
Modulation of DNA Damage Response by SAM and HD Domain Containing Deoxynucleoside Triphosphate Triphosphohydrolase (SAMHD1) Determines Prognosis and Treatment Efficacy in Different Solid Tumor Types

 , , , ,
, , , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Construction of Tissue Microarrays (TMA) and Immunohistochemical Methods
2.3. Cells Lines and SAMHD1 Knock-Out Generation
2.4. Western Blot Analysis
2.5. Cell Proliferation Assays
2.6. Cell Cycle Analysis
2.7. Quantitative RT-Polymerase Chain Reaction (qRT-PCR)
2.8. Drugs
2.9. Evaluation of Cytotoxicity
2.10. Evaluation of DNA Damage and Apoptosis
2.11. Statistical Analysis
3. Results
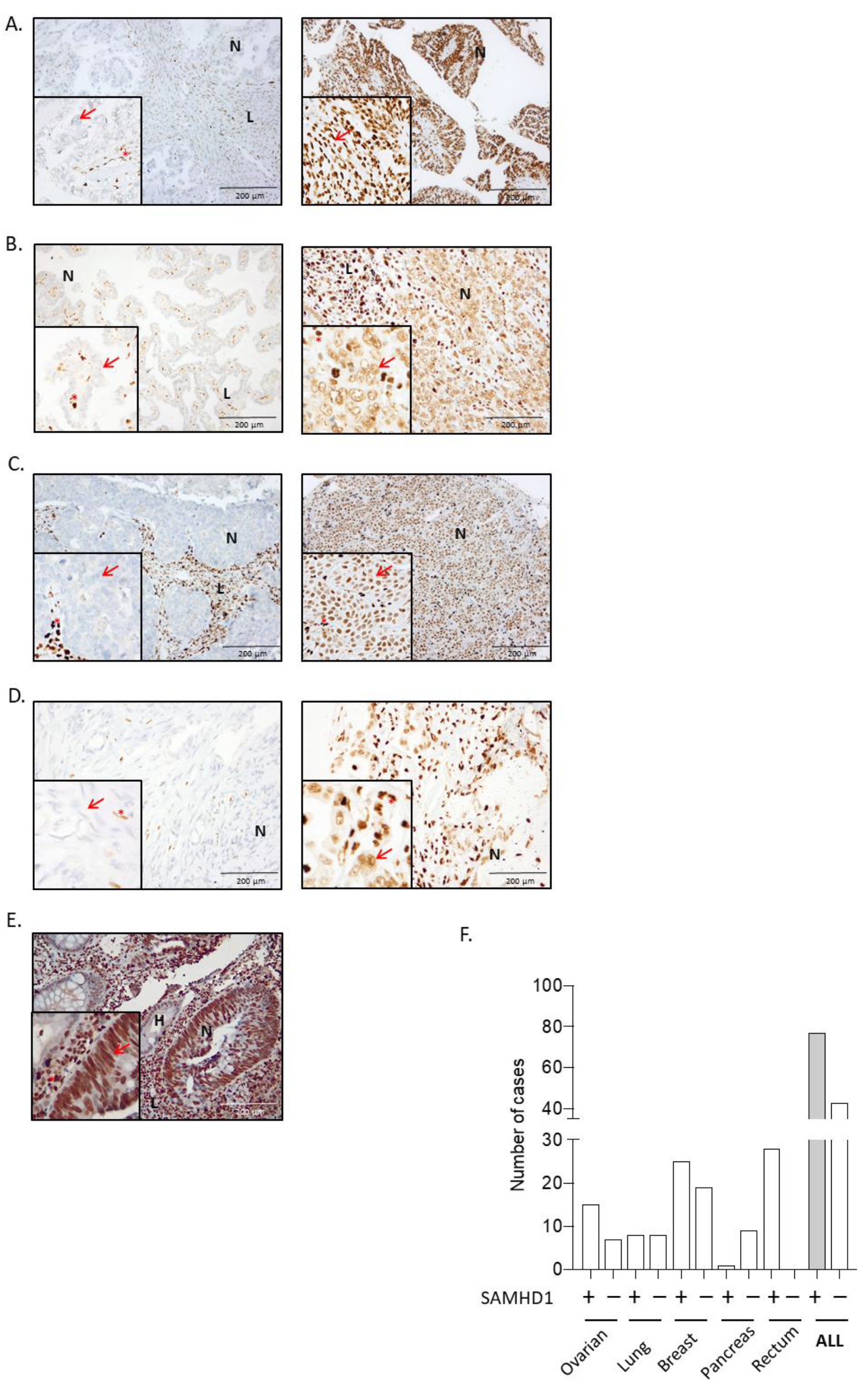
3.1. SAMHD1 Is Differentially Expressed in Solid Tumors and Correlates with Tumor Differentiation or Grade
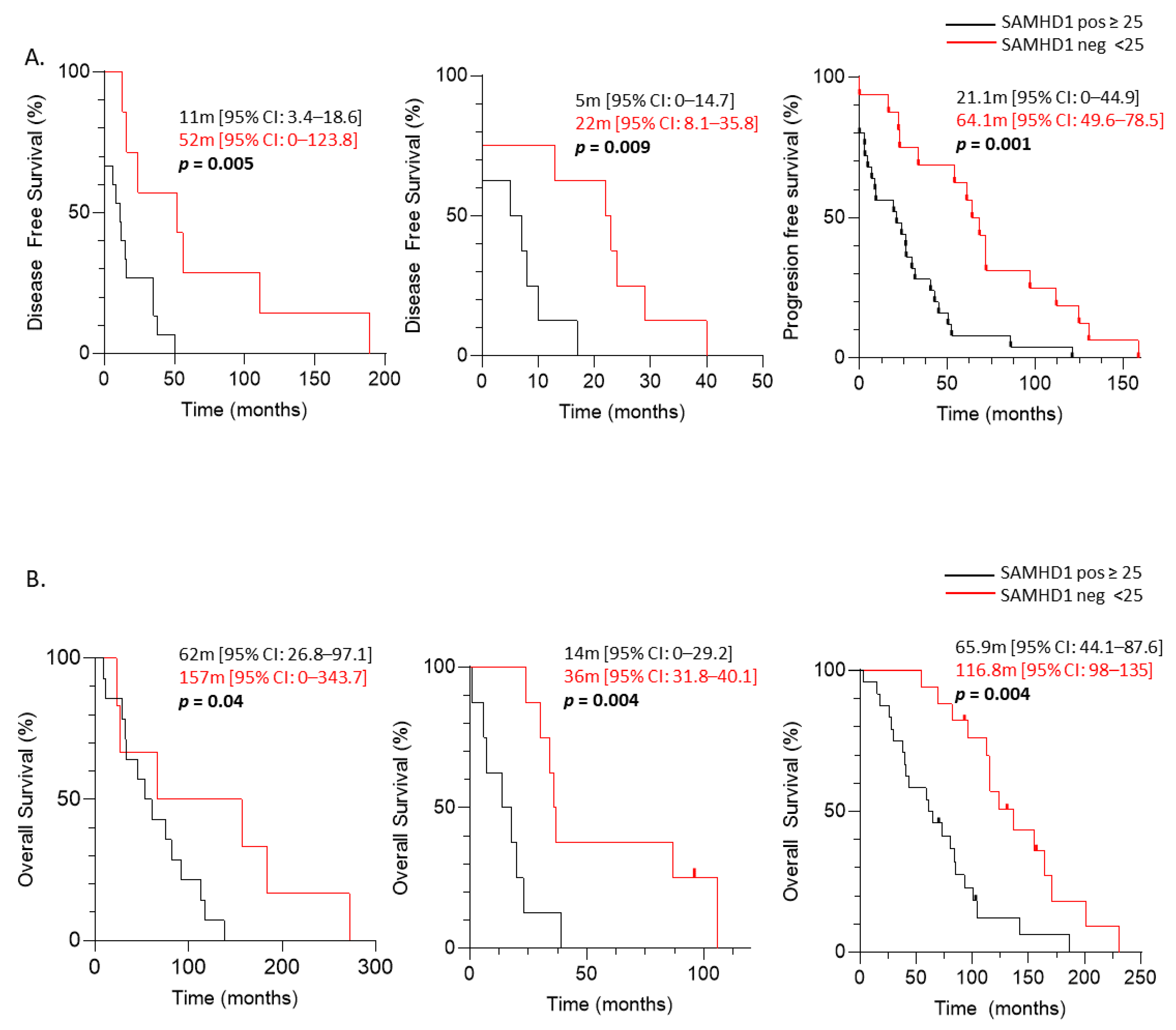
3.2. SAMHD1 Expression as a Negative Prognostic Factor in Breast, Ovarian, and NSCLC Patients
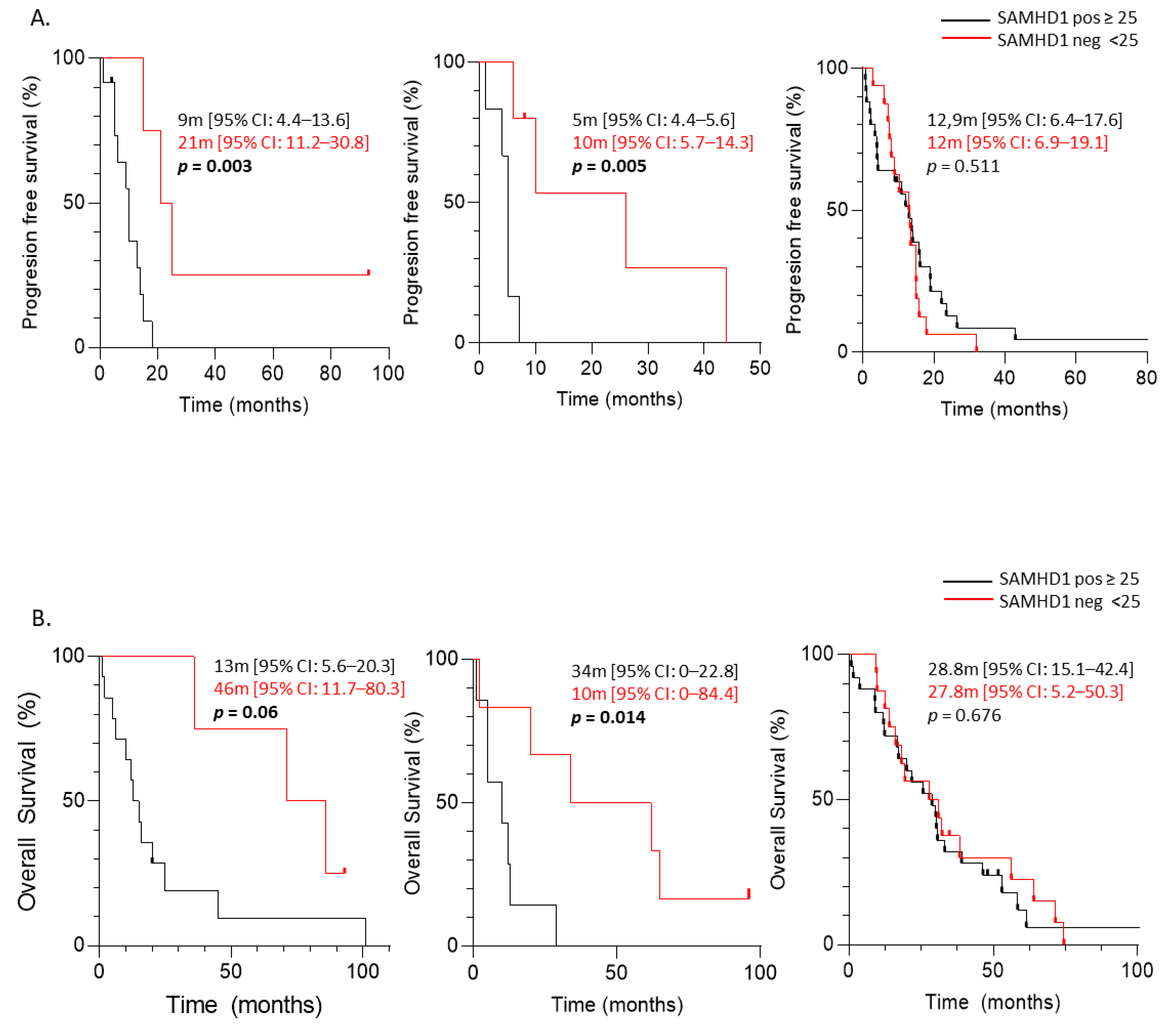
3.3. Predictive Significance of SAMHD1 Expression in Cancer Patients Treated with Antimetabolite- and/or Platin-Containing Regimens
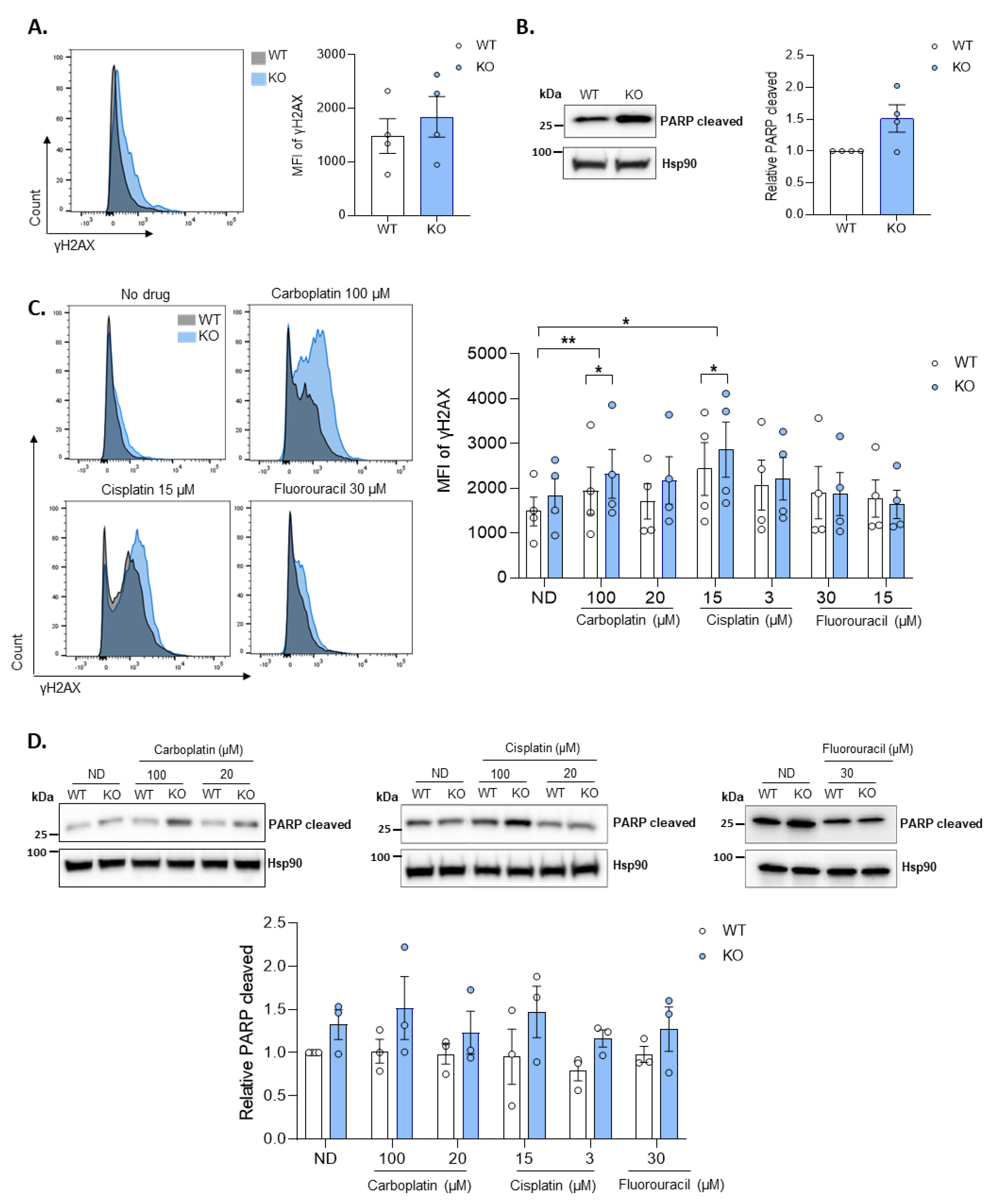
3.4. Loss of SAMHD1 Induced Cellular Apoptosis by Enhanced Genomic Instability and DNA Damage Response
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Reichard, P. Interactions between deoxyribonucleotide and DNA synthesis. Annu. Rev. Biochem. 1988, 57, 349–374. [Google Scholar] [CrossRef] [PubMed]
- Ballana, E.; Esté, J.A. SAMHD1: At the crossroads of cell proliferation, immune responses, and virus restriction. Trends Microbiol. 2015, 23, 680–692. [Google Scholar] [CrossRef] [PubMed]
