

Long Non-Coding RNAs Associated with Mitogen-Activated Protein Kinase in Human Pancreatic Cancer
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
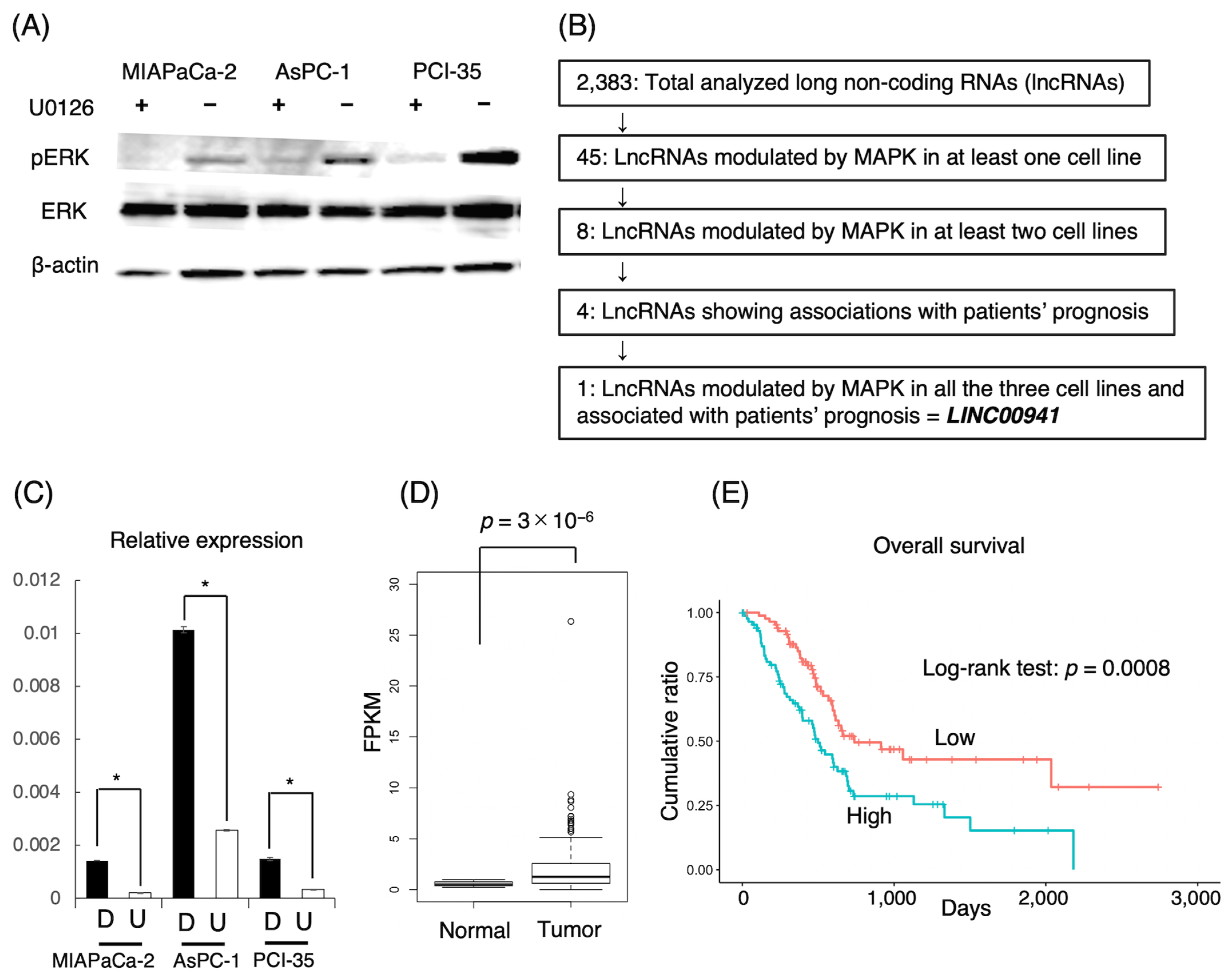
