
The Role of Single-Nucleotide Polymorphisms in Cholangiocarcinoma: A Systematic Review
 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
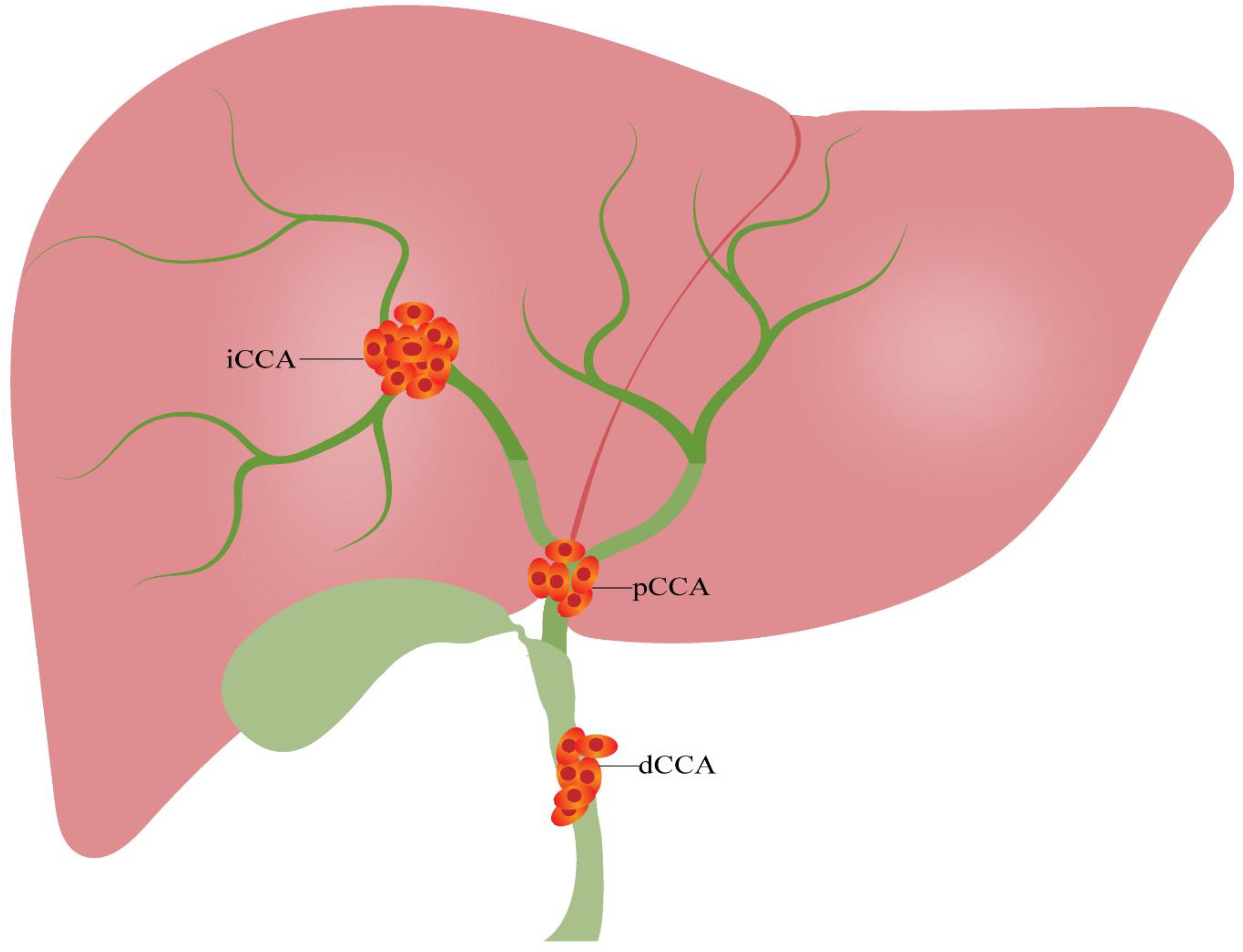
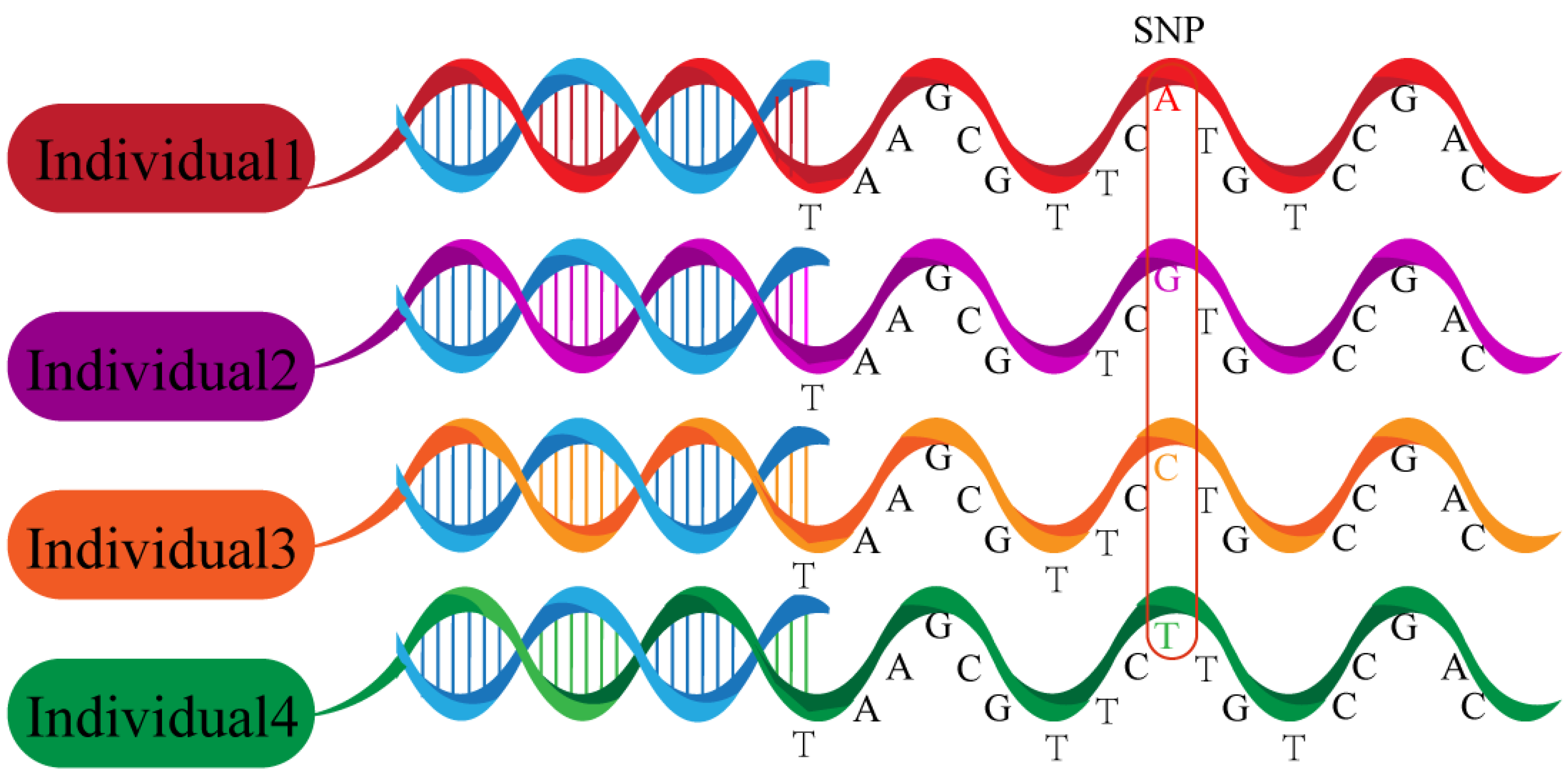
1. Introduction
2. Methods
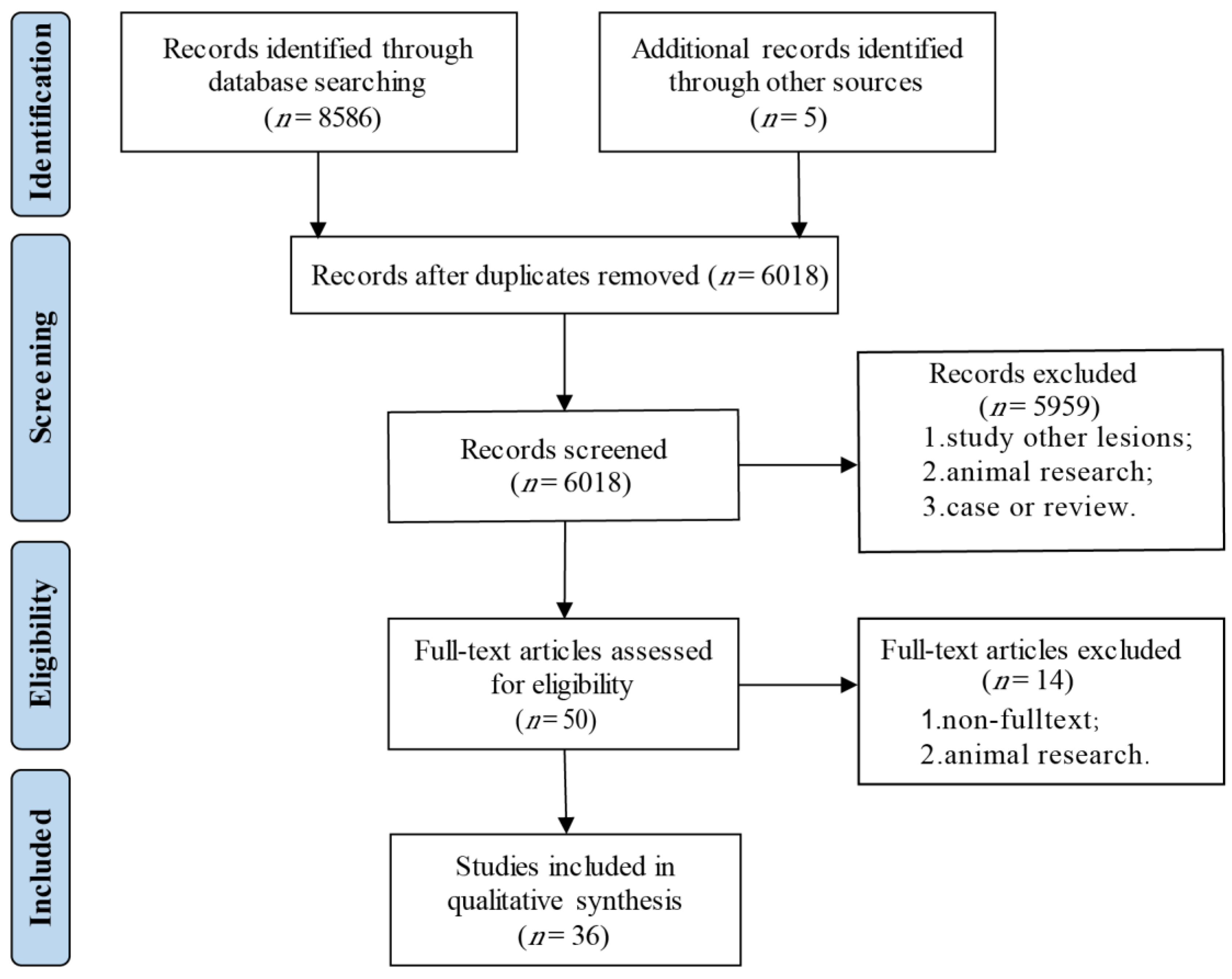
2.1. Registration and Protocol
2.2. Search Strategy
2.3. Inclusion and Exclusion Criteria
2.4. Data Extraction
2.5. Risk of Bias in Individual Studies
3. Results
3.1. Association of SNPs with Susceptibility to Cholangiocarcinoma
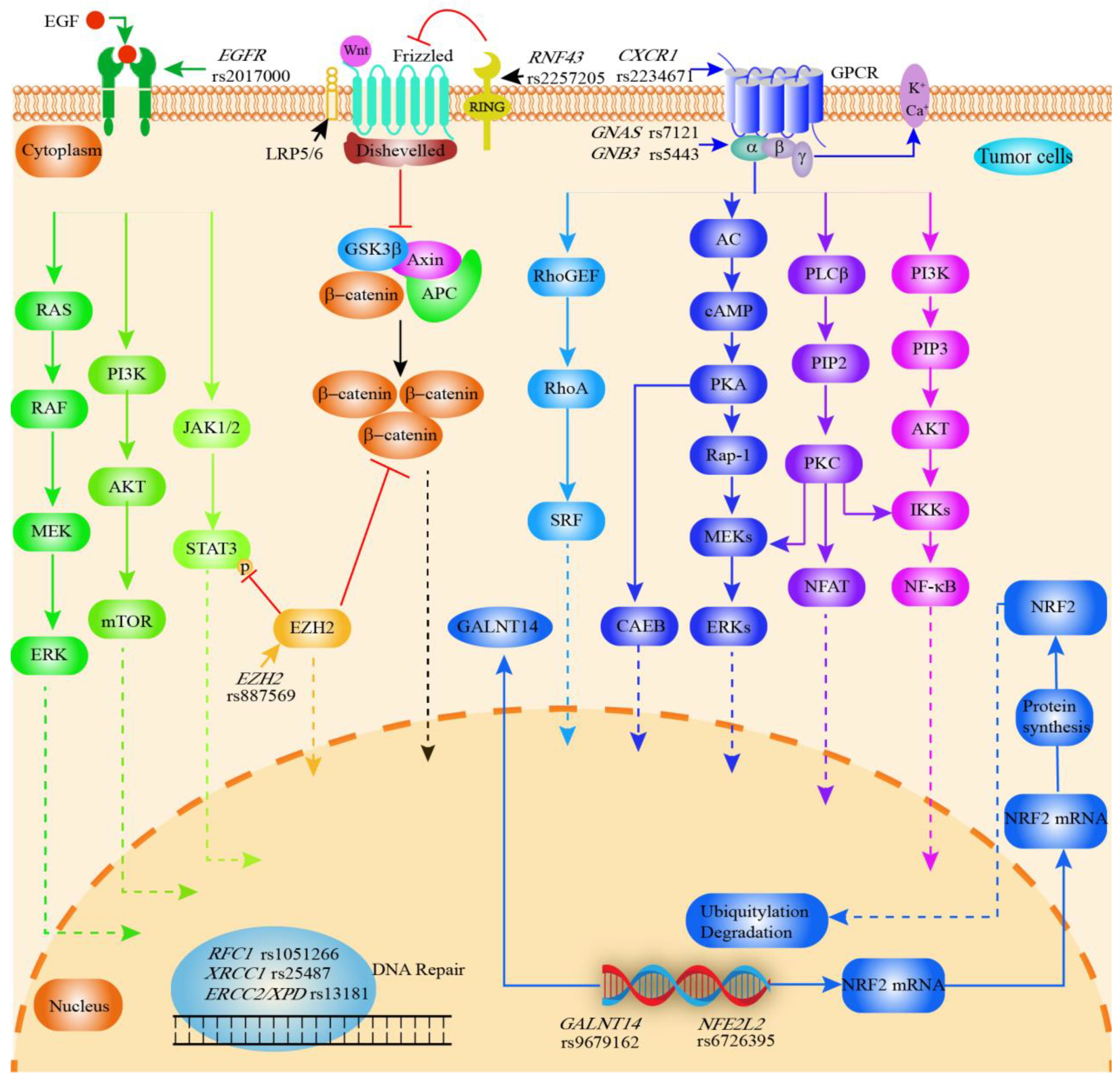
3.1.1. Inflammation-Related Genes
3.1.2. DNA Repair Genes
3.1.3. Cellular Protection against Toxin Genes
3.1.4. Other Enzyme-Related Genes
3.1.5. RNA-Related Polymorphisms
3.1.6. Membrane-Protein-Related Gene Polymorphisms
3.2. Correlation of SNPs with Cholangiocarcinoma Invasion and Metastasis
3.3. Relationship between SNPs and Prognosis of Cholangiocarcinoma
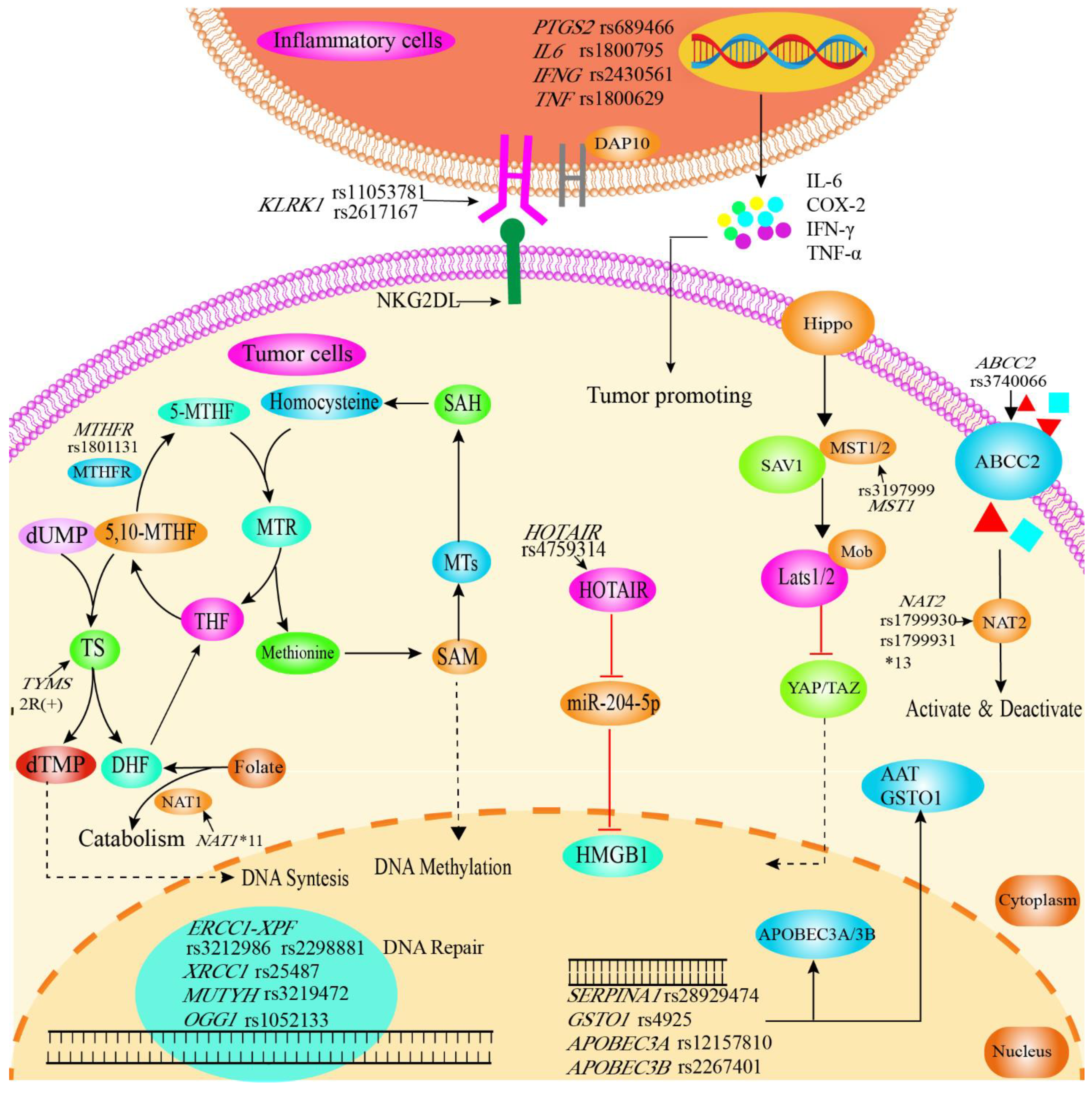
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kambakamba, P.; DeOliveira, M.L. Perihilar cholangiocarcinoma: Paradigms of surgical management. Am. J. Surg. 2014, 208, 563–570. [Google Scholar] [CrossRef]
- Banales, J.M.; Marin, J.J.G.; Lamarca, A.; Rodrigues, P.M.; Khan, S.A.; Roberts, L.R.; Cardinale, V.; Carpino, G.; Andersen, J.B.; Braconi, C.; et al. Cholangiocarcinoma 2020: The next horizon in mechanisms and management. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 557–588. [Google Scholar] [CrossRef]
- Blechacz, B.; Komuta, M.; Roskams, T.; Gores, G.J. Clinical diagnosis and staging of cholangiocarcinoma. Nat. Rev. Gastroenterol. Hepatol 2011, 8, 512–522. [Google Scholar] [CrossRef] [Green Version]
- Global Burden of Disease Cancer Collaboration. The Global Burden of Cancer 2013. JAMA Oncol. 2015, 1, 505–527. [Google Scholar] [CrossRef]
- Saha, S.K.; Zhu, A.X.; Fuchs, C.S.; Brooks, G.A.J.T. Forty-year trends in cholangiocarcinoma incidence in the US: Intrahepatic disease on the rise. Oncologist 2016, 21, 594. [Google Scholar] [CrossRef] [Green Version]
- Bertuccio, P.; Bosetti, C.; Levi, F.; Decarli, A.; Negri, E.; La Vecchia, C. A comparison of trends in mortality from primary liver cancer and intrahepatic cholangiocarcinoma in Europe. Ann. Oncol. 2013, 24, 1667–1674. [Google Scholar] [CrossRef]
- Patel, T. Increasing incidence and mortality of primary intrahepatic cholangiocarcinoma in the United States. Hepatology 2001, 33, 1353–1357. [Google Scholar] [CrossRef]
- Bertuccio, P.; Malvezzi, M.; Carioli, G.; Hashim, D.; Boffetta, P.; El-Serag, H.B.; La Vecchia, C.; Negri, E. Global trends in mortality from intrahepatic and extrahepatic cholangiocarcinoma. J. Hepatol. 2019, 71, 104–114. [Google Scholar] [CrossRef]
- Banales, J.M.; Cardinale, V.; Carpino, G.; Marzioni, M.; Andersen, J.B.; Invernizzi, P.; Lind, G.E.; Folseraas, T.; Forbes, S.J.; Fouassier, L.; et al. Expert consensus document: Cholangiocarcinoma: Current knowledge and future perspectives consensus statement from the European Network for the Study of Cholangiocarcinoma (ENS-CCA). Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 261–280. [Google Scholar] [CrossRef]
- Alabraba, E.; Joshi, H.; Bird, N.; Griffin, R.; Sturgess, R.; Stern, N.; Sieberhagen, C.; Cross, T.; Camenzuli, A.; Davis, R.; et al. Increased multimodality treatment options has improved survival for Hepatocellular carcinoma but poor survival for biliary tract cancers remains unchanged. Eur. J. Surg. Oncol. 2019, 45, 1660–1667. [Google Scholar] [CrossRef]
- Komaya, K.; Ebata, T.; Yokoyama, Y.; Igami, T.; Sugawara, G.; Mizuno, T.; Yamaguchi, J.; Nagino, M. Recurrence after curative-intent resection of perihilar cholangiocarcinoma: Analysis of a large cohort with a close postoperative follow-up approach. Surgery 2018, 163, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Lander, E.S. The new genomics: Global views of biology. Science 1996, 274, 536–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International HapMap, C.; Frazer, K.A.; Ballinger, D.G.; Cox, D.R.; Hinds, D.A.; Stuve, L.L.; Gibbs, R.A.; Belmont, J.W.; Boudreau, A.; Hardenbol, P.; et al. A second generation human haplotype map of over 3.1 million SNPs. Nature 2007, 449, 851–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, F.; Pelletier, J. Exploring the Impact of Single-Nucleotide Polymorphisms on Translation. Front. Genet. 2018, 9, 507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, N.K.; Scott, R.J.; Rowlings, P.; Enjeti, A.K. Clinical use of SNP-microarrays for the detection of genome-wide changes in haematological malignancies. Crit. Rev. Oncol. Hematol. 2019, 142, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Int. J. Surg. 2021, 88, 105906. [Google Scholar] [CrossRef] [PubMed]
- Deeks, J.J.; Dinnes, J.; D’Amico, R.; Sowden, A.J.; Sakarovitch, C.; Song, F.; Petticrew, M.; Altman, D.G.; International Stroke Trial Collaborative Group; European Carotid Surgery Trial Collaborative Group. Evaluating non-randomised intervention studies. Health Technol. Assess 2003, 7, 1–173. [Google Scholar] [CrossRef] [Green Version]
- Luo, A.H.; Zhang, G.F.; Yin, Y.F.; Li, X.; Feng, D.L.; Xie, C.H. Complete sequence of a cholangiocarcinoma inbred Sprague-Dawley rat model mitochondrial genome. Mitochondrial DNA A DNA Mapp. Seq. Anal. 2016, 27, 1732–1733. [Google Scholar] [CrossRef]
- Krawczyk, M.; Mihalache, F.; Hoblinger, A.; Acalovschi, M.; Lammert, F.; Zimmer, V. Pancreatic cancer risk variant ABO rs505922 in patients with cholangiocarcinoma. World J. Gastroenterol. 2011, 17, 4640–4642. [Google Scholar] [CrossRef]
- Li, J.S.; Song, Y.X.; Han, T.J.; Liu, L.; Gao, X.Y.; Jing, N.; Li, L.; Sui, S.J.; Wang, Q.C. FTO gene polymorphisms and cholangiocarcinoma risk: A case-control study. Int. J. Clin. Exp. Med. 2016, 9, 19974–19980. [Google Scholar]
