
Clinical Applications of Short Non-Coding RNA-Based Therapies in the Era of Precision Medicine

 , ,
, ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
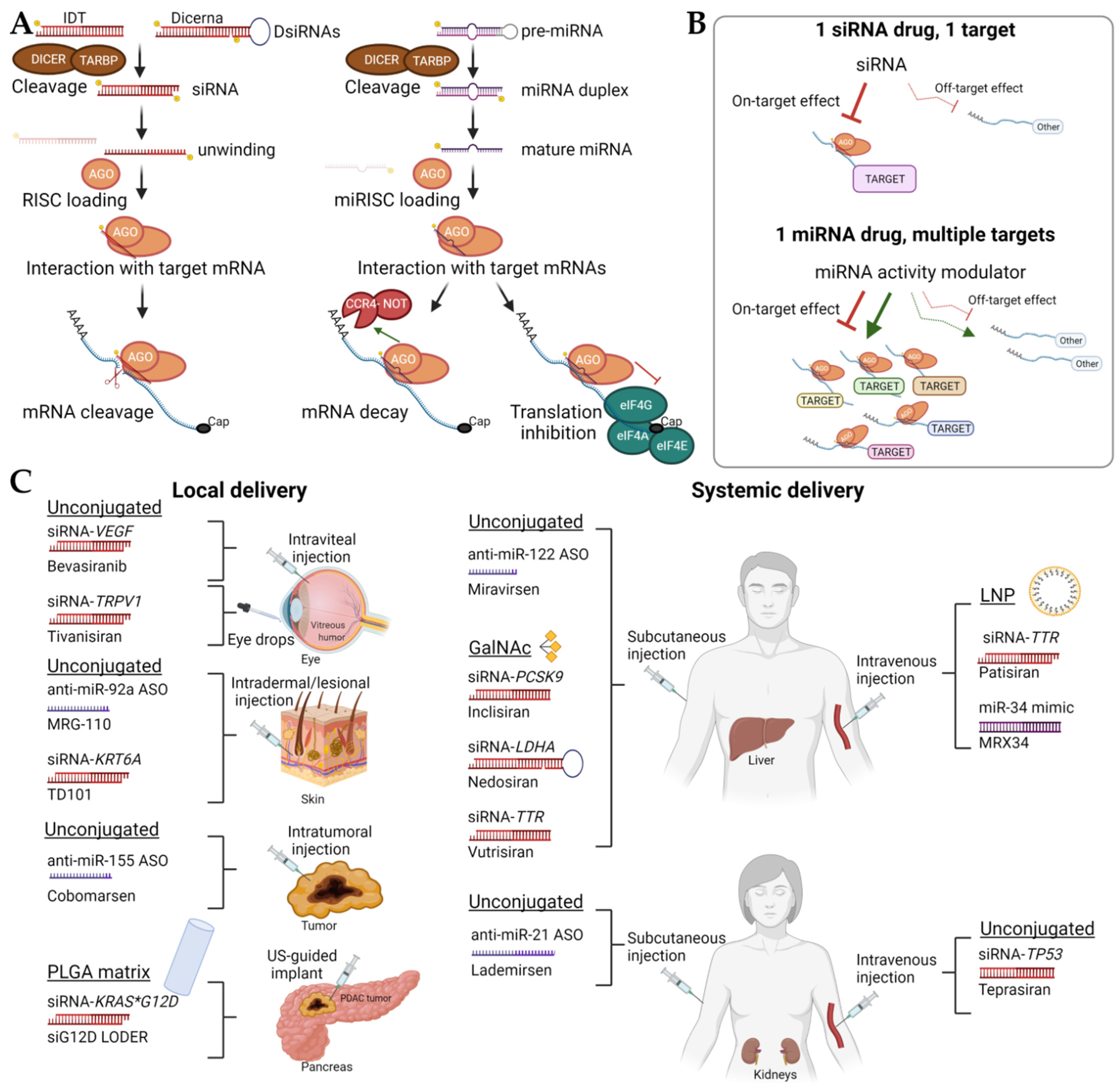
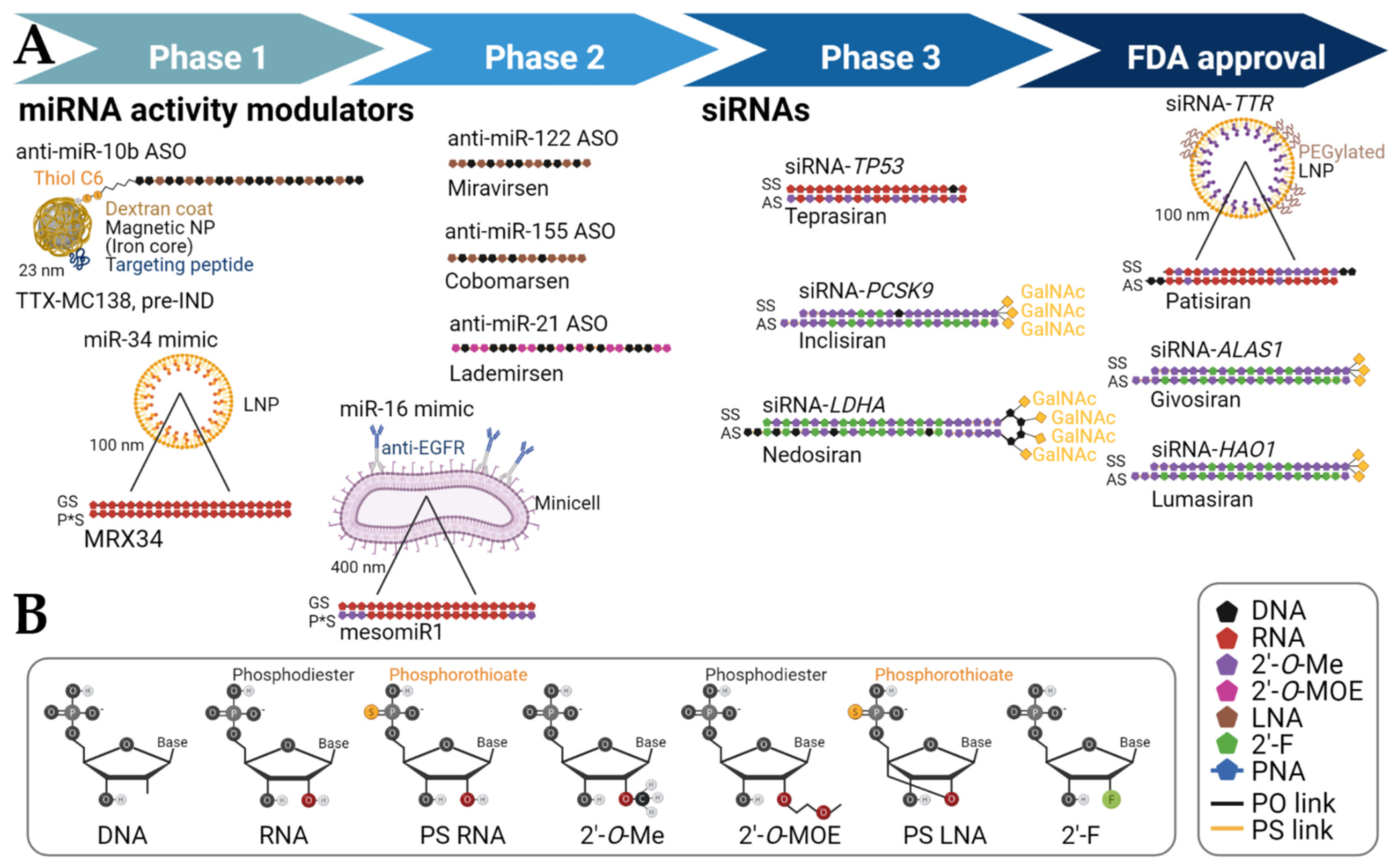
2. Delivery Strategies and Chemical Modifications to Reach the Target Organ or Tissues
3. Clinical Application of Short ncRNAs
3.1. Unconjugated sncRNAs
3.1.1. Diabetic Macular Edema and Age-Related Macular Degeneration
3.1.2. Respiratory Syncytial Infection
3.1.3. Pachyonychia Congenita
3.1.4. Hepatitis C
3.1.5. Acute Kidney Injury
3.1.6. Alport’s Disease
3.1.7. Cardiovascular Disease
3.1.8. Leukemias and Lymphomas
3.2. GalNAc-Conjugated sncRNAs
3.2.1. Porphyria
3.2.2. Alpha-1 Antitrypsin Deficiency
3.2.3. Primary Hyperoxaluria
3.2.4. Hemophilia
3.2.5. Hepatitis B
3.2.6. Cholesterol Metabolism and Atherosclerotic Cardiovascular Disease
3.2.7. Atypical Hemolytic Uremic Syndrome
3.3. Lipid Nanoparticle Therapies
3.3.1. Transthyretin Amyloidosis
3.3.2. Liver Fibrosis
3.3.3. Hepatocarcinoma and Liver Metastases
3.3.4. Pancreatic Cancer
3.4. DOPC Nanoliposomes
Solid Tumors
3.5. Mini-Cells, Larger Encapsulation
Mesothelioma
4. miRNA Therapeutics in Preclinical Investigation
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beg, M.S.; Brenner, A.J.; Sachdev, J.; Borad, M.; Kang, Y.K.; Stoudemire, J.; Smith, S.; Bader, A.G.; Kim, S.; Hong, D.S. Phase I study of MRX34, a liposomal miR-34a mimic, administered twice weekly in patients with advanced solid tumors. Investig. New Drugs 2017, 35, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Forterre, A.; Komuro, H.; Aminova, S.; Harada, M. A Comprehensive Review of Cancer MicroRNA Therapeutic Delivery Strategies. Cancers 2020, 12, 1852. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Zhang, Q.; Chen, W.; Wu, T.; Liu, S.; Li, X.; Luo, B.; Zhang, T.; Yan, G.; Lu, H.; et al. MicroRNA-301a promotes pancreatic cancer invasion and metastasis through the JAK/STAT3 signaling pathway by targeting SOCS5. Carcinogenesis 2020, 41, 502–514. [Google Scholar] [CrossRef]
- Kulkarni, J.A.; Witzigmann, D.; Thomson, S.B.; Chen, S.; Leavitt, B.R.; Cullis, P.R.; van der Meel, R. The current landscape of nucleic acid therapeutics. Nat. Nanotechnol. 2021, 16, 630–643. [Google Scholar] [CrossRef]
- Sempere, L.F.; Azmi, A.S.; Moore, A. microRNA-based diagnostic and therapeutic applications in cancer medicine. Wiley Interdiscip. Rev. RNA 2021, 12, e1662. [Google Scholar] [CrossRef]
