
Transcriptional Factor Repertoire of Breast Cancer in 3D Cell Culture Models


Abstract
:Simple Summary
Abstract
1. Introduction
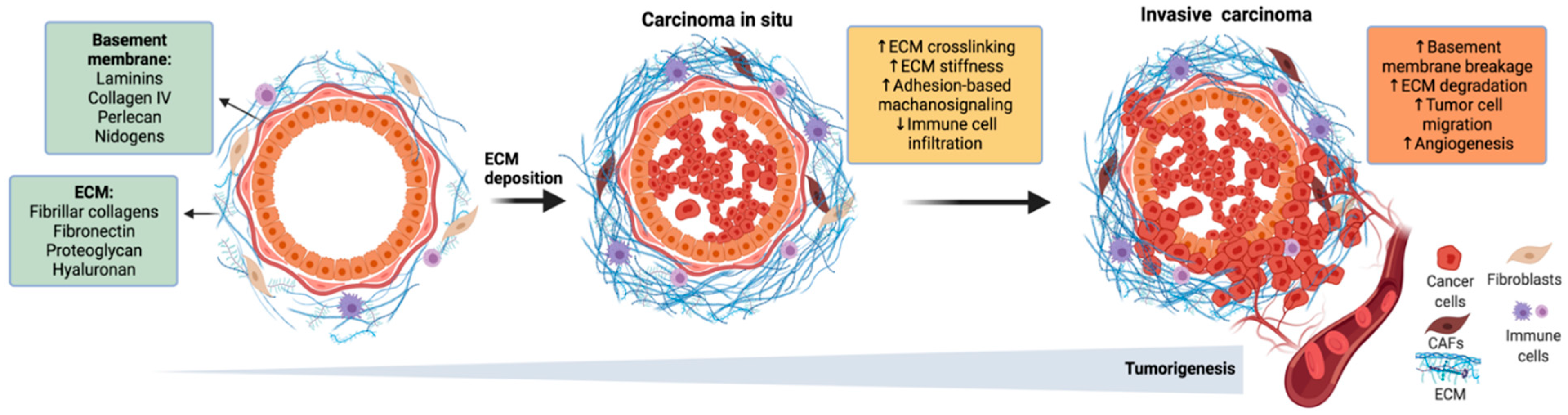
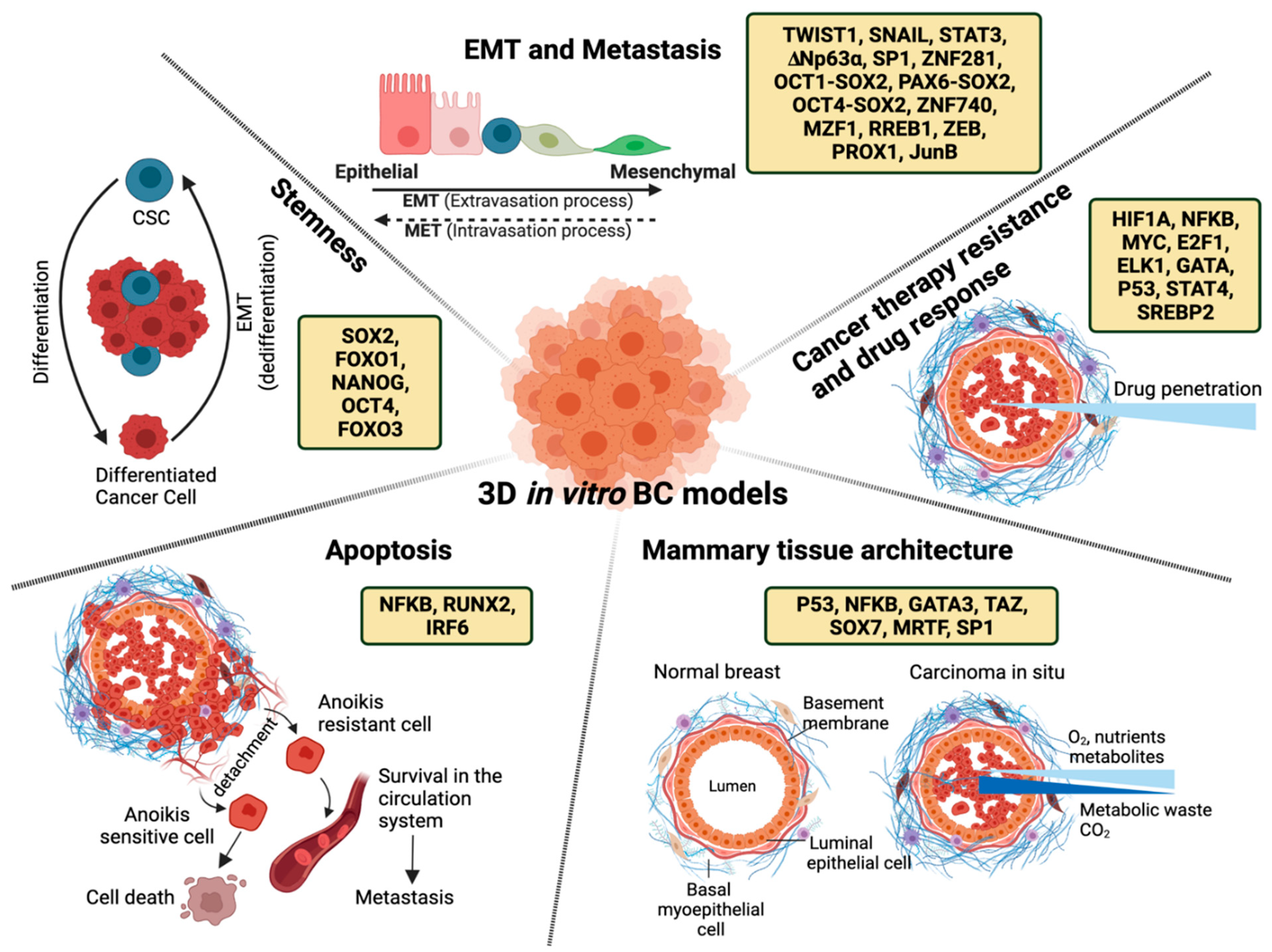
2. Transcriptional Regulation of Mammary Tissue Architecture
3. Transcriptional Regulation of Epithelial to Mesenchymal Transition (EMT) and Metastasis
4. Transcriptional Regulation of Apoptosis
5. Transcriptional Regulation of Cancer Therapy Resistance and Drug Response
6. Transcriptional Regulation of Stemness
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Polyak, K. Heterogeneity in Breast Cancer. J. Clin. Investig. 2011, 121, 3786–3788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turashvili, G.; Brogi, E. Tumor Heterogeneity in Breast Cancer. Front. Med. 2017, 4, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makki, J. Diversity of Breast Carcinoma: Histological Subtypes and Clinical Relevance. Clin. Med. Insights Pathol. 2015, 8, 23–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorlie, T.; Tibshirani, R.; Parker, J.; Hastie, T.; Marron, J.S.; Nobel, A.; Deng, S.; Johnsen, H.; Pesich, R.; Geisler, S.; et al. Repeated Observation of Breast Tumor Subtypes in Independent Gene Expression Data Sets. Proc. Natl. Acad. Sci. USA 2003, 100, 8418–8423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sørlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene Expression Patterns of Breast Carcinomas Distinguish Tumor Subclasses with Clinical Implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perou, C.M.; Sørlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular Portraits of Human Breast Tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef]
- Russnes, H.G.; Navin, N.; Hicks, J.; Borresen-Dale, A.-L. Insight into the Heterogeneity of Breast Cancer through next-Generation Sequencing. J. Clin. Investig. 2011, 121, 3810–3818. [Google Scholar] [CrossRef] [Green Version]
- Prat, A.; Parker, J.S.; Karginova, O.; Fan, C.; Livasy, C.; Herschkowitz, J.I.; He, X.; Perou, C.M. Phenotypic and Molecular Characterization of the Claudin-Low Intrinsic Subtype of Breast Cancer. Breast Cancer Res. 2010, 12, R68. [Google Scholar] [CrossRef] [Green Version]
- Curtis, C.; Shah, S.P.; Chin, S.-F.; Turashvili, G.; Rueda, O.M.; Dunning, M.J.; Speed, D.; Lynch, A.G.; Samarajiwa, S.; Yuan, Y.; et al. The Genomic and Transcriptomic Architecture of 2000 Breast Tumours Reveals Novel Subgroups. Nature 2012, 486, 346–352. [Google Scholar] [CrossRef]
- Park, S.Y.; Gönen, M.; Kim, H.J.; Michor, F.; Polyak, K. Cellular and Genetic Diversity in the Progression of in Situ Human Breast Carcinomas to an Invasive Phenotype. J. Clin. Investig. 2010, 120, 636–644. [Google Scholar] [CrossRef] [Green Version]
- Ellis, M.J.; Perou, C.M. The Genomic Landscape of Breast Cancer as a Therapeutic Roadmap. Cancer Discov. 2013, 3, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abd El-Rehim, D.M.; Ball, G.; Pinder, S.E.; Rakha, E.; Paish, C.; Robertson, J.F.R.; Macmillan, D.; Blamey, R.W.; Ellis, I.O. High-Throughput Protein Expression Analysis Using Tissue Microarray Technology of a Large Well-Characterised Series Identifies Biologically Distinct Classes of Breast Cancer Confirming Recent cDNA Expression Analyses. Int. J. Cancer 2005, 116, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Meacham, C.E.; Morrison, S.J. Tumour Heterogeneity and Cancer Cell Plasticity. Nature 2013, 501, 328–337. [Google Scholar] [CrossRef] [Green Version]
- Prat, A.; Perou, C.M. Deconstructing the Molecular Portraits of Breast Cancer. Mol. Oncol. 2011, 5, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Lüönd, F.; Tiede, S.; Christofori, G. Breast Cancer as an Example of Tumour Heterogeneity and Tumour Cell Plasticity during Malignant Progression. Br. J. Cancer 2021, 125, 164–175. [Google Scholar] [CrossRef]
- Poltavets, V.; Kochetkova, M.; Pitson, S.M.; Samuel, M.S. The Role of the Extracellular Matrix and Its Molecular and Cellular Regulators in Cancer Cell Plasticity. Front. Oncol. 2018, 8, 431. [Google Scholar] [CrossRef] [Green Version]
- Pickup, M.W.; Mouw, J.K.; Weaver, V.M. The Extracellular Matrix Modulates the Hallmarks of Cancer. EMBO Rep. 2014, 15, 1243–1253. [Google Scholar] [CrossRef] [Green Version]
- Winkler, J.; Abisoye-Ogunniyan, A.; Metcalf, K.J.; Werb, Z. Concepts of Extracellular Matrix Remodelling in Tumour Progression and Metastasis. Nat. Commun. 2020, 11, 5120. [Google Scholar] [CrossRef]
- Bissell, M.J.; Hall, H.G.; Parry, G. How Does the Extracellular Matrix Direct Gene Expression? J. Theor. Biol. 1982, 99, 31–68. [Google Scholar] [CrossRef]
- Xu, R.; Boudreau, A.; Bissell, M.J. Tissue Architecture and Function: Dynamic Reciprocity via Extra- and Intra-Cellular Matrices. Cancer Metastasis Rev. 2009, 28, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Le Beyec, J.; Xu, R.; Lee, S.-Y.; Nelson, C.M.; Rizki, A.; Alcaraz, J.; Bissell, M.J. Cell Shape Regulates Global Histone Acetylation in Human Mammary Epithelial Cells. Exp. Cell Res. 2007, 313, 3066–3075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, V.A.; Xu, R.; Bissell, M.J. Extracellular Matrix, Nuclear and Chromatin Structure, and Gene Expression in Normal Tissues and Malignant Tumors: A Work in Progress. Adv. Cancer Res. 2007, 97, 275–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roskelley, C.D.; Desprez, P.Y.; Bissell, M.J. Extracellular Matrix-Dependent Tissue-Specific Gene Expression in Mammary Epithelial Cells Requires Both Physical and Biochemical Signal Transduction. Proc. Natl. Acad. Sci. USA 1994, 91, 12378–12382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plachot, C.; Lelièvre, S.A. DNA Methylation Control of Tissue Polarity and Cellular Differentiation in the Mammary Epithelium. Exp. Cell Res. 2004, 298, 122–132. [Google Scholar] [CrossRef]
- Schmidhauser, C.; Casperson, G.F.; Myers, C.A.; Sanzo, K.T.; Bolten, S.; Bissell, M.J. A Novel Transcriptional Enhancer Is Involved in the Prolactin- and Extracellular Matrix-Dependent Regulation of Beta-Casein Gene Expression. Mol. Biol. Cell 1992, 3, 699–709. [Google Scholar] [CrossRef] [Green Version]
- Schatzmann, F.; Marlow, R.; Streuli, C.H. Integrin Signaling and Mammary Cell Function. J. Mammary Gland Biol. Neoplasia 2003, 8, 395–408. [Google Scholar] [CrossRef]
- Estrada, M.F.; Rebelo, S.P.; Davies, E.J.; Pinto, M.T.; Pereira, H.; Santo, V.E.; Smalley, M.J.; Barry, S.T.; Gualda, E.J.; Alves, P.M.; et al. Modelling the Tumour Microenvironment in Long-Term Microencapsulated 3D Co-Cultures Recapitulates Phenotypic Features of Disease Progression. Biomaterials 2016, 78, 50–61. [Google Scholar] [CrossRef] [Green Version]
- Xia, T.; Du, W.-L.; Chen, X.-Y.; Zhang, Y.-N. Organoid Models of the Tumor Microenvironment and Their Applications. J. Cell. Mol. Med. 2021, 25, 5829–5841. [Google Scholar] [CrossRef]
- Pampaloni, F.; Reynaud, E.G.; Stelzer, E.H.K. The Third Dimension Bridges the Gap between Cell Culture and Live Tissue. Nat. Rev. Mol. Cell Biol. 2007, 8, 839–845. [Google Scholar] [CrossRef]