- Wadsworth, C.A.; Dixon, P.H.; Taylor-Robinson, S.; Kim, J.U.; Zabron, A.A.; Wong, J.H.; Chapman, M.H.; McKay, S.C.; Spalding, D.R.; Wasan, H.S.; et al. Polymorphisms in Natural Killer Cell Receptor Protein 2D (NKG2D) as a Risk Factor for Cholangiocarcinoma. J. Clin. Exp. Hepatol. 2019, 9, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; You, G.; Tanaka, H.; Srivatanakul, P.; Ohta, E.; Viwatthanasittiphong, C.; Matharit, M.; Chenvidhya, D.; Jedpiyawongse, A.; Tanaka, M.; et al. Combined effects of polymorphisms of DNA-repair protein genes and metabolic enzyme genes on the risk of cholangiocarcinoma. Jpn. J. Clin. Oncol. 2013, 43, 1190–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmer, V.; Hoblinger, A.; Mihalache, F.; Assmann, G.; Acalovschi, M.; Lammert, F. Potential genotype-specific single nucleotide polymorphism interaction of common variation in p53 and its negative regulator mdm2 in cholangiocarcinoma susceptibility. Oncol. Lett. 2012, 4, 101–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riedmaier, S.; Klein, K.; Winter, S.; Hofmann, U.; Schwab, M.; Zanger, U.M. Paraoxonase (PON1 and PON3) Polymorphisms: Impact on Liver Expression and Atorvastatin-Lactone Hydrolysis. Front. Pharmacol. 2011, 2, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsing, A.W.; Sakoda, L.C.; Rashid, A.; Andreotti, G.; Chen, J.; Wang, B.S.; Shen, M.C.; Chen, B.E.; Rosenberg, P.S.; Zhang, M.; et al. Variants in inflammation genes and the risk of biliary tract cancers and stones: A population-based study in China. Cancer Res. 2008, 68, 6442–6452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaiteerakij, R.; Juran, B.D.; Aboelsoud, M.M.; Harmsen, W.S.; Moser, C.D.; Giama, N.H.; Allotey, L.K.; Mettler, T.A.; Baichoo, E.; Zhang, X.; et al. Association between variants in inflammation and cancer-associated genes and risk and survival of cholangiocarcinoma. Cancer Med. 2015, 4, 1599–1602. [Google Scholar] [CrossRef]
- Fingas, C.D.; Katsounas, A.; Kahraman, A.; Siffert, W.; Jochum, C.; Gerken, G.; Nuckel, H.; Canbay, A. Prognostic assessment of three single-nucleotide polymorphisms (GNB3 825C>T, BCL2-938C>A, MCL1-386C>G) in extrahepatic cholangiocarcinoma. Cancer Investig. 2010, 28, 472–478. [Google Scholar] [CrossRef]
- Hoblinger, A.; Grunhage, F.; Sauerbruch, T.; Lammert, F. Association of the c.3972C>T variant of the multidrug resistance-associated protein 2 Gene (MRP2/ABCC2) with susceptibility to bile duct cancer. Digestion 2009, 80, 36–39. [Google Scholar] [CrossRef]
- Honjo, S.; Srivatanakul, P.; Sriplung, H.; Kikukawa, H.; Hanai, S.; Uchida, K.; Todoroki, T.; Jedpiyawongse, A.; Kittiwatanachot, P.; Sripa, B.; et al. Genetic and environmental determinants of risk for cholangiocarcinoma via Opisthorchis viverrini in a densely infested area in Nakhon Phanom, northeast Thailand. Int. J. Cancer 2005, 117, 854–860. [Google Scholar] [CrossRef]
- Khunluck, T.; Kukongviriyapan, V.; Puapairoj, A.; Khuntikeo, N.; Senggunprai, L.; Zeekpudsa, P.; Prawan, A. Association of NRF2 polymorphism with cholangiocarcinoma prognosis in Thai patients. Asian Pac. J. Cancer Prev. 2014, 15, 299–304. [Google Scholar] [CrossRef]
- Ko, K.H.; Kim, N.K.; Yim, D.J.; Hong, S.P.; Park, P.W.; Rim, K.S.; Kim, S.; Hwang, S.G. Polymorphisms of 5,10-methylenetetrahydrofolate reductase (MTHFR C677T) and thymidylate synthase enhancer region (TSER) as a risk factor of cholangiocarcinoma in a Korean population. Anticancer Res. 2006, 26, 4229–4233. [Google Scholar] [PubMed]
- Krawczyk, M.; Hoblinger, A.; Mihalache, F.; Grunhage, F.; Acalovschi, M.; Lammert, F.; Zimmer, V. Macrophage stimulating protein variation enhances the risk of sporadic extrahepatic cholangiocarcinoma. Dig. Liver Dis. 2013, 45, 612–615. [Google Scholar] [CrossRef] [PubMed]
- Lampropoulou, D.I.; Laschos, K.; Aravantinos, G.; Georgiou, K.; Papiris, K.; Theodoropoulos, G.; Gazouli, M.; Filippou, D. Association between homeobox protein transcript antisense intergenic ribonucleic acid genetic polymorphisms and cholangiocarcinoma. World J. Clin. Cases 2021, 9, 1785–1792. [Google Scholar] [CrossRef] [PubMed]
- Melum, E.; Karlsen, T.H.; Schrumpf, E.; Bergquist, A.; Thorsby, E.; Boberg, K.M.; Lie, B.A. Cholangiocarcinoma in primary sclerosing cholangitis is associated with NKG2D polymorphisms. Hepatology 2008, 47, 90–96. [Google Scholar] [CrossRef]
- Meng, L.Q. Essential role of polymorphism of Gab1, EGFR, and EGF for the susceptibility of biliary tract cancer. Tumour Biol. 2014, 35, 12497–12508. [Google Scholar] [CrossRef]
- Mihalache, F.; Höblinger, A.; Acalovschi, M.; Sauerbruch, T.; Lammert, F.; Zimmer, V. A common variant in the precursor miR-146a sequence does not predispose to cholangiocarcinoma in a large European cohort. Hepatobiliary Pancreat. Dis. Int. 2012, 11, 412–417. [Google Scholar] [CrossRef]
- Mihalache, F.; Hoblinger, A.; Grunhage, F.; Krawczyk, M.; Gartner, B.C.; Acalovschi, M.; Sauerbruch, T.; Lammert, F.; Zimmer, V. Heterozygosity for the alpha1-antitrypsin Z allele may confer genetic risk of cholangiocarcinoma. Aliment. Pharmacol. Ther. 2011, 33, 389–394. [Google Scholar] [CrossRef]
- Prawan, A.; Kukongviriyapan, V.; Tassaneeyakul, W.; Pairojkul, C.; Bhudhisawasdi, V. Association between genetic polymorphisms of CYP1A2, arylamine N-acetyltransferase 1 and 2 and susceptibility to cholangiocarcinoma. Eur. J. Cancer Prev. Off. J. Eur. Cancer Prev. Organ. (ECP) 2005, 14, 245–250. [Google Scholar] [CrossRef]
- Songserm, N.; Promthet, S.; Sithithaworn, P.; Pientong, C.; Ekalaksananan, T.; Chopjitt, P.; Parkin, D.M. MTHFR polymorphisms and Opisthorchis viverrini infection: A relationship with increased susceptibility to cholangiocarcinoma in Thailand. Asian Pac. J. Cancer Prev. 2011, 12, 1341–1345. [Google Scholar]
- Sun, K.; Wang, W.; Wang, F.; Xiu, P.; Wang, H.; Liu, F. Association between ERCC1 and XPF polymorphisms and risk of extrahepatic cholangiocarcinoma. Int. J. Clin. Exp. Med. 2018, 11, 13813–13819. [Google Scholar]
- Surapaitoon, A.; Suttiprapa, S.; Mairiang, E.; Khuntikeo, N.; Pairojkul, C.; Bethony, J.; Brindley, P.J.; Sripa, B. Subsets of Inflammatory Cytokine Gene Polymorphisms are Associated with Risk of Carcinogenic Liver Fluke Opisthorchis viverrini-Associated Advanced Periductal Fibrosis and Cholangiocarcinoma. Korean J. Parasitol. 2017, 55, 295–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, S.H.; Wang, X.; Huang, S.; Wang, M.; Ji, G.Z.; Xia, J.R.; Fan, Z.N. MYH rs3219476 and rs3219472 polymorphisms and risk of cholangiocarcinoma. Mol. Med. Rep. 2013, 7, 347–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.Q.; Ma, H.X.; Su, M.S.; He, L. Osteopontin promoter polymorphisms at locus -443 are associated with metastasis and poor prognosis of human intrahepatic cholangiocarcinoma in Chinese population. Int. J. Clin. Exp. Pathol. 2014, 7, 6914–6921. [Google Scholar] [PubMed]
