
The Intricate Interplay between the ZNF217 Oncogene and Epigenetic Processes Shapes Tumor Progression
 ,
, 
 and
and 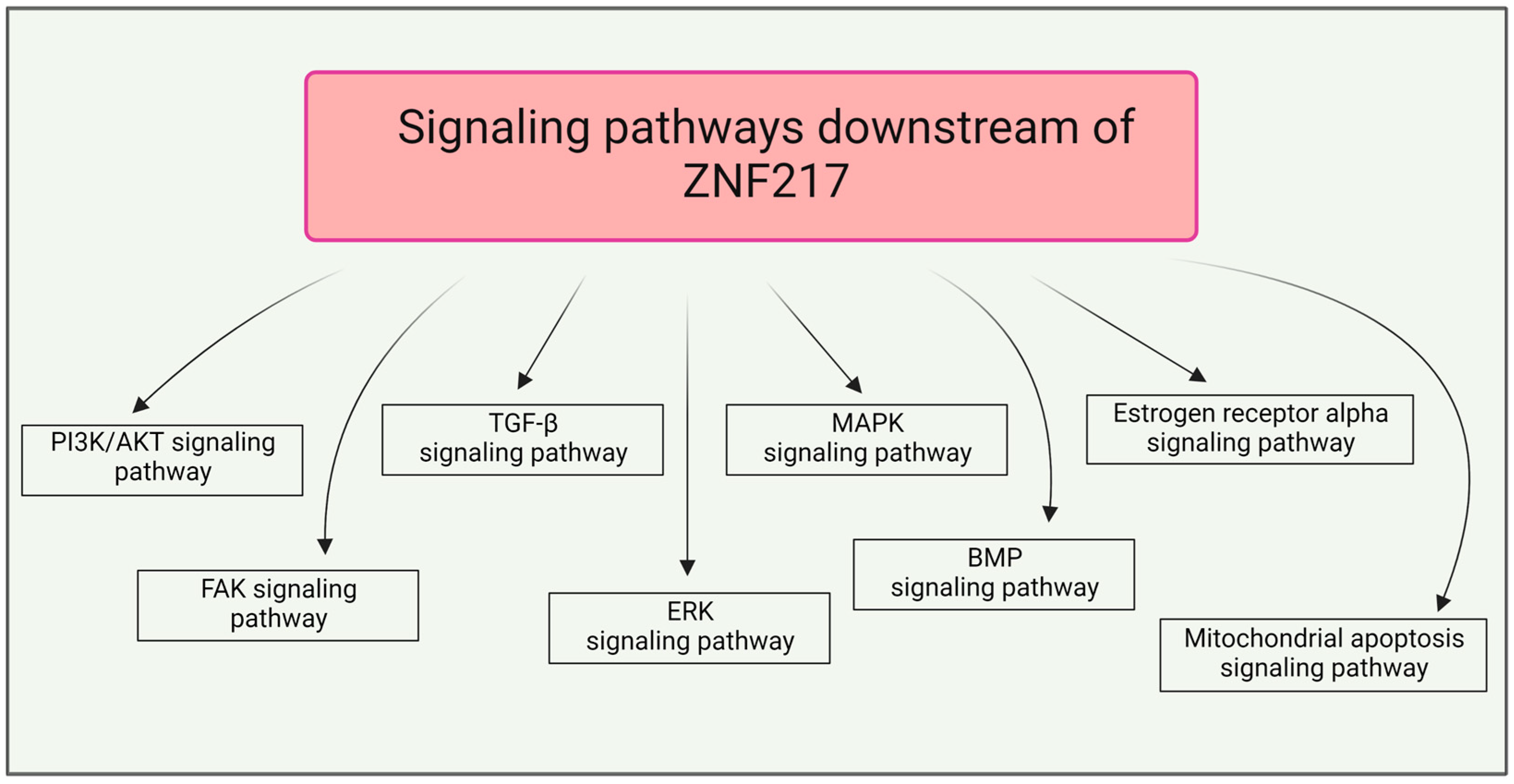{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. DNA Methylation Status at the ZNF217 Locus Inversely Correlates with ZNF217 Expression Levels
3. ZNF217 Fine Tunes DNA Methylation Status of the Tumor Suppressor P15INK4B/CDKN2B Gene
4. ZNF217/Zfp217 Regulates the Epitranscriptome to Promote Cell Reprograming and Pluripotency
5. Noncoding RNA Networks Target ZNF217 mRNA, Control ZNF217 Expression Levels and Govern ZNF217-Driven Functions
5.1. ZNF217-Driven EMT Could Be Epigenetically Regulated by Specific miRNAs and Intricate lncRNA Networks
5.2. Trastuzumab Resistance Is Mediated by a lncRNA-ATB/miR-200c/ZNF217/TGF-β Axis
5.3. Tumor Progression Is Accelerated by Oncogenic LncRNAs or Oncogenic circRNAs Sponging miRNA and Upregulating ZNF217 Expression
6. ZNF217-Associated Epigenetic Events Interfere with BCa Risk Factors
6.1. Estrogen Exposure
6.2. Obesity and Adipogenesis
6.3. Stroma Stiffness
7. NcRNAs Networks Targeting ZNF217 Are Also Involved in Programing Cancer Cells to Metastasize to Bone
8. Future Directions
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cohen, P.A.; Donini, C.F.; Nguyen, N.T.; Lincet, H.; Vendrell, J.A. The dark side of ZNF217, a key regulator of tumorigenesis with powerful biomarker value. Oncotarget 2015, 6, 41566–41581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wu, H.; Wang, Q.; Xu, S. ZNF217: The cerberus who fails to guard the gateway to lethal malignancy. Am. J. Cancer Res. 2021, 11, 3378–3405. [Google Scholar] [PubMed]
- Collins, C.; Rommens, J.M.; Kowbel, D.; Godfrey, T.; Tanner, M.; Hwang, S.I.; Polikoff, D.; Nonet, G.; Cochran, J.; Myambo, K.; et al. Positional cloning of ZNF217 and NABC1: Genes amplified at 20q13.2 and overexpressed in breast carcinoma. Proc. Natl. Acad. Sci. USA 1998, 95, 8703–8708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowger, J.J.; Zhao, Q.; Isovic, M.; Torchia, J. Biochemical characterization of the zinc-finger protein 217 transcriptional repressor complex: Identification of a ZNF217 consensus recognition sequence. Oncogene 2007, 26, 3378–3386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krig, S.R.; Jin, V.X.; Bieda, M.C.; O’Geen, H.; Yaswen, P.; Green, R.; Farnham, P.J. Identification of genes directly regulated by the oncogene ZNF217 using chromatin immunoprecipitation (ChIP)-chip assays. J. Biol. Chem. 2007, 282, 9703–9712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunez, N.; Clifton, M.M.K.; Funnell, A.P.W.; Artuz, C.; Hallal, S.; Quinlan, K.G.R.; Font, J.; Vandevenne, M.; Setiyaputra, S.; Pearson, R.C.M.; et al. The multi-zinc finger protein ZNF217 contacts DNA through a two-finger domain. J. Biol. Chem. 2011, 286, 38190–38201. [Google Scholar] [CrossRef] [Green Version]
- Quinlan, K.G.; Verger, A.; Yaswen, P.; Crossley, M. Amplification of zinc finger gene 217 (ZNF217) and cancer: When good fingers go bad. Biochim. Biophys. Acta 2007, 1775, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Thillainadesan, G.; Isovic, M.; Loney, E.; Andrews, J.; Tini, M.; Torchia, J. Genome analysis identifies the p15ink4b tumor suppressor as a direct target of the ZNF217/CoREST complex. Mol. Cell. Biol. 2008, 28, 6066–6077. [Google Scholar] [CrossRef] [Green Version]
- Vendrell, J.A.; Thollet, A.; Nguyen, N.T.; Ghayad, S.E.; Vinot, S.; Bieche, I.; Grisard, E.; Josserand, V.; Coll, J.L.; Roux, P.; et al. ZNF217 is a marker of poor prognosis in breast cancer that drives epithelial-mesenchymal transition and invasion. Cancer Res. 2012, 72, 3593–3606. [Google Scholar] [CrossRef] [Green Version]
- Littlepage, L.E.; Adler, A.S.; Kouros-Mehr, H.; Huang, G.; Chou, J.; Krig, S.R.; Griffith, O.L.; Korkola, J.E.; Qu, K.; Lawson, D.A.; et al. The transcription factor ZNF217 is a prognostic biomarker and therapeutic target during breast cancer progression. Cancer Discov. 2012, 2, 638–651. [Google Scholar] [CrossRef]
- Krig, S.R.; Miller, J.K.; Frietze, S.; Beckett, L.A.; Neve, R.M.; Farnham, P.J.; Yaswen, P.I.; Sweeney, C.A. ZNF217, a candidate breast cancer oncogene amplified at 20q13, regulates expression of the ErbB3 receptor tyrosine kinase in breast cancer cells. Oncogene 2010, 29, 5500–5510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilo, F.; Zhang, F.; Sancho, A.; Fidalgo, M.; Di Cecilia, S.; Vashisht, A.; Lee, D.F.; Chen, C.H.; Rengasamy, M.; Andino, B.; et al. Coordination of m(6)A mRNA Methylation and Gene Transcription by ZFP217 Regulates Pluripotency and Reprogramming. Cell Stem Cell 2015, 17, 689–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, T.; Yang, Y.; Wei, H.; Xie, X.; Lu, J.; Zeng, Q.; Peng, J.; Zhou, Y.; Jiang, S.; Peng, J. Zfp217 mediates m6A mRNA methylation to orchestrate transcriptional and post-transcriptional regulation to promote adipogenic differentiation. Nucleic Acids Res. 2019, 47, 6130–6144. [Google Scholar] [CrossRef] [Green Version]
- Ginzinger, D.G.; Godfrey, T.E.; Nigro, J.; Moore, D.H., 2nd; Suzuki, S.; Pallavicini, M.G.; Gray, J.W.; Jensen, R.H. Measurement of DNA copy number at microsatellite loci using quantitative PCR analysis. Cancer Res. 2000, 60, 5405–5409. [Google Scholar] [PubMed]
- Peiro, G.; Diebold, J.; Lohrs, U. CAS (cellular apoptosis susceptibility) gene expression in ovarian carcinoma: Correlation with 20q13.2 copy number and cyclin D1, p53, and Rb protein expression. Am. J. Clin. Pathol. 2002, 118, 922–929. [Google Scholar] [CrossRef] [Green Version]