- Knecht, K.M.; Buzovetsky, O.; Schneider, C.; Thomas, D.; Srikanth, V.; Kaderali, L.; Tofoleanu, F.; Reiss, K.; Ferreirós, N.; Geisslinger, G.; et al. The structural basis for cancer drug interactions with the catalytic and allosteric sites of SAMHD1. Proc. Natl. Acad. Sci. USA 2018, 115, E10022–E10031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, L.H.; Groom, H.C.T.; Kunzelmann, S.; Schwefel, D.; Caswell, S.J.; Ordonez, P.; Mann, M.C.; Rueschenbaum, S.; Goldstone, D.C.; Pennell, S.; et al. Phospho-dependent Regulation of SAMHD1 Oligomerisation Couples Catalysis and Restriction. PLoS Pathog. 2015, 11, e1005194. [Google Scholar] [CrossRef] [PubMed]
- Rice, G.I.; Bond, J.; Asipu, A.; Brunette, R.L.; Manfield, I.W.; Carr, I.M.; Fuller, J.C.; Jackson, R.M.; Lamb, T.; Briggs, T.A.; et al. Mutations involved in Aicardi-Goutières syndrome implicate SAMHD1 as regulator of the innate immune response. Nat. Genet. 2009, 41, 829–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clifford, R.; Louis, T.; Robbe, P.; Ackroyd, S.; Burns, A.; Timbs, A.T.; Wright Colopy, G.; Dreau, H.; Sigaux, F.; Judde, J.G.; et al. SAMHD1 is mutated recurrently in chronic lymphocytic leukemia and is involved in response to DNA damage. Blood 2014, 123, 1021–1031. [Google Scholar] [CrossRef]
- Landau, D.A.; Carter, S.L.; Stojanov, P.; McKenna, A.; Stevenson, K.; Lawrence, M.S.; Sougnez, C.; Stewart, C.; Sivachenko, A.; Wang, L.; et al. Evolution and impact of subclonal mutations in chronic lymphocytic leukemia. Cell 2013, 152, 714–726. [Google Scholar] [CrossRef] [Green Version]
- Rentoft, M.; Lindell, K.; Tran, P.; Chabes, A.L.; Buckland, R.J.; Watt, D.L.; Marjavaara, L.; Nilsson, A.K.; Melin, B.; Trygg, J.; et al. Heterozygous colon cancer-associated mutations of SAMHD1 have functional significance. Proc. Natl. Acad. Sci. USA 2016, 113, 4723–4728. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.A.; Wardell, C.P.; Melchor, L.; Hulkki, S.; Potter, N.E.; Johnson, D.C.; Fenwick, K.; Kozarewa, I.; Gonzalez, D.; Lord, C.J.; et al. Intraclonal heterogeneity and distinct molecular mechanisms characterize the development of t(4;14) and t(11;14) myeloma. Blood 2012, 120, 1077–1086. [Google Scholar] [CrossRef]