- Roberts, T.C.; Langer, R.; Wood, M.J.A. Advances in oligonucleotide drug delivery. Nat. Rev. Drug Discov. 2020, 19, 673–694. [Google Scholar] [CrossRef]
- Hickerson, R.P.; Leachman, S.A.; Pho, L.N.; Gonzalez-Gonzalez, E.; Smith, F.J.; McLean, W.I.; Contag, C.H.; Leake, D.; Milstone, L.M.; Kaspar, R.L. Development of quantitative molecular clinical end points for siRNA clinical trials. J. Investig. Dermatol. 2011, 131, 1029–1036. [Google Scholar] [CrossRef] [Green Version]
- Hanna, J.; Hossain, G.S.; Kocerha, J. The Potential for microRNA Therapeutics and Clinical Research. Front. Genet. 2019, 10, 478. [Google Scholar] [CrossRef] [Green Version]
- Zou, G.M.; Wu, W.; Chen, J.; Rowley, J.D. Duplexes of 21-nucleotide RNAs mediate RNA interference in differentiated mouse ES cells. Biol. Cell 2003, 95, 365–371. [Google Scholar] [CrossRef]
- Meister, G. Argonaute proteins: Functional insights and emerging roles. Nat. Rev. Genet. 2013, 14, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Monsellato, I.; Azienda Ospedaliera, S.S. Antonio e Biagio e Cesare Arrigo di Alessandria. Timisnar—Biomarkers Substudy (Timisnar-mirna). Available online: https://ClinicalTrials.gov/show/NCT03962088 (accessed on 24 February 2022).
- Crooke, S.T.; Witztum, J.L.; Bennett, C.F.; Baker, B.F. RNA-targeted therapeutics. Cell Metab. 2018, 27, 714–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crooke, S.T. Molecular mechanisms of antisense oligonucleotides. Nucleic Acid Ther. 2017, 27, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Quemener, A.M.; Bachelot, L.; Forestier, A.; Donnou-Fournet, E.; Gilot, D.; Galibert, M.D. The powerful world of antisense oligonucleotides: From bench to bedside. Wiley Interdiscip. Rev. RNA 2020, 11, e1594. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [Green Version]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef]
- Garba, A.O.; Mousa, S.A. Bevasiranib for the treatment of wet, age-related macular degeneration. Ophthalmol. Eye Dis. 2010, 2, 75–83. [Google Scholar] [CrossRef]
- Regulus. Regulus Announces Clinical Candidate Nomination for the Treatment of Glioblastoma Multiforme. Available online: http://ir.regulusrx.com/news-releases/news-release-details/regulus-announces-clinical-candidate-nomination-treatment (accessed on 14 March 2022).
- Transcode. Targeting microRNA-10b. Available online: https://www.transcodetherapeutics.com/ttx-mc138.html (accessed on 14 March 2022).