- Songserm, N.; Promthet, S.; Pientong, C.; Ekalaksananan, T.; Chopjitt, P.; Wiangnon, S. Gene-environment interaction involved in cholangiocarcinoma in the Thai population: Polymorphisms of DNA repair genes, smoking and use of alcohol. BMJ Open 2014, 4, e005447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Ji, H.; Zhao, J.; Song, J.; Zheng, S.; Chen, L.; Li, P.; Tan, X.; Ding, Y.; Pu, R.; et al. Transcriptional repression and apoptosis influence the effect of APOBEC3A/3B functional polymorphisms on biliary tract cancer risk. Int. J. Cancer 2022, 150, 1825–1837. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.S.; Dageforde, L.A. Cholangiocarcinoma. Surg. Clin. N. Am. 2019, 99, 315–335. [Google Scholar] [CrossRef]
- Hung, J.H.; Su, I.J.; Lei, H.Y.; Wang, H.C.; Lin, W.C.; Chang, W.T.; Huang, W.; Chang, W.C.; Chang, Y.S.; Chen, C.C.; et al. Endoplasmic reticulum stress stimulates the expression of cyclooxygenase-2 through activation of NF-kappaB and pp38 mitogen-activated protein kinase. J. Biol. Chem. 2004, 279, 46384–46392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, C.; Pereira, C.; Farinha, M.; Medeiros, R.; Dinis-Ribeiro, M. Genetic Variations in Prostaglandin E(2) Pathway Identified as Susceptibility Biomarkers for Gastric Cancer in an Intermediate Risk European Country. Int. J. Mol. Sci. 2021, 22, 648. [Google Scholar] [CrossRef]
- Zhang, X.; Miao, X.; Tan, W.; Ning, B.; Liu, Z.; Hong, Y.; Song, W.; Guo, Y.; Zhang, X.; Shen, Y. Identification of functional genetic variants in cyclooxygenase-2 and their association with risk of esophageal cancer. Gastroenterology 2005, 129, 565–576. [Google Scholar] [CrossRef]
- Fisher, D.T.; Appenheimer, M.M.; Evans, S.S. The two faces of IL-6 in the tumor microenvironment. Semin. Immunol. 2014, 26, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Balkwill, F. TNF-alpha in promotion and progression of cancer. Cancer Metastasis Rev. 2006, 25, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Maderuelo, D.; Arnalich, F.; Serantes, R.; Gonzalez, A.; Codoceo, R.; Madero, R.; Vazquez, J.J.; Montiel, C. Interferon-gamma and interleukin-10 gene polymorphisms in pulmonary tuberculosis. Am. J. Respir. Crit. Care Med. 2003, 167, 970–975. [Google Scholar] [CrossRef] [PubMed]
- Fishman, D.; Faulds, G.; Jeffery, R.; Mohamed-Ali, V.; Yudkin, J.S.; Humphries, S.; Woo, P. The effect of novel polymorphisms in the interleukin-6 (IL-6) gene on IL-6 transcription and plasma IL-6 levels, and an association with systemic-onset juvenile chronic arthritis. J. Clin. Investig. 1998, 102, 1369–1376. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.G.; Symons, J.A.; McDowell, T.L.; McDevitt, H.O.; Duff, G.W. Effects of a polymorphism in the human tumor necrosis factor alpha promoter on transcriptional activation. Proc. Natl. Acad. Sci. USA 1997, 94, 3195–3199. [Google Scholar] [CrossRef] [Green Version]
- Sijbers, A.M.; de Laat, W.L.; Ariza, R.R.; Biggerstaff, M.; Wei, Y.F.; Moggs, J.G.; Carter, K.C.; Shell, B.K.; Evans, E.; de Jong, M.C.; et al. Xeroderma pigmentosum group F caused by a defect in a structure-specific DNA repair endonuclease. Cell 1996, 86, 811–822. [Google Scholar] [CrossRef] [Green Version]
- Marsh, S.; McKay, J.A.; Cassidy, J.; McLeod, H.L. Polymorphism in the thymidylate synthase promoter enhancer region in colorectal cancer. Int. J. Oncol. 2001, 19, 383–386. [Google Scholar] [CrossRef]
- van der Put, N.M.; Gabreels, F.; Stevens, E.M.; Smeitink, J.A.; Trijbels, F.J.; Eskes, T.K.; van den Heuvel, L.P.; Blom, H.J. A second common mutation in the methylenetetrahydrofolate reductase gene: An additional risk factor for neural-tube defects? Am. J. Hum. Genet. 1998, 62, 1044–1051. [Google Scholar] [CrossRef] [Green Version]
- Pieroth, R.; Paver, S.; Day, S.; Lammersfeld, C. Folate and Its Impact on Cancer Risk. Curr. Nutr. Rep. 2018, 7, 70–84. [Google Scholar] [CrossRef] [Green Version]
- Wood, R.D.; Mitchell, M.; Sgouros, J.; Lindahl, T. Human DNA repair genes. Science 2001, 291, 1284–1289. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Fatah, T.; Sultana, R.; Abbotts, R.; Hawkes, C.; Seedhouse, C.; Chan, S.; Madhusudan, S. Clinicopathological and functional significance of XRCC1 expression in ovarian cancer. Int. J. Cancer 2013, 132, 2778–2786. [Google Scholar] [CrossRef] [Green Version]
- Floris, M.; Sanna, D.; Castiglia, P.; Putzu, C.; Sanna, V.; Pazzola, A.; De Miglio, M.R.; Sanges, F.; Pira, G.; Azara, A.; et al. MTHFR, XRCC1 and OGG1 genetic polymorphisms in breast cancer: A case-control study in a population from North Sardinia. BMC Cancer 2020, 20, 234. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, M.; Osawa, K.; Yoshida, K.; Miyaishi, A.; Osawa, Y.; Inoue, N.; Tsutou, A.; Tabuchi, Y.; Tanaka, K.; Yamamoto, M.; et al. Association of MUTYH Gln324His and APEX1 Asp148Glu with colorectal cancer and smoking in a Japanese population. J. Exp. Clin. Cancer Res. 2008, 27, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoer, N.C.; Botteri, E.; Thoresen, G.H.; Karlstad, O.; Weiderpass, E.; Friis, S.; Pottegard, A.; Andreassen, B.K. Drug Use and Cancer Risk: A Drug-Wide Association Study (DWAS) in Norway. Cancer Epidemiol. Biomark. Prev. 2021, 30, 682–689. [Google Scholar] [CrossRef] [PubMed]
- Vineis, P. Epidemiology of cancer from exposure to arylamines. Environ. Health Perspect. 1994, 102 (Suppl. S6), 7–10. [Google Scholar] [CrossRef]
- Hein, D.W.; Doll, M.A.; Fretland, A.J.; Leff, M.A.; Webb, S.J.; Xiao, G.H.; Devanaboyina, U.S.; Nangju, N.A.; Feng, Y. Molecular genetics and epidemiology of the NAT1 and NAT2 acetylation polymorphisms. Cancer Epidemiol. Biomark. Prev. 2000, 9, 29–42. [Google Scholar]
- Guengerich, F.P. Intersection of the Roles of Cytochrome P450 Enzymes with Xenobiotic and Endogenous Substrates: Relevance to Toxicity and Drug Interactions. Chem. Res. Toxicol. 2017, 30, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Nies, A.T.; Keppler, D. The apical conjugate efflux pump ABCC2 (MRP2). Pflug. Arch. 2007, 453, 643–659. [Google Scholar] [CrossRef]
