A.J.; Iyer, S.S.; Wang, J.; Ben-Sahra, I.; Byles, V.; Polynne-Stapornkul, T.; et al. Akt-mTORC1 signaling regulates Acly to integrate metabolic input to control of macrophage activation. Elife 2016, 5, e11612. [Google Scholar] [CrossRef]
- Namgaladze, D.; Zukunft, S.; Schnutgen, F.; Kurrle, N.; Fleming, I.; Fuhrmann, D.; Brune, B. Polarization of Human Macrophages by Interleukin-4 Does Not Require ATP-Citrate Lyase. Front. Immunol. 2018, 9, 2858. [Google Scholar] [CrossRef] [Green Version]
- Su, P.; Wang, Q.; Bi, E.; Ma, X.; Liu, L.; Yang, M.; Qian, J.; Yi, Q. Enhanced Lipid Accumulation and Metabolism Are Required for the Differentiation and Activation of Tumor-Associated Macrophages. Cancer Res. 2020, 80, 1438–1450. [Google Scholar] [CrossRef]
- Tariq, M.; Zhang, J.Q.; Liang, G.K.; He, Q.J.; Ding, L.; Yang, B. Gefitinib inhibits M2-like polarization of tumor-associated macrophages in Lewis lung cancer by targeting the STAT6 signaling pathway. Acta Pharmacol. Sin. 2017, 38, 1501–1511. [Google Scholar] [CrossRef] [Green Version]
- Chatham, J.C.; Zhang, J.; Wende, A.R. Role of O-Linked N-Acetylglucosamine Protein Modification in Cellular (Patho)Physiology. Physiol. Rev. 2021, 101, 427–493. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues Mantuano, N.; Stanczak, M.A.; Oliveira, I.A.; Kirchhammer, N.; Filardy, A.A.; Monaco, G.; Santos, R.C.; Fonseca, A.C.; Fontes, M.; Bastos, C.S., Jr.; et al. Hyperglycemia Enhances Cancer Immune Evasion by Inducing Alternative Macrophage Polarization through Increased O-GlcNAcylation. Cancer Immunol. Res. 2020, 8, 1262–1272. [Google Scholar] [CrossRef] [PubMed]
- Hinshaw, D.C.; Hanna, A.; Lama-Sherpa, T.; Metge, B.; Kammerud, S.C.; Benavides, G.A.; Kumar, A.; Alsheikh, H.A.; Mota, M.; Chen, D.; et al. Hedgehog signaling regulates metabolism and polarization of mammary tumor-associated macrophages. Cancer Res. 2021, 81, 5425–5437. [Google Scholar] [CrossRef] [PubMed]
- Mantuano, N.R.; Oliveira-Nunes, M.C.; Alisson-Silva, F.; Dias, W.B.; Todeschini, A.R. Emerging role of glycosylation in the polarization of tumor-associated macrophages. Pharmacol. Res. 2019, 146, 104285. [Google Scholar] [CrossRef] [PubMed]
- Enchev, R.I.; Schulman, B.A.; Peter, M. Protein neddylation: Beyond cullin-RING ligases. Nat. Rev. Mol. Cell Biol. 2015, 16, 30–44. [Google Scholar] [CrossRef]
- Zhou, L.; Jiang, Y.; Luo, Q.; Li, L.; Jia, L. Neddylation: A novel modulator of the tumor microenvironment. Mol. Cancer 2019, 18, 77. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Jia, L. Neddylation Pathway as a Novel Anti-cancer Target: Mechanistic Investigation and Therapeutic Implication. Anticancer Agents Med. Chem. 2015, 15, 1127–1133. [Google Scholar] [CrossRef]
- Asare, Y.; Ommer, M.; Azombo, F.A.; Alampour-Rajabi, S.; Sternkopf, M.; Sanati, M.; Gijbels, M.J.; Schmitz, C.; Sinitski, D.; Tilstam, P.V.; et al. Inhibition of atherogenesis by the COP9 signalosome subunit 5 in vivo. Proc. Natl. Acad. Sci. USA 2017, 114, E2766–E2775. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Chen, Y.; Feng, W.; Hua, R.; Zhang, J.; Huo, Y.; Jiang, H.; Yin, B.; Yang, X. Neddylation pathway alleviates chronic pancreatitis by reducing HIF1alpha-CCL5-dependent macrophage infiltration. Cell Death Dis. 2021, 12, 273. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, L.; Li, Y.; Liu, G.; Hoffman, R.M.; Jia, L. Neddylation Regulates Macrophages and Implications for Cancer Therapy. Front. Cell Dev. Biol. 2021, 9, 681186. [Google Scholar] [CrossRef]
