
ATP8B1 Knockdown Activated the Choline Metabolism Pathway and Induced High-Level Intracellular REDOX Homeostasis in Lung Squamous Cell Carcinoma

Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
2.1. Low Expression of ATP8B1 Was Associated with Poor Prognosis of LUSC
2.2. ATP8B1 Knockdown Promoted Proliferation, Inhibited Apoptosis, and Aggravated Invasion and Migration of LUSC Cells In Vitro and In Vivo
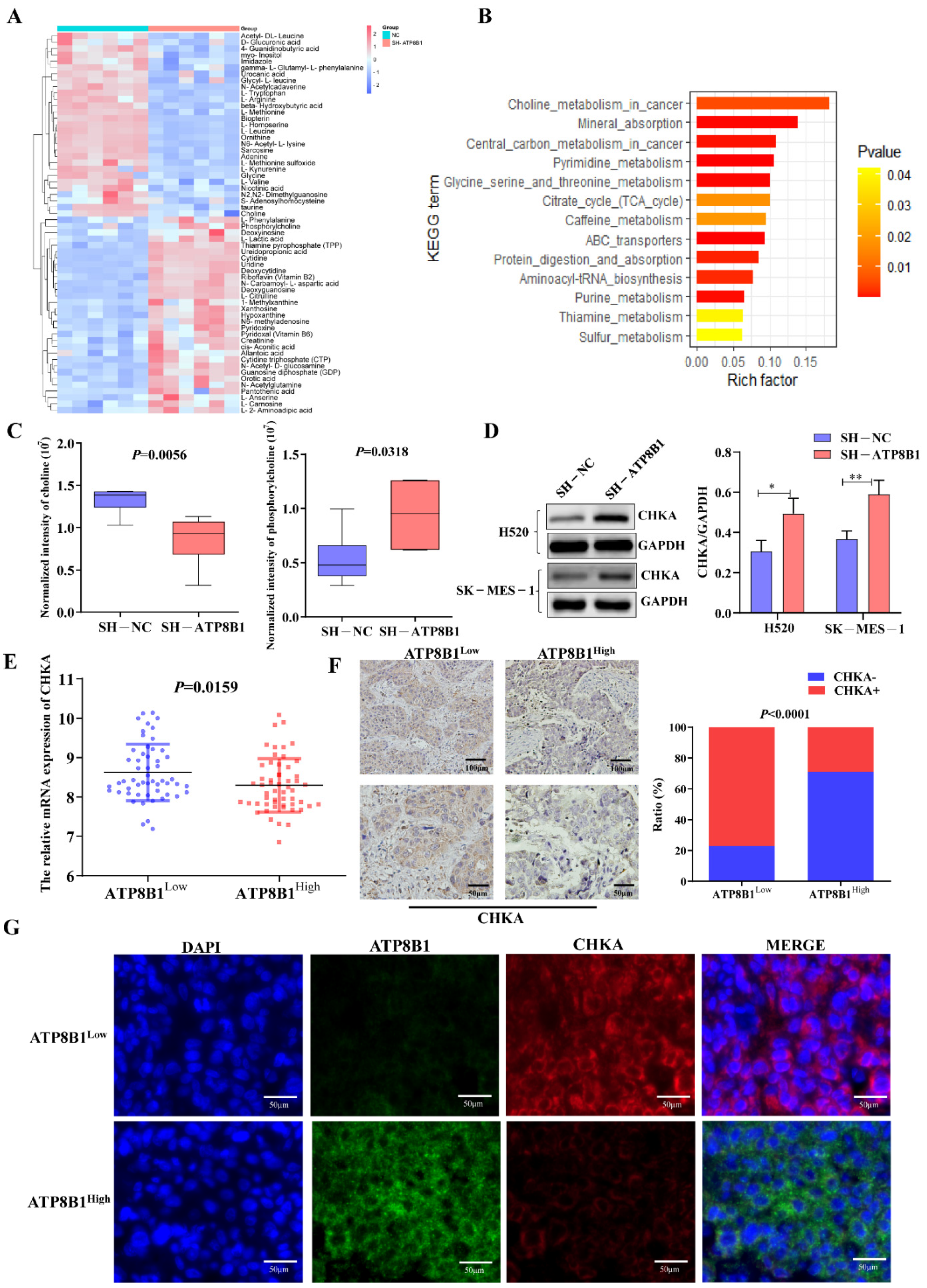
2.3. ATP8B1 Knockdown Activated the Choline Metabolic Pathway via Upregulation of Choline Kinase Expression
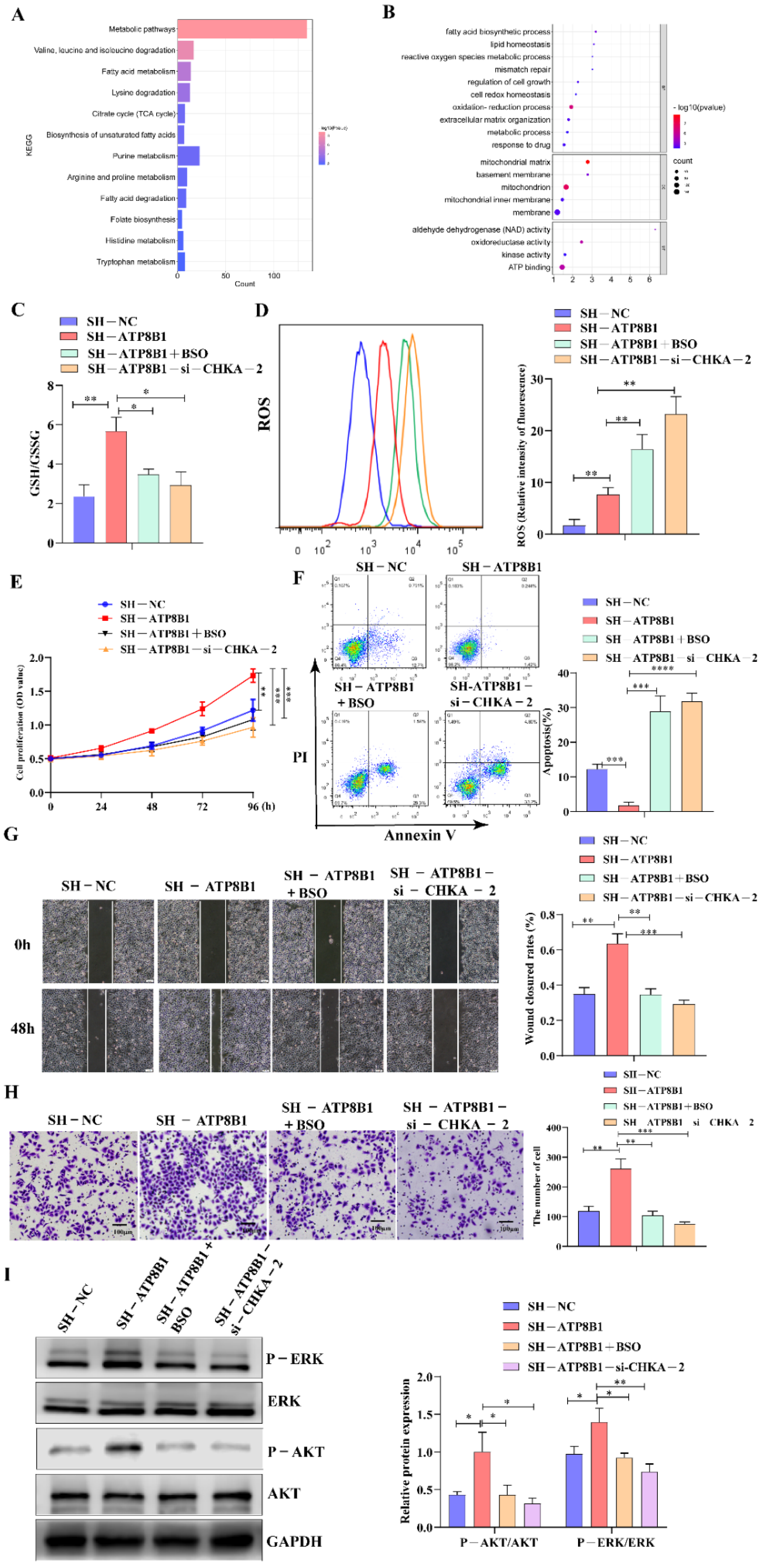
2.4. ATP8B1 Knockdown Promoted Cell Proliferation and Invasion in LUSC in A CHKA-Dependent Manner
2.5. ATP8B1 Knockdown Activated the Choline Metabolism Pathway and Induced High-Level Intracellular REDOX Homeostasis in LUSC
3. Discussion
4. Materials and Methods
4.1. Patient Information
4.2. Cell Lines and Cell Treatment
4.3. Mouse Models
4.4. Lentivirus Construction and Plasmid Transfection
4.5. RNA Extraction, Reverse Transcription-Polymerase Chain Reaction (RT-PCR), and Quantitative Real-Time PCR (qPCR) Analysis
4.6. RNA Library Preparation, Sequencing, and Enrichment Analysis
4.7. Immunohistochemistry
4.8. Multispectral Fluorescent Immunohistochemistry
4.9. Cell Proliferation Assay
4.10. Cell Apoptosis Assay
4.11. Wound Healing Assay
4.12. Trans-Well Invasion Assay
4.13. Western Blot
4.14. Metabonomics
4.15. Determination of Intracellular Reactive Oxygen Species (ROS)
4.16. Glutathione (GSH) Detection Assay
4.17. Phosphorylcholine Detection
4.18. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pan, Y.; Han, H.; Labbe, K.E.; Zhang, H.; Wong, K.-K. Recent advances in preclinical models for lung squamous cell carcinoma. Oncogene 2021, 40, 2817–2829. [Google Scholar] [CrossRef] [PubMed]
- Thai, A.A.; Solomon, B.J.; Sequist, L.V.; Gainor, J.F.; Heist, R.S. Lung cancer. Lancet 2021, 398, 535–554. [Google Scholar] [CrossRef]
- Alvarez, J.G.B.; González-Cao, M.; Karachaliou, N.; Santarpia, M.; Viteri, S.; Teixidó, C.; Rosell, R. Advances in immunotherapy for treatment of lung cancer. Cancer Biol. Med. 2015, 12, 209–222. [Google Scholar] [CrossRef]