- Brandi, G.; Rizzo, A.; Deserti, M.; Relli, V.; Indio, V.; Bin, S.; Pariali, M.; Palloni, A.; De Lorenzo, S.; Tovoli, F.; et al. Wilson disease, ABCC2 c.3972C > T polymorphism and primary liver cancers: Suggestions from a familial cluster. BMC Med. Genet. 2020, 21, 225. [Google Scholar] [CrossRef]
- Sookoian, S.; Castano, G.; Burgueno, A.; Gianotti, T.F.; Pirola, C.J. Association of the multidrug-resistance-associated protein gene (ABCC2) variants with intrahepatic cholestasis of pregnancy. J. Hepatol. 2008, 48, 125–132. [Google Scholar] [CrossRef]
- Meng, L.; Tian, Z.; Wang, Y.; Liu, Y.; Liu, J. Predictive and prognostic molecular markers for cholangiocarcinoma in Han Chinese population. Int. J. Clin. Exp. Med. 2015, 8, 13680–13689. [Google Scholar]
- Bulus, H.; Oguztuzun, S.; Güler Simsek, G.; Kilic, M.; Ada, A.O.; Göl, S.; Kocdogan, A.K.; Kaygın, P.; Bozer, B.; Iscan, M. Expression of CYP and GST in human normal and colon tumor tissues. Biotech. Histochem. 2019, 94, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chuang, J.J.; Dai, Y.C.; Lin, Y.L.; Chen, Y.Y.; Lin, W.H.; Chan, H.L.; Liu, Y.W. Downregulation of glutathione S-transferase M1 protein in N-butyl-N-(4-hydroxybutyl)nitrosamine-induced mouse bladder carcinogenesis. Toxicol. Appl. Pharmacol. 2014, 279, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Marahatta, S.B.; Punyarit, P.; Bhudisawasdi, V.; Paupairoj, A.; Wongkham, S.; Petmitr, S. Polymorphism of glutathione S-transferase omega gene and risk of cancer. Cancer Lett. 2006, 236, 276–281. [Google Scholar] [CrossRef]
- Melum, E.; Franke, A.; Schramm, C.; Weismuller, T.J.; Gotthardt, D.N.; Offner, F.A.; Juran, B.D.; Laerdahl, J.K.; Labi, V.; Bjornsson, E.; et al. Genome-wide association analysis in primary sclerosing cholangitis identifies two non-HLA susceptibility loci. Nat. Genet. 2011, 43, 17–19. [Google Scholar] [CrossRef] [Green Version]
- Karlsen, T.H.; Folseraas, T.; Thorburn, D.; Vesterhus, M. Primary sclerosing cholangitis-a comprehensive review. J. Hepatol. 2017, 67, 1298–1323. [Google Scholar] [CrossRef]
- Kim, E.Y.; Lorenzo-Redondo, R.; Little, S.J.; Chung, Y.S.; Phalora, P.K.; Maljkovic Berry, I.; Archer, J.; Penugonda, S.; Fischer, W.; Richman, D.D.; et al. Human APOBEC3 induced mutation of human immunodeficiency virus type-1 contributes to adaptation and evolution in natural infection. PLoS Pathog. 2014, 10, e1004281. [Google Scholar] [CrossRef] [Green Version]
- Bohn, M.F.; Shandilya, S.M.D.; Silvas, T.V.; Nalivaika, E.A.; Kouno, T.; Kelch, B.A.; Ryder, S.P.; Kurt-Yilmaz, N.; Somasundaran, M.; Schiffer, C.A. The ssDNA Mutator APOBEC3A Is Regulated by Cooperative Dimerization. Structure 2015, 23, 903–911. [Google Scholar] [CrossRef] [Green Version]
- Olson, M.E.; Harris, R.S.; Harki, D.A. APOBEC enzymes as targets for virus and cancer therapy. Cell Chem. Biol. 2018, 25, 36–49. [Google Scholar] [CrossRef] [Green Version]
- Burns, M.B.; Temiz, N.A.; Harris, R.S. Evidence for APOBEC3B mutagenesis in multiple human cancers. Nat. Genet. 2013, 45, 977–983. [Google Scholar] [CrossRef] [Green Version]
- Wickramasinghe, V.O.; Laskey, R.A. Control of mammalian gene expression by selective mRNA export. Nat. Rev. Mol. Cell Biol. 2015, 16, 431–442. [Google Scholar] [CrossRef]
- Dvinge, H.; Guenthoer, J.; Porter, P.L.; Bradley, R.K. RNA components of the spliceosome regulate tissue- and cancer-specific alternative splicing. Genome Res. 2019, 29, 1591–1604. [Google Scholar] [CrossRef] [Green Version]
- Qin, W.; Kang, P.; Xu, Y.; Leng, K.; Li, Z.; Huang, L.; Gao, J.; Cui, Y.; Zhong, X. Long non-coding RNA HOTAIR promotes tumorigenesis and forecasts a poor prognosis in cholangiocarcinoma. Sci. Rep. 2018, 8, 12176. [Google Scholar] [CrossRef]
- Pan, W.; Liu, L.; Wei, J.; Ge, Y.; Zhang, J.; Chen, H.; Zhou, L.; Yuan, Q.; Zhou, C.; Yang, M. A functional lncRNA HOTAIR genetic variant contributes to gastric cancer susceptibility. Mol. Carcinog. 2016, 55, 90–96. [Google Scholar] [CrossRef]
- Pan, W.; Wu, C.; Su, Z.; Duan, Z.; Li, L.; Mi, F.; Li, C. Genetic polymorphisms of non-coding RNAs associated with increased head and neck cancer susceptibility: A systematic review and meta-analysis. Oncotarget 2017, 8, 62508–62523. [Google Scholar] [CrossRef] [Green Version]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef]
- Xiong, X.; Yan, J.; Li, L.; Li, Y.; Cao, Y.; Tu, Y.; Mei, J. Relationship between miR-146a rs2910164 (G > C) polymorphism and digestive system cancer susceptibility: A meta-analysis. Ann. Clin. Lab. Sci. 2017, 47, 491–500. [Google Scholar]
- Arinaminpathy, Y.; Khurana, E.; Engelman, D.M.; Gerstein, M.B. Computational analysis of membrane proteins: The largest class of drug targets. Drug Discov. Today 2009, 14, 1130–1135. [Google Scholar] [CrossRef] [Green Version]
- Coudert, J.D.; Held, W. The role of the NKG2D receptor for tumor immunity. Semin. Cancer Biol. 2006, 16, 333–343. [Google Scholar] [CrossRef]
- Hayakawa, Y.; Smyth, M.J. NKG2D and cytotoxic effector function in tumor immune surveillance. Semin. Immunol. 2006, 18, 176–185. [Google Scholar] [CrossRef]
- Gu, H.; Neel, B.G. The “Gab” in signal transduction. Trends Cell Biol. 2003, 13, 122–130. [Google Scholar] [CrossRef]
- Sebastian, S.; Settleman, J.; Reshkin, S.J.; Azzariti, A.; Bellizzi, A.; Paradiso, A. The complexity of targeting EGFR signalling in cancer: From expression to turnover. Biochim. Biophys. Acta 2006, 1766, 120–139. [Google Scholar] [CrossRef]
- Silsirivanit, A. Glycosylation markers in cancer. Adv. Clin. Chem. 2019, 89, 189–213. [Google Scholar] [CrossRef]
- Rottger, S.; White, J.; Wandall, H.H.; Olivo, J.C.; Stark, A.; Bennett, E.P.; Whitehouse, C.; Berger, E.G.; Clausen, H.; Nilsson, T. Localization of three human polypeptide GalNAc-transferases in HeLa cells suggests initiation of O-linked glycosylation throughout the Golgi apparatus. J. Cell Sci. 1998, 111 Pt 1, 45–60. [Google Scholar] [CrossRef]