- Ross, E.M. Protein modification. Palmitoylation in G-protein signaling pathways. Curr. Biol. 1995, 5, 107–109. [Google Scholar] [CrossRef] [Green Version]
- Yeste-Velasco, M.; Linder, M.E.; Lu, Y.J. Protein S-palmitoylation and cancer. Biochim. Biophys. Acta 2015, 1856, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Ko, P.J.; Dixon, S.J. Protein palmitoylation and cancer. EMBO Rep. 2018, 19, e46666. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Zhi, X.; Wang, X.; Meng, D. Protein palmitoylation and its pathophysiological relevance. J. Cell. Physiol. 2021, 236, 3220–3233. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Ojha, R.; Noguera-Ortega, E.; Rebecca, V.W.; Attanasio, J.; Liu, S.; Piao, S.; Lee, J.J.; Nicastri, M.C.; Harper, S.L.; et al. PPT1 inhibition enhances the antitumor activity of anti-PD-1 antibody in melanoma. JCI Insight 2020, 5, e133225. [Google Scholar] [CrossRef] [PubMed]
- Tasmia, S.A.; Kibria, M.K.; Islam, M.A.; Khatun, M.S.; Haque Mollah, M.N. A comprehensive comparative review of protein sequence based computational prediction models of lysine succinylation sites. Curr. Protein Pept. Sci. 2022, 23, 1389–2037. [Google Scholar] [CrossRef]
- Zhao, Y.W.; Zhang, S.; Ding, H. Recent Development of Machine Learning Methods in Sumoylation Sites Prediction. Curr. Med. Chem. 2022, 29, 894–907. [Google Scholar] [CrossRef]
- Khalili, E.; Ramazi, S.; Ghanati, F.; Kouchaki, S. Predicting protein phosphorylation sites in soybean using interpretable deep tabular learning network. Brief. Bioinform. 2022, 23, bbac015. [Google Scholar] [CrossRef]
- Zhang, H.; He, J.; Hu, G.; Zhu, F.; Jiang, H.; Gao, J.; Zhou, H.; Lin, H.; Wang, Y.; Chen, K.; et al. Dynamics of Post-Translational Modification Inspires Drug Design in the Kinase Family. J. Med. Chem. 2021, 64, 15111–15125. [Google Scholar] [CrossRef]
- Meng, F.; Liang, Z.; Zhao, K.; Luo, C. Drug design targeting active posttranslational modification protein isoforms. Med. Res. Rev. 2021, 41, 1701–1750. [Google Scholar] [CrossRef]
- Lisi, S.; Trovato, M.; Vitaloni, O.; Fantini, M.; Chirichella, M.; Tognini, P.; Cornuti, S.; Costa, M.; Groth, M.; Cattaneo, A. Acetylation-Specific Interference by Anti-Histone H3K9ac Intrabody Results in Precise Modulation of Gene Expression. Int. J. Mol. Sci. 2022, 23, 8892. [Google Scholar] [CrossRef] [PubMed]
- Sumam de Oliveira, D.; Kronenberger, T.; Palmisano, G.; Wrenger, C.; de Souza, E.E. Targeting SUMOylation in Plasmodium as a Potential Target for Malaria Therapy. Front. Cell. Infect. Microbiol. 2021, 11, 685866. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, Y.; Takubo, K.; Kawada, K.; Ohue, M.; Higashi, M.; Ishiguro, T.; Hatano, S.; Toyomasu, Y.; Matsuyama, T.; Mochiki, E.; et al. Endocytoscopic Observation of Esophageal Lesions: Our Own Experience and a Review of the Literature. Diagnostics 2022, 12, 2222. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, C.; Liu, X.; Sun, W.; Li, L.; Gao, N.; Zhang, Y.; Yang, X.; Zhang, J.; Wang, H.; et al. Artificial intelligence-assisted system for precision diagnosis of PD-L1 expression in non-small cell lung cancer. Mod. Pathol. 2022, 35, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Li, X.; Mao, Q.; Wang, Y.; Li, H.; Wang, C.; Shen, Y.; Guo, E.; He, Q.; Tian, J.; et al. An artificial intelligence method to assess the tumor microenvironment with treatment outcomes for gastric cancer patients after gastrectomy. J. Transl. Med. 2022, 20, 100. [Google Scholar] [CrossRef]