- Hongisto, V.; Jernström, S.; Fey, V.; Mpindi, J.-P.; Kleivi Sahlberg, K.; Kallioniemi, O.; Perälä, M. High-Throughput 3D Screening Reveals Differences in Drug Sensitivities between Culture Models of JIMT1 Breast Cancer Cells. PLoS ONE 2013, 8, e77232. [Google Scholar] [CrossRef] [Green Version]
- Langhans, S.A. Three-Dimensional Cell Culture Models in Drug Discovery and Drug Repositioning. Front. Pharmacol. 2018, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- Hoarau-Véchot, J.; Rafii, A.; Touboul, C.; Pasquier, J. Halfway between 2D and Animal Models: Are 3D Cultures the Ideal Tool to Study Cancer-Microenvironment Interactions? Int. J. Mol. Sci. 2018, 19, 181. [Google Scholar] [CrossRef] [Green Version]
- Antoni, D.; Burckel, H.; Josset, E.; Noel, G. Three-Dimensional Cell Culture: A Breakthrough in Vivo. Int. J. Mol. Sci. 2015, 16, 5517–5527. [Google Scholar] [CrossRef]
- Mabry, K.M.; Payne, S.Z.; Anseth, K.S. Microarray Analyses to Quantify Advantages of 2D and 3D Hydrogel Culture Systems in Maintaining the Native Valvular Interstitial Cell Phenotype. Biomaterials 2016, 74, 31–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chitcholtan, K.; Asselin, E.; Parent, S.; Sykes, P.H.; Evans, J.J. Differences in Growth Properties of Endometrial Cancer in Three Dimensional (3D) Culture and 2D Cell Monolayer. Exp. Cell Res. 2013, 319, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Duval, K.; Grover, H.; Han, L.-H.; Mou, Y.; Pegoraro, A.F.; Fredberg, J.; Chen, Z. Modeling Physiological Events in 2D vs. 3D Cell Culture. Physiology 2017, 32, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Bissell, M.J.; Rizki, A.; Mian, I.S. Tissue Architecture: The Ultimate Regulator of Breast Epithelial Function. Curr. Opin. Cell Biol. 2003, 15, 753–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birgersdotter, A.; Sandberg, R.; Ernberg, I. Gene Expression Perturbation in Vitro--a Growing Case for Three-Dimensional (3D) Culture Systems. Semin. Cancer Biol. 2005, 15, 405–412. [Google Scholar] [CrossRef]
- Bokhari, M.; Carnachan, R.J.; Cameron, N.R.; Przyborski, S.A. Culture of HepG2 Liver Cells on Three Dimensional Polystyrene Scaffolds Enhances Cell Structure and Function during Toxicological Challenge. J. Anat. 2007, 211, 567–576. [Google Scholar] [CrossRef]
- Riedl, A.; Schlederer, M.; Pudelko, K.; Stadler, M.; Walter, S.; Unterleuthner, D.; Unger, C.; Kramer, N.; Hengstschläger, M.; Kenner, L.; et al. Comparison of Cancer Cells in 2D vs 3D Culture Reveals Differences in AKT-mTOR-S6K Signaling and Drug Responses. J. Cell Sci. 2017, 130, 203–218. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Huang, G.; Li, M.; Wang, L.; Elson, E.L.; Lu, T.J.; Genin, G.M.; Xu, F. An Approach to Quantifying 3D Responses of Cells to Extreme Strain. Sci. Rep. 2016, 6, 19550. [Google Scholar] [CrossRef] [PubMed]
- Breslin, S.; O’Driscoll, L. Three-Dimensional Cell Culture: The Missing Link in Drug Discovery. Drug Discov. Today 2013, 18, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Kapałczyńska, M.; Kolenda, T.; Przybyła, W.; Zajączkowska, M.; Teresiak, A.; Filas, V.; Ibbs, M.; Bliźniak, R.; Łuczewski, Ł.; Lamperska, K. 2D and 3D Cell Cultures—A Comparison of Different Types of Cancer Cell Cultures. Arch. Med. Sci. 2018, 14, 910–919. [Google Scholar] [CrossRef] [PubMed]
- Bissell, M.J.; Barcellos-Hoff, M.H. The Influence of Extracellular Matrix on Gene Expression: Is Structure the Message? J. Cell Sci. Suppl. 1987, 8, 327–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, O.W.; Rønnov-Jessen, L.; Howlett, A.R.; Bissell, M.J. Interaction with Basement Membrane Serves to Rapidly Distinguish Growth and Differentiation Pattern of Normal and Malignant Human Breast Epithelial Cells. Proc. Natl. Acad. Sci. USA 1992, 89, 9064–9068. [Google Scholar] [CrossRef] [Green Version]
- Weaver, V.M.; Petersen, O.W.; Wang, F.; Larabell, C.A.; Briand, P.; Damsky, C.; Bissell, M.J. Reversion of the Malignant Phenotype of Human Breast Cells in Three-Dimensional Culture and in Vivo by Integrin Blocking Antibodies. J. Cell Biol. 1997, 137, 231–245. [Google Scholar] [CrossRef] [Green Version]
- Hynes, R.O. Integrins: Versatility, Modulation, and Signaling in Cell Adhesion. Cell 1992, 69, 11–25. [Google Scholar] [CrossRef]
- Seguin, L.; Desgrosellier, J.S.; Weis, S.M.; Cheresh, D.A. Integrins and Cancer: Regulators of Cancer Stemness, Metastasis, and Drug Resistance. Trends Cell Biol. 2015, 25, 234–240. [Google Scholar] [CrossRef] [Green Version]
- Hamidi, H.; Pietilä, M.; Ivaska, J. The Complexity of Integrins in Cancer and New Scopes for Therapeutic Targeting. Br. J. Cancer 2016, 115, 1017–1023. [Google Scholar] [CrossRef] [Green Version]
- Cooper, J.; Giancotti, F.G. Integrin Signaling in Cancer: Mechanotransduction, Stemness, Epithelial Plasticity, and Therapeutic Resistance. Cancer Cell 2019, 35, 347–367. [Google Scholar] [CrossRef]
- Wang, F.; Weaver, V.M.; Petersen, O.W.; Larabell, C.A.; Dedhar, S.; Briand, P.; Lupu, R.; Bissell, M.J. Reciprocal Interactions between beta1-Integrin and Epidermal Growth Factor Receptor in Three-Dimensional Basement Membrane Breast Cultures: A Different Perspective in Epithelial Biology. Proc. Natl. Acad. Sci. USA 1998, 95, 14821–14826. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Radisky, D.C.; Wang, F.; Bissell, M.J. Polarity and Proliferation Are Controlled by Distinct Signaling Pathways Downstream of PI3-Kinase in Breast Epithelial Tumor Cells. J. Cell Biol. 2004, 164, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Kenny, P.A.; Bissell, M.J. Tumor Reversion: Correction of Malignant Behavior by Microenvironmental Cues. Int. J. Cancer 2003, 107, 688–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lelièvre, S.A.; Weaver, V.M.; Nickerson, J.A.; Larabell, C.A.; Bhaumik, A.; Petersen, O.W.; Bissell, M.J. Tissue Phenotype Depends on Reciprocal Interactions between the Extracellular Matrix and the Structural Organization of the Nucleus. Proc. Natl. Acad. Sci. USA 1998, 95, 14711–14716. [Google Scholar] [CrossRef] [Green Version]
- Kaminker, P.; Plachot, C.; Kim, S.-H.; Chung, P.; Crippen, D.; Petersen, O.W.; Bissell, M.J.; Campisi, J.; Lelièvre, S.A. Higher-Order Nuclear Organization in Growth Arrest of Human Mammary Epithelial Cells: A Novel Role for Telomere-Associated Protein TIN2. J. Cell Sci. 2005, 118, 1321–1330. [Google Scholar] [CrossRef] [Green Version]
- Weaver, V.M.; Lelièvre, S.; Lakins, J.N.; Chrenek, M.A.; Jones, J.C.R.; Giancotti, F.; Werb, Z.; Bissell, M.J. beta4 Integrin-Dependent Formation of Polarized Three-Dimensional Architecture Confers Resistance to Apoptosis in Normal and Malignant Mammary Epithelium. Cancer Cell 2002, 2, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Fournier, M.V.; Martin, K.J.; Kenny, P.A.; Xhaja, K.; Bosch, I.; Yaswen, P.; Bissell, M.J. Gene Expression Signature in Organized and Growth-Arrested Mammary Acini Predicts Good Outcome in Breast Cancer. Cancer Res. 2006, 66, 7095–7102. [Google Scholar] [CrossRef] [Green Version]