- Sarett, S.M.; Nelson, C.E.; Duvall, C.L. Technologies for controlled, local delivery of siRNA. J. Control. Release 2015, 218, 94–113. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.E.; Zuckerman, J.E.; Choi, C.H.; Seligson, D.; Tolcher, A.; Alabi, C.A.; Yen, Y.; Heidel, J.D.; Ribas, A. Evidence of RNAi in humans from systemically administered siRNA via targeted nanoparticles. Nature 2010, 464, 1067–1070. [Google Scholar] [CrossRef]
- Kavita, U.; Miller, W.; Ji, Q.C.; Pillutla, R.C. A Fit-for-Purpose Method for the Detection of Human Antibodies to Surface-Exposed Components of BMS-986263, a Lipid Nanoparticle-Based Drug Product Containing a siRNA Drug Substance. AAPS J. 2019, 21, 92. [Google Scholar] [CrossRef]
- Hu, B.; Zhong, L.; Weng, Y.; Peng, L.; Huang, Y.; Zhao, Y.; Liang, X.J. Therapeutic siRNA: State of the art. Signal Transduct. Target. Ther. 2020, 5, 101. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, C. PCSK9-gene-silencing, cholesterol-lowering drug impresses. Nat. Biotechnol. 2019, 37, 1385–1387. [Google Scholar] [CrossRef] [PubMed]
- Ray, K.K.; Wright, R.S.; Kallend, D.; Koenig, W.; Leiter, L.A.; Raal, F.J.; Bisch, J.A.; Richardson, T.; Jaros, M.; Wijngaard, P.L.J.; et al. Two Phase 3 Trials of Inclisiran in Patients with Elevated LDL Cholesterol. N. Engl. J. Med. 2020, 382, 1507–1519. [Google Scholar] [CrossRef] [PubMed]
- Lindow, M.; Kauppinen, S. Discovering the first microRNA-targeted drug. J. Cell Biol. 2012, 199, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Debacker, A.J.; Voutila, J.; Catley, M.; Blakey, D.; Habib, N. Delivery of Oligonucleotides to the Liver with GalNAc: From Research to Registered Therapeutic Drug. Mol. Ther. 2020, 28, 1759–1771. [Google Scholar] [CrossRef]
- Machiela, E.; Popkie, A.; Sempere, L. Individual Noncoding RNA Variations: Their Role in Shaping and Maintaining the Epigenetic Landscape. In Personalized Epigenetics; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Sempere, L.F.; Powell, K.; Rana, J.; Brock, A.A.; Schmittgen, T.D. Role of non-coding RNAs in tumor progression and metastasis in pancreatic cancer. Cancer Metastasis Rev. 2021, 40, 761–776. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef]
- Pardini, B.; Calin, G.A. MicroRNAs and Long Non-Coding RNAs and Their Hormone-Like Activities in Cancer. Cancers 2019, 11, 378. [Google Scholar] [CrossRef] [Green Version]
- Anfossi, S.; Babayan, A.; Pantel, K.; Calin, G.A. Clinical utility of circulating non-coding RNAs—An update. Nat. Rev. Clin. Oncol. 2018, 15, 541–563. [Google Scholar] [CrossRef]
- Dolgin, E. The tangled history of mRNA vaccines. Nature 2021, 597, 318–324. [Google Scholar] [CrossRef]
- Kaiser, P.K.; Symons, R.C.; Shah, S.M.; Quinlan, E.J.; Tabandeh, H.; Do, D.V.; Reisen, G.; Lockridge, J.A.; Short, B.; Guerciolini, R.; et al. RNAi-based treatment for neovascular age-related macular degeneration by Sirna-027. Am. J. Ophthalmol. 2010, 150, 33–39.e2. [Google Scholar] [CrossRef]
- DeVincenzo, J.; Lambkin-Williams, R.; Wilkinson, T.; Cehelsky, J.; Nochur, S.; Walsh, E.; Meyers, R.; Gollob, J.; Vaishnaw, A. A randomized, double-blind, placebo-controlled study of an RNAi-based therapy directed against respiratory syncytial virus. Proc. Natl. Acad. Sci. USA 2010, 107, 8800–8805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leachman, S.A.; Hickerson, R.P.; Schwartz, M.E.; Bullough, E.E.; Hutcherson, S.L.; Boucher, K.M.; Hansen, C.D.; Eliason, M.J.; Srivatsa, G.S.; Kornbrust, D.J.; et al. First-in-human mutation-targeted siRNA phase Ib trial of an inherited skin disorder. Mol. Ther. 2010, 18, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, X.; Jin, R.; Chen, L.; Dang, M.; Cao, H.; Dong, Y.; Cai, B.; Bai, G.; Gooding, J.J.; et al. Injectable hydrogel with MSNs/microRNA-21-5p delivery enables both immunomodification and enhanced angiogenesis for myocardial infarction therapy in pigs. Sci. Adv. 2021, 7, eabd6740. [Google Scholar] [CrossRef] [PubMed]
- Hinkel, R.; Ramanujam, D.; Kaczmarek, V.; Howe, A.; Klett, K.; Beck, C.; Dueck, A.; Thum, T.; Laugwitz, K.L.; Maegdefessel, L.; et al. AntimiR-21 Prevents Myocardial Dysfunction in a Pig Model of Ischemia/Reperfusion Injury. J. Am. Coll. Cardiol. 2020, 75, 1788–1800. [Google Scholar] [CrossRef] [PubMed]
- Ambati, J. Age-related macular degeneration and the other double helix the cogan lecture. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2166–2169. [Google Scholar] [CrossRef]
- du Castel, C. Safety and Efficacy Study of Small Interfering RNA Molecule (Cand5) to Treat Diabetic Macular Edema. Available online: https://ClinicalTrials.gov/show/NCT00306904 (accessed on 5 January 2022).
- Gottlieb, J.; Zamora, M.R.; Hodges, T.; Musk, A.W.; Sommerwerk, U.; Dilling, D.; Arcasoy, S.; DeVincenzo, J.; Karsten, V.; Shah, S.; et al. ALN-RSV01 for prevention of bronchiolitis obliterans syndrome after respiratory syncytial virus infection in lung transplant recipients. J. Heart Lung Transplant. Off. Publ. Int. Soc. Heart Transplant. 2016, 35, 213–221. [Google Scholar] [CrossRef] [PubMed]
- DeVincenzo, J.; Cehelsky, J.E.; Alvarez, R.; Elbashir, S.; Harborth, J.; Toudjarska, I.; Nechev, L.; Murugaiah, V.; Van Vliet, A.; Vaishnaw, A.K.; et al. Evaluation of the safety, tolerability and pharmacokinetics of ALN-RSV01, a novel RNAi antiviral therapeutic directed against respiratory syncytial virus (RSV). Antivir. Res. 2008, 77, 225–231. [Google Scholar] [CrossRef]
- Astor, T.L. RNA interference, RSV, and lung transplantation: A promising future for siRNA therapeutics. Am. J. Respir. Crit. Care Med. 2011, 183, 427–428. [Google Scholar] [CrossRef]
- Leachman, S.A. Study of TD101, a Small Interfering RNA (siRNA) Designed for Treatment of Pachyonychia Congenita. Available online: https://ClinicalTrials.gov/show/NCT00716014 (accessed on 6 January 2022).