- Rooney, P.H.; Boonsong, A.; McFadyen, M.C.; McLeod, H.L.; Cassidy, J.; Curran, S.; Murray, G.I. The candidate oncogene ZNF217 is frequently amplified in colon cancer. J. Pathol. 2004, 204, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Collins, C.; Volik, S.; Kowbel, D.; Ginzinger, D.; Ylstra, B.; Cloutier, T.; Hawkins, T.; Predki, P.; Martin, C.; Wernick, M.; et al. Comprehensive genome sequence analysis of a breast cancer amplicon. Genome Res. 2001, 11, 1034–1042. [Google Scholar] [CrossRef]
- Rahman, M.T.; Nakayama, K.; Rahman, M.; Nakayama, N.; Ishikawa, M.; Katagiri, A.; Iida, K.; Nakayama, S.; Otsuki, Y.; Shih Ie, M.; et al. Prognostic and therapeutic impact of the chromosome 20q13.2 ZNF217 locus amplification in ovarian clear cell carcinoma. Cancer 2012, 118, 2846–2857. [Google Scholar] [CrossRef] [Green Version]
- Ginestier, C.; Cervera, N.; Finetti, P.; Esteyries, S.; Esterni, B.; Adelaide, J.; Xerri, L.; Viens, P.; Jacquemier, J.; Charafe-Jauffret, E.; et al. Prognosis and gene expression profiling of 20q13-amplified breast cancers. Clin. Cancer Res. 2006, 12, 4533–4544. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.; Krig, S.; Kowbel, D.; Xu, H.; Hyun, B.; Volik, S.; Feuerstein, B.; Mills, G.B.; Stokoe, D.; Yaswen, P.; et al. ZNF217 suppresses cell death associated with chemotherapy and telomere dysfunction. Hum. Mol. Genet. 2005, 14, 3219–3225. [Google Scholar] [CrossRef]
- Jiang, X.; Zhang, C.; Qi, S.; Guo, S.; Chen, Y.; Du, E.; Zhang, H.; Wang, X.; Liu, R.; Qiao, B.; et al. Elevated expression of ZNF217 promotes prostate cancer growth by restraining ferroportin-conducted iron egress. Oncotarget 2016, 7, 84893–84906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, X.; Lv, S.; Qiao, Y.; Wang, F.; Miao, L. Interferon regulatory factor 5induced upregulation of zincfinger protein 217 promotes pancreatic carcinoma progression. Mol. Med. Rep. 2022, 25, 189. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, X.; Song, L.; Su, G.; Di, A.; Bai, C.; Wei, Z.; Li, G. Melatonin restores the pluripotency of long-term-cultured embryonic stem cells through melatonin receptor-dependent m6A RNA regulation. J. Pineal Res. 2020, 69, e12669. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.G.; Yan, M.; Xue, X.Y.; Zhang, X.; Ren, H.G.; Guo, G.; Wang, P.; Zhang, W.; Huo, J.L. Overexpression of ZNF217 in glioblastoma contributes to the maintenance of glioma stem cells regulated by hypoxia-inducible factors. Lab. Investig. 2011, 91, 1068–1078. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhi, W.I.; Lu, H.; Samanta, D.; Chen, I.; Gabrielson, E.; Semenza, G.L. Hypoxia-inducible factors regulate pluripotency factor expression by ZNF217- and ALKBH5-mediated modulation of RNA methylation in breast cancer cells. Oncotarget 2016, 7, 64527–64542. [Google Scholar] [CrossRef] [Green Version]
- Herceg, Z.; Ushijima, T. Introduction: Epigenetics and cancer. Adv. Genet. 2010, 70, 1–23. [Google Scholar] [CrossRef]
- Lee, J.E.; Kim, M.Y. Cancer epigenetics: Past, present and future. Semin. Cancer Biol. 2022, 83, 4–14. [Google Scholar] [CrossRef]
- Thillainadesan, G.; Chitilian, J.M.; Isovic, M.; Ablack, J.N.; Mymryk, J.S.; Tini, M.; Torchia, J. TGF-beta-dependent active demethylation and expression of the p15ink4b tumor suppressor are impaired by the ZNF217/CoREST complex. Mol. Cell 2012, 46, 636–649. [Google Scholar] [CrossRef] [Green Version]
- Banck, M.S.; Li, S.; Nishio, H.; Wang, C.; Beutler, A.S.; Walsh, M.J. The ZNF217 oncogene is a candidate organizer of repressive histone modifiers. Epigenetics 2009, 4, 100–106. [Google Scholar] [CrossRef] [Green Version]
- Kuppuswamy, M.; Vijayalingam, S.; Zhao, L.J.; Zhou, Y.; Subramanian, T.; Ryerse, J.; Chinnadurai, G. Role of the PLDLS-Binding Cleft Region of CtBP1 in Recruitment of Core and Auxiliary Components of the Corepressor Complex. Mol. Cell. Biol. 2008, 28, 269–281. [Google Scholar] [CrossRef]
- Quinlan, K.G.; Nardini, M.; Verger, A.; Francescato, P.; Yaswen, P.; Corda, D.; Bolognesi, M.; Crossley, M. Specific recognition of ZNF217 and other zinc finger proteins at a surface groove of C-terminal binding proteins. Mol. Cell. Biol. 2006, 26, 8159–8172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plass, C.; Pfister, S.M.; Lindroth, A.M.; Bogatyrova, O.; Claus, R.; Lichter, P. Mutations in regulators of the epigenome and their connections to global chromatin patterns in cancer. Nat. Rev. Genet. 2013, 14, 765–780. [Google Scholar] [CrossRef] [PubMed]
- Gama-Sosa, M.A.; Slagel, V.A.; Trewyn, R.W.; Oxenhandler, R.; Kuo, K.C.; Gehrke, C.W.; Ehrlich, M. The 5-methylcytosine content of DNA from human tumors. Nucleic Acids Res. 1983, 11, 6883–6894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feinberg, A.P.; Vogelstein, B. Hypomethylation distinguishes genes of some human cancers from their normal counterparts. Nature 1983, 301, 89–92. [Google Scholar] [CrossRef]
- Ferreira, H.J.; Esteller, M. CpG Islands in Cancer: Heads, Tails, and Sides. Methods Mol. Biol. 2018, 1766, 49–80. [Google Scholar] [CrossRef]