- Kenny, P.A.; Lee, G.Y.; Myers, C.A.; Neve, R.M.; Semeiks, J.R.; Spellman, P.T.; Lorenz, K.; Lee, E.H.; Barcellos-Hoff, M.H.; Petersen, O.W.; et al. The Morphologies of Breast Cancer Cell Lines in Three-Dimensional Assays Correlate with Their Profiles of Gene Expression. Mol. Oncol. 2007, 1, 84–96. [Google Scholar] [CrossRef]
- Rizki, A.; Weaver, V.M.; Lee, S.-Y.; Rozenberg, G.I.; Chin, K.; Myers, C.A.; Bascom, J.L.; Mott, J.D.; Semeiks, J.R.; Grate, L.R.; et al. A Human Breast Cell Model of Preinvasive to Invasive Transition. Cancer Res. 2008, 68, 1378–1387. [Google Scholar] [CrossRef] [Green Version]
- Koedoot, E.; Wolters, L.; Smid, M.; Stoilov, P.; Burger, G.A.; Herpers, B.; Yan, K.; Price, L.S.; Martens, J.W.M.; Le Dévédec, S.E.; et al. Differential Reprogramming of Breast Cancer Subtypes in 3D Cultures and Implications for Sensitivity to Targeted Therapy. Sci. Rep. 2021, 11, 7259. [Google Scholar] [CrossRef]
- Lee, T.I.; Young, R.A. Transcriptional Regulation and Its Misregulation in Disease. Cell 2013, 152, 1237–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duronio, R.J.; Xiong, Y. Signaling Pathways That Control Cell Proliferation. Cold Spring Harb. Perspect. Biol. 2013, 5, a008904. [Google Scholar] [CrossRef] [PubMed]
- Ell, B.; Kang, Y. Transcriptional Control of Cancer Metastasis. Trends Cell Biol. 2013, 23, 603–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aubrey, B.J.; Kelly, G.L.; Janic, A.; Herold, M.J.; Strasser, A. How Does p53 Induce Apoptosis and How Does This Relate to p53-Mediated Tumour Suppression? Cell Death Differ. 2018, 25, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Cakouros, D. Transcriptional Control of the Core Cell-Death Machinery. Trends Biochem. Sci. 2004, 29, 193–199. [Google Scholar] [CrossRef]
- Kohno, K.; Uchiumi, T.; Niina, I.; Wakasugi, T.; Igarashi, T.; Momii, Y.; Yoshida, T.; Matsuo, K.-I.; Miyamoto, N.; Izumi, H. Transcription Factors and Drug Resistance. Eur. J. Cancer 2005, 41, 2577–2586. [Google Scholar] [CrossRef]
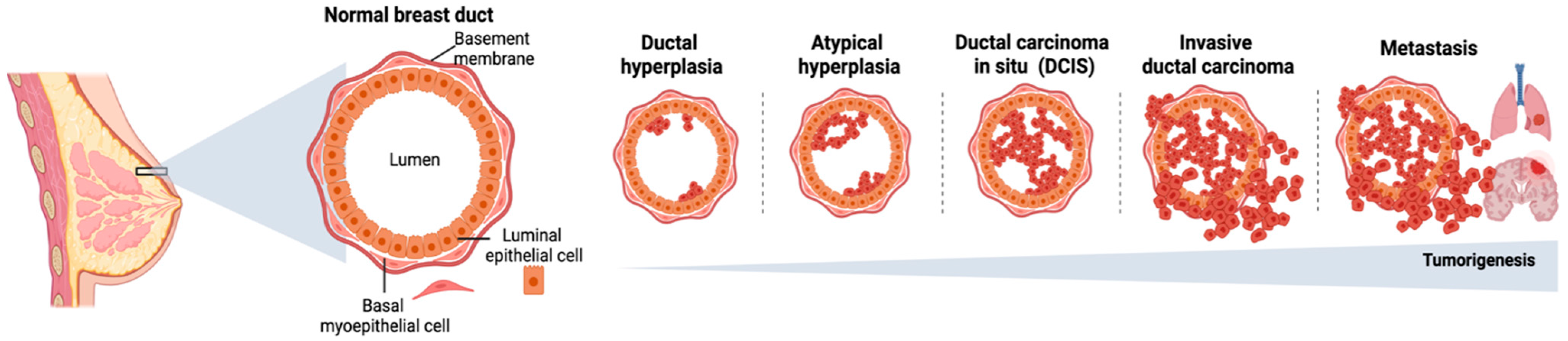
- Russo, J.; Lynch, H.; Russo, I.H. Mammary Gland Architecture as a Determining Factor in the Susceptibility of the Human Breast to Cancer. Breast J. 2001, 7, 278–291. [Google Scholar] [CrossRef]
- Hansen, R.K.; Bissell, M.J. Tissue Architecture and Breast Cancer: The Role of Extracellular Matrix and Steroid Hormones. Endocr. Relat. Cancer 2000, 7, 95–113. [Google Scholar] [CrossRef]
- Ghajar, C.M.; Bissell, M.J. Extracellular Matrix Control of Mammary Gland Morphogenesis and Tumorigenesis: Insights from Imaging. Histochem. Cell Biol. 2008, 130, 1105–1118. [Google Scholar] [CrossRef] [Green Version]
- Weaver, V.M.; Fischer, A.H.; Peterson, O.W.; Bissell, M.J. The Importance of the Microenvironment in Breast Cancer Progression: Recapitulation of Mammary Tumorigenesis Using a Unique Human Mammary Epithelial Cell Model and a Three-Dimensional Culture Assay. Biochem. Cell Biol. 1996, 74, 833–851. [Google Scholar] [CrossRef] [Green Version]
- Cichon, M.A.; Degnim, A.C.; Visscher, D.W.; Radisky, D.C. Microenvironmental Influences That Drive Progression from Benign Breast Disease to Invasive Breast Cancer. J. Mammary Gland Biol. Neoplasia 2010, 15, 389–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debnath, J.; Brugge, J.S. Modelling Glandular Epithelial Cancers in Three-Dimensional Cultures. Nat. Rev. Cancer 2005, 5, 675–688. [Google Scholar] [CrossRef] [PubMed]
- Eeles, R.A.; Bartkova, J.; Lane, D.P.; Bartek, J. The Role of TP53 in Breast Cancer Development. Cancer Surv. 1993, 18, 57–75. [Google Scholar] [PubMed]
- Kaur, R.P.; Vasudeva, K.; Kumar, R.; Munshi, A. Role of p53 Gene in Breast Cancer: Focus on Mutation Spectrum and Therapeutic Strategies. Curr. Pharm. Des. 2018, 24, 3566–3575. [Google Scholar] [CrossRef]
- Oren, M.; Rotter, V. Mutant p53 Gain-of-Function in Cancer. Cold Spring Harb. Perspect. Biol. 2010, 2, a001107. [Google Scholar] [CrossRef]
- Freed-Pastor, W.A.; Prives, C. Mutant p53: One Name, Many Proteins. Genes Dev. 2012, 26, 1268–1286. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Wang, C.; Ma, B.; Xu, M.; Xu, S.; Liu, J.; Tian, Y.; Fu, Y.; Luo, Y. Mutant p53 Drives Cancer Metastasis via RCP-Mediated Hsp90α Secretion. Cell Rep. 2020, 32, 107879. [Google Scholar] [CrossRef]
- Novo, D.; Heath, N.; Mitchell, L.; Caligiuri, G.; MacFarlane, A.; Reijmer, D.; Charlton, L.; Knight, J.; Calka, M.; McGhee, E.; et al. Mutant p53s Generate pro-Invasive Niches by Influencing Exosome Podocalyxin Levels. Nat. Commun. 2018, 9, 5069. [Google Scholar] [CrossRef] [Green Version]
- Freed-Pastor, W.A.; Mizuno, H.; Zhao, X.; Langerød, A.; Moon, S.-H.; Rodriguez-Barrueco, R.; Barsotti, A.; Chicas, A.; Li, W.; Polotskaia, A.; et al. Mutant p53 Disrupts Mammary Tissue Architecture via the Mevalonate Pathway. Cell 2012, 148, 244–258. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, B.B. Nuclear Factor-kappaB: The Enemy within. Cancer Cell 2004, 6, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Pacifico, F.; Leonardi, A. NF-kappaB in Solid Tumors. Biochem. Pharmacol. 2006, 72, 1142–1152. [Google Scholar] [CrossRef] [PubMed]
- Sethi, G.; Sung, B.; Aggarwal, B.B. Nuclear Factor-kappaB Activation: From Bench to Bedside. Exp. Biol. Med. 2008, 233, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Becker-Weimann, S.; Xiong, G.; Furuta, S.; Han, J.; Kuhn, I.; Akavia, U.-D.; Pe’er, D.; Bissell, M.J.; Xu, R. NFkB Disrupts Tissue Polarity in 3D by Preventing Integration of Microenvironmental Signals. Oncotarget 2013, 4, 2010–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruvberger, S.; Ringnér, M.; Chen, Y.; Panavally, S.; Saal, L.H.; Borg, A.; Fernö, M.; Peterson, C.; Meltzer, P.S. Estrogen Receptor Status in Breast Cancer Is Associated with Remarkably Distinct Gene Expression Patterns. Cancer Res. 2001, 61, 5979–5984. [Google Scholar] [PubMed]
- Voduc, D.; Cheang, M.; Nielsen, T. GATA-3 Expression in Breast Cancer Has a Strong Association with Estrogen Receptor but Lacks Independent Prognostic Value. Cancer Epidemiol. Biomark. Prev. 2008, 17, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Eeckhoute, J.; Keeton, E.K.; Lupien, M.; Krum, S.A.; Carroll, J.S.; Brown, M. Positive Cross-Regulatory Loop Ties GATA-3 to Estrogen Receptor Alpha Expression in Breast Cancer. Cancer Res. 2007, 67, 6477–6483. [Google Scholar] [CrossRef] [Green Version]
- Emmanuel, N.; Lofgren, K.A.; Peterson, E.A.; Meier, D.R.; Jung, E.H.; Kenny, P.A. Mutant GATA3 Actively Promotes the Growth of Normal and Malignant Mammary Cells. Anticancer Res. 2018, 38, 4435–4441. [Google Scholar] [CrossRef]
- Pfleger, C.M. The Hippo Pathway: A Master Regulatory Network Important in Development and Dysregulated in Disease. Curr. Top. Dev. Biol. 2017, 123, 181–228. [Google Scholar] [CrossRef]
- Yu, F.-X.; Zhao, B.; Guan, K.-L. Hippo Pathway in Organ Size Control, Tissue Homeostasis, and Cancer. Cell 2015, 163, 811–828. [Google Scholar] [CrossRef] [Green Version]
- Janse van Rensburg, H.J.; Lai, D.; Azad, T.; Hao, Y.; Yang, X. TAZ Enhances Mammary Cell Proliferation in 3D Culture through Transcriptional Regulation of IRS1. Cell. Signal. 2018, 52, 12–22. [Google Scholar] [CrossRef]