- Sjöblom, T.; Jones, S.; Wood, L.D.; Parsons, D.W.; Lin, J.; Barber, T.D.; Mandelker, D.; Leary, R.J.; Ptak, J.; Silliman, N.; et al. The consensus coding sequences of human breast and colorectal cancers. Science 2006, 314, 268–274. [Google Scholar] [CrossRef]
- Liu, J.; Lee, W.; Jiang, Z.; Chen, Z.; Jhunjhunwala, S.; Haverty, P.M.; Gnad, F.; Guan, Y.; Gilbert, H.N.; Stinson, J.; et al. Genome and transcriptome sequencing of lung cancers reveal diverse mutational and splicing events. Genome Res. 2012, 22, 2315–2327. [Google Scholar] [CrossRef] [Green Version]
- Jones, S.; Zhang, X.; Parsons, D.W.; Lin, J.C.H.; Leary, R.J.; Angenendt, P.; Mankoo, P.; Carter, H.; Kamiyama, H.; Jimeno, A.; et al. Core signaling pathways in human pancreatic cancers revealed by global genomic analyses. Science 2008, 321, 1801–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, D.W.; Jones, S.; Zhang, X.; Lin, J.C.H.; Leary, R.J.; Angenendt, P.; Mankoo, P.; Carter, H.; Siu, I.M.; Gallia, G.L.; et al. An integrated genomic analysis of human glioblastoma multiforme. Science 2008, 321, 1807–1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coggins, S.A.; Mahboubi, B.; Schinazi, R.F.; Kim, B. SAMHD1 functions and human diseases. Viruses 2020, 12, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.A.; Huang, H.Y.; Chang, Y.S.; Lin, C.L.; Lai, I.L.; Chang, J.G. DNA-Sensing and Nuclease Gene Expressions as Markers for Colorectal Cancer Progression. Oncology 2017, 92, 115–124. [Google Scholar] [CrossRef]