- Song, K.H.; Park, M.S.; Nandu, T.S.; Gadad, S.; Kim, S.C.; Kim, M.Y. GALNT14 promotes lung-specific breast cancer metastasis by modulating self-renewal and interaction with the lung microenvironment. Nat. Commun. 2016, 7, 13796. [Google Scholar] [CrossRef]
- Liang, K.H.; Yeh, T.S.; Wu, R.C.; Yeh, C.N.; Yeh, C.T. GALNT14 genotype is associated with perineural invasion, lymph node metastasis and overall survival in resected cholangiocarcinoma. Oncol. Lett. 2017, 13, 4215–4223. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Chen, W.; Athavale, D.; Ge, X.; Desert, R.; Das, S.; Han, H.; Nieto, N. Osteopontin Takes Center Stage in Chronic Liver Disease. Hepatology 2021, 73, 1594–1608. [Google Scholar] [CrossRef]
- Rittling, S.R.; Singh, R. Osteopontin in Immune-mediated Diseases. J. Dent. Res. 2015, 94, 1638–1645. [Google Scholar] [CrossRef] [Green Version]
- Pacetti, P.; Giovannetti, E.; Mambrini, A.; Nannizzi, S.; Orlandi, M.; Tartarini, R.; Del Freo, A.; Del Tacca, M.; Danesi, R.; Cantore, M. Single nucleotide polymorphisms and clinical outcome in patients with biliary tract carcinoma treated with epirubicin, cisplatin and capecitabine. Anticancer Res. 2009, 29, 1835–1840. [Google Scholar]
- Paolicchi, E.; Pacetti, P.; Giovannetti, E.; Mambrini, A.; Orlandi, M.; Crea, F.; Romani, A.A.; Tartarini, R.; Danesi, R.; Peters, G.J.; et al. A single nucleotide polymorphism in EZH2 predicts overall survival rate in patients with cholangiocarcinoma. Oncol. Lett. 2013, 6, 1487–1491. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, K.J.; Lang, H.; Frey, U.H.; Sotiropoulos, G.C.; Wohlschlaeger, J.; Reis, H.; Takeda, A.; Siffert, W.; Schmid, K.W.; Baba, H.A. GNAS1 T393C polymorphism is associated with clinical course in patients with intrahepatic cholangiocarcinoma. Neoplasia 2007, 9, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Lurje, I.; Czigany, Z.; Bednarsch, J.; Gaisa, N.T.; Dahl, E.; Knuchel, R.; Miller, H.; Ulmer, T.F.; Strnad, P.; Trautwein, C.; et al. Genetic Variant of CXCR1 (rs2234671) Associates with Clinical Outcome in Perihilar Cholangiocarcinoma. Liver Cancer 2022, 11, 162–173. [Google Scholar] [CrossRef]
- Rosskopf, D.; Busch, S.; Manthey, I.; Siffert, W. G protein beta 3 gene: Structure, promoter, and additional polymorphisms. Hypertension 2000, 36, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Siffert, W.; Rosskopf, D.; Moritz, A.; Wieland, T.; Kaldenberg-Stasch, S.; Kettler, N.; Hartung, K.; Beckmann, S.; Jakobs, K. Enhanced G protein activation in immortalized lymphoblasts from patients with essential hypertension. J. Clin. Investig. 1995, 96, 759–766. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Yamamoto, M. Nrf2-Keap1 regulation of cellular defense mechanisms against electrophiles and reactive oxygen species. Adv. Enzym. Regul. 2006, 46, 113–140. [Google Scholar] [CrossRef]
- Caldecott, K.W. XRCC1 and DNA strand break repair. DNA Repair 2003, 2, 955–969. [Google Scholar] [CrossRef]
- Gong, Y.; Qi, M.; Chen, J.; Fang, R.; Mai, C.; Chen, T.; Tang, H.; Tang, Y. XRCC1 Arg194Trp and Arg399Gln polymorphisms and risk of extrahepatic cholangiocarcinoma: A hospital-based case-control study in China. Int. J. Clin. Exp. Med. 2015, 8, 19339–19345. [Google Scholar]
- Nakagawa, S.; Okabe, H.; Sakamoto, Y.; Hayashi, H.; Hashimoto, D.; Yokoyama, N.; Sakamoto, K.; Kuroki, H.; Mima, K.; Nitta, H.; et al. Enhancer of zeste homolog 2 (EZH2) promotes progression of cholangiocarcinoma cells by regulating cell cycle and apoptosis. Ann. Surg. Oncol. 2013, 20 (Suppl. S3), S667–S675. [Google Scholar] [CrossRef]
- Vashist, Y.K.; Kutup, A.; Musici, S.; Yekebas, E.F.; Mina, S.; Uzunoglu, G.; Zehler, O.; Koenig, A.; Cataldegirmen, G.; Bockhorn, M.; et al. The GNAS1 T393C single nucleotide polymorphism predicts the natural postoperative course of complete resected esophageal cancer. Cell Oncol. 2011, 34, 281–288. [Google Scholar] [CrossRef]
- Moruzzi, S.; Udali, S.; Guarini, P.; Guglielmi, A.; Ruzzenente, A.; Pattini, P.; Conci, S.; Tammen, S.; Olivieri, O.; Choi, S.W.; et al. The RFC1 80g>a relates to survival rate according to PBMCS DNA global methylation in primary liver cancer. FASEB J. 2015, 29, e0167534. [Google Scholar] [CrossRef]
- Zebisch, M.; Xu, Y.; Krastev, C.; MacDonald, B.T.; Chen, M.; Gilbert, R.J.; He, X.; Jones, E.Y. Structural and molecular basis of ZNRF3/RNF43 transmembrane ubiquitin ligase inhibition by the Wnt agonist R-spondin. Nat. Commun. 2013, 4, 2787. [Google Scholar] [CrossRef] [Green Version]
- Xing, C.; Zhou, W.; Ding, S.; Xie, H.; Zhang, W.; Yang, Z.; Wei, B.; Chen, K.; Su, R.; Cheng, J.; et al. Reversing effect of ring finger protein 43 inhibition on malignant phenotypes of human hepatocellular carcinoma. Mol. Cancer Ther. 2013, 12, 94–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talabnin, C.; Janthavon, P.; Thongsom, S.; Suginta, W.; Talabnin, K.; Wongkham, S. Ring finger protein 43 expression is associated with genetic alteration status and poor prognosis among patients with intrahepatic cholangiocarcinoma. Hum. Pathol. 2016, 52, 47–54. [Google Scholar] [CrossRef]
- Hosono, M.; Koma, Y.I.; Takase, N.; Urakawa, N.; Higashino, N.; Suemune, K.; Kodaira, H.; Nishio, M.; Shigeoka, M.; Kakeji, Y.; et al. CXCL8 derived from tumor-associated macrophages and esophageal squamous cell carcinomas contributes to tumor progression by promoting migration and invasion of cancer cells. Oncotarget 2017, 8, 106071–106088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Li, A.; Tian, Y.; Wu, J.D.; Liu, Y.; Li, T.; Chen, Y.; Han, X.; Wu, K. The CXCL8-CXCR1/2 pathways in cancer. Cytokine Growth Factor Rev. 2016, 31, 61–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.H.; Das, B.B.; Casagrande, F.; Tian, Y.; Nothnagel, H.J.; Chu, M.; Kiefer, H.; Maier, K.; De Angelis, A.A.; Marassi, F.M.; et al. Structure of the chemokine receptor CXCR1 in phospholipid bilayers. Nature 2012, 491, 779–783. [Google Scholar] [CrossRef] [Green Version]
- Razumilava, N.; Gores, G.J. Cholangiocarcinoma. Lancet 2014, 383, 2168–2179. [Google Scholar] [CrossRef] [Green Version]