- Pratap, J.; Imbalzano, K.M.; Underwood, J.M.; Cohet, N.; Gokul, K.; Akech, J.; van Wijnen, A.J.; Stein, J.L.; Imbalzano, A.N.; Nickerson, J.A.; et al. Ectopic runx2 Expression in Mammary Epithelial Cells Disrupts Formation of Normal Acini Structure: Implications for Breast Cancer Progression. Cancer Res. 2009, 69, 6807–6814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anbazhagan, R.; Fujii, H.; Gabrielson, E. Allelic Loss of Chromosomal Arm 8p in Breast Cancer Progression. Am. J. Pathol. 1998, 152, 815–819. [Google Scholar] [PubMed]
- Stovall, D.B.; Wan, M.; Miller, L.D.; Cao, P.; Maglic, D.; Zhang, Q.; Stampfer, M.R.; Liu, W.; Xu, J.; Sui, G. The Regulation of SOX7 and Its Tumor Suppressive Role in Breast Cancer. Am. J. Pathol. 2013, 183, 1645–1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panayiotou, R.; Miralles, F.; Pawlowski, R.; Diring, J.; Flynn, H.R.; Skehel, M.; Treisman, R. Phosphorylation Acts Positively and Negatively to Regulate MRTF-A Subcellular Localisation and Activity. eLife 2016, 5, e15460. [Google Scholar] [CrossRef]
- Baarlink, C.; Grosse, R. Formin’ Actin in the Nucleus. Nucleus 2014, 5, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Seifert, A.; Posern, G. Tightly Controlled MRTF-A Activity Regulates Epithelial Differentiation during Formation of Mammary Acini. Breast Cancer Res. 2017, 19, 68. [Google Scholar] [CrossRef] [Green Version]
- Chaudhuri, O.; Koshy, S.T.; Branco da Cunha, C.; Shin, J.-W.; Verbeke, C.S.; Allison, K.H.; Mooney, D.J. Extracellular Matrix Stiffness and Composition Jointly Regulate the Induction of Malignant Phenotypes in Mammary Epithelium. Nat. Mater. 2014, 13, 970–978. [Google Scholar] [CrossRef]
- Uhler, C.; Shivashankar, G.V. Regulation of Genome Organization and Gene Expression by Nuclear Mechanotransduction. Nat. Rev. Mol. Cell Biol. 2017, 18, 717–727. [Google Scholar] [CrossRef]
- Danovi, S. Facing up to a Hard Truth. Nat. Rev. Cancer 2019, 19, 542–543. [Google Scholar] [CrossRef]
- Stowers, R.S.; Shcherbina, A.; Israeli, J.; Gruber, J.J.; Chang, J.; Nam, S.; Rabiee, A.; Teruel, M.N.; Snyder, M.P.; Kundaje, A.; et al. Matrix Stiffness Induces a Tumorigenic Phenotype in Mammary Epithelium through Changes in Chromatin Accessibility. Nat. Biomed. Eng. 2019, 3, 1009–1019. [Google Scholar] [CrossRef]
- Kang, Y.; Massagué, J. Epithelial-Mesenchymal Transitions: Twist in Development and Metastasis. Cell 2004, 118, 277–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhou, B.P. Epithelial-Mesenchymal Transition in Breast Cancer Progression and Metastasis. Chin. J. Cancer 2011, 30, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Weinberg, R.A. The Basics of Epithelial-Mesenchymal Transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.J.; Nieto, M.A. Epithelial-Mesenchymal Transitions in Development and Disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the Extracellular Matrix in Development and Disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef]
- Kai, F.; Drain, A.P.; Weaver, V.M. The Extracellular Matrix Modulates the Metastatic Journey. Dev. Cell 2019, 49, 332–346. [Google Scholar] [CrossRef]
- Li, Q.; Chen, C.; Kapadia, A.; Zhou, Q.; Harper, M.K.; Schaack, J.; LaBarbera, D.V. 3D Models of Epithelial-Mesenchymal Transition in Breast Cancer Metastasis: High-Throughput Screening Assay Development, Validation, and Pilot Screen. J. Biomol. Screen. 2011, 16, 141–154. [Google Scholar] [CrossRef] [Green Version]
- Bidarra, S.J.; Oliveira, P.; Rocha, S.; Saraiva, D.P.; Oliveira, C.; Barrias, C.C. A 3D in Vitro Model to Explore the Inter-Conversion between Epithelial and Mesenchymal States during EMT and Its Reversion. Sci. Rep. 2016, 6, 27072. [Google Scholar] [CrossRef]
- Zhang, Y.-Q.; Wei, X.-L.; Liang, Y.-K.; Chen, W.-L.; Zhang, F.; Bai, J.-W.; Qiu, S.-Q.; Du, C.-W.; Huang, W.-H.; Zhang, G.-J. Over-Expressed Twist Associates with Markers of Epithelial Mesenchymal Transition and Predicts Poor Prognosis in Breast Cancers via ERK and Akt Activation. PLoS ONE 2015, 10, e0135851. [Google Scholar] [CrossRef]
- Eckert, M.A.; Lwin, T.M.; Chang, A.T.; Kim, J.; Danis, E.; Ohno-Machado, L.; Yang, J. Twist1-Induced Invadopodia Formation Promotes Tumor Metastasis. Cancer Cell 2011, 19, 372–386. [Google Scholar] [CrossRef] [Green Version]
- Narita, D.; Seclaman, E.; Ursoniu, S.; Anghel, A. Increased Expression of ADAM12 and ADAM17 Genes in Laser-Capture Microdissected Breast Cancers and Correlations with Clinical and Pathological Characteristics. Acta Histochem. 2012, 114, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Duhachek-Muggy, S.; Qi, Y.; Hong, Y.; Behbod, F.; Zolkiewska, A. An Essential Role of Metalloprotease-Disintegrin ADAM12 in Triple-Negative Breast Cancer. Breast Cancer Res. Treat. 2012, 135, 759–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckert, M.A.; Santiago-Medina, M.; Lwin, T.M.; Kim, J.; Courtneidge, S.A.; Yang, J. ADAM12 Induction by Twist1 Promotes Tumor Invasion and Metastasis via Regulation of Invadopodia and Focal Adhesions. J. Cell Sci. 2017, 130, 2036–2048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Mani, S.A.; Donaher, J.L.; Ramaswamy, S.; Itzykson, R.A.; Come, C.; Savagner, P.; Gitelman, I.; Richardson, A.; Weinberg, R.A. Twist, a Master Regulator of Morphogenesis, Plays an Essential Role in Tumor Metastasis. Cell 2004, 117, 927–939. [Google Scholar] [CrossRef] [Green Version]
- Franz-Wachtel, M.; Eisler, S.A.; Krug, K.; Wahl, S.; Carpy, A.; Nordheim, A.; Pfizenmaier, K.; Hausser, A.; Macek, B. Global Detection of Protein Kinase D-Dependent Phosphorylation Events in Nocodazole-Treated Human Cells. Mol. Cell. Proteom. 2012, 11, 160–170. [Google Scholar] [CrossRef] [Green Version]
- Georgess, D.; Padmanaban, V.; Sirka, O.K.; Coutinho, K.; Choi, A.; Frid, G.; Neumann, N.M.; Inoue, T.; Ewald, A.J. Twist1-Induced Epithelial Dissemination Requires Prkd1 Signaling. Cancer Res. 2020, 80, 204–218. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.; Liu, Z.; Chen, H.; Xu, J. The Steroid Receptor Coactivator-1 Regulates Twist Expression and Promotes Breast Cancer Metastasis. Cancer Res. 2009, 69, 3819–3827. [Google Scholar] [CrossRef] [Green Version]
- Mittal, V. Epithelial Mesenchymal Transition in Tumor Metastasis. Annu. Rev. Pathol. 2018, 13, 395–412. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, J.; Chai, K.; Ying, X.; Zhou, B.P. The Role of Snail in EMT and Tumorigenesis. Curr. Cancer Drug Targets 2013, 13, 963–972. [Google Scholar] [CrossRef]
- Vistain, L.F.; Yamamoto, N.; Rathore, R.; Cha, P.; Meade, T.J. Targeted Inhibition of Snail Activity in Breast Cancer Cells by Using a Co(III) -Ebox Conjugate. ChemBioChem 2015, 16, 2065–2072. [Google Scholar] [CrossRef] [Green Version]