- Shang, Z.; Qian, L.; Liu, S.; Niu, X.; Qiao, Z.; Sun, Y.; Zhang, Y.; Fan, L.Y.; Guan, X.; Cao, C.X.; et al. Graphene Oxide-Facilitated Comprehensive Analysis of Cellular Nucleic Acid Binding Proteins for Lung Cancer. ACS Appl. Mater. Interfaces 2018, 10, 17756–17770. [Google Scholar] [CrossRef]
- Wang, J.L.; Lu, F.Z.; Shen, X.Y.; Wu, Y.; Zhao, L.T. SAMHD1 is down regulated in lung cancer by methylation and inhibits tumor cell proliferation. Biochem. Biophys. Res. Commun. 2014, 455, 229–233. [Google Scholar] [CrossRef]
- Ballana, E.; Badia, R.; Terradas, G.; Torres-Torronteras, J.; Ruiz, A.; Pauls, E.; Riveira-Muñoz, E.; Clotet, B.; Martí, R.; Esté, J.A. SAMHD1 specifically affects the antiviral potency of thymidine analog HIV reverse transcriptase inhibitors. Antimicrob. Agents Chemother. 2014, 58, 4804–4813. [Google Scholar] [CrossRef] [Green Version]
- Ordonez, P.; Kunzelmann, S.; Groom, H.C.T.; Yap, M.W.; Weising, S.; Meier, C.; Bishop, K.N.; Taylor, I.A.; Stoye, J.P. SAMHD1 enhances nucleoside-analogue efficacy against HIV-1 in myeloid cells. Sci. Rep. 2017, 7, 42824. [Google Scholar] [CrossRef]
- Herold, N.; Rudd, S.G.; Ljungblad, L.; Sanjiv, K.; Myrberg, I.H.; Paulin, C.B.J.; Heshmati, Y.; Hagenkort, A.; Kutzner, J.; Page, B.D.G.; et al. Targeting SAMHD1 with the Vpx protein to improve cytarabine therapy for hematological malignancies. Nat. Med. 2017, 23, 256–263. [Google Scholar] [CrossRef]
- Schneider, C.; Oellerich, T.; Baldauf, H.M.; Schwarz, S.M.; Thomas, D.; Flick, R.; Bohnenberger, H.; Kaderali, L.; Stegmann, L.; Cremer, A.; et al. SAMHD1 is a biomarker for cytarabine response and a therapeutic target in acute myeloid leukemia. Nat. Med. 2017, 23, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Castellví, M.; Felip, E.; Ezeonwumelu, I.; Badia, R.; Garcia-Vidal, E.; Pujantell, M.; Gutiérrez-Chamorro, L.; Teruel, I.; Martínez-Cardús, A.; Clotet, B.; et al. Pharmacological Modulation of