- Leu, Y.W.; Yan, P.S.; Fan, M.; Jin, V.X.; Liu, J.C.; Curran, E.M.; Welshons, W.V.; Wei, S.H.; Davuluri, R.V.; Plass, C.; et al. Loss of estrogen receptor signaling triggers epigenetic silencing of downstream targets in breast cancer. Cancer Res. 2004, 64, 8184–8192. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.T.; Vendrell, J.A.; Poulard, C.; Gyorffy, B.; Goddard-Leon, S.; Bieche, I.; Corbo, L.; Le Romancer, M.; Bachelot, T.; Treilleux, I.; et al. A functional interplay between ZNF217 and estrogen receptor alpha exists in luminal breast cancers. Mol. Oncol. 2014, 8, 1441–1457. [Google Scholar] [CrossRef]
- Bellanger, A.; Le, D.T.; Vendrell, J.; Wierinckx, A.; Pongor, L.S.; Solassol, J.; Lachuer, J.; Clezardin, P.; Gyorffy, B.; Cohen, P.A. Exploring the Significance of the Exon 4-Skipping Isoform of the ZNF217 Oncogene in Breast Cancer. Front. Oncol. 2021, 11, 647269. [Google Scholar] [CrossRef]
- Widschwendter, M.; Apostolidou, S.; Raum, E.; Rothenbacher, D.; Fiegl, H.; Menon, U.; Stegmaier, C.; Jacobs, I.J.; Brenner, H. Epigenotyping in peripheral blood cell DNA and breast cancer risk: A proof of principle study. PLoS ONE 2008, 3, e2656. [Google Scholar] [CrossRef]
- Khakpour, G.; Pooladi, A.; Izadi, P.; Noruzinia, M.; Tavakkoly Bazzaz, J. DNA methylation as a promising landscape: A simple blood test for breast cancer prediction. Tumour Biol. 2015, 36, 4905–4912. [Google Scholar] [CrossRef]
- An, G.; Feng, L.; Hou, L.; Li, X.; Bai, J.; He, L.; Gu, S.; Zhao, X. A bioinformatics analysis of zinc finger protein family reveals potential oncogenic biomarkers in breast cancer. Gene 2022, 828, 146471. [Google Scholar] [CrossRef] [PubMed]
- Etcheverry, A.; Aubry, M.; de Tayrac, M.; Vauleon, E.; Boniface, R.; Guenot, F.; Saikali, S.; Hamlat, A.; Riffaud, L.; Menei, P.; et al. DNA methylation in glioblastoma: Impact on gene expression and clinical outcome. BMC Genom. 2010, 11, 701. [Google Scholar] [CrossRef] [Green Version]
- Ness, C.; Katta, K.; Garred, O.; Kumar, T.; Olstad, O.K.; Petrovski, G.; Moe, M.C.; Noer, A. Integrated differential DNA methylation and gene expression of formalin-fixed paraffin-embedded uveal melanoma specimens identifies genes associated with early metastasis and poor prognosis. Exp. Eye Res. 2021, 203, 108426. [Google Scholar] [CrossRef] [PubMed]
- Renner, M.; Wolf, T.; Meyer, H.; Hartmann, W.; Penzel, R.; Ulrich, A.; Lehner, B.; Hovestadt, V.; Czwan, E.; Egerer, G.; et al. Integrative DNA methylation and gene expression analysis in high-grade soft tissue sarcomas. Genome Biol. 2013, 14, r137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.W.; Kang, B.H.; Jang, H.; Kwak, S.; Shin, J.; Kim, H.; Lee, S.E.; Lee, S.M.; Lee, J.H.; Kim, J.H.; et al. Ctbp2 Modulates NuRD-Mediated Deacetylation of H3K27 and Facilitates PRC2-Mediated H3K27me3 in Active Embryonic Stem Cell Genes During Exit from Pluripotency. Stem Cells 2015, 33, 2442–2455. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.Y.; Sharpless, N.E. The regulation of INK4/ARF in cancer and aging. Cell 2006, 127, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, I.; Much, C.; Di Giacomo, M.; Azzi, C.; Morgan, M.; Moreira, P.N.; Monahan, J.; Carrieri, C.; Enright, A.J.; O’Carroll, D. The RNA m(6)A Reader YTHDF2 Is Essential for the Post-transcriptional Regulation of the Maternal Transcriptome and Oocyte Competence. Mol. Cell 2017, 67, 1059–1067 e1054. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Li, H.; Wu, A.; Peng, Y.; Shu, G.; Yin, G. Functions of N6-methyladenosine and its role in cancer. Mol. Cancer 2019, 18, 176. [Google Scholar] [CrossRef] [Green Version]
- Zaccara, S.; Ries, R.J.; Jaffrey, S.R. Reading, writing and erasing mRNA methylation. Nat. Rev. Mol. Cell Biol. 2019, 20, 608–624. [Google Scholar] [CrossRef]
- Lan, Q.; Liu, P.Y.; Haase, J.; Bell, J.L.; Huttelmaier, S.; Liu, T. The Critical Role of RNA m(6)A Methylation in Cancer. Cancer Res. 2019, 79, 1285–1292. [Google Scholar] [CrossRef]
- Liu, Q.; Zhao, Y.; Wu, R.; Jiang, Q.; Cai, M.; Bi, Z.; Liu, Y.; Yao, Y.; Feng, J.; Wang, Y.; et al. ZFP217 regulates adipogenesis by controlling mitotic clonal expansion in a METTL3-m(6)A dependent manner. RNA Biol. 2019, 16, 1785–1793. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, N.; Du, Z.; Ling, Z.; Zhang, P.; Yang, J.; Khaleel, M.; Khoury, A.N.; Li, J.; Li, S.; et al. Bioinformatics analysis integrating metabolomics of m(6)A RNA microarray in intervertebral disc degeneration. Epigenomics 2020, 12, 1419–1441. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhong, L.; Zou, Z.; Liang, G.; Tang, Z.; Li, K.; Tan, S.; Huang, Y.; Zhu, X. Clinical and Prognostic Pan-Cancer Analysis of N6-Methyladenosine Regulators in Two Types of Hematological Malignancies: A Retrospective Study Based on TCGA and GTEx Databases. Front. Oncol. 2021, 11, 623170. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.F.; Walsh, M.J.; Aguilo, F. ZNF217/ZFP217 Meets Chromatin and RNA. Trends Biochem. Sci. 2016, 41, 986–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oskarsson, T.; Batlle, E.; Massague, J. Metastatic stem cells: Sources, niches, and vital pathways. Cell Stem Cell 2014, 14, 306–321. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Lewis, M.T.; Huang, J.; Gutierrez, C.; Osborne, C.K.; Wu, M.F.; Hilsenbeck, S.G.; Pavlick, A.; Zhang, X.; Chamness, G.C.; et al. Intrinsic resistance of tumorigenic breast cancer cells to chemotherapy. J. Natl. Cancer Inst. 2008, 100, 672–679. [Google Scholar] [CrossRef]
- Seymour, T.; Twigger, A.J.; Kakulas, F. Pluripotency Genes and Their Functions in the Normal and Aberrant Breast and Brain. Int. J. Mol. Sci. 2015, 16, 27288–27301. [Google Scholar] [CrossRef] [Green Version]