- Henry, W.S.; Hendrickson, D.G.; Beca, F.; Glass, B.; Lindahl-Allen, M.; He, L.; Ji, Z.; Struhl, K.; Beck, A.H.; Rinn, J.L.; et al. LINC00520 Is Induced by Src, STAT3, and PI3K and Plays a Functional Role in Breast Cancer. Oncotarget 2016, 7, 81981–81994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katz, E.; Sims, A.H.; Sproul, D.; Caldwell, H.; Dixon, M.J.; Meehan, R.R.; Harrison, D.J. Targeting of Rac GTPases Blocks the Spread of Intact Human Breast Cancer. Oncotarget 2012, 3, 608–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koster, M.I.; Kim, S.; Mills, A.A.; DeMayo, F.J.; Roop, D.R. p63 Is the Molecular Switch for Initiation of an Epithelial Stratification Program. Genes Dev. 2004, 18, 126–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, A.; Kaghad, M.; Wang, Y.; Gillett, E.; Fleming, M.D.; Dötsch, V.; Andrews, N.C.; Caput, D.; McKeon, F. p63, a p53 Homolog at 3q27-29, Encodes Multiple Products with Transactivating, Death-Inducing, and Dominant-Negative Activities. Mol. Cell 1998, 2, 305–316. [Google Scholar] [CrossRef]
- Lindsay, J.; McDade, S.S.; Pickard, A.; McCloskey, K.D.; McCance, D.J. Role of DeltaNp63gamma in Epithelial to Mesenchymal Transition. J. Biol. Chem. 2011, 286, 3915–3924. [Google Scholar] [CrossRef] [Green Version]
- Yoh, K.E.; Regunath, K.; Guzman, A.; Lee, S.-M.; Pfister, N.T.; Akanni, O.; Kaufman, L.J.; Prives, C.; Prywes, R. Repression of p63 and Induction of EMT by Mutant Ras in Mammary Epithelial Cells. Proc. Natl. Acad. Sci. USA 2016, 113, E6107–E6116. [Google Scholar] [CrossRef] [Green Version]
- Gramolelli, S.; Cheng, J.; Martinez-Corral, I.; Vähä-Koskela, M.; Elbasani, E.; Kaivanto, E.; Rantanen, V.; Tuohinto, K.; Hautaniemi, S.; Bower, M.; et al. PROX1 Is a Transcriptional Regulator of MMP14. Sci. Rep. 2018, 8, 9531. [Google Scholar] [CrossRef]
- Wright, R.H.G.; Lioutas, A.; Le Dily, F.; Soronellas, D.; Pohl, A.; Bonet, J.; Nacht, A.S.; Samino, S.; Font-Mateu, J.; Vicent, G.P.; et al. ADP-Ribose-Derived Nuclear ATP Synthesis by NUDIX5 Is Required for Chromatin Remodeling. Science 2016, 352, 1221–1225. [Google Scholar] [CrossRef]
- Page, B.D.G.; Valerie, N.C.K.; Wright, R.H.G.; Wallner, O.; Isaksson, R.; Carter, M.; Rudd, S.G.; Loseva, O.; Jemth, A.-S.; Almlöf, I.; et al. Targeted NUDT5 Inhibitors Block Hormone Signaling in Breast Cancer Cells. Nat. Commun. 2018, 9, 250. [Google Scholar] [CrossRef] [Green Version]
- Pickup, K.E.; Pardow, F.; Carbonell-Caballero, J.; Lioutas, A.; Villanueva-Cañas, J.L.; Wright, R.H.G.; Beato, M. Expression of Oncogenic Drivers in 3D Cell Culture Depends on Nuclear ATP Synthesis by NUDT5. Cancers 2019, 11, 1337. [Google Scholar] [CrossRef] [Green Version]
- Gaboli, M.; Kotsi, P.A.; Gurrieri, C.; Cattoretti, G.; Ronchetti, S.; Cordon-Cardo, C.; Broxmeyer, H.E.; Hromas, R.; Pandolfi, P.P. Mzf1 Controls Cell Proliferation and Tumorigenesis. Genes Dev. 2001, 15, 1625–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beishline, K.; Azizkhan-Clifford, J. Sp1 and the “Hallmarks of Cancer”. FEBS J. 2015, 282, 224–258. [Google Scholar] [CrossRef] [PubMed]
- Hahn, S.; Hermeking, H. ZNF281/ZBP-99: A New Player in Epithelial-Mesenchymal Transition, Stemness, and Cancer. J. Mol. Med. 2014, 92, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Burkhart, D.L.; Sage, J. Cellular Mechanisms of Tumour Suppression by the Retinoblastoma Gene. Nat. Rev. Cancer 2008, 8, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Egger, J.V.; Lane, M.V.; Antonucci, L.A.; Dedi, B.; Krucher, N.A. Dephosphorylation of the Retinoblastoma Protein (Rb) Inhibits Cancer Cell EMT via Zeb. Cancer Biol. Ther. 2016, 17, 1197–1205. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Elenbaas, B.; Weinberg, R.A. Heterotypic Signaling between Epithelial Tumor Cells and Fibroblasts in Carcinoma Formation. Exp. Cell Res. 2001, 264, 169–184. [Google Scholar] [CrossRef]
- Buess, M.; Nuyten, D.S.A.; Hastie, T.; Nielsen, T.; Pesich, R.; Brown, P.O. Characterization of Heterotypic Interaction Effects in Vitro to Deconvolute Global Gene Expression Profiles in Cancer. Genome Biol. 2007, 8, R191. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.; Wang, L.; Liu, S.; Wang, Y.; Liu, Y.; Wang, M.; Nakshatri, H.; Li, B.-Y.; Yokota, H. Attraction and Compaction of Migratory Breast Cancer Cells by Bone Matrix Proteins through Tumor-Osteocyte Interactions. Sci. Rep. 2018, 8, 5420. [Google Scholar] [CrossRef] [Green Version]
- Pang, M.-F.; Georgoudaki, A.-M.; Lambut, L.; Johansson, J.; Tabor, V.; Hagikura, K.; Jin, Y.; Jansson, M.; Alexander, J.S.; Nelson, C.M.; et al. TGF-β1-Induced EMT Promotes Targeted Migration of Breast Cancer Cells through the Lymphatic System by the Activation of CCR7/CCL21-Mediated Chemotaxis. Oncogene 2016, 35, 748–760. [Google Scholar] [CrossRef]
- Bissell, M.J.; Radisky, D. Putting Tumours in Context. Nat. Rev. Cancer 2001, 1, 46–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reginato, M.J.; Mills, K.R.; Becker, E.B.E.; Lynch, D.K.; Bonni, A.; Muthuswamy, S.K.; Brugge, J.S. Bim Regulation of Lumen Formation in Cultured Mammary Epithelial Acini Is Targeted by Oncogenes. Mol. Cell. Biol. 2005, 25, 4591–4601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muthuswamy, S.K.; Li, D.; Lelievre, S.; Bissell, M.J.; Brugge, J.S. ErbB2, but Not ErbB1, Reinitiates Proliferation and Induces Luminal Repopulation in Epithelial Acini. Nat. Cell Biol. 2001, 3, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Liu, Z.; Myers, D.P.; Terada, L.S. Mechanotransduction and Anoikis: Death and the Homeless Cell. Cell Cycle 2008, 7, 2462–2465. [Google Scholar] [CrossRef] [Green Version]
- Frisch, S.M.; Francis, H. Disruption of Epithelial Cell-Matrix Interactions Induces Apoptosis. J. Cell Biol. 1994, 124, 619–626. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-N.; Koo, K.H.; Sung, J.Y.; Yun, U.-J.; Kim, H. Anoikis Resistance: An Essential Prerequisite for Tumor Metastasis. Int. J. Cell Biol. 2012, 2012, 306879. [Google Scholar] [CrossRef] [Green Version]
- Simpson, C.D.; Anyiwe, K.; Schimmer, A.D. Anoikis Resistance and Tumor Metastasis. Cancer Lett. 2008, 272, 177–185. [Google Scholar] [CrossRef]
- Adeshakin, F.O.; Adeshakin, A.O.; Afolabi, L.O.; Yan, D.; Zhang, G.; Wan, X. Mechanisms for Modulating Anoikis Resistance in Cancer and the Relevance of Metabolic Reprogramming. Front. Oncol. 2021, 11, 626577. [Google Scholar] [CrossRef]
- Akekawatchai, C.; Roytrakul, S.; Phaonakrop, N.; Jaresitthikunchai, J.; Jitrapakdee, S. Proteomic Analysis of the Anoikis-Resistant Human Breast Cancer Cell Lines. Methods Mol. Biol. 2020, 2138, 185–193. [Google Scholar] [CrossRef]
- Akekawatchai, C.; Roytrakul, S.; Kittisenachai, S.; Isarankura-Na-Ayudhya, P.; Jitrapakdee, S. Protein Profiles Associated with Anoikis Resistance of Metastatic MDA-MB-231 Breast Cancer Cells. Asian Pac. J. Cancer Prev. 2016, 17, 581–590. [Google Scholar] [CrossRef] [Green Version]
- Buchheit, C.L.; Weigel, K.J.; Schafer, Z.T. Cancer Cell Survival during Detachment from the ECM: Multiple Barriers to Tumour Progression. Nat. Rev. Cancer 2014, 14, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Frisch, S.M.; Screaton, R.A. Anoikis Mechanisms. Curr. Opin. Cell Biol. 2001, 13, 555–562. [Google Scholar] [CrossRef]
- Khan, I.A.; Yoo, B.H.; McPhee, M.; Masson, O.; Surette, A.; Dakin-Hache, K.; Younis, T.; Bethune, G.; Rosen, K.V. ErbB2-Driven Downregulation of the Transcription Factor Irf6 in Breast Epithelial Cells Is Required for Their 3D Growth. Breast Cancer Res. 2018, 20, 151. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.F.; Rao, S.S. Three-Dimensional Culture Models to Study Drug Resistance in Breast Cancer. Biotechnol. Bioeng. 2020, 117, 2262–2278. [Google Scholar] [CrossRef]