SAMHD1 Activity by CDK4/6 Inhibitors Improves Anticancer Therapy. Cancers 2020, 12, 713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauney, C.H.; Hollis, T. SAMHD1: Recurring roles in cell cycle, viral restriction, cancer, and innate immunity. Autoimmunity 2018, 51, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, S.; Wolf, C.; König, N.; Staroske, W.; Guck, J.; Häusler, M.; Luksch, H.; Nguyen, L.A.; Kim, B.; Alexopoulou, D.; et al. SAMHD1 prevents autoimmunity by maintaining genome stability. Ann. Rheum. Dis. 2015, 74, e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathews, C.K. DNA precursor metabolism and genomic stability. FASEB J. 2006, 20, 1300–1314. [Google Scholar] [CrossRef] [Green Version]
- Poli, J.; Tsaponina, O.; Crabbé, L.; Keszthelyi, A.; Pantesco, V.; Chabes, A.; Lengronne, A.; Pasero, P. dNTP pools determine fork progression and origin usage under replication stress. EMBO J. 2012, 31, 883–894. [Google Scholar] [CrossRef]
- Coquel, F.; Neumayer, C.; Lin, Y.L.; Pasero, P. SAMHD1 and the innate immune response to cytosolic DNA during DNA replication. Curr. Opin. Immunol. 2019, 56, 24–30. [Google Scholar] [CrossRef]
- Zhang, Z.; Zheng, L.; Yu, Y.; Wu, J.; Yang, F.; Xu, Y.; Guo, Q.; Wu, X.; Cao, S.; Cao, L.; et al. Involvement of SAMHD1 in dNTP homeostasis and the maintenance of genomic integrity and oncotherapy (Review). Int. J. Oncol. 2020, 56, 879–888. [Google Scholar] [CrossRef]
- Cabello-Lobato, M.J.; Wang, S.; Schmidt, C.K. SAMHD1 Sheds Moonlight on DNA Double-Strand Break Repair. Trends Genet. 2017, 33, 895–897. [Google Scholar] [CrossRef] [Green Version]
- Protocol Development; CTEP. Common Terminology Criteria for Adverse Events (CTCAE). Available online: https://ctep.cancer.gov/protocoldevelopment/electronic_applications/ctc.htm (accessed on 15 January 2022).