- van der Ree, M.H.; van der Meer, A.J.; van Nuenen, A.C.; de Bruijne, J.; Ottosen, S.; Janssen, H.L.; Kootstra, N.A.; Reesink, H.W. Miravirsen dosing in chronic hepatitis C patients results in decreased microRNA-122 levels without affecting other microRNAs in plasma. Aliment. Pharmacol. Ther. 2016, 43, 102–113. [Google Scholar] [CrossRef] [Green Version]
- Gebert, L.F.; Rebhan, M.A.; Crivelli, S.E.; Denzler, R.; Stoffel, M.; Hall, J. Miravirsen (SPC3649) can inhibit the biogenesis of miR-122. Nucleic Acids Res. 2014, 42, 609–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abplanalp, W.T.; Fischer, A.; John, D.; Zeiher, A.M.; Gosgnach, W.; Darville, H.; Montgomery, R.; Pestano, L.; Allée, G.; Paty, I.; et al. Efficiency and Target Derepression of Anti-miR-92a: Results of a First in Human Study. Nucleic Acid Ther. 2020, 30, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.P.; Van Pham, L.; Uzcategui, N.; Bukh, J. Functional analysis of microRNA-122 binding sequences of hepatitis C virus and identification of variants with high resistance against a specific antagomir. J. Gen. Virol. 2016, 97, 1381–1394. [Google Scholar] [CrossRef] [PubMed]
- Ottosen, S.; Parsley, T.B.; Yang, L.; Zeh, K.; van Doorn, L.J.; van der Veer, E.; Raney, A.K.; Hodges, M.R.; Patick, A.K. In vitro antiviral activity and preclinical and clinical resistance profile of miravirsen, a novel anti-hepatitis C virus therapeutic targeting the human factor miR-122. Antimicrob. Agents Chemother. 2015, 59, 599–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, S.H.; Wang, B.; Kota, J.; Yu, J.; Costinean, S.; Kutay, H.; Yu, L.; Bai, S.; La Perle, K.; Chivukula, R.R.; et al. Essential metabolic, anti-inflammatory, and anti-tumorigenic functions of miR-122 in liver. J. Clin. Investig. 2012, 122, 2871–2883. [Google Scholar] [CrossRef] [Green Version]
- Wonnacott, A.; Meran, S.; Amphlett, B.; Talabani, B.; Phillips, A. Epidemiology and outcomes in community-acquired versus hospital-acquired AKI. Clin. J. Am. Soc. Nephrol. 2014, 9, 1007–1014. [Google Scholar] [CrossRef] [Green Version]
- James, M.T.; Hemmelgarn, B.R.; Wiebe, N.; Pannu, N.; Manns, B.J.; Klarenbach, S.W.; Tonelli, M.; Alberta Kidney Disease Network. Glomerular filtration rate, proteinuria, and the incidence and consequences of acute kidney injury: A cohort study. Lancet 2010, 376, 2096–2103. [Google Scholar] [CrossRef]
- Thielmann, M.; Corteville, D.; Szabo, G.; Swaminathan, M.; Lamy, A.; Lehner, L.J.; Brown, C.D.; Noiseux, N.; Atta, M.G.; Squiers, E.C.; et al. Teprasiran, a Small Interfering RNA, for the Prevention of Acute Kidney Injury in High-Risk Patients Undergoing Cardiac Surgery: A Randomized Clinical Study. Circulation 2021, 144, 1133–1144. [Google Scholar] [CrossRef]
- Bautista-Sanchez, D.; Arriaga-Canon, C.; Pedroza-Torres, A.; De La Rosa-Velazquez, I.A.; Gonzalez-Barrios, R.; Contreras-Espinosa, L.; Montiel-Manriquez, R.; Castro-Hernandez, C.; Fragoso-Ontiveros, V.; Alvarez-Gomez, R.M.; et al. The Promising Role of miR-21 as a Cancer Biomarker and Its Importance in RNA-Based Therapeutics. Mol. Ther. Nucleic Acids 2020, 20, 409–420. [Google Scholar] [CrossRef]
- Sheedy, F.J. Turning 21: Induction of miR-21 as a Key Switch in the Inflammatory Response. Front. Immunol. 2015, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Gomez, I.G.; MacKenna, D.A.; Johnson, B.G.; Kaimal, V.; Roach, A.M.; Ren, S.; Nakagawa, N.; Xin, C.; Newitt, R.; Pandya, S.; et al. Anti—microRNA-21 oligonucleotides prevent Alport nephropathy progression by stimulating metabolic pathways. J. Clin. Investig. 2015, 125, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Song, W.; Boulanger, J.; Xu, E.Y.; Wang, F.; Zhang, Y.; He, Q.; Wang, S.; Yang, L.; Pryce, C.; et al. Dysregulated Expression of microRNA-21 and Disease-Related Genes in Human Patients and in a Mouse Model of Alport Syndrome. Hum. Gene Ther. 2019, 30, 865–881. [Google Scholar] [CrossRef] [PubMed]
- Gallant-Behm, C.L.; Piper, J.; Dickinson, B.A.; Dalby, C.M.; Pestano, L.A.; Jackson, A.L. A synthetic microRNA-92a inhibitor (MRG-110) accelerates angiogenesis and wound healing in diabetic and nondiabetic wounds. Wound Repair Regen. 2018, 26, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Seto, A.G.; Beatty, X.; Lynch, J.M.; Hermreck, M.; Tetzlaff, M.; Duvic, M.; Jackson, A.L. Cobomarsen, an oligonucleotide inhibitor of miR-155, co-ordinately regulates multiple survival pathways to reduce cellular proliferation and survival in cutaneous T-cell lymphoma. Br. J. Haematol. 2018, 183, 428–444. [Google Scholar] [CrossRef] [Green Version]
- miRagen. Miragen Announces Internal Review of Preliminary Topline Data for the Phase 2 Solar Clinical Trial of Cobomarsen in Patients with Cutaneous T-Cell Lymphoma (CTCL). Available online: http://investors.miragen.com/press-releases/press-release/2020/miRagen-Announces-Internal-Review-of-Preliminary-Topline-Data-for-the-Phase-2-SOLAR-Clinical-Trial-of-Cobomarsen-in-Patients-with-Cutaneous-T-Cell-Lymphoma-CTCL/default.aspx (accessed on 14 March 2022).