- Ding, Y.; Liu, W.; Yu, W.; Lu, S.; Liu, M.; Kaplan, D.L.; Wang, X. Three-Dimensional Tissue Culture Model of Human Breast Cancer for the Evaluation of Multidrug Resistance. J. Tissue Eng. Regen. Med. 2018, 12, 1959–1971. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.-Z.; Jin, W.-L. The Updated Landscape of Tumor Microenvironment and Drug Repurposing. Signal Transduct. Target. Ther. 2020, 5, 166. [Google Scholar] [CrossRef] [PubMed]
- Meads, M.B.; Gatenby, R.A.; Dalton, W.S. Environment-Mediated Drug Resistance: A Major Contributor to Minimal Residual Disease. Nat. Rev. Cancer 2009, 9, 665–674. [Google Scholar] [CrossRef]
- Picco, N.; Sahai, E.; Maini, P.K.; Anderson, A.R.A. Integrating Models to Quantify Environment-Mediated Drug Resistance. Cancer Res. 2017, 77, 5409–5418. [Google Scholar] [CrossRef] [Green Version]
- Imamura, Y.; Mukohara, T.; Shimono, Y.; Funakoshi, Y.; Chayahara, N.; Toyoda, M.; Kiyota, N.; Takao, S.; Kono, S.; Nakatsura, T.; et al. Comparison of 2D- and 3D-Culture Models as Drug-Testing Platforms in Breast Cancer. Oncol. Rep. 2015, 33, 1837–1843. [Google Scholar] [CrossRef] [Green Version]
- Kitaeva, K.V.; Rutland, C.S.; Rizvanov, A.A.; Solovyeva, V.V. Cell Culture Based Test Systems for Anticancer Drug Screening. Front. Bioeng. Biotechnol. 2020, 8, 322. [Google Scholar] [CrossRef] [Green Version]
- Dhiman, H.K.; Ray, A.R.; Panda, A.K. Three-Dimensional Chitosan Scaffold-Based MCF-7 Cell Culture for the Determination of the Cytotoxicity of Tamoxifen. Biomaterials 2005, 26, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Horning, J.L.; Sahoo, S.K.; Vijayaraghavalu, S.; Dimitrijevic, S.; Vasir, J.K.; Jain, T.K.; Panda, A.K.; Labhasetwar, V. 3-D Tumor Model for in Vitro Evaluation of Anticancer Drugs. Mol. Pharm. 2008, 5, 849–862. [Google Scholar] [CrossRef]
- Lovitt, C.J.; Shelper, T.B.; Avery, V.M. Doxorubicin Resistance in Breast Cancer Cells Is Mediated by Extracellular Matrix Proteins. BMC Cancer 2018, 18, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickl, M.; Ries, C.H. Comparison of 3D and 2D Tumor Models Reveals Enhanced HER2 Activation in 3D Associated with an Increased Response to Trastuzumab. Oncogene 2009, 28, 461–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirschhaeuser, F.; Menne, H.; Dittfeld, C.; West, J.; Mueller-Klieser, W.; Kunz-Schughart, L.A. Multicellular Tumor Spheroids: An Underestimated Tool Is Catching up Again. J. Biotechnol. 2010, 148, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Freyer, J.P.; Sutherland, R.M. Regulation of Growth Saturation and Development of Necrosis in EMT6/Ro Multicellular Spheroids by the Glucose and Oxygen Supply. Cancer Res. 1986, 46, 3504–3512. [Google Scholar] [PubMed]
- Mueller-Klieser, W. Multicellular Spheroids. A Review on Cellular Aggregates in Cancer Research. J. Cancer Res. Clin. Oncol. 1987, 113, 101–122. [Google Scholar] [CrossRef] [PubMed]
- Däster, S.; Amatruda, N.; Calabrese, D.; Ivanek, R.; Turrini, E.; Droeser, R.A.; Zajac, P.; Fimognari, C.; Spagnoli, G.C.; Iezzi, G.; et al. Induction of Hypoxia and Necrosis in Multicellular Tumor Spheroids Is Associated with Resistance to Chemotherapy Treatment. Oncotarget 2017, 8, 1725–1736. [Google Scholar] [CrossRef] [Green Version]
- Vidi, P.-A.; Bissell, M.J.; Lelièvre, S.A. Three-Dimensional Culture of Human Breast Epithelial Cells: The How and the Why. Methods Mol. Biol. 2013, 945, 193–219. [Google Scholar] [CrossRef] [Green Version]
- Djomehri, S.I.; Burman, B.; Gonzalez, M.E.; Takayama, S.; Kleer, C.G. A Reproducible Scaffold-Free 3D Organoid Model to Study Neoplastic Progression in Breast Cancer. J. Cell Commun. Signal. 2019, 13, 129–143. [Google Scholar] [CrossRef]
- Semenza, G.L. HIF-1 Mediates Metabolic Responses to Intratumoral Hypoxia and Oncogenic Mutations. J. Clin. Investig. 2013, 123, 3664–3671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohwer, N.; Cramer, T. Hypoxia-Mediated Drug Resistance: Novel Insights on the Functional Interaction of HIFs and Cell Death Pathways. Drug Resist. Updat. 2011, 14, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Doublier, S.; Belisario, D.C.; Polimeni, M.; Annaratone, L.; Riganti, C.; Allia, E.; Ghigo, D.; Bosia, A.; Sapino, A. HIF-1 Activation Induces Doxorubicin Resistance in MCF7 3-D Spheroids via P-Glycoprotein Expression: A Potential Model of the Chemo-Resistance of Invasive Micropapillary Carcinoma of the Breast. BMC Cancer 2012, 12, 4. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.Y.; Cusack, J.C., Jr.; Liu, R.; Baldwin, A.S., Jr. Control of Inducible Chemoresistance: Enhanced Anti-Tumor Therapy through Increased Apoptosis by Inhibition of NF-kappaB. Nat. Med. 1999, 5, 412–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karin, M. Nuclear Factor-kappaB in Cancer Development and Progression. Nature 2006, 441, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, K.M.; Zhang, H.; Park, C.C. NF-κB Regulates Radioresistance Mediated by β1-Integrin in Three-Dimensional Culture of Breast Cancer Cells. Cancer Res. 2013, 73, 3737–3748. [Google Scholar] [CrossRef] [Green Version]
- Mann, A.P.; Verma, A.; Sethi, G.; Manavathi, B.; Wang, H.; Fok, J.Y.; Kunnumakkara, A.B.; Kumar, R.; Aggarwal, B.B.; Mehta, K. Overexpression of Tissue Transglutaminase Leads to Constitutive Activation of Nuclear Factor-kappaB in Cancer Cells: Delineation of a Novel Pathway. Cancer Res. 2006, 66, 8788–8795. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Xu, A.-M.; Liu, W. Transglutaminase 2 in Cancer. Am. J. Cancer Res. 2015, 5, 2756–2776. [Google Scholar]
- Kumar, A.; Xu, J.; Brady, S.; Gao, H.; Yu, D.; Reuben, J.; Mehta, K. Tissue Transglutaminase Promotes Drug Resistance and Invasion by Inducing Mesenchymal Transition in Mammary Epithelial Cells. PLoS ONE 2010, 5, e13390. [Google Scholar] [CrossRef] [Green Version]
- Bihani, T.; Ezell, S.A.; Ladd, B.; Grosskurth, S.E.; Mazzola, A.M.; Pietras, M.; Reimer, C.; Zinda, M.; Fawell, S.; D’Cruz, C.M. Resistance to Everolimus Driven by Epigenetic Regulation of MYC in ER+ Breast Cancers. Oncotarget 2015, 6, 2407–2420. [Google Scholar] [CrossRef] [Green Version]
- White, M.E.; Fenger, J.M.; Carson, W.E., 3rd. Emerging Roles of and Therapeutic Strategies Targeting BRD4 in Cancer. Cell. Immunol. 2019, 337, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Hart, T.; Brown, K.R.; Sircoulomb, F.; Rottapel, R.; Moffat, J. Measuring Error Rates in Genomic Perturbation Screens: Gold Standards for Human Functional Genomics. Mol. Syst. Biol. 2014, 10, 733. [Google Scholar] [CrossRef] [PubMed]