- Kwak, S.; Kim, T.W.; Kang, B.H.; Kim, J.H.; Lee, J.S.; Lee, H.T.; Hwang, I.Y.; Shin, J.; Lee, J.H.; Cho, E.J.; et al. Zinc finger proteins orchestrate active gene silencing during embryonic stem cell differentiation. Nucleic Acids Res. 2018, 46, 6592–6607. [Google Scholar] [CrossRef]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Grillone, K.; Riillo, C.; Scionti, F.; Rocca, R.; Tradigo, G.; Guzzi, P.H.; Alcaro, S.; Di Martino, M.T.; Tagliaferri, P.; Tassone, P. Non-coding RNAs in cancer: Platforms and strategies for investigating the genomic “dark matter”. J. Exp. Clin. Cancer Res. 2020, 39, 117. [Google Scholar] [CrossRef]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Warnefors, M.; Liechti, A.; Halbert, J.; Valloton, D.; Kaessmann, H. Conserved microRNA editing in mammalian evolution, development and disease. Genome Biol. 2014, 15, R83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebert, M.S.; Sharp, P.A. Roles for microRNAs in conferring robustness to biological processes. Cell 2012, 149, 515–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grixti, J.M.; Ayers, D. Long noncoding RNAs and their link to cancer. Noncoding RNA Res. 2020, 5, 77–82. [Google Scholar] [CrossRef]
- Huarte, M. The emerging role of lncRNAs in cancer. Nat. Med. 2015, 21, 1253–1261. [Google Scholar] [CrossRef]
- Pang, B.; Wang, Q.; Ning, S.; Wu, J.; Zhang, X.; Chen, Y.; Xu, S. Landscape of tumor suppressor long noncoding RNAs in breast cancer. J. Exp. Clin. Cancer Res. 2019, 38, 79. [Google Scholar] [CrossRef] [Green Version]
- Patop, I.L.; Wust, S.; Kadener, S. Past, present, and future of circRNAs. EMBO J. 2019, 38, e100836. [Google Scholar] [CrossRef]
- Huang, A.; Zheng, H.; Wu, Z.; Chen, M.; Huang, Y. Circular RNA-protein interactions: Functions, mechanisms, and identification. Theranostics 2020, 10, 3503–3517. [Google Scholar] [CrossRef]
- Dweep, H.; Gretz, N. miRWalk2.0: A comprehensive atlas of microRNA-target interactions. Nat. Methods 2015, 12, 697. [Google Scholar] [CrossRef]
- Xiang, H.; Zhong, Z.X.; Peng, Y.D.; Jiang, S.W. The Emerging Role of Zfp217 in Adipogenesis. Int. J. Mol. Sci. 2017, 18, 1367. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.M.; Zhang, J.; Ma, Y.; Yuan, Y.J.; Yu, H.; Wang, J.; Cao, X.C.; Zhu, L.; Wang, P. MicroRNA-135 inhibits initiation of epithelial-mesenchymal transition in breast cancer by targeting ZNF217 and promoting m6A modification of NANOG. Oncogene 2022, 41, 1742–1751. [Google Scholar] [CrossRef] [PubMed]
- Burk, U.; Schubert, J.; Wellner, U.; Schmalhofer, O.; Vincan, E.; Spaderna, S.; Brabletz, T. A reciprocal repression between ZEB1 and members of the miR-200 family promotes EMT and invasion in cancer cells. EMBO Rep. 2008, 9, 582–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, W.D.; Ye, X.M.; Zhang, M.Y.; Zhu, H.Y.; Xi, W.J.; Huang, X.; Zhao, J.; Gu, B.; Zheng, G.X.; Yang, A.G.; et al. MiR-200c suppresses TGF-beta signaling and counteracts trastuzumab resistance and metastasis by targeting ZNF217 and ZEB1 in breast cancer. Int. J. Cancer 2014, 135, 1356–1368. [Google Scholar] [CrossRef]
- Gregory, P.A.; Bert, A.G.; Paterson, E.L.; Barry, S.C.; Tsykin, A.; Farshid, G.; Vadas, M.A.; Khew-Goodall, Y.; Goodall, G.J. The miR-200 family and miR-205 regulate epithelial to mesenchymal transition by targeting ZEB1 and SIP1. Nat. Cell Biol. 2008, 10, 593–601. [Google Scholar] [CrossRef]
- Zhu, H.Y.; Bai, W.D.; Li, C.; Zheng, Z.; Guan, H.; Liu, J.Q.; Yang, X.K.; Han, S.C.; Gao, J.X.; Wang, H.T.; et al. Knockdown of lncRNA-ATB suppresses autocrine secretion of TGF-beta2 by targeting ZNF217 via miR-200c in keloid fibroblasts. Sci. Rep. 2016, 6, 24728. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.H.; Yang, F.; Wang, F.; Ma, J.Z.; Guo, Y.J.; Tao, Q.F.; Liu, F.; Pan, W.; Wang, T.T.; Zhou, C.C.; et al. A long noncoding RNA activated by TGF-beta promotes the invasion-metastasis cascade in hepatocellular carcinoma. Cancer Cell 2014, 25, 666–681. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Yi, X.M.; Tang, C.P.; Ge, J.P.; Zhang, Z.Y.; Zhou, W.Q. Long non-coding RNA ATB promotes growth and epithelial-mesenchymal transition and predicts poor prognosis in human prostate carcinoma. Oncol. Rep. 2016, 36, 10–22. [Google Scholar] [CrossRef] [Green Version]
- Liang, Z.H.; Pan, Y.C.; Lin, S.S.; Qiu, Z.Y.; Zhang, Z. LncRNA MALAT1 promotes wound healing via regulating miR-141-3p/ZNF217 axis. Regen. Ther. 2020, 15, 202–209. [Google Scholar] [CrossRef]
- Qin, S.; Li, Y.; Cao, X.; Du, J.; Huang, X. NANOG regulates epithelial-mesenchymal transition and chemoresistance in ovarian cancer. Biosci. Rep. 2017, 37, BSR20160247. [Google Scholar] [CrossRef]
- Jiang, X.; Chen, Y.; Du, E.; Yang, K.; Zhang, Z.; Qi, S.; Xu, Y. GATA3-driven expression of miR-503 inhibits prostate cancer progression by repressing ZNF217 expression. Cell Signal. 2016, 28, 1216–1224. [Google Scholar] [CrossRef] [PubMed]