- Anastasiadou, E.; Seto, A.; Beatty, X.; Hermreck, M.; Gilles, M.E.; Stroopinsky, D.; Pinter-Brown, L.C.; Pestano, L.; Marchese, C.; Avigan, D.; et al. Cobomarsen, an oligonucleotide inhibitor of miR-155, slows DLBCL tumor cell growth in vitro and in vivo. Clin. Cancer Res. 2020, 27, 1139–1149. [Google Scholar] [CrossRef]
- Sardh, E.; Balwani, M.; Rees, D.C.; Stein, P.; Stölzel, U.; Peiro, P.A.; Bissell, D.M.; Bonkovsky, H.L.; Keel, S.; Parker, C.; et al. S1169 Twelve-Month Interim Analysis of Efficacy and Safety of Givosiran, an Investigational RNAi Therapeutic for Acute Hepatic Porphyria, in the ENVISION Open Label Extension. Off. J. Am. Coll. Gastroenterol. ACG 2020, 115, S584–S585. [Google Scholar] [CrossRef]
- Neeleman, R.A.; Wensink, D.; Wagenmakers, M.; Mijnhout, G.S.; Friesema, E.C.H.; Langendonk, J.G. Diagnostic and therapeutic strategies for porphyrias. Neth. J. Med. 2020, 78, 149–160. [Google Scholar]
- Springer, A.D.; Dowdy, S.F. GalNAc-siRNA Conjugates: Leading the Way for Delivery of RNAi Therapeutics. Nucleic Acid Ther. 2018, 28, 109–118. [Google Scholar] [CrossRef]
- Balwani, M.; Sardh, E.; Ventura, P.; Peiró, P.A.; Rees, D.C.; Stölzel, U.; Bissell, D.M.; Bonkovsky, H.L.; Windyga, J.; Anderson, K.E.; et al. Phase 3 Trial of RNAi Therapeutic Givosiran for Acute Intermittent Porphyria. N. Engl. J. Med. 2020, 382, 2289–2301. [Google Scholar] [CrossRef]
- Turner, A.M.; Stolk, J.; Bals, R.; Lickliter, J.D.; Hamilton, J.; Christianson, D.R.; Given, B.D.; Burdon, J.G.; Loomba, R.; Stoller, J.K.; et al. Hepatic-targeted RNA interference provides robust and persistent knockdown of alpha-1 antitrypsin levels in ZZ patients. J. Hepatol. 2018, 69, 378–384. [Google Scholar] [CrossRef]
- Wood, K.D.; Holmes, R.P.; Erbe, D.; Liebow, A.; Fargue, S.; Knight, J. Reduction in urinary oxalate excretion in mouse models of Primary Hyperoxaluria by RNA interference inhibition of liver lactate dehydrogenase activity. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2019, 1865, 2203–2209. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.J.; Keam, S.J. Lumasiran: First Approval. Drugs 2021, 81, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Frishberg, Y.; Deschenes, G.; Cochat, P.; Magen, D.; Groothoff, J.; Hulton, S.A.; Harambat, J.; vant Hoff, W.; Hoppe, B.; Lieske, J.C.; et al. Mp12-14 Safety and Efficacy Study of Lumasiran, an Investigational RNA Interference (RNAI) Therapeutic, in Adult and Pediatric Patients with Primary Hyperoxaluria Type 1 (PH1). J. Urol. 2019, 201, e174. [Google Scholar] [CrossRef] [Green Version]
- Garrelfs, S.F.; Frishberg, Y.; Hulton, S.A.; Koren, M.J.; O’Riordan, W.D.; Cochat, P.; Deschênes, G.; Shasha-Lavsky, H.; Saland, J.M.; Van’t Hoff, W.G.; et al. Lumasiran, an RNAi Therapeutic for Primary Hyperoxaluria Type 1. N. Engl. J. Med. 2021, 384, 1216–1226. [Google Scholar] [CrossRef] [PubMed]
- Frishberg, Y.; Deschenes, G.; Groothoff, J.W.; Hulton, S.A.; Magen, D.; Harambat, J.; Van’t Hoff, W.G.; Lorch, U.; Milliner, D.S.; Lieske, J.C.; et al. Phase 1/2 Study of Lumasiran for Treatment of Primary Hyperoxaluria Type 1: A Placebo-Controlled Randomized Clinical Trial. Clin. J. Am. Soc. Nephrol. 2021, 16, 1025–1036. [Google Scholar] [CrossRef]
- Machin, N.; Ragni, M.V. An investigational RNAi therapeutic targeting antithrombin for the treatment of hemophilia A and B. J. Blood Med. 2018, 9, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Pasi, K.J.; Georgiev, P.; Mant, T.; Lissitchkov, T.; Creagh, M.D.; Bevan, D.; Austin, S.; Hay, C.R.; Hegemann, I.; Kazmi, R.; et al. Fitusiran, an Investigational RNAi Therapeutic Targeting Antithrombin for the Treatment of Hemophilia: Updated Results from a Phase 1 and Phase 1/2 Extension Study in Patients with Inhibitors. Blood 2016, 128, 1397. [Google Scholar] [CrossRef]
- Schluep, T.; Lickliter, J.; Hamilton, J.; Lewis, D.L.; Lai, C.L.; Lau, J.Y.; Locarnini, S.A.; Gish, R.G.; Given, B.D. Safety, Tolerability, and Pharmacokinetics of ARC-520 Injection, an RNA Interference-Based Therapeutic for the Treatment of Chronic Hepatitis B Virus Infection, in Healthy Volunteers. Clin. Pharmacol. Drug Dev. 2017, 6, 350–362. [Google Scholar] [CrossRef] [Green Version]
- Yuen, M.F.; Schiefke, I.; Yoon, J.H.; Ahn, S.H.; Heo, J.; Kim, J.H.; Lik Yuen Chan, H.; Yoon, K.T.; Klinker, H.; Manns, M.; et al. RNA Interference Therapy with ARC-520 Results in Prolonged Hepatitis B Surface Antigen Response in Patients with Chronic Hepatitis B Infection. Hepatology 2020, 72, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Pirner, M. Study of Safety and Tolerability of DCR HBVS. Available online: https://ClinicalTrials.gov/show/NCT03772249 (accessed on 1 April 2021).
- Wright, R.S.; Collins, M.G.; Stoekenbroek, R.M.; Robson, R.; Wijngaard, P.L.J.; Landmesser, U.; Leiter, L.A.; Kastelein, J.J.P.; Ray, K.K.; Kallend, D. Effects of Renal Impairment on the Pharmacokinetics, Efficacy, and Safety of Inclisiran: An Analysis of the ORION-7 and ORION-1 Studies. Mayo Clin. Proc. 2020, 95, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Raal, F.J.; Kallend, D.; Ray, K.K.; Turner, T.; Koenig, W.; Wright, R.S.; Wijngaard, P.L.J.; Curcio, D.; Jaros, M.J.; Leiter, L.A.; et al. Inclisiran for the Treatment of Heterozygous Familial Hypercholesterolemia. N. Engl. J. Med. 2020, 382, 1520–1530. [Google Scholar] [CrossRef] [PubMed]
- Ray, K.K.; Landmesser, U.; Leiter, L.A.; Kallend, D.; Dufour, R.; Karakas, M.; Hall, T.; Troquay, R.P.; Turner, T.; Visseren, F.L.; et al. Inclisiran in Patients at High Cardiovascular Risk with Elevated LDL Cholesterol. N. Engl. J. Med. 2017, 376, 1430–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, L.; Mafham, M.; Preiss, D.; Landray, M. A Randomized Trial Assessing the Effects of Inclisiran on Clinical Outcomes Among People with Cardiovascular Disease. Available online: https://ClinicalTrials.gov/show/NCT03705234 (accessed on 1 February 2022).
- Wright, S. Trial to Assess the Effect of Long Term Dosing of Inclisiran in Subjects with High CV Risk and Elevated LDL-C. Available online: https://ClinicalTrials.gov/show/NCT03814187 (accessed on 1 February 2022).
- Mario Negri Institute for Pharmacological Research. Eculizumab to Cemdisiran Switch in aHUS. Available online: https://ClinicalTrials.gov/show/NCT03999840 (accessed on 7 March 2022).
- Adams, D.; Gonzalez-Duarte, A.; O’Riordan, W.D.; Yang, C.-C.; Ueda, M.; Kristen, A.V.; Tournev, I.; Schmidt, H.H.; Coelho, T.; Berk, J.L.; et al. Patisiran, an RNAi therapeutic, for hereditary transthyretin amyloidosis. N. Engl. J. Med. 2018, 379, 11–21. [Google Scholar] [CrossRef]
- Emdin, M.; Aimo, A.; Rapezzi, C.; Fontana, M.; Perfetto, F.; Seferović, P.M.; Barison, A.; Castiglione, V.; Vergaro, G.; Giannoni, A.; et al. Treatment of cardiac transthyretin amyloidosis: An update. Eur. Heart J. 2019, 40, 3699–3706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urits, I.; Swanson, D.; Swett, M.C.; Patel, A.; Berardino, K.; Amgalan, A.; Berger, A.A.; Kassem, H.; Kaye, A.; Viswanath, O. A Review of Patisiran (ONPATTRO(R)) for the Treatment of Polyneuropathy in People with Hereditary Transthyretin Amyloidosis. Neurol. Ther. 2020, 9, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Judge, D.P.; Kristen, A.V.; Grogan, M.; Maurer, M.S.; Falk, R.H.; Hanna, M.; Gillmore, J.; Garg, P.; Vaishnaw, A.K.; Harrop, J.; et al. Phase 3 Multicenter Study of Revusiran in Patients with Hereditary Transthyretin-Mediated (hATTR) Amyloidosis with Cardiomyopathy (ENDEAVOUR). Cardiovasc. Drugs Ther. 2020, 34, 357–370. [Google Scholar] [CrossRef] [Green Version]
- Squibb, B.-M. Safety and Effectiveness of BMS-986263 in Adults with Compensated Cirrhosis (Liver Disease) From Nonalcoholic Steatohepatitis (NASH). Available online: https://ClinicalTrials.gov/show/NCT04267393 (accessed on 16 February 2022).