- Marcotte, R.; Brown, K.R.; Suarez, F.; Sayad, A.; Karamboulas, K.; Krzyzanowski, P.M.; Sircoulomb, F.; Medrano, M.; Fedyshyn, Y.; Koh, J.L.Y.; et al. Essential Gene Profiles in Breast, Pancreatic, and Ovarian Cancer Cells. Cancer Discov. 2012, 2, 172–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashima, T.; Seimiya, H.; Tsuruo, T. De Novo Fatty-Acid Synthesis and Related Pathways as Molecular Targets for Cancer Therapy. Br. J. Cancer 2009, 100, 1369–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sethunath, V.; Hu, H.; De Angelis, C.; Veeraraghavan, J.; Qin, L.; Wang, N.; Simon, L.M.; Wang, T.; Fu, X.; Nardone, A.; et al. Targeting the Mevalonate Pathway to Overcome Acquired Anti-HER2 Treatment Resistance in Breast Cancer. Mol. Cancer Res. 2019, 17, 2318–2330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bathaie, S.Z.; Ashrafi, M.; Azizian, M.; Tamanoi, F. Mevalonate Pathway and Human Cancers. Curr. Mol. Pharmacol. 2017, 10, 77–85. [Google Scholar] [CrossRef]
- Pandyra, A.A.; Mullen, P.J.; Goard, C.A.; Ericson, E.; Sharma, P.; Kalkat, M.; Yu, R.; Pong, J.T.; Brown, K.R.; Hart, T.; et al. Genome-Wide RNAi Analysis Reveals That Simultaneous Inhibition of Specific Mevalonate Pathway Genes Potentiates Tumor Cell Death. Oncotarget 2015, 6, 26909–26921. [Google Scholar] [CrossRef] [Green Version]
- Weiss, M.S.; Peñalver Bernabé, B.; Shin, S.; Asztalos, S.; Dubbury, S.J.; Mui, M.D.; Bellis, A.D.; Bluver, D.; Tonetti, D.A.; Saez-Rodriguez, J.; et al. Dynamic Transcription Factor Activity and Networks during ErbB2 Breast Oncogenesis and Targeted Therapy. Integr. Biol. 2014, 6, 1170–1182. [Google Scholar] [CrossRef] [Green Version]
- Crabtree, J.S.; Miele, L. Breast Cancer Stem Cells. Biomedicines 2018, 6, 77. [Google Scholar] [CrossRef] [Green Version]
- Korkaya, H.; Liu, S.; Wicha, M.S. Breast Cancer Stem Cells, Cytokine Networks, and the Tumor Microenvironment. J. Clin. Investig. 2011, 121, 3804–3809. [Google Scholar] [CrossRef]
- Plaks, V.; Kong, N.; Werb, Z. The Cancer Stem Cell Niche: How Essential Is the Niche in Regulating Stemness of Tumor Cells? Cell Stem Cell 2015, 16, 225–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Xiong, L.; Li, Q.; Lin, L.; Miao, X.; Yan, S.; Hong, Z.; Yang, L.; Wen, Y.; Deng, X. 3D Modeling of Cancer Stem Cell Niche. Oncotarget 2018, 9, 1326–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.-M.; Sun, W.; Wang, Z.-H.; Liang, X.; Hua, F.; Li, K.; Lv, X.-X.; Zhang, X.-W.; Liu, Y.-Y.; Yu, J.-J.; et al. TRIB3 Supports Breast Cancer Stemness by Suppressing FOXO1 Degradation and Enhancing SOX2 Transcription. Nat. Commun. 2019, 10, 5720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wennemers, M.; Bussink, J.; Scheijen, B.; Nagtegaal, I.D.; van Laarhoven, H.W.M.; Raleigh, J.A.; Varia, M.A.; Heuvel, J.J.T.M.; Rouschop, K.M.; Sweep, F.C.G.J.; et al. Tribbles Homolog 3 Denotes a Poor Prognosis in Breast Cancer and Is Involved in Hypoxia Response. Breast Cancer Res. 2011, 13, R82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, W.; Huang, X.; Chen, C.; Gray, J.; Huang, T. TBX3 and Its Isoform TBX3+2a Are Functionally Distinctive in Inhibition of Senescence and Are Overexpressed in a Subset of Breast Cancer Cell Lines. Cancer Res. 2004, 64, 5132–5139. [Google Scholar] [CrossRef] [PubMed]
- Yarosh, W.; Barrientos, T.; Esmailpour, T.; Lin, L.; Carpenter, P.M.; Osann, K.; Anton-Culver, H.; Huang, T. TBX3 Is Overexpressed in Breast Cancer and Represses p14 ARF by Interacting with Histone Deacetylases. Cancer Res. 2008, 68, 693–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Tian, Y.; Li, J.; Lu, B.; Sun, M.; Zou, Y.; Kong, R.; Luo, Y.; Shi, Y.; Wang, K.; et al. miR-206 Is down-Regulated in Breast Cancer and Inhibits Cell Proliferation through the up-Regulation of cyclinD2. Biochem. Biophys. Res. Commun. 2013, 433, 207–212. [Google Scholar] [CrossRef]
- Amir, S.; Simion, C.; Umeh-Garcia, M.; Krig, S.; Moss, T.; Carraway, K.L., 3rd; Sweeney, C. Regulation of the T-Box Transcription Factor Tbx3 by the Tumour Suppressor microRNA-206 in Breast Cancer. Br. J. Cancer 2016, 114, 1125–1134. [Google Scholar] [CrossRef] [Green Version]
- Yousefnia, S.; Ghaedi, K.; Seyed Forootan, F.; Nasr Esfahani, M.H. Characterization of the Stemness Potency of Isolated from the Breast Cancer Cell Lines. Tumour Biol. 2019, 41, 1010428319869101. [Google Scholar] [CrossRef] [Green Version]
- Maric, G.; Annis, M.G.; MacDonald, P.A.; Russo, C.; Perkins, D.; Siwak, D.R.; Mills, G.B.; Siegel, P.M. GPNMB Augments Wnt-1 Mediated Breast Tumor Initiation and Growth by Enhancing PI3K/AKT/mTOR Pathway Signaling and β-Catenin Activity. Oncogene 2019, 38, 5294–5307. [Google Scholar] [CrossRef]
- Chen, C.; Okita, Y.; Watanabe, Y.; Abe, F.; Fikry, M.A.; Ichikawa, Y.; Suzuki, H.; Shibuya, A.; Kato, M. Glycoprotein Nmb Is Exposed on the Surface of Dormant Breast Cancer Cells and Induces Stem Cell-like Properties. Cancer Res. 2018, 78, 6424–6435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaicharoenaudomrung, N.; Kunhorm, P.; Noisa, P. Three-Dimensional Cell Culture Systems as an Platform for Cancer and Stem Cell Modeling. World J. Stem Cells 2019, 11, 1065–1083. [Google Scholar] [CrossRef] [PubMed]
- Jensen, C.; Teng, Y. Is It Time to Start Transitioning From 2D to 3D Cell Culture? Front. Mol. Biosci. 2020, 7, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolas, J.; Magli, S.; Rabbachin, L.; Sampaolesi, S.; Nicotra, F.; Russo, L. 3D Extracellular Matrix Mimics: Fundamental Concepts and Role of Materials Chemistry to Influence Stem Cell Fate. Biomacromolecules 2020, 21, 1968–1994. [Google Scholar] [CrossRef] [PubMed]
- Burdett, E.; Kasper, F.K.; Mikos, A.G.; Ludwig, J.A. Engineering Tumors: A Tissue Engineering Perspective in Cancer Biology. Tissue Eng. Part B Rev. 2010, 16, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Nii, T.; Makino, K.; Tabata, Y. Three-Dimensional Culture System of Cancer Cells Combined with Biomaterials for Drug Screening. Cancers 2020, 12, 2754. [Google Scholar] [CrossRef]
- Liu, C.; Lewin Mejia, D.; Chiang, B.; Luker, K.E.; Luker, G.D. Hybrid Collagen Alginate Hydrogel as a Platform for 3D Tumor Spheroid Invasion. Acta Biomater. 2018, 75, 213–225. [Google Scholar] [CrossRef]
- Nii, T.; Kuwahara, T.; Makino, K.; Tabata, Y. A Co-Culture System of Three-Dimensional Tumor-Associated Macrophages and Three-Dimensional Cancer-Associated Fibroblasts Combined with Biomolecule Release for Cancer Cell Migration. Tissue Eng. Part A 2020, 26, 1272–1282. [Google Scholar] [CrossRef]
- Pinto, M.P.; Dye, W.W.; Jacobsen, B.M.; Horwitz, K.B. Malignant Stroma Increases Luminal Breast Cancer Cell Proliferation and Angiogenesis through Platelet-Derived Growth Factor Signaling. BMC Cancer 2014, 14, 735. [Google Scholar] [CrossRef] [Green Version]
- Bersini, S.; Jeon, J.S.; Dubini, G.; Arrigoni, C.; Chung, S.; Charest, J.L.; Moretti, M.; Kamm, R.D. A Microfluidic 3D in Vitro Model for Specificity of Breast Cancer Metastasis to Bone. Biomaterials 2014, 35, 2454–2461. [Google Scholar] [CrossRef]