- Faubert, B.; Solmonson, A.; DeBerardinis, R.J. Metabolic reprogramming and cancer progression. Science 2020, 368, eaaw5473. [Google Scholar] [CrossRef] [PubMed]
- Vanhove, K.; Derveaux, E.; Graulus, G.-J.; Mesotten, L.; Thomeer, M.; Noben, J.-P.; Guedens, W.; Adriaensens, P. Glutamine Addiction and Therapeutic Strategies in Lung Cancer. Int. J. Mol. Sci. 2019, 20, 252. [Google Scholar] [CrossRef] [Green Version]
- DeBerardinis, R.J.; Chandel, N.S. Fundamentals of cancer metabolism. Sci. Adv. 2016, 2, e1600200. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, R.; Yu, J. New Understanding of the Relevant Role of LINE-1 Retrotransposition in Human Disease and Immune Modulation. Front. Cell Dev. Biol. 2020, 8, 657. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, F.; Sun, Z.; Liu, P.; Zhang, X.; Ye, Y.; Cai, B.; Walsh, M.J.; Ren, X.; Hao, X.; et al. LINE-1 Retrotransposition Promotes the Development and Progression of Lung Squamous Cell Carcinoma by Disrupting the Tumor-Suppressor Gene FGGY. Cancer Res. 2019, 79, 4453–4465. [Google Scholar] [CrossRef] [Green Version]
- Korneenko, T.V.; Pestov, N.B.; Okkelman, I.A.; Modyanov, N.N.; Shakhparonov, M.I. P4-ATP-ase Atp8b1/FIC1: Structural properties and (patho)physiological functions. Bioorg. Khim. 2015, 41, 3–12. [Google Scholar] [CrossRef]
- Deng, L.; Niu, G.-M.; Ren, J.; Ke, C.-W.; Loura, L. Identification of ATP8B1 as a Tumor Suppressor Gene for Colorectal Cancer and Its Involvement in Phospholipid Homeostasis. BioMed Res. Int. 2020, 2020, 2015648. [Google Scholar] [CrossRef]
- Alyabsi, M.; Alhumaid, A.; Allah-Bakhsh, H.; Alkelya, M.; Aziz, M.A. Colorectal cancer in Saudi Arabia as the proof-of-principle model for implementing strategies of predictive, preventive, and personalized medicine in healthcare. EPMA J. 2019, 11, 119–131. [Google Scholar] [CrossRef]
- Fukumoto, J.; Leung, J.; Cox, R.; Czachor, A.; Parthasarathy, P.T.; Lagishetty, V.; Mandry, M.; Hosseinian, N.; Patel, P.; Perry, B.; et al. Oxidative stress induces club cell proliferation and pulmonary fibrosis in Atp8b1 mutant mice. Aging 2019, 11, 209–229. [Google Scholar] [CrossRef]