- Petrick, J.L.; Campbell, P.T.; Koshiol, J.; Thistle, J.E.; Andreotti, G.; Beane-Freeman, L.E.; Buring, J.E.; Chan, A.T.; Chong, D.Q.; Doody, M.M. Tobacco, alcohol use and risk of hepatocellular carcinoma and intrahepatic cholangiocarcinoma: The Liver Cancer Pooling Project. Br. J. Cancer 2018, 118, 1005–1012. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.A.; Tavolari, S.; Brandi, G. Cholangiocarcinoma: Epidemiology and risk factors. Liver Int. Off. J. Int. Assoc. Study Liver 2019, 39 (Suppl. S1), 19–31. [Google Scholar] [CrossRef]
- Viet, N.H.; Trung, N.Q.; Dong, L.T.; Trung, L.Q.; Espinoza, J.L. Genetic variants in NKG2D axis and susceptibility to Epstein-Barr virus-induced nasopharyngeal carcinoma. J. Cancer Res. Clin. Oncol. 2021, 147, 713–723. [Google Scholar] [CrossRef]
- Gimeno, L.; Martinez-Banaclocha, H.; Bernardo, M.V.; Bolarin, J.M.; Marin, L.; Lopez-Hernandez, R.; Lopez-Alvarez, M.R.; Moya-Quiles, M.R.; Muro, M.; Frias-Iniesta, J.F.; et al. NKG2D Polymorphism in Melanoma Patients from Southeastern Spain. Cancers 2019, 11, 438. [Google Scholar] [CrossRef] [Green Version]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.J.; Humphries, S.E. Cytokine and cytokine receptor gene polymorphisms and their functionality. Cytokine Growth Factor Rev. 2009, 20, 43–59. [Google Scholar] [CrossRef] [PubMed]
- Hakem, R. DNA-damage repair; the good, the bad, and the ugly. EMBO J. 2008, 27, 589–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, R.; Zhou, P.K. DNA damage repair: Historical perspectives, mechanistic pathways and clinical translation for targeted cancer therapy. Signal Transduct. Target. Ther. 2021, 6, 254. [Google Scholar] [CrossRef] [PubMed]
- Goode, E.L.; Ulrich, C.M.; Potter, J.D. Polymorphisms in DNA repair genes and associations with cancer risk. Cancer Epidemiol. Biomark. Prev. 2002, 11, 1513–1530. [Google Scholar]
- Houtsmuller, A.B.; Rademakers, S.; Nigg, A.L.; Hoogstraten, D.; Hoeijmakers, J.H.; Vermeulen, W. Action of DNA repair endonuclease ERCC1/XPF in living cells. Science 1999, 284, 958–961. [Google Scholar] [CrossRef]
- Zienolddiny, S.; Campa, D.; Lind, H.; Ryberg, D.; Skaug, V.; Stangeland, L.; Phillips, D.H.; Canzian, F.; Haugen, A. Polymorphisms of DNA repair genes and risk of non-small cell lung cancer. Carcinogenesis 2006, 27, 560–567. [Google Scholar] [CrossRef]
- He, J.; Zhuo, Z.J.; Zhang, A.; Zhu, J.; Hua, R.X.; Xue, W.Q.; Zhang, S.D.; Zhang, J.B.; Li, X.Z.; Jia, W.H. Genetic variants in the nucleotide excision repair pathway genes and gastric cancer susceptibility in a southern Chinese population. Cancer Manag. Res. 2018, 10, 765–774. [Google Scholar] [CrossRef] [Green Version]
- Woelfelschneider, A.; Popanda, O.; Lilla, C.; Linseisen, J.; Mayer, C.; Celebi, O.; Debus, J.; Bartsch, H.; Chang-Claude, J.; Schmezer, P. A distinct ERCC1 haplotype is associated with mRNA expression levels in prostate cancer patients. Carcinogenesis 2008, 29, 1758–1764. [Google Scholar] [CrossRef]
- Helleday, T.; Petermann, E.; Lundin, C.; Hodgson, B.; Sharma, R.A. DNA repair pathways as targets for cancer therapy. Nature Rev. Cancer 2008, 8, 193–204. [Google Scholar] [CrossRef]
- Ghorbani, M.; Azghandi, M.; Khayami, R.; Baharara, J.; Kerachian, M.A. Association of MTHFR C677T variant genotype with serum folate and Vit B12 in Iranian patients with colorectal cancer or adenomatous polyps. BMC Med. Genom. 2021, 14, 246. [Google Scholar] [CrossRef] [PubMed]
- Belkouz, A.; de Vos-Geelen, J.; Mathot, R.A.A.; Eskens, F.; van Gulik, T.M.; van Oijen, M.G.H.; Punt, C.J.A.; Wilmink, J.W.; Klumpen, H.J. Efficacy and safety of FOLFIRINOX as salvage treatment in advanced biliary tract cancer: An open-label, single arm, phase 2 trial. Br. J. Cancer 2020, 122, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Jordan, P.; Carmo-Fonseca, M. Molecular mechanisms involved in cisplatin cytotoxicity. Cell Mol. Life Sci. 2000, 57, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.P.; Hamilton, T.C.; Schilder, R.J. Platinum resistance: The role of DNA repair pathways. Clin. Cancer Res. 2008, 14, 1291–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matullo, G.; Palli, D.; Peluso, M.; Guarrera, S.; Carturan, S.; Celentano, E.; Krogh, V.; Munnia, A.; Tumino, R.; Polidoro, S.; et al. XRCC1, XRCC3, XPD gene polymorphisms, smoking and (32)P-DNA adducts in a sample of healthy subjects. Carcinogenesis 2001, 22, 1437–1445. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Ouyang, Y.; Chang, J.; Liu, P.; Tian, X.; Yu, J. Pharmacogenetic Association between XRCC1 Polymorphisms and Response to Platinum-Based Chemotherapy in Asian Patients with NSCLC: A Meta-Analysis. Biomed Res. Int. 2020, 2020, 3520764. [Google Scholar] [CrossRef]
- Zipprich, J.; Terry, M.B.; Brandt-Rauf, P.; Freyer, G.A.; Liao, Y.; Agrawal, M.; Gurvich, I.; Senie, R.; Santella, R.M. XRCC1 polymorphisms and breast cancer risk from the New York Site of the Breast Cancer Family Registry: A family-based case-control study. J. Carcinog. 2010, 9, 4. [Google Scholar] [CrossRef]
- Suh, K.W.; Kim, J.H.; Kim, D.Y.; Kim, Y.B.; Lee, C.; Choi, S. Which gene is a dominant predictor of response during FOLFOX chemotherapy for the treatment of metastatic colorectal cancer, the MTHFR or XRCC1 gene? Ann. Surg. Oncol. 2006, 13, 1379–1385. [Google Scholar] [CrossRef]
- Sparreboom, A.; Danesi, R.; Ando, Y.; Chan, J.; Figg, W.D. Pharmacogenomics of ABC transporters and its role in cancer chemotherapy. Drug Resist. Updat. 2003, 6, 71–84. [Google Scholar] [CrossRef]
- de Jong, F.A.; Scott-Horton, T.J.; Kroetz, D.L.; McLeod, H.L.; Friberg, L.E.; Mathijssen, R.H.; Verweij, J.; Marsh, S.; Sparreboom, A. Irinotecan-induced diarrhea: Functional significance of the polymorphic ABCC2 transporter protein. Clin. Pharmacol. Ther. 2007, 81, 42–49. [Google Scholar] [CrossRef]