- Schutgens, F.; Clevers, H. Human Organoids: Tools for Understanding Biology and Treating Diseases. Annu. Rev. Pathol. 2020, 15, 211–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachs, N.; de Ligt, J.; Kopper, O.; Gogola, E.; Bounova, G.; Weeber, F.; Balgobind, A.V.; Wind, K.; Gracanin, A.; Begthel, H.; et al. A Living Biobank of Breast Cancer Organoids Captures Disease Heterogeneity. Cell 2018, 172, 373–386.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekkers, J.F.; Whittle, J.R.; Vaillant, F.; Chen, H.-R.; Dawson, C.; Liu, K.; Geurts, M.H.; Herold, M.J.; Clevers, H.; Lindeman, G.J.; et al. Modeling Breast Cancer Using CRISPR-Cas9-Mediated Engineering of Human Breast Organoids. J. Natl. Cancer Inst. 2020, 112, 540–544. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Koo, B.-K.; Knoblich, J.A. Human Organoids: Model Systems for Human Biology and Medicine. Nat. Rev. Mol. Cell Biol. 2020, 21, 571–584. [Google Scholar] [CrossRef]
- Pérez-González, C.; Ceada, G.; Greco, F.; Matejčić, M.; Gómez-González, M.; Castro, N.; Menendez, A.; Kale, S.; Krndija, D.; Clark, A.G.; et al. Mechanical Compartmentalization of the Intestinal Organoid Enables Crypt Folding and Collective Cell Migration. Nat. Cell Biol. 2021, 23, 745–757. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristics | 2D Culture | 3D Culture | References |
|---|---|---|---|
| Cellular morphology | Cells cultured as monolayers; changed morphology; loss of epithelial cell polarity and diverse phenotype | Cells cultured in 3D aggregates, with preserved original morphology and polarization of cells; diverse phenotype | [33] |
| Proliferation | Often proliferate more rapidly than in vivo | Depends on 3D matrix interactions | [34] |
| Differentiation | Poorly or moderately differentiated | Well differentiated | [35] |
| Cell interactions | Limited cell–cell and cell–microenvironment interactions | Cell junctions enable cell communication | [36,37] |
| Gene expression | Cell adhesion-, proliferation-, and survival-related genes are usually modified | Similar gene expression profiles to those in vivo | [38] |
| Drug sensitivity | More sensitive to drugs, high efficacy Drugs are not well metabolized | More resistant to drugs, low potency Enhanced drug metabolism | [39] |
| Response to stimuli | Limited representation of the response to mechanical and biochemical cues | Accurate representation of in vivo stimuli | [40,41] |
| Cell exposure to medium | Equal access to nutrients and growth factors | Differential nutrient and growth factor availability, mimicking hypoxic conditions, very similar to in vivo tissues | [42] |
| Cost | Inexpensive, suitable for large-scale studies, commercially available materials | More expensive than 2D culture, time consuming | [43] |
| Quality | High performance, reproducible, easily interpretable, suitable for long-term culture | Worse performance and reproducibility, difficult to interpret, difficult to maintain long-term culture | [43] |
| Transcriptional Regulation of | TFs | Target Gene | Proposed Function in breast Carcinogenesis | Cell Lines Used for 3D Culture | Type of ECM | References |
|---|---|---|---|---|---|---|
| Mammary tissue architecture | P53 | SREBP-1 and SREBP-2 | Disruption of normal acinar morphology via mutant p53 | MDA-468, MDA-231, and MCF10A | rBM matrix | [79] |
| NF-κB | Disorganization genes | Disruption of tissue polarity and organization | HMT-3522 S1 and T4-2 cells | rBM matrix | [83] | |
| GATA3 | Induces proliferation via mutant GATA3s | ZR751 | Xenograft | [87] | ||
| TAZ | IRS1 | Induces proliferation | MCF10A | rBM matrix | [90] | |
| SOX7 | SOX7 depletion causes loss of polarity and increased proliferation | MCF10A | rBM matrix | [93] | ||
| TBX3 | Branching morphology | MDA-MB-231 | rBM matrix | [198] | ||
| MRTF-A and MRTF-B | integrin α5, integrin α6 | Disruption of normal acinar morphology | MCF10A | rBM matrix | [96] | |
| Sp1 | Stiffness-induced tumorigenic phenotype and chromatin organization | MCF7 and MDA-MB-231 | rBM matrix | [100] | ||
| EMT and metastasis | TWIST1 | ADAM12 | Induces tumor invasion and metastasis | Hs578T and HMLE cells | Matrigel–collagen mixture | [113] |
| TWIST1 | PRKD1 | Promotes invasion, epithelial dissemination, and metastasis | murine and human mammary tumor tissues | Organoid | [116] | |
| SNAIL | SNAIL inhibition reduces invasion and metastasis | MCF7 | [120] | |||
| SNAIL | Downregulation of Snail induces a reversal of EMT in the bone | MDA-MB-231, mammary tumor cells, osteoblast-like cells | rBM matrix | [139] | ||
| STAT3 | LINC00520 | Invasion and metastasis | MCF10A-Src transformed cells | rBM matrix, bioprinting | [121] | |
| STAT3 | STAT3 inhibition blocks invasion | primary tumor tissue explant culture | rBM matrix | [122] | ||
| ∆Np63α | ∆Np63α inhibition induces EMT | MCF10A | 3D collagen gel | [126] | ||
| OCT1-SOX2, PAX6-SOX2, OCT4-SOX2 | aggressive cancer phenotype | T47D | [130] | |||
| SP1, ZNF281, ZNF740, MZF1 and RREB1 | aggressive cancer phenotype | T47D | [130] | |||
| ZEB1 | E- and N-cadherins | Induces EMT | HT1080 | rBM matrix | [135] | |
| PROX1 | MMP14 | Inhibits invasion | MDA-MB-231 | 3D fibrin matrix | [127] | |
| JunB | EMT-induction | NMuMG and iLEC cells | 3D co-culture fibrin beads | [140] | ||
| Apoptosis | NF-κB | Induced by tissue polarity and drives resistance to apoptosis | HMT-3522 sourced S-1 and T4-2 cells | rBM matrix | [56] | |
| RUNX2 | Bcl-2 and IL-8 | Disruption of normal acinar morphology, increased cell proliferation | MCF10A and MDA-MB-231 | rBM matrix | [91] | |
| IRF6 | Induced by detachment and downregulation causes anoikis resistance | MCF10A, HMEC | rBM matrix | [153] | ||
| Therapy resistance and drug response | HIF-1α | MDR-1 | Increased resistance to doxorubicin | MCF7 | [173] | |
| NF-κB | β1-integrin | Increased radioresistance | HMT-3522 S1 and T4-2 cells | rBM matrix | [176] | |
| MYC | Drives resistance to everolimus | MCF7, ZR75, CAMA-1 | rBM matrix | [180] | ||
| E2F1, ELK1, GATA, P53 and STAT4 | Increased activity upon lapatinib treatment | MCF10A/ErbBB2 | rBM matrix | [188] | ||
| SREBP2 | HMGCR and HMGCS1 | Sensitizes to fluvastatin-induced apoptosis | MDA-MB-231 and MCF7 | rBM matrix | [187] | |
| Stemness | SOX2 and FOXO1 | Accumulation of FOXO1 and enhanced SOX2 transcription supports BC stemness | Human breast cancer tissues | ULA plates | [193] | |
| NANOG and OCT4 | Increased expression in mammospheres | MCF7 and SKBR3, and MDA-MB-231 | Agar-coated plates | [199] | ||
| SOX2, NANOG, OCT4, and FOXO3 | Increased expression in cell surface-GPNMBhigh cells | BT-474, Hs578T, and MDA-MB-468 | ULA or HEMA-coated plates | [201] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Özkan, H.; Öztürk, D.G.; Korkmaz, G. Transcriptional Factor Repertoire of Breast Cancer in 3D Cell Culture Models. Cancers 2022, 14, 1023. https://doi.org/10.3390/cancers14041023
Özkan H, Öztürk DG, Korkmaz G. Transcriptional Factor Repertoire of Breast Cancer in 3D Cell Culture Models. Cancers. 2022; 14(4):1023. https://doi.org/10.3390/cancers14041023
Chicago/Turabian StyleÖzkan, Hande, Deniz Gülfem Öztürk, and Gozde Korkmaz. 2022. "Transcriptional Factor Repertoire of Breast Cancer in 3D Cell Culture Models" Cancers 14, no. 4: 1023. https://doi.org/10.3390/cancers14041023
APA StyleÖzkan, H., Öztürk, D. G., & Korkmaz, G. (2022). Transcriptional Factor Repertoire of Breast Cancer in 3D Cell Culture Models. Cancers, 14(4), 1023. https://doi.org/10.3390/cancers14041023