
Elevated MACC1 Expression in Colorectal Cancer Is Driven by Chromosomal Instability and Is Associated with Molecular Subtype and Worse Patient Survival

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Oncotrack Cohort of CRC Patients
2.2. SCNA and Gene Expression Analysis of the Oncotrack Cohort
2.3. Charité Metastasis Cohort of CRC Patients
2.4. DNA Isolation and ddPCR
2.5. TCGA COAD-READ Cohorts of CRC Patients
2.6. Statistical Analysis
2.7. Data Availability
3. Results
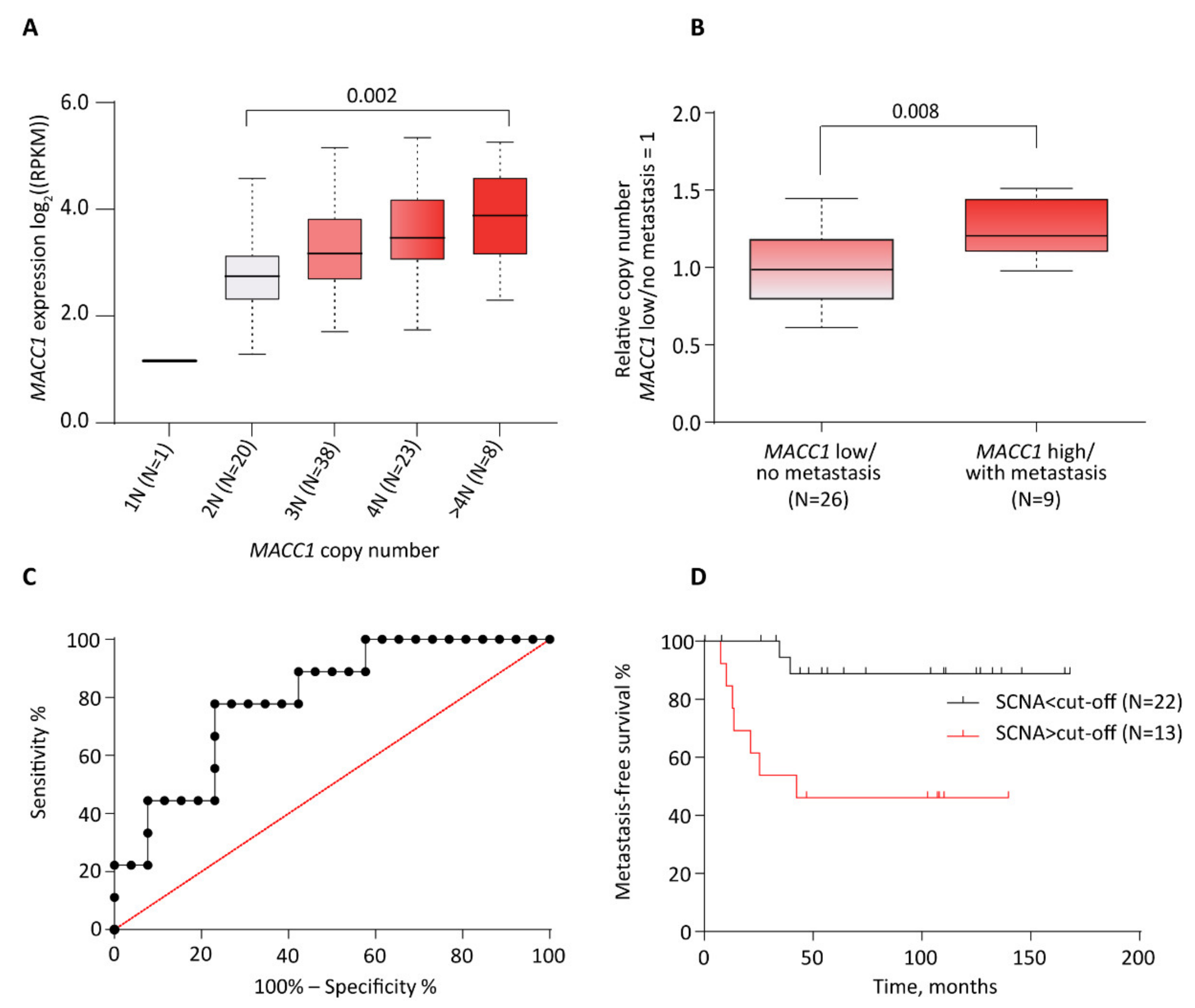
3.1. Correlation between Increased MACC1 SCNAs and Elevated Expression Levels in CRC Patient Samples (Oncotrack Cohort)
3.2. Correlation between Increased MACC1 SCNAs, Elevated Expression Levels and Metastasis Formation in CRC Patients (Charité Cohort)
3.3. Correlation between Increased MACC1 SCNA, Elevated Expression Levels and Clinical, Molecular Subtypes, and Patient Survival Variables (TCGA COAD-READ Cohort)
3.3.1. Association with Anatomopathological Parameters
3.3.2. Association with Tumor Subtypes and Molecular Characteristics
3.3.3. Association with CRC Patient DFS and OS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: Globocan Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Pineros, M.; Znaor, A.; Bray, F. Estimating the Global Cancer Incidence and Mortality in 2018: Globocan Sources and Methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, U.; Schlag, P.M. Clinical, Biological, and Molecular Aspects of Metastasis in Colorectal Cancer. Recent Results Cancer Res. 2007, 176, 61–80. [Google Scholar] [PubMed]
- Stein, U.; Walther, W.; Arlt, F.; Schwabe, H.; Smith, J.; Fichtner, I.; Birchmeier, W.; Schlag, P.M. Macc1, a Newly Identified Key Regulator of Hgf-Met Signaling, Predicts Colon Cancer Metastasis. Nat. Med. 2009, 15, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Lemos, C.; Hardt, M.S.; Juneja, M.; Voss, C.; Förster, S.; Jerchow, B.; Haider, W.; Bläker, H.; Stein, U. Macc1 Induces Tumor Progression in Transgenic Mice and Colorectal Cancer Patients Via Increased Pluripotency Markers Nanog and Oct4. Clin. Cancer Res. 2016, 22, 2812–2824. [Google Scholar] [CrossRef] [Green Version]
- Radhakrishnan, H.; Walther, W.; Zincke, F.; Kobelt, D.; Imbastari, F.; Erdem, M.; Kortüm, B.; Dahlmann, M.; Stein, U. Macc1-the First Decade of a Key Metastasis Molecule from Gene Discovery to Clinical Translation. Cancer Metastasis Rev. 2018, 37, 805–820. [Google Scholar] [CrossRef]
- Jin, Y.; Zhou, K.; Zhao, W.; Han, R.; Huo, X.; Yang, F.; Chen, J. Clinicopathological and Prognostic Significance of Metastasis-Associated in Colon Cancer-1 in Gastric Cancer: A Meta-Analysis. Int. J. Biol. Markers 2019, 34, 27–32. [Google Scholar] [CrossRef]