- Squibb, B.-M. A Study of Experimental Medication BMS-986263 in Adults with Advanced Hepatic Fibrosis After Cure of Hepatitis C. Available online: https://ClinicalTrials.gov/show/NCT03420768 (accessed on 16 February 2022).
- Hong, D.S.; Kang, Y.K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H.Y.; Brenner, A.J.; Park, K.; Lee, J.L.; Kim, T.Y.; et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br. J. Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef]
- Synlogic. Pipeline + Programs. Available online: https://www.synlogictx.com/pipeline-programs/ (accessed on 14 March 2022).
- Golan, T.; Khvalevsky, E.Z.; Hubert, A.; Gabai, R.M.; Hen, N.; Segal, A.; Domb, A.; Harari, G.; David, E.B.; Raskin, S.; et al. RNAi therapy targeting KRAS in combination with chemotherapy for locally advanced pancreatic cancer patients. Oncotarget 2015, 6, 24560–24570. [Google Scholar] [CrossRef] [Green Version]
- Mizrahi, J.D.; Surana, R.; Valle, J.W.; Shroff, R.T. Pancreatic cancer. Lancet 2020, 395, 2008–2020. [Google Scholar] [CrossRef]
- Kleeff, J.; Korc, M.; Apte, M.; La Vecchia, C.; Johnson, C.D.; Biankin, A.V.; Neale, R.E.; Tempero, M.; Tuveson, D.A.; Hruban, R.H.; et al. Pancreatic cancer. Nat. Rev. Dis. Primers 2016, 2, 16022. [Google Scholar] [CrossRef]
- Heidel, J.D.; Liu, J.Y.; Yen, Y.; Zhou, B.; Heale, B.S.; Rossi, J.J.; Bartlett, D.W.; Davis, M.E. Potent siRNA inhibitors of ribonucleotide reductase subunit RRM2 reduce cell proliferation in vitro and in vivo. Clin. Cancer Res. 2007, 13, 2207–2215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duxbury, M.S.; Ito, H.; Zinner, M.J.; Ashley, S.W.; Whang, E.E. RNA interference targeting the M2 subunit of ribonucleotide reductase enhances pancreatic adenocarcinoma chemosensitivity to gemcitabine. Oncogene 2004, 23, 1539–1548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartlett, D.W.; Davis, M.E. Impact of tumor-specific targeting and dosing schedule on tumor growth inhibition after intravenous administration of siRNA-containing nanoparticles. Biotechnol. Bioeng. 2008, 99, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Gilles, M.E.; Hao, L.; Brown, K.; Lim, J.; Bhatia, S.N.; Slack, F.J. Tumor penetrating nanomedicine targeting both an oncomiR and an oncogene in pancreatic cancer. Oncotarget 2019, 10, 5349–5358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, L.; Tao, H.; Xu, H.; Li, C.; Qiao, G.; Guo, M.; Cao, S.; Liu, M.; Lin, X. Exosomes-Coated miR-34a Displays Potent Antitumor Activity in Pancreatic Cancer Both in vitro and in vivo. Drug Des. Dev. Ther. 2020, 14, 3495–3507. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R. The biology and function of exosomes in cancer. J. Clin. Investig. 2016, 126, 1208–1215. [Google Scholar] [CrossRef] [PubMed]
- Kamerkar, S.; LeBleu, V.S.; Sugimoto, H.; Yang, S.; Ruivo, C.F.; Melo, S.A.; Lee, J.J.; Kalluri, R. Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer. Nature 2017, 546, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Luan, X.; Sansanaphongpricha, K.; Myers, I.; Chen, H.; Yuan, H.; Sun, D. Engineering exosomes as refined biological nanoplatforms for drug delivery. Acta Pharmacol. Sin. 2017, 38, 754–763. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Cao, F.; Sun, H.; Wang, Y.; Liu, S.; Wu, Y.; Cui, Q.; Mei, W.; Li, F. Exosomes derived from human umbilical cord mesenchymal stromal cells deliver exogenous miR-145-5p to inhibit pancreatic ductal adenocarcinoma progression. Cancer Lett. 2019, 442, 351–361. [Google Scholar] [CrossRef]
- Xie, Y.; Hang, Y.; Wang, Y.; Sleightholm, R.; Prajapati, D.R.; Bader, J.; Yu, A.; Tang, W.; Jaramillo, L.; Li, J.; et al. Stromal Modulation and Treatment of Metastatic Pancreatic Cancer with Local Intraperitoneal Triple miRNA/siRNA Nanotherapy. ACS Nano 2020, 14, 255–271. [Google Scholar] [CrossRef]
- Shang, S.; Wang, J.; Chen, S.; Tian, R.; Zeng, H.; Wang, L.; Xia, M.; Zhu, H.; Zuo, C. Exosomal miRNA-1231 derived from bone marrow mesenchymal stem cells inhibits the activity of pancreatic cancer. Cancer Med. 2019, 8, 7728–7740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.M.; Wen, X.; Han, X.R.; Wang, S.; Wang, Y.J.; Shen, M.; Fan, S.H.; Zhang, Z.F.; Shan, Q.; Li, M.Q.; et al. Bone Marrow Mesenchymal Stem Cell-Derived Exosomal MicroRNA-126-3p Inhibits Pancreatic Cancer Development by Targeting ADAM9. Mol. Ther. Nucleic Acids 2019, 16, 229–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendt, M.; Kamerkar, S.; Sugimoto, H.; McAndrews, K.M.; Wu, C.C.; Gagea, M.; Yang, S.; Blanko, E.V.R.; Peng, Q.; Ma, X.; et al. Generation and testing of clinical-grade exosomes for pancreatic cancer. JCI Insight 2018, 3, e99263. [Google Scholar] [CrossRef] [PubMed]
- Ramot, Y.; Rotkopf, S.; Gabai, R.M.; Zorde Khvalevsky, E.; Muravnik, S.; Marzoli, G.A.; Domb, A.J.; Shemi, A.; Nyska, A. Preclinical Safety Evaluation in Rats of a Polymeric Matrix Containing an siRNA Drug Used as a Local and Prolonged Delivery System for Pancreatic Cancer Therapy. Toxicol. Pathol. 2016, 44, 856–865. [Google Scholar] [CrossRef] [Green Version]
- Wagner, M.J.; Mitra, R.; McArthur, M.J.; Baze, W.; Barnhart, K.; Wu, S.Y.; Rodriguez-Aguayo, C.; Zhang, X.; Coleman, R.L.; Lopez-Berestein, G.; et al. Preclinical Mammalian Safety Studies of EPHARNA (DOPC Nanoliposomal EphA2-Targeted siRNA). Mol. Cancer Ther. 2017, 16, 1114–1123. [Google Scholar] [CrossRef] [Green Version]
- Coleman, R.L. EphA2 siRNA in Treating Patients with Advanced or Recurrent Solid Tumors. Available online: https://ClinicalTrials.gov/show/NCT01591356 (accessed on 21 January 2022).