- Falcon, S.C.; Hudson, C.S.; Huang, Y.; Mortimore, M.; Golec, J.M.; A Charlton, P.; Weber, P.; Sundaram, H. A non-catalytic role of choline kinase alpha is important in promoting cancer cell survival. Oncogenesis 2013, 2, e38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spadaro, F.; Ramoni, C.; Mezzanzanica, D.; Miotti, S.; Alberti, P.; Cecchetti, S.; Iorio, E.; Dolo, V.; Canevari, S.; Podo, F.; et al. Phosphatidylcholine-Specific Phospholipase C Activation in Epithelial Ovarian Cancer Cells. Cancer Res. 2008, 68, 6541–6549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paris, L.; Cecchetti, S.; Spadaro, F.; Abalsamo, L.; Lugini, L.; Pisanu, M.E.; Iorio, E.; Natali, P.G.; Ramoni, C.; Podo, F. Inhibition of phosphatidylcholine-specific phospholipase C downregulates HER2 overexpression on plasma membrane of breast cancer cells. Breast Cancer Res. 2010, 12, R27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glunde, K.; Bhujwalla, Z.M.; Ronen, S.M. Choline metabolism in malignant transformation. Nat. Cancer 2011, 11, 835–848. [Google Scholar] [CrossRef] [Green Version]
- Clem, B.F.; Clem, A.L.; Yalcin, A.; Goswami, U.; Arumugam, S.; Telang, S.; Trent, J.O.; Chesney, J. A novel small molecule antagonist of choline kinase-α that simultaneously suppresses MAPK and PI3K/AKT signaling. Oncogene 2011, 30, 3370–3380. [Google Scholar] [CrossRef] [Green Version]
- Chio, I.I.C.; Tuveson, D.A. ROS in Cancer: The Burning Question. Trends Mol. Med. 2017, 23, 411–429. [Google Scholar] [CrossRef] [Green Version]
- Vogg, A.T.J.; Drude, N.; Mottaghy, F.M.; Morgenroth, A.; Miran, T. Modulation of glutathione promotes apoptosis in triple-negative breast cancer cells. FASEB J. 2018, 32, 2803–2813. [Google Scholar] [CrossRef] [Green Version]
- Nayagam, J.S.; Williamson, C.; Joshi, D.; Thompson, R.J. Review article: Liver disease in adults with variants in the cholestasis-related genes ABCB11, ABCB4 and ATP8B1. Aliment. Pharmacol. Ther. 2020, 52, 1628–1639. [Google Scholar]
- Cheng, M.; Rizwan, A.; Jiang, L.; Bhujwalla, Z.M.; Glunde, K. Molecular Effects of Doxorubicin on Choline Metabolism in Breast Cancer. Neoplasia 2017, 19, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Sonkar, K.; Ayyappan, V.; Tressler, C.; Adelaja, O.; Cai, R.; Cheng, M.; Glunde, K. Focus on the glycerophosphocholine pathway in choline phospholipid metabolism of cancer. NMR Biomed. 2019, 32, e4112. [Google Scholar] [CrossRef] [PubMed]