- Kawakami, E.; Tabata, J.; Yanaihara, N.; Ishikawa, T.; Koseki, K.; Iida, Y.; Saito, M.; Komazaki, H.; Shapiro, J.S.; Goto, C.; et al. Application of Artificial Intelligence for Preoperative Diagnostic and Prognostic Prediction in Epithelial Ovarian Cancer Based on Blood Biomarkers. Clin. Cancer Res. 2019, 25, 3006–3015. [Google Scholar] [CrossRef] [Green Version]
- Su, T.H.; Wu, C.H.; Kao, J.H. Artificial intelligence in precision medicine in hepatology. J. Gastroenterol. Hepatol. 2021, 36, 569–580. [Google Scholar] [CrossRef]
- Yu, S.C.; Chang, K.C.; Wang, H.; Li, M.F.; Yang, T.L.; Chen, C.N.; Chen, C.J.; Chen, K.C.; Shen, C.Y.; Kuo, P.Y.; et al. Distinguishing lupus lymphadenitis from Kikuchi disease based on clinicopathological features and C4d immunohistochemistry. Rheumatology 2021, 60, 1543–1552. [Google Scholar] [CrossRef]
- Kawaguchi, Y.; Shimada, Y.; Murakami, K.; Omori, T.; Kudo, Y.; Makino, Y.; Maehara, S.; Hagiwara, M.; Kakihana, M.; Yamada, T.; et al. Prognostic impact of artificial intelligence-based volumetric quantification of the solid part of the tumor in clinical stage 0-I adenocarcinoma. Lung Cancer 2022, 170, 85–90. [Google Scholar] [CrossRef]
- Zimmer-Stelmach, A.; Zak, J.; Pawlosek, A.; Rosner-Tenerowicz, A.; Budny-Winska, J.; Pomorski, M.; Fuchs, T.; Zimmer, M. The Application of Artificial Intelligence-Assisted Colposcopy in a Tertiary Care Hospital within a Cervical Pathology Diagnostic Unit. Diagnostics 2022, 12, 106. [Google Scholar] [CrossRef]
- Cancian, P.; Cortese, N.; Donadon, M.; Di Maio, M.; Soldani, C.; Marchesi, F.; Savevski, V.; Santambrogio, M.D.; Cerina, L.; Laino, M.E.; et al. Development of a Deep-Learning Pipeline to Recognize and Characterize Macrophages in Colo-Rectal Liver Metastasis. Cancers 2021, 13, 3313. [Google Scholar] [CrossRef] [PubMed]
- Carreras, J.; Hiraiwa, S.; Kikuti, Y.Y.; Miyaoka, M.; Tomita, S.; Ikoma, H.; Ito, A.; Kondo, Y.; Roncador, G.; Garcia, J.F.; et al. Artificial Neural Networks Predicted the Overall Survival and Molecular Subtypes of Diffuse Large B-Cell Lymphoma Using a Pancancer Immune-Oncology Panel. Cancers 2021, 13, 6384. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, S.; Liang, Y.; Tan, Y.; Li, L.; Liu, Q.; Liu, T.; Lu, X. Small Tweaks, Major Changes: Post-Translational Modifications That Occur within M2 Macrophages in the Tumor Microenvironment. Cancers 2022, 14, 5532. https://doi.org/10.3390/cancers14225532
Zheng S, Liang Y, Tan Y, Li L, Liu Q, Liu T, Lu X. Small Tweaks, Major Changes: Post-Translational Modifications That Occur within M2 Macrophages in the Tumor Microenvironment. Cancers. 2022; 14(22):5532. https://doi.org/10.3390/cancers14225532
Chicago/Turabian StyleZheng, Shutao, Yan Liang, Yiyi Tan, Lu Li, Qing Liu, Tao Liu, and Xiaomei Lu. 2022. "Small Tweaks, Major Changes: Post-Translational Modifications That Occur within M2 Macrophages in the Tumor Microenvironment" Cancers 14, no. 22: 5532. https://doi.org/10.3390/cancers14225532
APA StyleZheng, S., Liang, Y., Tan, Y., Li, L., Liu, Q., Liu, T., & Lu, X. (2022). Small Tweaks, Major Changes: Post-Translational Modifications That Occur within M2 Macrophages in the Tumor Microenvironment. Cancers, 14(22), 5532. https://doi.org/10.3390/cancers14225532