- Reid, G.; Kao, S.C.; Pavlakis, N.; Brahmbhatt, H.; MacDiarmid, J.; Clarke, S.; Boyer, M.; van Zandwijk, N. Clinical development of TargomiRs, a miRNA mimic-based treatment for patients with recurrent thoracic cancer. Epigenomics 2016, 8, 1079–1085. [Google Scholar] [CrossRef] [Green Version]
- Kao, S.C.; Fulham, M.; Wong, K.; Cooper, W.; Brahmbhatt, H.; MacDiarmid, J.; Pattison, S.; Sagong, J.O.; Huynh, Y.; Leslie, F.; et al. A Significant Metabolic and Radiological Response after a Novel Targeted MicroRNA-based Treatment Approach in Malignant Pleural Mesothelioma. Am. J. Respir. Crit. Care Med. 2015, 191, 1467–1469. [Google Scholar] [CrossRef]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Yoo, B.; Meka, N.; Sheedy, P.; Billig, A.M.; Pantazopoulos, P.; Medarova, Z. MicroRNA-710 regulates multiple pathways of carcinogenesis in murine metastatic breast cancer. PLoS ONE 2019, 14, e0226356. [Google Scholar] [CrossRef] [Green Version]
- Yoo, B.; Kavishwar, A.; Wang, P.; Ross, A.; Pantazopoulos, P.; Dudley, M.; Moore, A.; Medarova, Z. Therapy targeted to the metastatic niche is effective in a model of stage IV breast cancer. Sci. Rep. 2017, 7, 45060. [Google Scholar] [CrossRef] [Green Version]
- Yoo, B.; Kavishwar, A.; Ross, A.; Wang, P.; Tabassum, D.P.; Polyak, K.; Barteneva, N.; Petkova, V.; Pantazopoulos, P.; Tena, A.; et al. Combining miR-10b—Targeted Nanotherapy with Low-Dose Doxorubicin Elicits Durable Regressions of Metastatic Breast Cancer. Cancer Res. 2015, 75, 4407–4415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.Y.; Lee, J.H.; Ha, M.; Nam, J.W.; Kim, V.N. miR-29 miRNAs activate p53 by targeting p85 alpha and CDC42. Nat. Struct. Mol. Biol. 2009, 16, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Schipper, J.; Westerhuis, J.J.; Beddows, I.; Madaj, Z.; Monsma, D.; Hostetter, G.; Kiupel, M.; Conejo-Garcia, J.R.; Sempere, L.F. Loss of microRNA-21 leads to profound stromal remodeling and short survival in K-Ras-driven mouse models of pancreatic cancer. Int. J. Cancer 2020, 147, 2265–2278. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Xu, F.M.; Zhang, G.Q.; Li, S.B.; Wen, Y.Y.; Zeng, F. Down-regulation of microRNA-21 inhibits cell proliferation and invasion of high-invasion liver cancer stem cells. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7832–7840. [Google Scholar] [CrossRef]
- Zhou, R.; Wang, C.; Wen, C.; Wang, D. miR-21 promotes collagen production in keloid via Smad7. Burns 2017, 43, 555–561. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Target Gene | Drug Name | Chemistry | Platform | Delivery | Treatment (Organ Site) | Sponsor | References |
|---|---|---|---|---|---|---|---|
| ALAS1 | ALN-AS1 (Givosiran) * | siRNA (2′-O-Me, 2′F, partial PS backbone) | GalNAc conjugation, 2.5 mg/kg | Subcutaneous | Acute Hepatic Porphyrias (Liver) | Alnylam Pharmaceuticals (Cambridge, MA, USA) | NCT03338816, Completed |
| AT | Fitusiran ALN-AT3SC (Fitusiran) | siRNA (2′-O-Me, 2′-F, partial PS backbone) | GalNAc conjugation | Subcutaneous | Hemophilia A or B (Liver) | Genzyme, a Sanofi Company (Cambridge, MA, USA) | NCT03417102/03417245, Completed; NCT03754790/NCT03549871, Active |
| CASP2 | QPI-1007 | siRNA (2′-O-Me) | Up to 3 mg | Intraviteal | Acute Nonarteritic Anterior Ischemic Optic Neuropathy (Eye) | Quark Pharmaceuticals (Newark, CA, USA) | NCT02341560, Terminated |
| HAO1 | ALN-GO1 (Lumasiran) * | siRNA (2′-O-Me, 2′F, partial PS backbone) | GalNAc conjugation, up to 3 mg/kg | Subcutaneous | Primary Hyperoxaluria Type 1 (Liver) | Alnylam Pharmaceuticals (Cambridge, MA, USA) | NCT03681184, Active; NCT03905694, Active; NCT04152200, Active |
| LDHA | DCR-PHXC (Nedosiran) | DsiRNA pseudo-hairpin (2′-O-Me, 2′F, DNA, partial PS backbone) | GalXC | Subcutaneous | Hyperoxaluria (Liver) | Dicerna Pharmaceuticals (Lexington, MA, USA) | NCT04042402, Enrolling by invitation |
| PCSK9 | Inclisiran | siRNA (2′-O-Me, 2′F, internal DNA, partial PS backbone) | GalNAc conjugation, 300 mg | Subcutaneous | Homozygous Familial Hypercholesterolemia (Liver) | Novartis Pharmaceuticals (Basel, Switzerland) | NCT03851705, Active; NCT04659863, Recruiting |
| PCSK9 | Inclisiran | siRNA (2′-O-Me, 2′F, internal DNA, partial PS) | GalNAc conjugation, 300 mg | Subcutaneous | Atherosclerotic Cardiovascular Disease (ASCVD) or ASCVD High Risk and Elevated LDL-C (Liver) | Novartis Pharmaceuticals (Basel, Switzerland) | NCT04765657, Recruiting |
| PCSK9 | Inclisiran | siRNA (2′-O-Me, 2′F, internal DNA, partial PS backbone) | GalNAc conjugation, 300 mg | Subcutaneous | Prevent Cardiovascular events in Participants with Established Cardiovascular Disease (Liver) | Novartis Pharmaceuticals (Basel, Switzerland) | NCT05030428, Recruiting |
| TP53 | QPI-1002 (Teprasiran) | siRNA (2′-O-Me) | - | Intravenous | Improved Graft Function after Donor Kidney Transplant (Kidney) | Quark Pharmaceuticals (Newark, CA, USA) | NCT02610296, Completed |
| TP53 | QPI-1002 (Teprasiran) | siRNA (2′-O-Me) | - | Intravenous | Prevention of acute kidney injury after cardiac surgery (Kidney) | Quark Pharmaceuticals (Newark, CA, USA) | NCT03510897, Terminated |
| TRPV1 | SYL1001 (Tivanisiran) | siRNA | Ophthalmic solution | Periocular | Sjögren′s Syndrome, Dry eye (Eye) | Sylentis, S.A. (Madrid, Spain) | NCT04819269, Recruiting |
| TRPV1 | SYL1001 (Tivanisiran) | siRNA | Ophthalmic solution, 11.25 mg/mL | Periocular | Moderate to Severe Dry Eye Disease (Eye) | Sylentis, S.A. (Madrid, Spain) | NCT03108664, Completed |