- Szczyrba, J.; Nolte, E.; Hart, M.; Doll, C.; Wach, S.; Taubert, H.; Keck, B.; Kremmer, E.; Stohr, R.; Hartmann, A.; et al. Identification of ZNF217, hnRNP-K, VEGF-A and IPO7 as targets for microRNAs that are downregulated in prostate carcinoma. Int. J. Cancer 2013, 132, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Si, W.; Zhao, Y.; Zhou, J.; Zhang, Q.; Zhang, Y. The coordination between ZNF217 and LSD1 contributes to hepatocellular carcinoma progress and is negatively regulated by miR-101. Exp. Cell Res. 2019, 379, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Toda, H.; Seki, N.; Kurozumi, S.; Shinden, Y.; Yamada, Y.; Nohata, N.; Moriya, S.; Idichi, T.; Maemura, K.; Fujii, T.; et al. RNA-sequence-based microRNA expression signature in breast cancer: Tumor-suppressive miR-101-5p regulates molecular pathogenesis. Mol. Oncol. 2020, 14, 426–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zhu, K.; Han, Q.; Wang, Q.; Yang, B. lncRNA PCAT1 might coordinate ZNF217 to promote CRC adhesion and invasion through regulating MTA2/MTA3/Snai1/E-cadherin signaling. Cell. Mol. Biol. 2022, 67, 1–9. [Google Scholar] [CrossRef]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.; Nieto, M.A. Epithelial-mesenchymal transitions in development and disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef]
- Massague, J. TGFbeta in Cancer. Cell 2008, 134, 215–230. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.J.; Wang, L.J.; Yu, B.; Li, Y.H.; Jin, Y.; Bai, X.Z. LncRNA-ATB promotes trastuzumab resistance and invasion-metastasis cascade in breast cancer. Oncotarget 2015, 6, 11652–11663. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.R.; Xu, Y.L.; Qian, J.; Wang, Y. Long non-coding RNA SNHG15 accelerates the progression of non-small cell lung cancer by absorbing miR-211-3p. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1536–1544. [Google Scholar] [CrossRef]
- Ge, P.; Cao, L.; Zheng, M.; Yao, Y.; Wang, W.; Chen, X. LncRNA SNHG1 contributes to the cisplatin resistance and progression of NSCLC via miR-330-5p/DCLK1 axis. Exp. Mol. Pathol. 2021, 120, 104633. [Google Scholar] [CrossRef]
- Xiong, X.; Feng, Y.; Li, L.; Yao, J.; Zhou, M.; Zhao, P.; Huang, F.; Zeng, L.; Yuan, L. Long noncoding RNA SNHG1 promotes breast cancer progression by regulation of LMO4. Oncol. Rep. 2020, 43, 1503–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Zhang, N.; Lv, C.; Li, N.; Li, X.; Li, W. lncRNA SNHG1 Knockdown Alleviates Amyloid-beta-Induced Neuronal Injury by Regulating ZNF217 via Sponging miR-361-3p in Alzheimer’s Disease. J. Alzheimers. Dis. 2020, 77, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Shi, F.; Du, Y.; Wang, Z.; Feng, Y.; Song, J.; Liu, Y.; Xiao, M. Long non-coding RNA CTBP1-AS2 enhances cervical cancer progression via up-regulation of ZNF217 through sponging miR-3163. Cancer Cell Int. 2020, 20, 343. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yue, W.; Li, M.; Jiang, Z.; Hou, Z.; Liu, W.; Ma, N.; Gan, W.; Li, Y.; Zhou, T.; et al. Downregulating Long Non-coding RNAs CTBP1-AS2 Inhibits Colorectal Cancer Development by Modulating the miR-93-5p/TGF-beta/SMAD2/3 Pathway. Front. Oncol. 2021, 11, 626620. [Google Scholar] [CrossRef]
- Yang, G.; Zhang, C. CTBP1-AS2 promoted non-small cell lung cancer progression via sponging the miR-623/MMP3 axis. Environ. Sci. Pollut. Res. Int. 2022, 29, 38385–38394. [Google Scholar] [CrossRef]
- Yang, Y.; Gao, M.; Li, Y.; Li, M.; Ma, Q. LncRNA CTBP1-AS2 Facilitates Gastric Cancer Progression via Regulating the miR-139-3p/MMP11 Axis. Onco Targets Ther. 2020, 13, 11537–11547. [Google Scholar] [CrossRef]
- Guo, L.; Chen, J.; Liu, D.; Liu, L. OIP5-AS1/miR-137/ZNF217 Axis Promotes Malignant Behaviors in Epithelial Ovarian Cancer. Cancer Manag. Res. 2020, 12, 6707–6717. [Google Scholar] [CrossRef]
- Jiang, Z.; Cheng, P.; Luo, B.; Huang, J. Construction and Analysis of a Long Non-Coding RNA-Associated Competing Endogenous RNA Network Identified Potential Prognostic Biomarkers in Luminal Breast Cancer. Onco Targets Ther. 2020, 13, 4271–4282. [Google Scholar] [CrossRef]
- Tan, S.; Khumalo, N.; Bayat, A. Understanding Keloid Pathobiology From a Quasi-Neoplastic Perspective: Less of a Scar and More of a Chronic Inflammatory Disease With Cancer-Like Tendencies. Front. Immunol. 2019, 10, 1810. [Google Scholar] [CrossRef]
- Zou, A.; Liu, P.; Liu, T.; Li, Q. Long non-coding RNA HOXA11-AS contributes to the formation of keloid by relieving the inhibition of miR-182-5p on ZNF217. Burns 2022, in press. [Google Scholar] [CrossRef]
- Qiang, F.; Li, J. CircCSNK1G1 Contributes to the Tumorigenesis of Gastric Cancer by Sponging miR-758 and Regulating ZNF217 Expression. Cancer Manag. Res. 2021, 13, 5027–5038. [Google Scholar] [CrossRef] [PubMed]
- Ou, H.; Li, J.; Lv, Q.; Feng, D. Hsa_circ_0069094 positively regulates the expression of oncogenic ZNF217 by competitively targeting miR-758-3p to promote the development of breast cancer. Reprod. Biol. 2022, 22, 100708. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Du, Q.; Wu, C. CircLPAR1/miR-212-3p/ZNF217 feedback loop promotes amyloid beta-induced neuronal injury in Alzheimer’s Disease. Brain Res. 2021, 1770, 147622. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Sheng, H.; Xie, H.; Zheng, Q.; Shen, Y.; Shi, G.; Ye, D. circLPAR1 is a novel biomarker of prognosis for muscle-invasive bladder cancer with invasion and metastasis by miR-762. Oncol. Lett. 2019, 17, 3537–3547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, R.; Zhang, K.; Tan, S.; Gao, F.; Zhang, Y.; Xu, W.; Wang, H.; Gu, D.; Zhu, L.; Li, S.; et al. Exosomal circLPAR1 functions in colorectal cancer diagnosis and tumorigenesis through suppressing BRD4 via METTL3-eIF3h interaction. Mol. Cancer 2022, 21, 49. [Google Scholar] [CrossRef]
- McIntyre, A.J.; Angel, C.Z.; Smith, J.S.; Templeman, A.; Beattie, K.; Beattie, S.; Ormrod, A.; Devlin, E.; McGreevy, C.; Bothwell, C.; et al. TBX2 acts as a potent transcriptional silencer of tumour suppressor genes through interaction with the CoREST complex to sustain the proliferation of breast cancers. Nucleic Acids Res. 2022, 50, 6154–6173. [Google Scholar] [CrossRef]