- Granata, A.; Nicoletti, R.; Perego, P.; Iorio, E.; Krishnamachary, B.; Benigni, F.; Ricci, A.; Podo, F.; Bhujwalla, Z.M.; Canevari, S.; et al. Global metabolic profile identifies choline kinase alpha as a key regulator of glutathione-dependent antioxidant cell defense in ovarian carcinoma. Oncotarget 2015, 6, 11216–11230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trousil, S.; Lee, P.; Pinato, D.J.; Ellis, J.K.; Dina, R.; Aboagye, E.O.; Keun, H.C.; Sharma, R. Alterations of Choline Phospholipid Metabolism in Endometrial Cancer Are Caused by Choline Kinase Alpha Overexpression and a Hyperactivated Deacylation Pathway. Cancer Res. 2014, 74, 6867–6877. [Google Scholar] [CrossRef] [Green Version]
- de Molina, A.R.; Estrada, J.S.; Iniesta, C.B.; Tarón, M.; de Molina, V.R.; Cejas, P.; Skrzypski, M.; Gallego-Ortega, D.; de Castro, J.; Casado, E.; et al. Expression of choline kinase alpha to predict outcome in patients with early-stage non-small-cell lung cancer: A retrospective study. Lancet Oncol. 2007, 8, 889–897. [Google Scholar] [CrossRef]
- De Molina, A.R.; De La Cueva, A.; Machado-Pinilla, R.; Rodriguez-Fanjul, V.; Del Pulgar, T.G.; Cebrian, A.; Perona, R.; Lacal, J.C. Acid ceramidase as a chemotherapeutic target to overcome resistance to the antitumoral effect of choline kinase α inhibition. Curr. Cancer Drug Targets 2012, 12, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Kall, S.L.; Delikatny, E.J.; Lavie, A. Identification of a Unique Inhibitor-Binding Site on Choline Kinase α. Biochemistry 2018, 57, 1316–1325. [Google Scholar] [CrossRef]
- Lacal, J.C.; Campos, J. Preclinical Characterization of RSM-932A, a Novel Anticancer Drug Targeting the Human Choline Kinase Alpha, an Enzyme Involved in Increased Lipid Metabolism of Cancer Cells. Mol. Cancer Ther. 2014, 14, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Ruiz, B.; Figuerola-Conchas, A.; Ramos-Torrecillas, J.; Capitán-Cañadas, F.; Ríos-Marco, P.; Carrasco, M.P.; Gallo, M.; Espinosa, A.; Marco, C.; Ruiz, C.; et al. Discovery of a New Binding Site on Human Choline Kinase α1: Design, Synthesis, Crystallographic Studies, and Biological Evaluation of Asymmetrical Bispyridinium Derivatives. J. Med. Chem. 2014, 57, 507–515. [Google Scholar] [CrossRef]