- Christian, W.; Asamura, H.; Sobin, L.H. TNM Atlas (Union for International Cancer Control); Wiley-Blackwell: Hoboken, NJ, USA, 2014. [Google Scholar]
- Rassidakis, G.Z.; Herold, N.; Myrberg, I.H.; Tsesmetzis, N.; Rudd, S.G.; Henter, J.I.; Schaller, T.; Ng, S.B.; Chng, W.J.; Yan, B.; et al. Low-level expression of SAMHD1 in acute myeloid leukemia (AML) blasts correlates with improved outcome upon consolidation chemotherapy with high-dose cytarabine-based regimens. Blood Cancer J. 2018, 8, 98. [Google Scholar] [CrossRef]
- Badia, R.; Pujantell, M.; Torres-Torronteras, J.; Menéndez-Arias, L.; Martí, R.; Ruzo, A.; Pauls, E.; Clotet, B.; Ballana, E.; Esté, J.A.; et al. SAMHD1 is active in cycling cells permissive to HIV-1 infection. Antivir. Res. 2017, 142, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Herold, N.; Rudd, S.G.; Sanjiv, K.; Kutzner, J.; Myrberg, I.H.; Paulin, C.B.J.; Olsen, T.K.; Helleday, T.; Henter, J.I.; Schaller, T. With me or against me: Tumor suppressor and drug resistance activities of SAMHD1. Exp. Hematol. 2017, 52, 32–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daddacha, W.; Koyen, A.E.; Bastien, A.J.; Head, P.S.E.; Dhere, V.R.; Nabeta, G.N.; Connolly, E.C.; Werner, E.; Madden, M.Z.; Daly, M.B.; et al. SAMHD1 Promotes DNA End Resection to Facilitate DNA Repair by Homologous Recombination. Cell Rep. 2017, 20, 1921–1935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.Z.; Xu, M.; Pradhan, M.; Gorshkov, K.; Petersen, J.; Straus, M.R.; Zhu, W.; Shinn, P.; Guo, H.; Shen, M.; et al. Identifying SARS-CoV-2 entry inhibitors through drug repurposing screens of SARS-S and MERS-S pseudotyped particles. bioRxiv 2020, bioRixv:197988. [Google Scholar] [CrossRef]
- Caruso, L.B.; Martin, K.A.; Lauretti, E.; Hulse, M.; Siciliano, M.; Lupey-Green, L.N.; Abraham, A.; Skorski, T.; Tempera, I. Poly(ADP-ribose) Polymerase 1, PARP1, modifies EZH2 and inhibits EZH2 histone methyltransferase activity after DNA damage. Oncotarget 2018, 9, 10585–10605. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.Q.; Li, Y.; Yang, C.; Li, J.; Tan, S.H.; Chin, D.W.L.; Nakamura-Ishizu, A.; Yang, H.; Suda, T. PRMT5 Modulates Splicing for Genome Integrity and Preserves Proteostasis of Hematopoietic Stem Cells. Cell Rep. 2019, 26, 2316–2328.e6. [Google Scholar] [CrossRef] [Green Version]
- Miranda-Filho, A.; Bray, F.; Charvat, H.; Rajaraman, S.; Soerjomataram, I. The world cancer patient population (WCPP): An updated standard for international comparisons of population-based survival. Cancer Epidemiol. 2020, 69, 101802. [Google Scholar] [CrossRef]
- Schmidt, S.; Schenkova, K.; Adam, T.; Erikson, E.; Lehmann-Koch, J.; Sertel, S.; Verhasselt, B.; Fackler, O.T.; Lasitschka, F.; Keppler, O.T. SAMHD1′s protein expression profile in humans. J. Leukoc. Biol. 2015, 98, 5–14. [Google Scholar] [CrossRef]
- Kohnken, R.; Kodigepalli, K.M.; Wu, L. Regulation of deoxynucleotide metabolism in cancer: Novel mechanisms and therapeutic implications. Mol. Cancer 2015, 14, 176. [Google Scholar] [CrossRef] [Green Version]
- Kodigepalli, K.M.; Li, M.; Liu, S.L.; Wu, L. Exogenous expression of SAMHD1 inhibits proliferation and induces apoptosis in cutaneous T-cell lymphoma-derived HuT78 cells. Cell Cycle 2017, 16, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Bester, A.C.; Roniger, M.; Oren, Y.S.; Im, M.M.; Sarni, D.; Chaoat, M.; Bensimon, A.; Zamir, G.; Shewach, D.S.; Kerem, B. Nucleotide deficiency promotes genomic instability in early stages of cancer development. Cell 2011, 145, 435–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzolin, E.; Pontarin, G.; Rampazzo, C.; Miazzi, C.; Ferraro, P.; Palumbo, E.; Reichard, P.; Bianchi, V. The deoxynucleotide triphosphohydrolase SAMHD1 is a major regulator of DNA precursor pools in mammalian cells. Proc. Natl. Acad. Sci. USA 2013, 110, 14272–14277. [Google Scholar] [CrossRef] [Green Version]
- Bonifati, S.; Daly, M.B.; St. Gelais, C.; Kim, S.H.; Hollenbaugh, J.A.; Shepard, C.; Kennedy, E.M.; Kim, D.H.; Schinazi, R.F.; Kim, B.; et al. SAMHD1 controls cell cycle status, apoptosis and HIV-1 infection in monocytic THP-1 cells. Virology 2016, 495, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Chabes, A.; Stillman, B. Constitutively high dNTP concentration inhibits cell cycle progression and the DNA damage checkpoint in yeast Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2007, 104, 1183–1188. [Google Scholar] [CrossRef] [Green Version]