- Sun, D.-W.; Zhang, Y.-Y.; Qi, Y.; Liu, G.-Q.; Chen, Y.-G.; Ma, J.; Lv, G.-Y. Prognostic and Clinicopathological Significance of Macc1 Expression in Hepatocellular Carcinoma Patients: A Meta-Analysis. Int. J. Clin. Exp. Med. 2015, 8, 4769–4777. [Google Scholar]
- Wang, G.; Fu, Z.; Li, D. Macc1 Overexpression and Survival in Solid Tumors: A Meta-Analysis. Tumour Biol. 2015, 36, 1055–1065. [Google Scholar] [CrossRef]
- Wu, Z.; Zhou, R.; Su, Y.; Sun, L.; Liao, Y.; Liao, W. Prognostic Value of Macc1 in Digestive System Neoplasms: A Systematic Review and Meta-Analysis. Biomed Res. Int. 2015, 2015, 252043. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Dai, C.; Wang, M.; Kang, H.; Lin, S.; Yang, P.; Liu, X.; Liu, K.; Xu, P.; Zheng, Y.; et al. Clinicopathological and Prognostic Significance of Metastasis-Associated in Colon Cancer-1 (Macc1) Overexpression in Colorectal Cancer: A Meta-Analysis. Oncotarget 2016, 7, 62966–62975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Fan, L.; Xu, H.; Jiang, H. Prognostic Significance of the Expression of Metastasis-Associated in Colon Cancer-1 in Gynecologic Cancers and Breast Cancer: A Protocol for Systematic Review and Meta-Analysis. Medicine 2021, 100, e24255. [Google Scholar] [CrossRef] [PubMed]
- Stein, U. Macc1—A Novel Target for Solid Cancers. Expert Opin. Ther. Targets 2013, 17, 1039–1052. [Google Scholar] [CrossRef] [PubMed]
- Ilm, K.; Fuchs, S.; Mudduluru, G.; Stein, U. Macc1 Is Post-Transcriptionally Regulated by Mir-218 in Colorectal Cancer. Oncotarget 2016, 7, 53443–53458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juneja, M.; Ilm, K.; Schlag, P.M.; Stein, U. Promoter Identification and Transcriptional Regulation of the Metastasis Gene Macc1 in Colorectal Cancer. Mol. Oncol. 2013, 7, 929–943. [Google Scholar] [CrossRef]
- Budczies, J.; Kluck, K.; Walther, W.; Stein, U. Decoding and Targeting the Molecular Basis of Macc1-Driven Metastatic Spread: Lessons from Big Data Mining and Clinical-Experimental Approaches. Semin. Cancer Biol. 2020, 60, 365–379. [Google Scholar] [CrossRef]
- Horvat, M.; Potocnik, U.; Repnik, K.; Kavalar, R.; Zadnik, V.; Potrc, S.; Stabuc, B. Single Nucleotide Polymorphisms in Genes Macc1, Rad18, Mmp7 and Sdf-1a as Prognostic Factors in Resectable Colorectal Cancer. Radiol. Oncol. 2017, 51, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Lang, A.H.; Geller-Rhomberg, S.; Winder, T.; Stark, N.; Gasser, K.; Hartmann, B.; Kohler, B.; Grizelj, I.; Drexel, H.; Muendlein, A. A Common Variant of the Macc1 Gene Is Significantly Associated with Overall Survival in Colorectal Cancer Patients. BMC Cancer 2012, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Schmid, F.; Burock, S.; Klockmeier, K.; Schlag, P.M.; Stein, U. Snps in the Coding Region of the Metastasis-Inducing Gene Macc1 and Clinical Outcome in Colorectal Cancer. Mol. Cancer 2012, 11, 49. [Google Scholar] [CrossRef] [Green Version]