- Sahún-Roncero, M.; Rubio-Ruiz, B.; Saladino, G.; Conejo-García, A.; Espinosa, A.; Velazquez-Campoy, A.; Gervasio, F.L.; Entrena, A.; Hurtado-Guerrero, R. The Mechanism of Allosteric Coupling in Choline Kinase α1 Revealed by the Action of a Rationally Designed Inhibitor. Angew. Chem. 2013, 52, 4582–4586. [Google Scholar] [CrossRef]
- Moloney, J.N.; Cotter, T.G. ROS signalling in the biology of cancer. Semin. Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Schraut, K.; Kalnytska, O.; Lamp, D.; Jastroch, M.; Eder, M.; Hausch, F.; Gassen, N.C.; Moore, S.; Nagaraj, N.; Lopez, J.P.; et al. Loss of the psychiatric risk factor SLC6A15 is associated with increased metabolic functions in primary hippocampal neurons. Eur. J. Neurosci. 2020, 53, 390–401. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, J.C.; Amaral, A.U.; Cecatto, C.; Wajner, A.; Godoy, K.D.S.; Ribeiro, R.T.; Gonçalves, A.D.M.; Zanatta, Â.; Da Rosa, M.S.; Loureiro, S.O.; et al. α-Ketoadipic Acid and α-Aminoadipic Acid Cause Disturbance of Glutamatergic Neurotransmission and Induction of Oxidative Stress In Vitro in Brain of Adolescent Rats. Neurotox. Res. 2017, 32, 276–290. [Google Scholar] [CrossRef]
- Bolaños, J.P. Bioenergetics and redox adaptations of astrocytes to neuronal activity. J. Neurochem. 2016, 139, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Yi, J.; Lu, J.; Nie, M.; Huang, M.; Rong, J.; Zhu, Z.; Chen, J.; Zhou, X.; Li, B.; et al. N-Acetylcysteine Reduces ROS-Mediated Oxidative DNA Damage and PI3K/Akt Pathway Activation Induced by Helicobacter pylori Infection. Oxidative Med. Cell. Longev. 2018, 2018, 1874985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, A.-R.; Ku, S.-Y.; Cho, M.S.; Kim, Y.Y.; Kim, Y.J.; Oh, S.K.; Kim, S.H.; Moon, S.Y.; Choi, Y.M. Reactive oxygen species enhance differentiation of human embryonic stem cells into mesendodermal lineage. Exp. Mol. Med. 2010, 42, 175–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Pathological Parameters | Number of Patients |
|---|---|
| Total | 109 |
| Gender | |
| Male | 81 |
| Female | 28 |
| Age | |
| <60 | 52 |
| ≥60 | 57 |
| Stage | |