- Amin, N.A.; Seymour, E.; Saiya-Cork, K.; Parkin, B.; Shedden, K.; Malek, S.N. A quantitative analysis of subclonal and clonal gene mutations before and after therapy in chronic lymphocytic leukemia. Clin. Cancer Res. 2016, 22, 4525–4535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Silva, S.; Wang, F.; Hake, T.S.; Porcu, P.; Wong, H.K.; Wu, L. Downregulation of SAMHD1 expression correlates with promoter DNA methylation in Sézary syndrome patients. J. Investig. Dermatol. 2014, 134, 562–565. [Google Scholar] [CrossRef] [Green Version]
- Schuh, A.; Becq, J.; Humphray, S.; Alexa, A.; Burns, A.; Clifford, R.; Feller, S.M.; Grocock, R.; Henderson, S.; Khrebtukova, I.; et al. Monitoring chronic lymphocytic leukemia progression by whole genome sequencing reveals heterogeneous clonal evolution patterns. Blood 2012, 120, 4191–4196. [Google Scholar] [CrossRef] [Green Version]
- Park, K.; Ryoo, J.; Jeong, H.; Kim, M.; Lee, S.; Hwang, S.Y.; Ahn, J.; Kim, D.; Moon, H.C.; Baek, D.; et al. Aicardi-Goutières syndrome-associated gene SAMHD1 preserves genome integrity by preventing R-loop formation at transcription-replication conflict regions. PLoS Genet. 2021, 17, e1009523. [Google Scholar] [CrossRef]
- Schott, K.; Majer, C.; Bulashevska, A.; Childs, L.; Schmidt, M.H.H.; Rajalingam, K.; Munder, M.; König, R. SAMHD1 in cancer: Curse or cure? J. Mol. Med. 2021. [Google Scholar] [CrossRef]
- Xagoraris, I.; Vassilakopoulos, T.P.; Drakos, E.; Angelopoulou, M.K.; Panitsas, F.; Herold, N.; Medeiros, L.J.; Giakoumis, X.; Pangalis, G.A.; Rassidakis, G.Z. Expression of the novel tumour suppressor sterile alpha motif and HD domain-containing protein 1 is an independent adverse prognostic factor in classical Hodgkin lymphoma. Br. J. Haematol. 2021, 193, 488–496. [Google Scholar] [CrossRef]
- Herold, N.; Rudd, S.G.; Sanjiv, K.; Kutzner, J.; Bladh, J.; Paulin, C.B.J.; Helleday, T.; Henter, J.I.; Schaller, T. SAMHD1 protects cancer cells from various nucleoside-based antimetabolites. Cell Cycle 2017, 16, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Davenne, T.; Klintman, J.; Sharma, S.; Rigby, R.E.; Blest, H.T.W.; Cursi, C.; Bridgeman, A.; Dadonaite, B.; De Keersmaecker, K.; Hillmen, P.; et al. SAMHD1 Limits the Efficacy of Forodesine in Leukemia by Protecting Cells against the Cytotoxicity of dGTP. Cell Rep. 2020, 31, 107640. [Google Scholar] [CrossRef] [PubMed]
- Rudd, S.G.; Tsesmetzis, N.; Sanjiv, K.; Paulin, C.B.; Sandhow, L.; Kutzner, J.; Hed Myrberg, I.; Bunten, S.S.; Axelsson, H.; Zhang, S.M.; et al. Ribonucleotide reductase inhibitors suppress SAMHD 1 ara- CTP ase activity enhancing cytarabine efficacy. EMBO Mol. Med. 2020, 12, e10419. [Google Scholar] [CrossRef] [PubMed]
- Coquel, F.; Silva, M.J.; Técher, H.; Zadorozhny, K.; Sharma, S.; Nieminuszczy, J.; Mettling, C.; Dardillac, E.; Barthe, A.; Schmitz, A.L.; et al. SAMHD1 acts at stalled replication forks to prevent interferon induction. Nature 2018, 557, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Bermejo, M.; López-Huertas, M.R.; García-Pérez, J.; Climent, N.; Descours, B.; Ambrosioni, J.; Mateos, E.; Rodríguez-Mora, S.; Rus-Bercial, L.; Benkirane, M.; et al. Dasatinib inhibits HIV-1 replication through the interference of SAMHD1 phosphorylation in CD4+ T cells. Biochem. Pharmacol. 2016, 106, 30–45. [Google Scholar] [CrossRef]
- Lee, E.J.; Seo, J.H.; Park, J.-H.; Vo, T.T.L.; An, S.; Bae, S.-J.; Le, H.; Lee, H.S.; Wee, H.-J.; Lee, D.; et al. SAMHD1 acetylation enhances its deoxynucleotide triphosphohydrolase activity and promotes cancer cell proliferation. Oncotarget 2017, 8, 68517–68529. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Type of Tumor | ||||
|---|---|---|---|---|---|
| Rectal (n = 30) | Ovarian (n = 22) | Lung (n = 16) | Breast (n = 46) | Pancreas(n = 14) | |
| Age (y), Mean Interquartile range (IQR) | 62.77 (37–80) | 62.82 (51–82) | 61.88 (47–83) | 54.02 (29–84) | 63.57 (45–80) |
| Gender | |||||
| Male, n (%) Female, n (%) | 19(63.3%) 11(36.7%) | - 22 (100%) | 13 (81.3%) 3 (18.7%) | - 46 (100%) | 9 (64.3%) 5 (35.7%) |
| Line of Therapy | |||||
| Neoadjuvant, n (%) | 30(100%) | ||||
| First, n (%) | 1(4.5%) | 15 (32.6%) | 13 (92.9%) | ||
| Second, n (%) | 7 (31.8%) | 16 (100%) | 15 (32.6%) | 1 (7.1%) | |
| Third, n (%) | 6 (27.3%) | 8 (17.4%) | |||
| ≥Fourth, n (%) | 8 (36.4%) | 8 (17.4%) | |||
| Overall Response Rate (ORR) * | |||||
| Complete response (CR), n (%) | 3(13.6%) | 0 (0%) | 3 (18.7%) | ||
| Partial response (PR), n (%) | 6 (20%) | 8 (36.4%) | 5 (31.3%) | 1 (2.2%) | |
| Stable disease (SD), n (%) | 21(70%) | 3 (13.6%) | 6 (37.5%) | 17 (37%) | 2 (14.3%) |
| Progressive disease (PD), n (%) | 2 (6.7%) | 5 (22.8%) | 2 (12.5%) | 15 (32.6%) | 4 (28.6%) |