- Dai, Z.J.; Liu, X.H.; Kang, H.F.; Wang, X.J.; Jin, T.B.; Zhang, S.Q.; Feng, T.; Ma, X.B.; Wang, M.; Feng, Y.J.; et al. Genetic Variation in Metastasis-Associated in Colon Cancer-1 and the Risk of Breast Cancer among the Chinese Han Population: A Strobe-Compliant Observational Study. Medicine 2016, 95, e2801. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, H.; Cai, J.; Fang, L.; Wu, J.; Ye, C.; Zhu, X.; Li, M. Overexpression of Macc1 and Its Significance in Human Breast Cancer Progression. Cell Biosci. 2013, 3, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muendlein, A.; Hubalek, M.; Geller-Rhomberg, S.; Gasser, K.; Winder, T.; Drexel, H.; Decker, T.; Mueller-Holzner, E.; Chamson, M.; Marth, C.; et al. Significant Survival Impact of Macc1 Polymorphisms in Her2 Positive Breast Cancer Patients. Eur. J. Cancer 2014, 50, 2134–2141. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Gao, S.; Yang, Z.; Xie, H.; Zhang, C.; Lin, B.; Wu, L.; Zheng, S.; Zhou, L. Single Nucleotide Polymorphisms in the Metastasis-Associated in Colon Cancer-1 Gene Predict the Recurrence of Hepatocellular Carcinoma after Transplantation. Int. J. Med. Sci. 2014, 11, 142–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schütte, M.; Risch, T.; Abdavi-Azar, N.; Boehnke, K.; Schumacher, D.; Keil, M.; Yildiriman, R.; Jandrasits, C.; Borodina, T.; Amstislavskiy, V.; et al. Molecular Dissection of Colorectal Cancer in Pre-Clinical Models Identifies Biomarkers Predicting Sensitivity to Egfr Inhibitors. Nat. Commun. 2017, 8, 14262. [Google Scholar] [CrossRef] [Green Version]
- Fritah, S.; Niclou, S.P.; Azuaje, F. Databases for Lncrnas: A Comparative Evaluation of Emerging Tools. RNA 2014, 20, 1655–1665. [Google Scholar] [CrossRef] [Green Version]
- Xi, R.; Hadjipanayis, A.G.; Luquette, L.J.; Kim, T.M.; Lee, E.; Zhang, J.; Johnson, M.D.; Muzny, D.M.; Wheeler, D.A.; Gibbs, R.A.; et al. Copy Number Variation Detection in Whole-Genome Sequencing Data Using the Bayesian Information Criterion. Proc. Natl. Acad. Sci. USA 2011, 108, E1128–E1136. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Armenia, J.; Zhang, C.; Penson, A.V.; Reznik, E.; Zhang, L.; Minet, T.; Ochoa, A.; Gross, B.E.; Iacobuzio-Donahue, C.A.; et al. Unifying Cancer and Normal Rna Sequencing Data from Different Sources. Sci. Data 2018, 5, 180061. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Sethi, N.S.; Hinoue, T.; Schneider, B.G.; Cherniack, A.D.; Sanchez-Vega, F.; Seoane, J.A.; Farshidfar, F.; Bowlby, R.; Islam, M.; et al. Comparative Molecular Analysis of Gastrointestinal Adenocarcinomas. Cancer Cell 2018, 33, 721–735.e8. [Google Scholar] [CrossRef] [Green Version]
- A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. R Core Team. 2020, Volume 2018. Available online: https://Www.R-Project.Org/ (accessed on 3 January 2021).