- Smolarz, B.; Nowak, A.Z.; Romanowicz, H. Breast Cancer-Epidemiology, Classification, Pathogenesis and Treatment (Review of Literature). Cancers 2022, 14, 2569. [Google Scholar] [CrossRef]
- Vendrell, J.A.; Solassol, J.; Gyorffy, B.; Vilquin, P.; Jarlier, M.; Donini, C.F.; Gamba, L.; Maudelonde, T.; Rouanet, P.; Cohen, P.A. Evaluating ZNF217 mRNA Expression Levels as a Predictor of Response to Endocrine Therapy in ER+ Breast Cancer. Front. Pharmacol. 2018, 9, 1581. [Google Scholar] [CrossRef] [Green Version]
- Vendrell, J.A.; Magnino, F.; Danis, E.; Duchesne, M.J.; Pinloche, S.; Pons, M.; Birnbaum, D.; Nguyen, C.; Theillet, C.; Cohen, P.A. Estrogen regulation in human breast cancer cells of new downstream gene targets involved in estrogen metabolism, cell proliferation and cell transformation. J. Mol. Endocrinol. 2004, 32, 397–414. [Google Scholar] [CrossRef] [Green Version]
- Cicatiello, L.; Mutarelli, M.; Grober, O.M.; Paris, O.; Ferraro, L.; Ravo, M.; Tarallo, R.; Luo, S.; Schroth, G.P.; Seifert, M.; et al. Estrogen receptor alpha controls a gene network in luminal-like breast cancer cells comprising multiple transcription factors and microRNAs. Am. J. Pathol. 2010, 176, 2113–2130. [Google Scholar] [CrossRef]
- Baran-Gale, J.; Purvis, J.E.; Sethupathy, P. An integrative transcriptomics approach identifies miR-503 as a candidate master regulator of the estrogen response in MCF-7 breast cancer cells. RNA 2016, 22, 1592–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prestat, E.; de Morais, S.R.; Vendrell, J.A.; Thollet, A.; Gautier, C.; Cohen, P.A.; Aussem, A. Learning the local Bayesian network structure around the ZNF217 oncogene in breast tumours. Comput. Biol. Med. 2013, 43, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Wang, N.; Zhang, Y.; Yang, Y.; Li, S.; Zheng, R.; Chai, J.; Qiao, T.; Jiang, S. Partial Deficiency of Zfp217 Resists High-Fat Diet-Induced Obesity by Increasing Energy Metabolism in Mice. Int. J. Mol. Sci. 2021, 22, 5390. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Ji, X.; Guo, X.; Ji, S. Regulatory Role of N(6)-methyladenosine (m(6) A) Methylation in RNA Processing and Human Diseases. J. Cell Biochem. 2017, 118, 2534–2543. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Frazier, K.; Zhang, J.; Gan, Z.; Wang, T.; Zhong, X. Emerging role of m(6) A RNA methylation in nutritional physiology and metabolism. Obes. Rev. 2020, 21, e12942. [Google Scholar] [CrossRef] [PubMed]
- McCormack, V.A.; dos Santos Silva, I. Breast density and parenchymal patterns as markers of breast cancer risk: A meta-analysis. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1159–1169. [Google Scholar] [CrossRef] [Green Version]
- Palomares, M.R.; Machia, J.R.; Lehman, C.D.; Daling, J.R.; McTiernan, A. Mammographic density correlation with Gail model breast cancer risk estimates and component risk factors. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1324–1330. [Google Scholar] [CrossRef] [Green Version]
- Acerbi, I.; Cassereau, L.; Dean, I.; Shi, Q.; Au, A.; Park, C.; Chen, Y.Y.; Liphardt, J.; Hwang, E.S.; Weaver, V.M. Human breast cancer invasion and aggression correlates with ECM stiffening and immune cell infiltration. Integr. Biol. 2015, 7, 1120–1134. [Google Scholar] [CrossRef] [Green Version]
- Northey, J.J.; Barrett, A.S.; Acerbi, I.; Hayward, M.K.; Talamantes, S.; Dean, I.S.; Mouw, J.K.; Ponik, S.M.; Lakins, J.N.; Huang, P.J.; et al. Stiff stroma increases breast cancer risk by inducing the oncogene ZNF217. J. Clin. Investig. 2020, 130, 5721–5737. [Google Scholar] [CrossRef]
- Clezardin, P.; Coleman, R.; Puppo, M.; Ottewell, P.; Bonnelye, E.; Paycha, F.; Confavreux, C.B.; Holen, I. Bone metastasis: Mechanisms, therapies, and biomarkers. Physiol. Rev. 2021, 101, 797–855. [Google Scholar] [CrossRef]
- D’Oronzo, S.; Coleman, R.; Brown, J.; Silvestris, F. Metastatic bone disease: Pathogenesis and therapeutic options: Up-date on bone metastasis management. J. Bone Oncol. 2019, 15, 004. [Google Scholar] [CrossRef] [PubMed]
- Coleman, R.E.; Croucher, P.I.; Padhani, A.R.; Clezardin, P.; Chow, E.; Fallon, M.; Guise, T.; Colangeli, S.; Capanna, R.; Costa, L. Bone metastases. Nat. Rev. Dis. Primers 2020, 6, 83. [Google Scholar] [CrossRef] [PubMed]
- Bellanger, A.; Donini, C.F.; Vendrell, J.A.; Lavaud, J.; Machuca-Gayet, I.; Ruel, M.; Vollaire, J.; Grisard, E.; Gyorffy, B.; Bieche, I.; et al. The critical role of the ZNF217 oncogene in promoting breast cancer metastasis to the bone. J. Pathol. 2017, 242, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Vollaire, J.; Machuca-Gayet, I.; Lavaud, J.; Bellanger, A.; Bouazza, L.; El Moghrabi, S.; Treilleux, I.; Coll, J.L.; Peyruchaud, O.; Josserand, V.; et al. The Bone Morphogenetic Protein Signaling Inhibitor LDN-193189 Enhances Metastasis Development in Mice. Front. Pharmacol. 2019, 10, 667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browne, G.; Taipaleenmaki, H.; Stein, G.S.; Stein, J.L.; Lian, J.B. MicroRNAs in the control of metastatic bone disease. Trends Endocrinol. Metab. 2014, 25, 320–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Oronzo, S.; Brown, J.; Coleman, R. The value of biomarkers in bone metastasis. Eur. J. Cancer Care 2017, 26, e12725. [Google Scholar] [CrossRef]