| TTR | ALN-TTR02 (patisiran)* | siRNA (2′-O-Me, DNA overhangs) | Lipid nanoparticle | Intravenous | Transthyretin-Mediated Polyneuropathy (Liver) | Alnylam Pharmaceuticals (Cambridge, MA, USA) | NCT01960348, Completed |
| TTR | ALN-TTR02 (patisiran) | siRNA (2′-O-Me, DNA overhangs) | Lipid nanoparticle, 0.3 mg/kg | Intravenous | hATTR amyloidosis with disease progression after liver transplant (Liver) | Alnylam Pharmaceuticals (Cambridge, MA, USA) | NCT03862807, Completed |
| TTR | ALN-TTR02 (patisiran) | siRNA (2′-O-Me, DNA overhangs) | Lipid nanoparticle | Intravenous | ATTR Amyloidosis with Cardiomyopathy (Liver) | Alnylam Pharmaceuticals (Cambridge, MA, USA) | NCT03997383, Active |
| TTR | ALN-TTRSC (Revusiran) | siRNA (2′-O-Me, 2′-F) | GalNAc conjugation | Subcutaneous | Transthyretin-Mediated Familial Amyloidotic Cardiomyopathy (Liver) | Alnylam Pharmaceuticals (Cambridge, MA, USA) | NCT02319005, Completed |
| TTR | ALN-TTRSC02 (Vutrisiran) | siRNA (2′-O-Me, 2′-F, partial PS backbone) | GalNAc conjugation, 25 mg | Subcutaneous | Transthyretin Amyloidosis with Cardiomyopathy (Liver) | Alnylam Pharmaceuticals (Cambridge, MA, USA) | NCT04153149, Active |
| TTR | ALN-TTRSC02 (Vutrisiran) | siRNA (2′-O-Me, 2′-F, partial PS backbone) | GalNAc conjugation | Subcutaneous | hATTR Amyloidosis (Liver) | Alnylam Pharmaceuticals (Cambridge, MA, USA) | NCT03759379, Active |
| VEGF | Bevasiranib | siRNA | Up to 2.5 mg | Intraviteal | Age-Related Macular Degeneration following initiation of anti-VEGF Lucentis® antibody therapy (Eye) | OPKO Health, Inc. (Miami, FL, USA) | NCT00557791, Withdrawn |
| miRNA Modulation | Drug Name | Chemistry | Platform | Delivery | Disease (Organ Site) | Sponsor | Clinical Status | References |
|---|---|---|---|---|---|---|---|---|
| miR-10b inhibition | RGLS5579 | ASO (2′-O-MOE, partial PS backbone) | - | Intravenous or intracranial | Glioblastoma (Brain) | Regulus Therapeutics (San Diego, CA, USA) | Pre-IND filing | [19] |
| miR-10b inhibition | TTX-MC138 | ASO (partial LNA, partial PS backbone) | Dextran-coated iron oxide magnetic nanoparticle | Intravenous | Metastatic breast cancer (Lung, other organs) | Transcode Therapeutics (Boston, MA, USA) | Pre-IND filing, scheduled 2022 | [20] |
| miR-16 restoration | mesomiR1 (TargomiR) | dsRNA mimic (2′-O-Me on passenger strand only) | Bacterial minicells with anti-EGFR bispecific antibody | Intravenous | Recurrent malignant pleural mesothelioma and non-small cell lung cancer (Lung) | Asbestos Diseases Research Foundation (New South Wales, Australia), EnGeneIC Limited (Lane Cave West, Australia) | Phase 1 | NCT02369198, Competed |
| miR-21 inhibition | Lademirsen (SAR339375; previously known as RG-012 [Regulus]) | ASO (sugar 2′ position modifications, PS backbone) | Unconjugated | Subcutaneous, 1.5 mg/kg | Alport syndrome (Kidney) | Genzyme, a Sanofi Company (Cambridge, MA, USA) | Phase 1 | NCT02855268, Completed |
| miR-21 inhibition | Lademirsen (SAR339375; previously known as RG-012 [Regulus]) | ASO (sugar 2′ position modifications, PS backbone) | Unconjugated | Subcutaneous | Alport syndrome (Kidney) | Genzyme, a Sanofi Company (Cambridge, MA, USA) | Phase 2 | NCT02855268, Recruiting |
| miR-34a restoration | MRX34 | dsRNA mimic | Liposome | Intravenous | Primary liver cancer or other selected solid tumors or hematologic malignancies (Liver, other organs) | Mirna Therapeutics (Austin, TX, USA) | Phase 1 | NCT01829971, Terminated; NCT02862145, Withdrawn |
| miR-92a inhibition | MRG-110 | ASO (LNA-modified) | - | Intradermal | Wound healing | miRagen Therapeutics, Inc. (Boulder, CO, USA) | Phase 1 | NCT03603431, Completed |
| miR-122 inhibition | Miravirsen (SPC3649) | ASO (partial LNA, PS backbone) | Unconjugated | Subcutaneous | HCV chronic infection (Liver) | Copenhagen, Denmark | Phase 2 | NCT01200420, Completed |
| miR-155 inhibition | MRG-106 (Cobomarsen) | ASO (partial LNA) | Unconjugated | Intratumoral and/or intravenous or subcutaneous | Certain lymphomas and leukemias, including CTCL [mycosis fungoides subtype], CLL, DLBCL [activated B-cell (ABC) subtype], and ATLL | miRagen Therapeutics, Inc. (Boulder, CO, USA) | Phase 1 | NCT02580552, Completed |
| miR-155 inhibition | MRG-106 (Cobomarsen) | ASO (partial LNA) | Unconjugated | Intravenous | CTCL [mycosis fungoides subtype] | miRagen Therapeutics, Inc. (Boulder, CO, USA) | Phase 2 | NCT03713320 and NCT03837457, Terminated |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, E.S.; Whitty, E.; Yoo, B.; Moore, A.; Sempere, L.F.; Medarova, Z. Clinical Applications of Short Non-Coding RNA-Based Therapies in the Era of Precision Medicine. Cancers 2022, 14, 1588. https://doi.org/10.3390/cancers14061588
Smith ES, Whitty E, Yoo B, Moore A, Sempere LF, Medarova Z. Clinical Applications of Short Non-Coding RNA-Based Therapies in the Era of Precision Medicine. Cancers. 2022; 14(6):1588. https://doi.org/10.3390/cancers14061588
Chicago/Turabian StyleSmith, Ellen S., Eric Whitty, Byunghee Yoo, Anna Moore, Lorenzo F. Sempere, and Zdravka Medarova. 2022. "Clinical Applications of Short Non-Coding RNA-Based Therapies in the Era of Precision Medicine" Cancers 14, no. 6: 1588. https://doi.org/10.3390/cancers14061588
APA StyleSmith, E. S., Whitty, E., Yoo, B., Moore, A., Sempere, L. F., & Medarova, Z. (2022). Clinical Applications of Short Non-Coding RNA-Based Therapies in the Era of Precision Medicine. Cancers, 14(6), 1588. https://doi.org/10.3390/cancers14061588