- Vasaikar, S.; Huang, C.; Wang, X.; Petyuk, V.A.; Savage, S.R.; Wen, B.; Dou, Y.; Zhang, Y.; Shi, Z.; Arshad, O.A.; et al. Proteogenomic Analysis of Human Colon Cancer Reveals New Therapeutic Opportunities. Cell 2019, 177, 1035–1049.e19. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. Rsem: Accurate Transcript Quantification from Rna-Seq Data with or without a Reference Genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Toyota, M.; Ahuja, N.; Ohe-Toyota, M.; Herman, J.G.; Baylin, S.B.; Issa, J.-P. Cpg Island Methylator Phenotype in Colorectal Cancer. Proc. Natl. Acad. Sci. USA 1999, 96, 8681–8686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guinney, J.; Dienstmann, R.; Wang, X.; De Reyniès, A.; Schlicker, A.; Soneson, C.; Marisa, L.; Roepman, P.; Nyamundanda, G.; Angelino, P.; et al. The Consensus Molecular Subtypes of Colorectal Cancer. Nat. Med. 2015, 21, 1350–1356. [Google Scholar] [CrossRef] [PubMed]
- Finotello, F.; Mayer, C.; Plattner, C.; Laschober, G.; Rieder, D.; Hackl, H.; Krogsdam, A.; Loncova, Z.; Posch, W.; Wilflingseder, D.; et al. Molecular and Pharmacological Modulators of the Tumor Immune Contexture Revealed by Deconvolution of Rna-Seq Data. Genome Med. 2019, 11, 34. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Lichtenberg, T.M.; Hoadley, K.A.; Poisson, L.M.; Lazar, A.J.; Cherniack, A.D.; Kovatich, A.J.; Benz, C.C.; Levine, D.A.; Lee, A.V.; et al. An Integrated Tcga Pan-Cancer Clinical Data Resource to Drive High-Quality Survival Outcome Analytics. Cell 2018, 173, 400–416.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichorner, A.; Sack, U.; Kobelt, D.; Kelch, I.; Arlt, F.; Smith, J.; Walther, W.; Schlag, P.M.; Stein, U. In Vivo Imaging of Colorectal Cancer Growth and Metastasis by Targeting Macc1 with Shrna in Xenografted Mice. Clin. Exp. Metastasis 2012, 29, 573–583. [Google Scholar] [CrossRef]
- Pino, M.S.; Chung, D.C. The Chromosomal Instability Pathway in Colon Cancer. Gastroenterology 2010, 138, 2059–2072. [Google Scholar] [CrossRef] [Green Version]
- Galimi, F.; Torti, D.; Sassi, F.; Isella, C.; Corà, D.; Gastaldi, S.; Ribero, D.; Muratore, A.; Massucco, P.; Siatis, D.; et al. Genetic and Expression Analysis of Met, Macc1, and Hgf in Metastatic Colorectal Cancer: Response to Met Inhibition in Patient Xenografts and Pathologic Correlations. Clin. Cancer Res. 2011, 17, 3146–3156. [Google Scholar] [CrossRef] [Green Version]
- Shimokawa, H.; Uramoto, H.; Onitsuka, T.; Chundong, G.; Hanagiri, T.; Oyama, T.; Yasumoto, K. Overexpression of Macc1 Mrna in Lung Adenocarcinoma Is Associated with Postoperative Recurrence. J. Thorac. Cardiovasc. Surg. 2011, 141, 895–898. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.; Zhang, X.; Zhang, R.L.; He, X.-F.; Zhang, J.-G.; Yan, W.-H. Prognostic and Risk Stratification Value of Lesion Macc1 Expression in Colorectal Cancer Patients. Front. Oncol. 2019, 9, 28. [Google Scholar] [CrossRef]