- Padmanabhan, B.; Tong, K.I.; Ohta, T.; Nakamura, Y.; Scharlock, M.; Ohtsuji, M.; Kang, M.I.; Kobayashi, A.; Yokoyama, S.; Yamamoto, M. Structural basis for defects of Keap1 activity provoked by its point mutations in lung cancer. Mol. Cell 2006, 21, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Misra, V.; Thimmulappa, R.K.; Lee, H.; Ames, S.; Hoque, M.O.; Herman, J.G.; Baylin, S.B.; Sidransky, D.; Gabrielson, E.; et al. Dysfunctional KEAP1-NRF2 interaction in non-small-cell lung cancer. PLoS Med. 2006, 3, e420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taguchi, K.; Motohashi, H.; Yamamoto, M. Molecular mechanisms of the Keap1-Nrf2 pathway in stress response and cancer evolution. Genes Cells 2011, 16, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Esquivel, A.; Chinchilla-Monge, R.; Abbas, J.; Valle, M. C677T and A1298C MTHFR gene polymorphisms and response to fluoropyrimidine-based chemotherapy in Mestizo patients with metastatic colorectal cancer. Pharm. Genom. 2021, 31, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Lan, G.; Lin, L.; Chen, X.; Chen, L.; Chen, X. Correlation Between Methylenetetrahydrofolate Reductase (MTHFR) C677T Polymorphisms and Pemetrexed Chemotherapy Efficacy/Toxicity in Non-Squamous Non-Small Cell Lung Cancer. Med. Sci. Monit. 2017, 23, 5683–5689. [Google Scholar] [CrossRef] [Green Version]
- Henriquez-Hernandez, L.A.; Murias-Rosales, A.; Gonzalez-Hernandez, A.; de Leon, A.C.; Diaz-Chico, N.; Fernandez-Perez, L. Distribution of TYMS, MTHFR, p53 and MDR1 gene polymorphisms in patients with breast cancer treated with neoadjuvant chemotherapy. Cancer Epidemiol. 2010, 34, 634–638. [Google Scholar] [CrossRef]
- Valle, J.; Wasan, H.; Palmer, D.H.; Cunningham, D.; Anthoney, A.; Maraveyas, A.; Madhusudan, S.; Iveson, T.; Hughes, S.; Pereira, S.P.; et al. Cisplatin plus gemcitabine versus gemcitabine for biliary tract cancer. N. Engl. J. Med. 2010, 362, 1273–1281. [Google Scholar] [CrossRef] [Green Version]
- Lamarca, A.; Palmer, D.H.; Wasan, H.S.; Ross, P.J.; Ma, Y.T.; Arora, A.; Falk, S.; Gillmore, R.; Wadsley, J.; Patel, K.; et al. Second-line FOLFOX chemotherapy versus active symptom control for advanced biliary tract cancer (ABC-06): A phase 3, open-label, randomised, controlled trial. Lancet Oncol. 2021, 22, 690–701. [Google Scholar] [CrossRef]
- Olaussen, K.A.; Dunant, A.; Fouret, P.; Brambilla, E.; Andre, F.; Haddad, V.; Taranchon, E.; Filipits, M.; Pirker, R.; Popper, H.H.; et al. DNA repair by ERCC1 in non-small-cell lung cancer and cisplatin-based adjuvant chemotherapy. N. Engl. J. Med. 2006, 355, 983–991. [Google Scholar] [CrossRef]
- Yin, M.; Yan, J.; Martinez-Balibrea, E.; Graziano, F.; Lenz, H.J.; Kim, H.J.; Robert, J.; Im, S.A.; Wang, W.S.; Etienne-Grimaldi, M.C.; et al. ERCC1 and ERCC2 polymorphisms predict clinical outcomes of oxaliplatin-based chemotherapies in gastric and colorectal cancer: A systemic review and meta-analysis. Clin. Cancer Res. 2011, 17, 1632–1640. [Google Scholar] [CrossRef] [Green Version]
- Lau, K.S.; Dennis, J.W. N-Glycans in cancer progression. Glycobiology 2008, 18, 750–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, E.P.; Mandel, U.; Clausen, H.; Gerken, T.A.; Fritz, T.A.; Tabak, L.A. Control of mucin-type O-glycosylation: A classification of the polypeptide GalNAc-transferase gene family. Glycobiology 2012, 22, 736–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, T.C.; Chen, S.T.; Huang, M.C.; Huang, J.; Hsu, C.L.; Juan, H.F.; Lin, H.H.; Chen, C.H. GALNT6 expression enhances aggressive phenotypes of ovarian cancer cells by regulating EGFR activity. Oncotarget 2017, 8, 42588–42601. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Hsu, C.W.; Chen, Y.C.; Chang, M.L.; Liang, K.H.; Lai, M.W.; Lin, C.L.; Chien, R.N.; Lin, K.H.; Yeh, C.T. A GALNT14 rs9679162 genotype-guided therapeutic strategy for advanced hepatocellular carcinoma: Systemic or hepatic arterial infusion chemotherapy. Pharm. J. 2020, 20, 57–68. [Google Scholar] [CrossRef]
- Tummala, P.; Munigala, S.; Eloubeidi, M.A.; Agarwal, B. Patients with obstructive jaundice and biliary stricture ± mass lesion on imaging: Prevalence of malignancy and potential role of EUS-FNA. J. Clin. Gastroenterol. 2013, 47, 532–537. [Google Scholar] [CrossRef]
- Cillo, U.; Fondevila, C.; Donadon, M.; Gringeri, E.; Mocchegiani, F.; Schlitt, H.J.; Ijzermans, J.N.M.; Vivarelli, M.; Zieniewicz, K.; Olde Damink, S.W.M.; et al. Surgery for cholangiocarcinoma. Liver Int. 2019, 39 (Suppl. S1), 143–155. [Google Scholar] [CrossRef] [Green Version]
- Tanisaka, Y.; Mizuide, M.; Fujita, A.; Ogawa, T.; Suzuki, M.; Katsuda, H.; Saito, Y.; Miyaguchi, K.; Tashima, T.; Mashimo, Y.; et al. Diagnostic Process Using Endoscopy for Biliary Strictures: A Narrative Review. J. Clin. Med. 2021, 10, 1048. [Google Scholar] [CrossRef]
- Park, J.S.; Son, J.H.; Park, C.S.; Chang, H.S. Clinical Implications of Single Nucleotide Polymorphisms in Diagnosis of Asthma and its Subtypes. Yonsei Med. J. 2019, 60, 1–9. [Google Scholar] [CrossRef]
- Cardinale, V.; Semeraro, R.; Torrice, A.; Gatto, M.; Napoli, C.; Bragazzi, M.C.; Gentile, R.; Alvaro, D. Intra-hepatic and extra-hepatic cholangiocarcinoma: New insight into epidemiology and risk factors. World J. Gastrointest. Oncol. 2010, 2, 407–416. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author/Year/Ref. | Selection | Comparability | Outcomes | Quality Score |
|---|---|---|---|---|
| Chaiteerakij, 2015 [26] | ★★★★ | ★★ | ★★★ | 9/9 |
| Fingas, 2010 [27] | ★★★★ | ★★ | ★★★ | 9/9 |
| Hoblinger, 2009 [28] | ★★★★ | ★ | ★★★ | 8/9 |
| Honjo, 2005 [29] | ★★★★ | ★ | ★★★ | 8/9 |
| Khunluck, 2014 [30] | ★★★★ | ★★ | ★★★ | 9/9 |
| Ko, 2006 [31] | ★★★★ | ★★ | ★★★ | 9/9 |
| Krawczyk, 2013 [32] | ★★★★ | ★★ | ★★★ | 9/9 |
| Krawczyk, 2011 [19] | ★★★ | ★ | ★★★ | 7/9 |
| Lampropoulou, 2021 [33] | ★★★★ | ★★ | ★★★ | 9/9 |
| Melum, 2007 [34] | ★★★★ | ★ | ★★★ | 8/9 |
| Meng, 2014 [35] | ★★★★ | ★ | ★★ | 7/9 |
| Mihalache, 2012 [36] | ★★★★ | ★★ | ★★★ | 9/9 |
| Mihalache, 2011 [37] | ★★★★ | ★★ | ★★★ | 9/9 |
| Prawan, 2005 [38] | ★★★★ | ★ | ★★★ | 8/9 |
| Songserm, 2011 [39] | ★★★★ | ★★ | ★★★ | 9/9 |
| Sun, 2018 [40] | ★★★★ | ★ | ★★★ | 8/9 |
| Surapaitoon, 2017 [41] | ★★★★ | ★★ | ★★★ | 9/9 |
| Wadsworth, 2019 [21] | ★★★★ | ★★ | ★★★ | 9/9 |
| You, 2012 [42] | ★★★★ | ★★ | ★★★ | 9/9 |
| Zhao, 2014 [43] | ★★★★ | ★★ | ★★★ | 9/9 |
| Zeng, 2013 [22] | ★★★★ | ★★ | ★★★★ | 9/9 |
| Songserm, 2014 [44] | ★★★★ | ★★ | ★★★★ | 9/9 |
| Liu et al., 2022 [45] | ★★★★ | ★★ | ★★★★ | 9/9 |
| Hsing et al., 2008 [25] | ★★★★ | ★★ | ★★★ | 8/9 |
| Gene | SNP | Factor | Method | Sample | Case (%) | Control (%) | OR (95% CI) | p Value | Susceptibility | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| PTGS2 | rs689466 | C | TaqMan | Blood | 163 (22) | 252 (17) | 1.36 (1.10–1.69) | 0.005 | Increase | Chaiteerakij et al., 2015 [26] |