| I–II | 67 |
| III–IV | 42 |
| T stage | |
| 1–2 | 76 |
| 3–4 | 33 |
| N stage | |
| 0–1 | 77 |
| 2–3 | 32 |
| M stage | |
| 0 | 96 |
| 1 | 13 |
| Metastatic site | |
| Negative | 96 |
| Lung | 6 |
| Bone | 5 |
| Brain | 1 |
| Others | 2 |
| Location | |
| Central | 50 |
| Periphery | 59 |
| Smoking | |
| Negative | 20 |
| Positive | 89 |
| Clinical Pathological Parameters | ATP8B1 | p | |
|---|---|---|---|
| Low Expression | High Expression | ||
| No. of patients | 54 | 55 | - |
| Gender | 0.094 | ||
| Male | 40 | 48 | |
| Female | 14 | 7 | |
| Age | 0.567 | ||
| <60 | 26 | 30 | |
| ≥60 | 28 | 25 | |
| OS | 0.004 | ||
| ≤40 | 38 | 23 | |
| >40 | 16 | 32 | |
| T stage | 0.002 | ||
| 1–2 | 30 | 46 | |
| 3–4 | 24 | 9 | |
| N stage | 0.003 | ||
| 0–1 | 31 | 23 | |
| 2–3 | 23 | 9 | |
| M stage | 0.042 | ||
| 0 | 44 | 52 | |
| 1 | 10 | 3 | |
| Location | 0.848 | ||
| Periphery | 30 | 29 | |
| Central | 24 | 26 | |
| Smoking | 0.332 | ||
| Negative | 12 | 8 | |
| Positive | 42 | 47 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Zhang, R.; Liu, P.; Zhang, R.; Ning, J.; Ye, Y.; Yu, W.; Yu, J. ATP8B1 Knockdown Activated the Choline Metabolism Pathway and Induced High-Level Intracellular REDOX Homeostasis in Lung Squamous Cell Carcinoma. Cancers 2022, 14, 835. https://doi.org/10.3390/cancers14030835
Zhang X, Zhang R, Liu P, Zhang R, Ning J, Ye Y, Yu W, Yu J. ATP8B1 Knockdown Activated the Choline Metabolism Pathway and Induced High-Level Intracellular REDOX Homeostasis in Lung Squamous Cell Carcinoma. Cancers. 2022; 14(3):835. https://doi.org/10.3390/cancers14030835
Chicago/Turabian StyleZhang, Xiao, Rui Zhang, Pengpeng Liu, Runjiao Zhang, Junya Ning, Yingnan Ye, Wenwen Yu, and Jinpu Yu. 2022. "ATP8B1 Knockdown Activated the Choline Metabolism Pathway and Induced High-Level Intracellular REDOX Homeostasis in Lung Squamous Cell Carcinoma" Cancers 14, no. 3: 835. https://doi.org/10.3390/cancers14030835
APA StyleZhang, X., Zhang, R., Liu, P., Zhang, R., Ning, J., Ye, Y., Yu, W., & Yu, J. (2022). ATP8B1 Knockdown Activated the Choline Metabolism Pathway and Induced High-Level Intracellular REDOX Homeostasis in Lung Squamous Cell Carcinoma. Cancers, 14(3), 835. https://doi.org/10.3390/cancers14030835