- Koelzer, V.H.; Herrmann, P.; Zlobec, I.; Karamitopoulou, E.; Lugli, A.; Stein, U. Heterogeneity Analysis of Metastasis Associated in Colon Cancer 1 (Macc1) for Survival Prognosis of Colorectal Cancer Patients: A Retrospective Cohort Study. BMC Cancer 2015, 15, 160. [Google Scholar] [CrossRef] [Green Version]
- Greenman, C.D.; Bignell, G.; Butler, A.; Edkins, S.; Hinton, J.; Beare, D.; Swamy, S.; Santarius, T.; Chen, L.; Widaa, S.; et al. Picnic: An Algorithm to Predict Absolute Allelic Copy Number Variation with Microarray Cancer Data. Biostatistics 2010, 11, 164–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoorn, S.T.; de Back, T.R.; Sommeijer, D.W.; Vermeulen, L. Clinical Value of Consensus Molecular Subtypes in Colorectal Cancer: A Systematic Review and Meta-Analysis. J. Natl. Cancer Inst. 2021, djab106. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Li, H.; Shi, H.; Yang, Q.; Zhang, F.; Yang, Y.; Kang, L.; Zhen, T.; Dai, S.; Dong, Y.; et al. Macc1 Down-Regulation Inhibits Proliferation and Tumourigenicity of Nasopharyngeal Carcinoma Cells through Akt/Beta-Catenin Signaling Pathway. PLoS ONE 2013, 8, e60821. [Google Scholar]
- Zhen, T.; Dai, S.; Li, H.; Yang, Y.; Kang, L.; Shi, H.; Zhang, F.; Yang, D.; Cai, S.; He, Y.; et al. Macc1 Promotes Carcinogenesis of Colorectal Cancer Via Beta-Catenin Signaling Pathway. Oncotarget 2014, 5, 3756–3769. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Liu, W.; Zeng, X.-C.; Jiang, N.; Fu, B.-S.; Guo, Y.; Yi, H.-M.; Li, H.; Zhang, Q.; Chen, W.-J.; et al. Down-Regulation of Microrna-338-3p Promoted Angiogenesis in Hepatocellular Carcinoma. Biomed. Pharmacother. 2016, 84, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Ou, S.; Ma, X.; Zhang, S.; Lai, Y. Macc1 Silencing Inhibits Cell Proliferation and Induces Cell Apoptosis of Lung Adenocarcinoma Cells through the Beta-Catenin Pathway. Neoplasma 2018, 65, 552–560. [Google Scholar] [CrossRef]
- Kim, H.J.; Moon, S.J.; Kim, S.H.; Heo, K.; Kim, J.H. Dbc1 Regulates Wnt/Beta-Catenin-Mediated Expression of Macc1, a Key Regulator of Cancer Progression, in Colon Cancer. Cell Death Dis. 2018, 9, 831. [Google Scholar] [CrossRef]
- Zhang, Q.; Xu, P.; Lu, Y.; Dou, H. Correlation of Macc1/C-Myc Expression in Endometrial Carcinoma with Clinical/Pathological Features or Prognosis. Med. Sci. Monit. 2018, 24, 4738–4744. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, Y.; Chen, S.; Li, W.; Chen, W.; Gu, W. Lncrna Macc1-As1 Sponges Multiple Mirnas and Rna-Binding Protein Ptbp1. Oncogenesis 2019, 8, 73. [Google Scholar] [CrossRef]