- Puppo, M.; Taipaleenmaki, H.; Hesse, E.; Clezardin, P. Non-coding RNAs in bone remodelling and bone metastasis: Mechanisms of action and translational relevance. Br. J. Pharmacol. 2021, 178, 1936–1954. [Google Scholar] [CrossRef] [PubMed]
- Puppo, M.; Valluru, M.K.; Clezardin, P. MicroRNAs and Their Roles in Breast Cancer Bone Metastasis. Curr. Osteoporos. Rep. 2021, 19, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Saini, S.; Majid, S.; Yamamura, S.; Tabatabai, L.; Suh, S.O.; Shahryari, V.; Chen, Y.; Deng, G.; Tanaka, Y.; Dahiya, R. Regulatory Role of mir-203 in Prostate Cancer Progression and Metastasis. Clin. Cancer Res. 2011, 17, 5287–5298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siu, M.K.; Abou-Kheir, W.; Yin, J.J.; Chang, Y.S.; Barrett, B.; Suau, F.; Casey, O.; Chen, W.Y.; Fang, L.; Hynes, P.; et al. Loss of EGFR signaling regulated miR-203 promotes prostate cancer bone metastasis and tyrosine kinase inhibitors resistance. Oncotarget 2014, 5, 3770–3784. [Google Scholar] [CrossRef] [PubMed]
- Taipaleenmaki, H.; Browne, G.; Akech, J.; Zustin, J.; van Wijnen, A.J.; Stein, J.L.; Hesse, E.; Stein, G.S.; Lian, J.B. Targeting of Runx2 by miR-135 and miR-203 Impairs Progression of Breast Cancer and Metastatic Bone Disease. Cancer Res. 2015, 75, 1433–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Wa, Q.; Pan, J.; Peng, X.; Ren, D.; Huang, Y.; Chen, X.; Tang, Y. Downregulation of miR-141-3p promotes bone metastasis via activating NF-kappaB signaling in prostate cancer. J. Exp. Clin. Cancer Res. 2017, 36, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Y.; Li, S.L.; Ma, Y.Y.; Diao, Y.J.; Yang, L.; Su, M.Q.; Li, Z.; Ji, Y.; Wang, J.; Lei, L.; et al. Exosomal miR-141-3p regulates osteoblast activity to promote the osteoblastic metastasis of prostate cancer. Oncotarget 2017, 8, 94834–94849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Sun, W.; Liu, Y.; Dong, X. The role of lncRNA MALAT1 in bone metastasis in patients with non-small cell lung cancer. Oncol. Rep. 2016, 36, 1679–1685. [Google Scholar] [CrossRef] [Green Version]
- Ren, S.; Liu, Y.; Xu, W.; Sun, Y.; Lu, J.; Wang, F.; Wei, M.; Shen, J.; Hou, J.; Gao, X.; et al. Long noncoding RNA MALAT-1 is a new potential therapeutic target for castration resistant prostate cancer. J. Urol. 2013, 190, 2278–2287. [Google Scholar] [CrossRef]
- Misawa, A.; Kondo, Y.; Takei, H.; Takizawa, T. Long Noncoding RNA HOXA11-AS and Transcription Factor HOXB13 Modulate the Expression of Bone Metastasis-Related Genes in Prostate Cancer. Genes 2021, 12, 182. [Google Scholar] [CrossRef]
- Anfossi, S.; Babayan, A.; Pantel, K.; Calin, G.A. Clinical utility of circulating non-coding RNAs—An update. Nat. Rev. Clin. Oncol. 2018, 15, 541–563. [Google Scholar] [CrossRef]
- An, X.; Quan, H.; Lv, J.; Meng, L.; Wang, C.; Yu, Z.; Han, J. Serum microRNA as potential biomarker to detect breast atypical hyperplasia and early-stage breast cancer. Future Oncol. 2018, 14, 3145–3161. [Google Scholar] [CrossRef]
- Cavallari, I.; Ciccarese, F.; Sharova, E.; Urso, L.; Raimondi, V.; Silic-Benussi, M.; D’Agostino, D.M.; Ciminale, V. The miR-200 Family of microRNAs: Fine Tuners of Epithelial-Mesenchymal Transition and Circulating Cancer Biomarkers. Cancers 2021, 13, 5874. [Google Scholar] [CrossRef]
- Gal, Z.; Gezsi, A.; Semsei, A.F.; Nagy, A.; Sultesz, M.; Csoma, Z.; Tamasi, L.; Galffy, G.; Szalai, C. Investigation of circulating lncRNAs as potential biomarkers in chronic respiratory diseases. J. Transl. Med. 2020, 18, 422. [Google Scholar] [CrossRef]
- Han, P.; Zhao, J.; Gao, L. Increased serum exosomal long non-coding RNA SNHG15 expression predicts poor prognosis in non-small cell lung cancer. J. Clin. Lab. Anal. 2021, 35, e23979. [Google Scholar] [CrossRef]
- Jin, W.; Fei, X.; Wang, X.; Chen, F.; Song, Y. Circulating miRNAs as Biomarkers for Prostate Cancer Diagnosis in Subjects with Benign Prostatic Hyperplasia. J. Immunol. Res. 2020, 2020, 5873056. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.R.; Kim, G.; Tak, W.Y.; Jang, S.Y.; Kweon, Y.O.; Park, J.G.; Lee, H.W.; Han, Y.S.; Chun, J.M.; Park, S.Y.; et al. Circulating exosomal noncoding RNAs as prognostic biomarkers in human hepatocellular carcinoma. Int J. Cancer 2019, 144, 1444–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.Y.; Li, Y.M.; Li, Y.; Shi, X.W.; Chen, H. Circulating microRNA-137 is a potential biomarker for human glioblastoma. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 3599–3604. [Google Scholar] [PubMed]
- Nassar, F.J.; Msheik, Z.S.; Itani, M.M.; Helou, R.E.; Hadla, R.; Kreidieh, F.; Bejjany, R.; Mukherji, D.; Shamseddine, A.; Nasr, R.R.; et al. Circulating miRNA as Biomarkers for Colorectal Cancer Diagnosis and Liver Metastasis. Diagnostics 2021, 11, 341. [Google Scholar] [CrossRef]
- Roth, C.; Stuckrath, I.; Pantel, K.; Izbicki, J.R.; Tachezy, M.; Schwarzenbach, H. Low levels of cell-free circulating miR-361-3p and miR-625* as blood-based markers for discriminating malignant from benign lung tumors. PLoS ONE 2012, 7, e38248. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.P.; Su, F.; Zhang, S.P.; Chen, H.K.; Li, Z.J.; Xing, G.Q.; Liu, H.J.; Li, Y.Y. miR-212 as potential biomarker suppresses the proliferation of gastric cancer via targeting SOX4. J. Clin. Lab. Anal. 2020, 34, e23511. [Google Scholar] [CrossRef]