| Non-evaluable (NE), n (%) | 1 (3.3%) | 3 (13.6%) | 13 (28.2%) | 8 (57.1%) | |
| TTP #, months, median (IQR) | 26.00 (0.00–56.79) | 10.00 (4.68–15.31) | 6.00 (2.75–9.23) | 12.97 (9.06–16.89) | 6 (4.3–7.46) |
| OS £, months, median (IQR) | 77.85 (68.11–87.59) | 18.00 (9.68–26.31) | 13.00 (1.25–24.74) | 28.81 (18.95–38.67) | 11.00 (7.33–14.66) |
| DFS &, months, median (IQR) | |||||
| All | nd | 15.00 (10.40–19.59) | 8 (2.1–13.88) | 29.83 (17.83–41.82) | 6 (4.3–7.46) |
| Excluding “de novo” ¥ | nd | 24.00 (7.86–40.13) | 17 (4.05–29.94) | 40.41 (22.77–58.04) | 15 (11.90–18.09) |
| OSCD α, months, median (IQR) | nd | 66 (33.03–98.96) | 24.00 10.28–37.72 | 93.83 72.33–115.32 | 17.00 0–51.83 |
| SAMHD1 Expression | Type of Tumor (n) | ||||
|---|---|---|---|---|---|
| Rectal (n = 30) | Ovarian (n = 22) | Lung (n = 16) | Breast (n = 46) | Pancreas (n = 14) | |
| % Expression, mean (IQR) | 64.64 (25–90) | 55.82 (0–100) | 34.06 (0–100) | 27.88 (0–80) | 16.11 (0–75) |
| Positivity ratio evaluable patients,n (%) | 28 (100%) | 15 (68.2%) | 8 (50%) | 25 (61%) | 1 (11.1%) |
| Non evaluable,n | 2 | 0 | 0 | 5 | 5 |
| Tumor | Variable | SAMHD1 Positivity Ratio | p-Value * | |
|---|---|---|---|---|
| Positive (≥25%) | Negative (<25%) | |||
| Ovarian | Histology High grade serous papillary, n (%) Clear cell carcinoma, n (%) Low grade serous papillary, n (%) | 14 (82.3%) 1 (25%) 0 (0%) | 3 (17.7%) 3 (75%) 1 (100%) | 0.028 * |
| Lung | Histology Squamous, n (%) Adenocarcinoma, n (%) Poorly differentiated, n (%) | 2 (33.4%) 1 (20%) 5 (100%) | 4 (66.6%) 4 (80%) 0 (0%) | 0.024 * |
| Tumor grade I, n (%) II, n (%) III, n (%) | 1 (100%) 1 (14.3%) 7 (87.5%) | 0 (0%) 6 (85.7%) 1 (12.5%) | 0.011 * | |
| Breast | Tumor grade£ I, n (%) II, n (%) III, n (%) | 0 (%) 8 (40%) 16 (84.2%) | 0 (0%) 12 (60%) 3 (15. 8%) | 0.017 * |
| Variable | Type of Tumor (n) | |||||
|---|---|---|---|---|---|---|
| Ovarian (22) | Lung (16) | Breast (46) | ||||
| Pos. (≥25) | Neg. (<25) | Pos. (≥25) | Neg. (<25) | Pos. (≥25) | Neg. (<25) | |
| Clinical response rate | ||||||
| Complete response (CR), n (%) | 2 (66.6%) | 1 (33.4%) | 0 (0%) | 0 (0%) | 1 (100%) | 0 (0%) |
| Partial response (PR), n (%) | 4 (50%) | 4 (50%) | 0 (0%) | 5 (100%) | 11 (68.8%) | 5 (31.2%) |
| Stable disease (SD), n (%) | 3 (100%) | 0 (0%) | 4 (60%) | 2 (40%) | 2 (16.8%) | 10 (83.2%) |
| Progressive disease (PD), n (%) | 5 (100%) | 0 (0%) | 2 (100%) | 0 (0%) | 11 (91.6%) | 1 (8.4%) |
| Overall response rate (ORR) √ | ||||||
| Yes, n (%) | 6 (54.5.5%) | 5 (45.5%) | 0 (0%) | 5 (100%) | 12 (70.5%) | 5 (29.5%) |
| No, n (%) | 8 (100%) | 0 (0%) | 6 (75%) | 2 (25%) | 13 (54.2%) | 11 (45.8%) |
| p-value * | 0.04 * | 0.016 * | 0.232 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Felip, E.; Gutiérrez-Chamorro, L.; Gómez, M.; Garcia-Vidal, E.; Romeo, M.; Morán, T.; Layos, L.; Pérez-Roca, L.; Riveira-Muñoz, E.; Clotet, B.; et al. Modulation of DNA Damage Response by SAM and HD Domain Containing Deoxynucleoside Triphosphate Triphosphohydrolase (SAMHD1) Determines Prognosis and Treatment Efficacy in Different Solid Tumor Types. Cancers 2022, 14, 641. https://doi.org/10.3390/cancers14030641
Felip E, Gutiérrez-Chamorro L, Gómez M, Garcia-Vidal E, Romeo M, Morán T, Layos L, Pérez-Roca L, Riveira-Muñoz E, Clotet B, et al. Modulation of DNA Damage Response by SAM and HD Domain Containing Deoxynucleoside Triphosphate Triphosphohydrolase (SAMHD1) Determines Prognosis and Treatment Efficacy in Different Solid Tumor Types. Cancers. 2022; 14(3):641. https://doi.org/10.3390/cancers14030641
Chicago/Turabian StyleFelip, Eudald, Lucía Gutiérrez-Chamorro, Maica Gómez, Edurne Garcia-Vidal, Margarita Romeo, Teresa Morán, Laura Layos, Laia Pérez-Roca, Eva Riveira-Muñoz, Bonaventura Clotet, and et al. 2022. "Modulation of DNA Damage Response by SAM and HD Domain Containing Deoxynucleoside Triphosphate Triphosphohydrolase (SAMHD1) Determines Prognosis and Treatment Efficacy in Different Solid Tumor Types" Cancers 14, no. 3: 641. https://doi.org/10.3390/cancers14030641
APA StyleFelip, E., Gutiérrez-Chamorro, L., Gómez, M., Garcia-Vidal, E., Romeo, M., Morán, T., Layos, L., Pérez-Roca, L., Riveira-Muñoz, E., Clotet, B., Fernandez, P. L., Mesía, R., Martínez-Cardús, A., Ballana, E., & Margelí, M. (2022). Modulation of DNA Damage Response by SAM and HD Domain Containing Deoxynucleoside Triphosphate Triphosphohydrolase (SAMHD1) Determines Prognosis and Treatment Efficacy in Different Solid Tumor Types. Cancers, 14(3), 641. https://doi.org/10.3390/cancers14030641