- He, W.; Liang, B.; Wang, C.; Li, S.; Zhao, Y.; Huang, Q.; Liu, Z.; Yao, Z.; Wu, Q.; Liao, W.; et al. Msc-Regulated Lncrna Macc1-As1 Promotes Stemness and Chemoresistance through Fatty Acid Oxidation in Gastric Cancer. Oncogene 2019, 38, 4637–4654. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, Y.; Zhang, T. Long Non-Coding Rna Macc1-As1 Is Involved in Distant Recurrence of Hepatocellular Carcinoma after Surgical Resection. Med. Sci. Monit. 2020, 26, e921175. [Google Scholar] [CrossRef]
- Peng, T.; Li, Z.; Li, D.; Wang, S. Macc1 Promotes Angiogenesis in Cholangiocarcinoma by Upregulating Vegfa. Onco Targets Ther. 2019, 12, 1893–1903. [Google Scholar] [CrossRef] [PubMed]
- Harpaz, N.; Taboada, S.; Ko, H.M.; Yu, J.; Yang, Q.; Xu, H.; Cao, W. Expression of Macc1 and Met in Inflammatory Bowel Disease-Associated Colonic Neoplasia. Inflamm. Bowel Dis. 2014, 20, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Kobelt, D.; Zhang, C.; Clayton-Lucey, I.A.; Glauben, R.; Voss, C.; Siegmund, B.; Stein, U. Pro-Inflammatory Tnf-Alpha and Ifn-Gamma Promote Tumor Growth and Metastasis Via Induction of Macc1. Front. Immunol. 2020, 11, 980. [Google Scholar] [CrossRef] [PubMed]
- Tijhuis, A.E.; Johnson, S.C.; McClelland, S.E. The Emerging Links between Chromosomal Instability (Cin), Metastasis, Inflammation and Tumour Immunity. Mol. Cytogenet. 2019, 12, 17. [Google Scholar] [CrossRef] [Green Version]
- Raskov, H.; Orhan, A.; Christensen, J.P.; Gogenur, I. Cytotoxic Cd8(+) T Cells in Cancer and Cancer Immunotherapy. Br. J. Cancer 2021, 124, 359–367. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vuaroqueaux, V.; Musch, A.; Kobelt, D.; Risch, T.; Herrmann, P.; Burock, S.; Peille, A.-L.; Yaspo, M.-L.; Fiebig, H.-H.; Stein, U. Elevated MACC1 Expression in Colorectal Cancer Is Driven by Chromosomal Instability and Is Associated with Molecular Subtype and Worse Patient Survival. Cancers 2022, 14, 1749. https://doi.org/10.3390/cancers14071749
Vuaroqueaux V, Musch A, Kobelt D, Risch T, Herrmann P, Burock S, Peille A-L, Yaspo M-L, Fiebig H-H, Stein U. Elevated MACC1 Expression in Colorectal Cancer Is Driven by Chromosomal Instability and Is Associated with Molecular Subtype and Worse Patient Survival. Cancers. 2022; 14(7):1749. https://doi.org/10.3390/cancers14071749
Chicago/Turabian StyleVuaroqueaux, Vincent, Alexandra Musch, Dennis Kobelt, Thomas Risch, Pia Herrmann, Susen Burock, Anne-Lise Peille, Marie-Laure Yaspo, Heinz-Herbert Fiebig, and Ulrike Stein. 2022. "Elevated MACC1 Expression in Colorectal Cancer Is Driven by Chromosomal Instability and Is Associated with Molecular Subtype and Worse Patient Survival" Cancers 14, no. 7: 1749. https://doi.org/10.3390/cancers14071749
APA StyleVuaroqueaux, V., Musch, A., Kobelt, D., Risch, T., Herrmann, P., Burock, S., Peille, A. -L., Yaspo, M. -L., Fiebig, H. -H., & Stein, U. (2022). Elevated MACC1 Expression in Colorectal Cancer Is Driven by Chromosomal Instability and Is Associated with Molecular Subtype and Worse Patient Survival. Cancers, 14(7), 1749. https://doi.org/10.3390/cancers14071749