| rs2143417 | T | TaqMan | Blood | 148 (20) | 207 (14) | 1.52 (1.21–1.91) | 0.0003 | Increase | ||
| IL6 | rs1800795 | GC | PCR-RFLP | Blood | 45 (40.9) | 67 (33.5) | 2.35 (1.31–4.21) | Increase | Surapaitoon et al., 2017 [41] | |
| GC | PCR-RFLP | Blood | 45 (40.9) | 59 (29.5) | 2.95 (1.64–5.31) | Increase | ||||
| C | PCR-RFLP | Blood | 111 (50.5) | 93 (23.3) | 3.36 (2.32–4.85) | Increase | ||||
| IFNG | rs2430561 | AA | PCR-RFLP | Blood | 54 (49.1) | 65 (32.5) | 2.20 (1.13–4.20) | Increase | ||
| TNF | rs1800629 | A | PCR-RFLP | Blood | 173 (78.6) | 284 (71.0) | 1.50 (1.00–2.26) | Increase | ||
| A | PCR-RFLP | Blood | 173 (78.6) | 278 (69.5) | 1.61 (1.08–2.43) | Increase |
| Gene | SNP | Factor | Method | Sample | Case (%) | Control (%) | OR (95% CI) | p Value | Susceptibility | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| ERCC1 | rs3212986(ECCA) | AC + AA | PCR-RFLP | Blood | 68 (53.5) | 59 (40.7) | 1.68 (1.04–2.72) | 0.03 | Increase | Sun et al., 2018 [40] |
| MTHFR | rs1801131 | CC | PCR-HRM | Tissues | 62 (35.4) | 72 (20.6) | 2.00 (1.14–3.48) | Increase | Songserm et al., 2011 [39] | |
| NAT1 | * 11 | PCR-RFLP | Blood | 1 (0.2) | 11 (2.4) | 0.10 (0.00–0.58) | 0.005 | Decrease | Prawan et al., 2005 [38] | |
| NAT2 | * 13 | PCR-RFLP | Blood | 8 (1.9) | 24 (5.2) | 0.35 (0.16–0.77) | 0.008 | Decrease | ||
| rs1799930 | * 6B | PCR-RFLP | Blood | 6 (1.4) | 22 (4.7) | 0.28 (0.12–0.69) | 0.004 | Decrease | ||
| rs1799931 | * 7A | PCR-RFLP | Blood | 9 (2.1) | 28 (6.0) | 0.33 (0.16–0.70) | 0.003 | Decrease | ||
| MUTYH | rs3219476 | TG | PCR-RFLP | Blood | 20 (30.9) | 58 (58.0) | 0.36 (0.17–0.76) | 0.006 | Decrease | You et al., 2013 [42] |
| rs3219472 | AA | PCR-RFLP | Blood | 12 (20.3) | 7 (7.0) | 2.82 (0.99–8.00) | 0.047 | Increase | ||
| ABCC2 | rs3740066 | T | TaqMan | Blood | 47 (39.2) | 38 (26.0) | 1.83 (1.09–3.08) | 0.022 | Increase | Höblinger et al., 2009 [28] |
| Gene | SNP | Factor | Method | Sample | Case (%) | Control (%) | OR (95% CI) | p Value | Susceptibility | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| SERPINA1 | rs28929474 (Z) | T | TaqMan | Blood | 15 (4.0) | 12 (2.0) | 2.46 (1.14–5.32) | 0.036 | Increase | Mihalache et al., 2015 [37] |
| GSTO1 | rs4925 | D140 | PCR | Tissues | 18 (30.0) | 4 (6.7) | 8.50 (2.07–37.85) | <0.05 | Increase | Marahatta et al., 2005 |
| MST1 | rs3197999 | GG | TaqMan | Blood | 115 (52.0) | 194 (55.0) | 1.97 (1.09–3.36) | 0.023 | Increase | Krawczyk et al., 2013 [32] |
| AA (ECCA) | TaqMan | Blood | 22 (12.0) | 24 (6.0) | 2.04 (1.09–3.84) | 0.024 | Increase | |||
| HOTAIR | rs4759314 | GG | PCR-RFLP | Blood | 7 (5.7) | 1 (0.6) | 12.31 (1.48–101.87) | 0.005 | Increase | Lampropoulou et al., 2021 [33] |
| AG | PCR-RFLP | Blood | 32 (26.3) | 18 (10.9) | 3.13 (1.65–5.91) | 0.0004 | Increase | |||
| KLRK1 | rs11053781 | G/A | TaqMan | Blood | 32 (66.0) | 184 (50.0) | 1.95 (1.23–3.07) | 0.0038 | Increase | Melum et al., 2007 [34] |
| rs2617167 | A/G | TaqMan | Blood | 19 (39.0) | 85 (23.0) | 2.20 (1.40–3.44) | 0.00046 | Increase | ||
| GAB1 | rs3805246 | AA + AG VS GG | TaqMan | Tissues | 154 (68.4) | 71 (31.6) | 2.15 (1.28–3.71) | 0.016 | Increase | Lingqin et al., 2014 [35] |
| AA VS (AG + GG) | TaqMan | Tissues | 35 (15.5) | 190 (84.5) | 1.98 (1.21–2.84) | 0.012 | Increase | |||
| EGFR | rs2017000 | AA | TaqMan | Blood | 26 (11.6) | 1.92 (1.14–2.59) | 0.002 | Increase | Lingqin et al., 2015 [70] | |
| APOBEC3B | rs2267401 | TG | Taqman | Blood | *** | *** | 0.51 (0.36–0.72) | 0.00016 | Decrease | Liu et al., 2022 [45] |
| TG + GG | Taqman | Blood | *** | *** | 0.69 (0.51–0.94) | 0.0189 | Decrease | |||
| APOBEC3A | rs12157810 | CC | Taqman | Blood | *** | *** | 0.44 (0.33–0.60) | <0.001 | Decrease | |
| AC + CC | Taqman | Blood | *** | *** | 0.80 (0.66–0.97) | 0.025 | Decrease |
| Gene | SNP | Factor | Method | Samples | Case (%) | Control (%) | p Value | Prognosis (OS) | Reference |
|---|---|---|---|---|---|---|---|---|---|
| GNB3 | rs5443 | CC VS (CT + TT) | PCR | Blood | 17 (42.5) | 23 (57.5) | <0.05 | Improved | Fingas et al., 2009 [27] |
| NFE2L2 | rs6726395 | GG VS (AA/GG) | TaqMan | Blood | 34 (38.6) | 54 (61.4) | 0.006 | Improved | Khunluck et al., 2014 [30] |
| GALNT14 | rs9679162 | TT VS (TG + GG) | Sanger sequencing | Tissues | 35 (31.3) | 77 (38.7) | 0.023 | Reduced | Liang et al., 2017 [95] |
| EGFR | rs2017000 | AA VS (GG + GA) | TaqMan | Blood | 21 (10.2) | 105 (88.8) | 0.021 | Reduced | Lingqin et al., 2015 [70] |
| XRCC1 | rs25487 | Arg/Arg VS(Arg/Gln + Gln/Gln) | TaqMan | Blood | 17 (51.5) | 16 (48.5) | 0.013 | Reduced | Pacetti et al., [98] |
| EZH2 | rs887569 | TT VS (CC + CT) | TaqMan | Blood | *** | *** | 0.036 | Improved | Paolicchi et al., 2013 [99] |
| GNAS1 | rs7121 | TT VS (CT + CC) | PCR | Tissues | 7 (14.0) | 40 (85.1) | <0.008 | Reduced | Schmitz et al., 2007 [100] |
| CXCR1 | rs2234671 | CC VS CG | PCR-RFLP | Tissues | 92 (83.6) | 18 (16.4) | 0.002 | Improved | Lurje et al. [101] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.; Heij, L.R.; Liu, D.; Dahl, E.; LANG, S.A.; Ulmer, T.F.; LUEDDE, T.; Neumann, U.P.; Bednarsch, J. The Role of Single-Nucleotide Polymorphisms in Cholangiocarcinoma: A Systematic Review. Cancers 2022, 14, 5969. https://doi.org/10.3390/cancers14235969
Wang G, Heij LR, Liu D, Dahl E, LANG SA, Ulmer TF, LUEDDE T, Neumann UP, Bednarsch J. The Role of Single-Nucleotide Polymorphisms in Cholangiocarcinoma: A Systematic Review. Cancers. 2022; 14(23):5969. https://doi.org/10.3390/cancers14235969
Chicago/Turabian StyleWang, Guanwu, Lara Rosaline Heij, Dong Liu, Edgar Dahl, Sven Arke LANG, Tom Florian Ulmer, Tom LUEDDE, Ulf Peter Neumann, and Jan Bednarsch. 2022. "The Role of Single-Nucleotide Polymorphisms in Cholangiocarcinoma: A Systematic Review" Cancers 14, no. 23: 5969. https://doi.org/10.3390/cancers14235969
APA StyleWang, G., Heij, L. R., Liu, D., Dahl, E., LANG, S. A., Ulmer, T. F., LUEDDE, T., Neumann, U. P., & Bednarsch, J. (2022). The Role of Single-Nucleotide Polymorphisms in Cholangiocarcinoma: A Systematic Review. Cancers, 14(23), 5969. https://doi.org/10.3390/cancers14235969