- Toraih, E.A.; El-Wazir, A.; Ageeli, E.A.; Hussein, M.H.; Eltoukhy, M.M.; Killackey, M.T.; Kandil, E.; Fawzy, M.S. Unleash multifunctional role of long noncoding RNAs biomarker panel in breast cancer: A predictor classification model. Epigenomics 2020, 12, 1215–1237. [Google Scholar] [CrossRef]
- Wu, K.; Feng, J.; Lyu, F.; Xing, F.; Sharma, S.; Liu, Y.; Wu, S.Y.; Zhao, D.; Tyagi, A.; Deshpande, R.P.; et al. Exosomal miR-19a and IBSP cooperate to induce osteolytic bone metastasis of estrogen receptor-positive breast cancer. Nat. Commun. 2021, 12, 5196. [Google Scholar] [CrossRef]
- Yamana, K.; Inoue, J.; Yoshida, R.; Sakata, J.; Nakashima, H.; Arita, H.; Kawaguchi, S.; Gohara, S.; Nagao, Y.; Takeshita, H.; et al. Extracellular vesicles derived from radioresistant oral squamous cell carcinoma cells contribute to the acquisition of radioresistance via the miR-503-3p-BAK axis. J. Extracell. Vesicles 2021, 10, e12169. [Google Scholar] [CrossRef]
- Zheng, D.; Zhu, Y.; Zhang, J.; Zhang, W.; Wang, H.; Chen, H.; Wu, C.; Ni, J.; Xu, X.; Nian, B.; et al. Identification and evaluation of circulating small extracellular vesicle microRNAs as diagnostic biomarkers for patients with indeterminate pulmonary nodules. J. Nanobiotechnol. 2022, 20, 172. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kong, G. Roles and epigenetic regulation of epithelial-mesenchymal transition and its transcription factors in cancer initiation and progression. Cell Mol. Life Sci. 2016, 73, 4643–4660. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Chai, G.; Wu, Y.; Li, J.; Chen, F.; Liu, J.; Luo, G.; Tauler, J.; Du, J.; Lin, S.; et al. RNA m(6)A methylation regulates the epithelial mesenchymal transition of cancer cells and translation of Snail. Nat. Commun. 2019, 10, 2065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghasemi, S. Cancer’s epigenetic drugs: Where are they in the cancer medicines? Pharm. J. 2020, 20, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Slaby, O.; Laga, R.; Sedlacek, O. Therapeutic targeting of non-coding RNAs in cancer. Biochem. J. 2017, 474, 4219–4251. [Google Scholar] [CrossRef]
- Hong, D.S.; Kang, Y.K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H.Y.; Brenner, A.J.; Park, K.; Lee, J.L.; Kim, T.Y.; et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br. J. Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef]
- Tang, C.; Liu, J.; Hu, Q.; Zeng, S.; Yu, L. Metastatic colorectal cancer: Perspectives on long non-coding RNAs and promising therapeutics. Eur. J. Pharmacol. 2021, 908, 174367. [Google Scholar] [CrossRef]
- Abulwerdi, F.A.; Xu, W.; Ageeli, A.A.; Yonkunas, M.J.; Arun, G.; Nam, H.; Schneekloth, J.S., Jr.; Dayie, T.K.; Spector, D.; Baird, N.; et al. Selective Small-Molecule Targeting of a Triple Helix Encoded by the Long Noncoding RNA, MALAT1. ACS Chem. Biol. 2019, 14, 223–235. [Google Scholar] [CrossRef]
- Mercatelli, N.; Fortini, D.; Palombo, R.; Paronetto, M.P. Small molecule inhibition of Ewing sarcoma cell growth via targeting the long non coding RNA HULC. Cancer Lett. 2020, 469, 111–123. [Google Scholar] [CrossRef]
- Huang, H.; Weng, H.; Chen, J. m(6)A Modification in Coding and Non-coding RNAs: Roles and Therapeutic Implications in Cancer. Cancer Cell 2020, 37, 270–288. [Google Scholar] [CrossRef]
- Vandevenne, M.; Jacques, D.A.; Artuz, C.; Nguyen, C.D.; Kwan, A.H.; Segal, D.J.; Matthews, J.M.; Crossley, M.; Guss, J.M.; Mackay, J.P. New insights into DNA recognition by zinc fingers revealed by structural analysis of the oncoprotein ZNF217. J. Biol. Chem. 2013, 288, 10616–10627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smeester, B.A.; Draper, G.M.; Slipek, N.J.; Larsson, A.T.; Stratton, N.; Pomeroy, E.J.; Becklin, K.L.; Yamamoto, K.; Williams, K.B.; Laoharawee, K.; et al. Implication of ZNF217 in Accelerating Tumor Development and Therapeutically Targeting ZNF217-Induced PI3K-AKT Signaling for the Treatment of Metastatic Osteosarcoma. Mol. Cancer Ther. 2020, 19, 2528–2541. [Google Scholar] [CrossRef] [PubMed]
- Suarez, C.D.; Wu, J.; Badve, S.S.; Sparano, J.A.; Kaliney, W.; Littlepage, L.E. The AKT inhibitor triciribine in combination with paclitaxel has order-specific efficacy against Zfp217-induced breast cancer chemoresistance. Oncotarget 2017, 8, 108534–108547. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fahmé, P.; Ramadan, F.; Le, D.T.; Thi, K.-O.N.; Ghayad, S.E.; Hussein, N.; Diaz, C.; Croset, M.; Clézardin, P.; Cohen, P.A. The Intricate Interplay between the ZNF217 Oncogene and Epigenetic Processes Shapes Tumor Progression. Cancers 2022, 14, 6043. https://doi.org/10.3390/cancers14246043
Fahmé P, Ramadan F, Le DT, Thi K-ON, Ghayad SE, Hussein N, Diaz C, Croset M, Clézardin P, Cohen PA. The Intricate Interplay between the ZNF217 Oncogene and Epigenetic Processes Shapes Tumor Progression. Cancers. 2022; 14(24):6043. https://doi.org/10.3390/cancers14246043
Chicago/Turabian StyleFahmé, Pia, Farah Ramadan, Diep Tien Le, Kieu-Oanh Nguyen Thi, Sandra E. Ghayad, Nader Hussein, Chantal Diaz, Martine Croset, Philippe Clézardin, and Pascale A. Cohen. 2022. "The Intricate Interplay between the ZNF217 Oncogene and Epigenetic Processes Shapes Tumor Progression" Cancers 14, no. 24: 6043. https://doi.org/10.3390/cancers14246043
APA StyleFahmé, P., Ramadan, F., Le, D. T., Thi, K. -O. N., Ghayad, S. E., Hussein, N., Diaz, C., Croset, M., Clézardin, P., & Cohen, P. A. (2022). The Intricate Interplay between the ZNF217 Oncogene and Epigenetic Processes Shapes Tumor Progression. Cancers, 14(24), 6043. https://doi.org/10.3390/cancers14246043