2.1. Methodological Flowchart
2.2. Cell Lines
2.3. Inhibition of MAPK
2.4. Immunoblotting
2.5. Whole Transcriptome Sequencing
2.6. Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR)
2.7. TCGA Open-Access Database Analysis
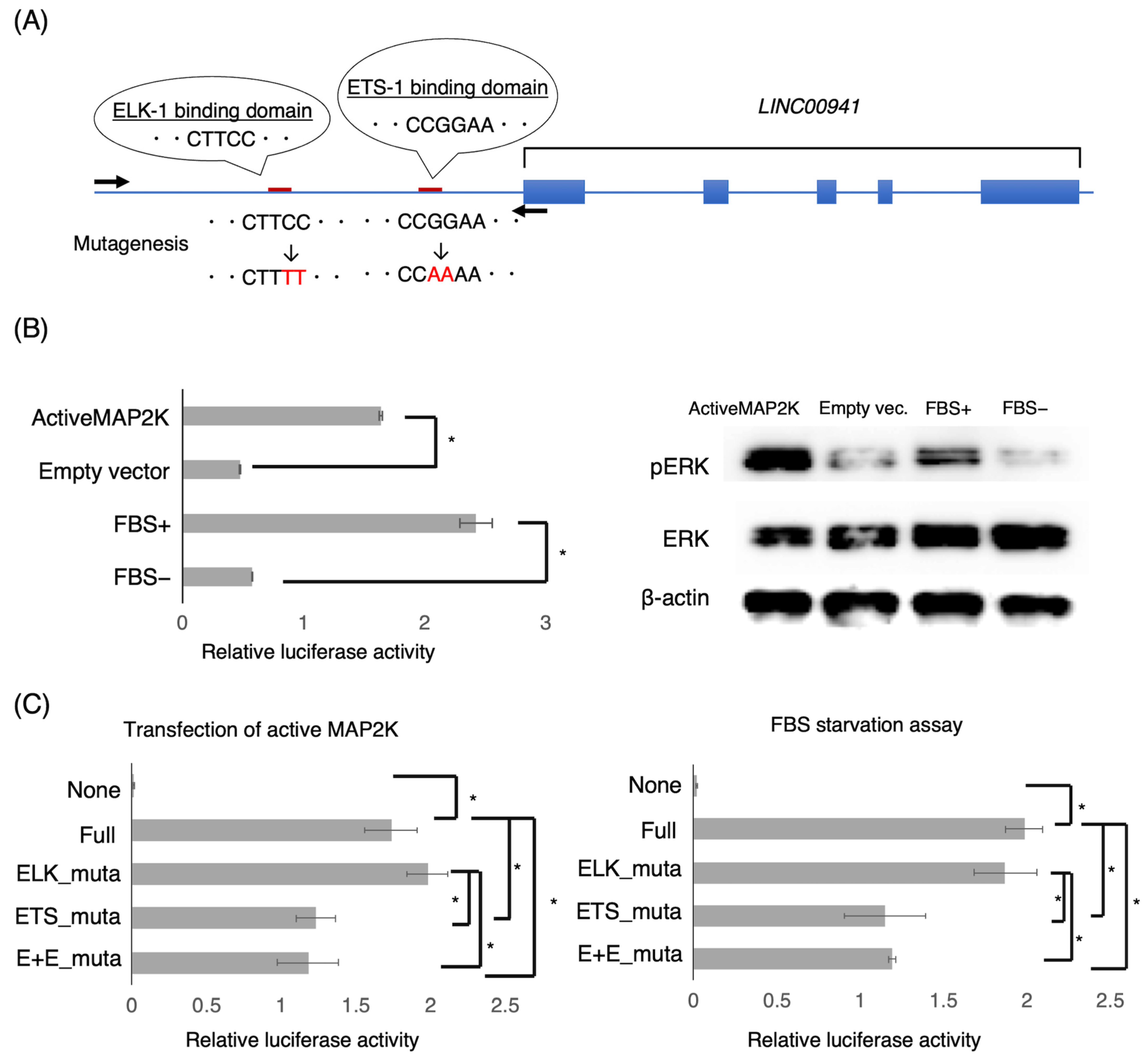
2.8. Promoter Assay
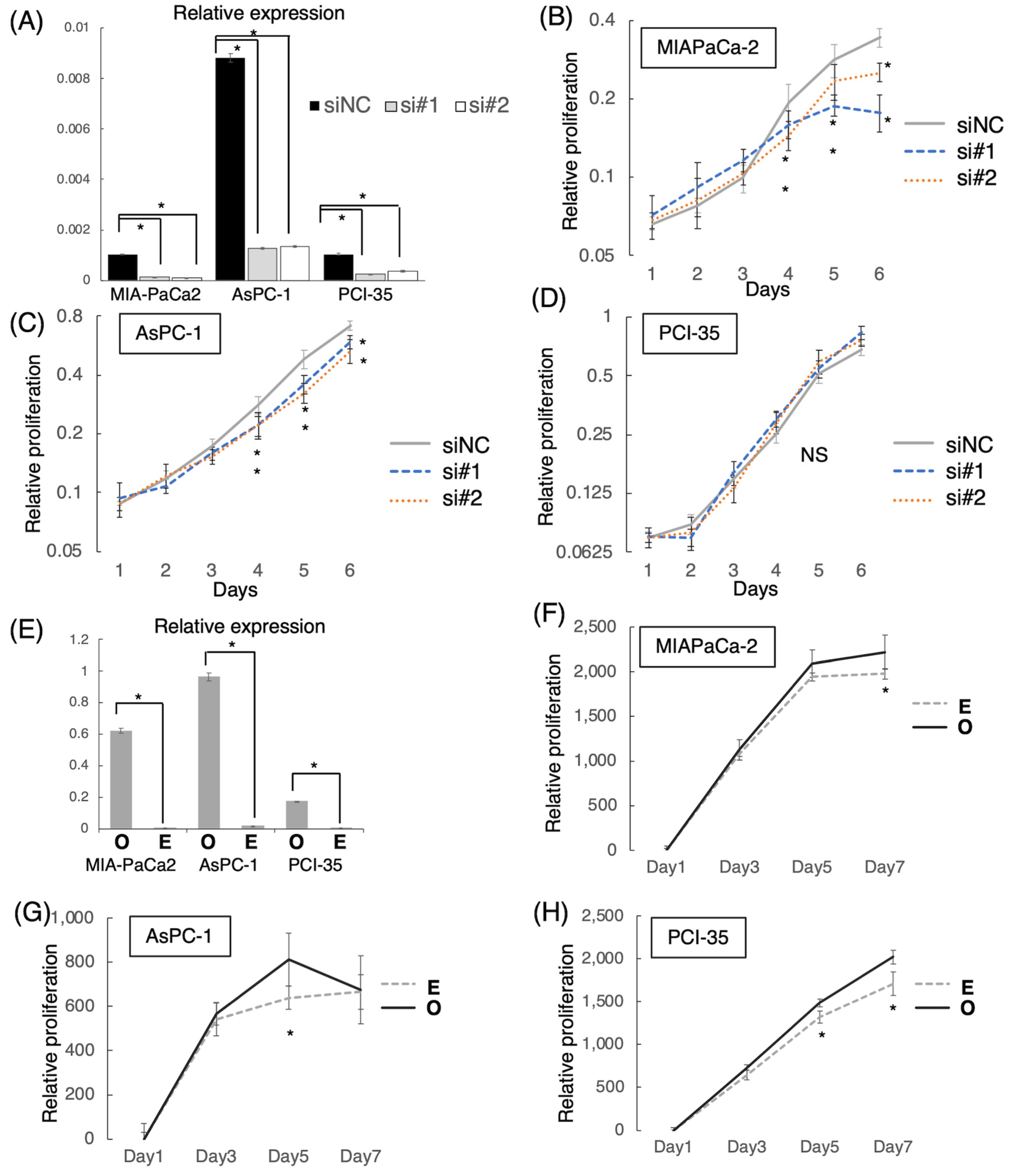
2.9. Cell Proliferation Assay with Altering LINC00941 Expression
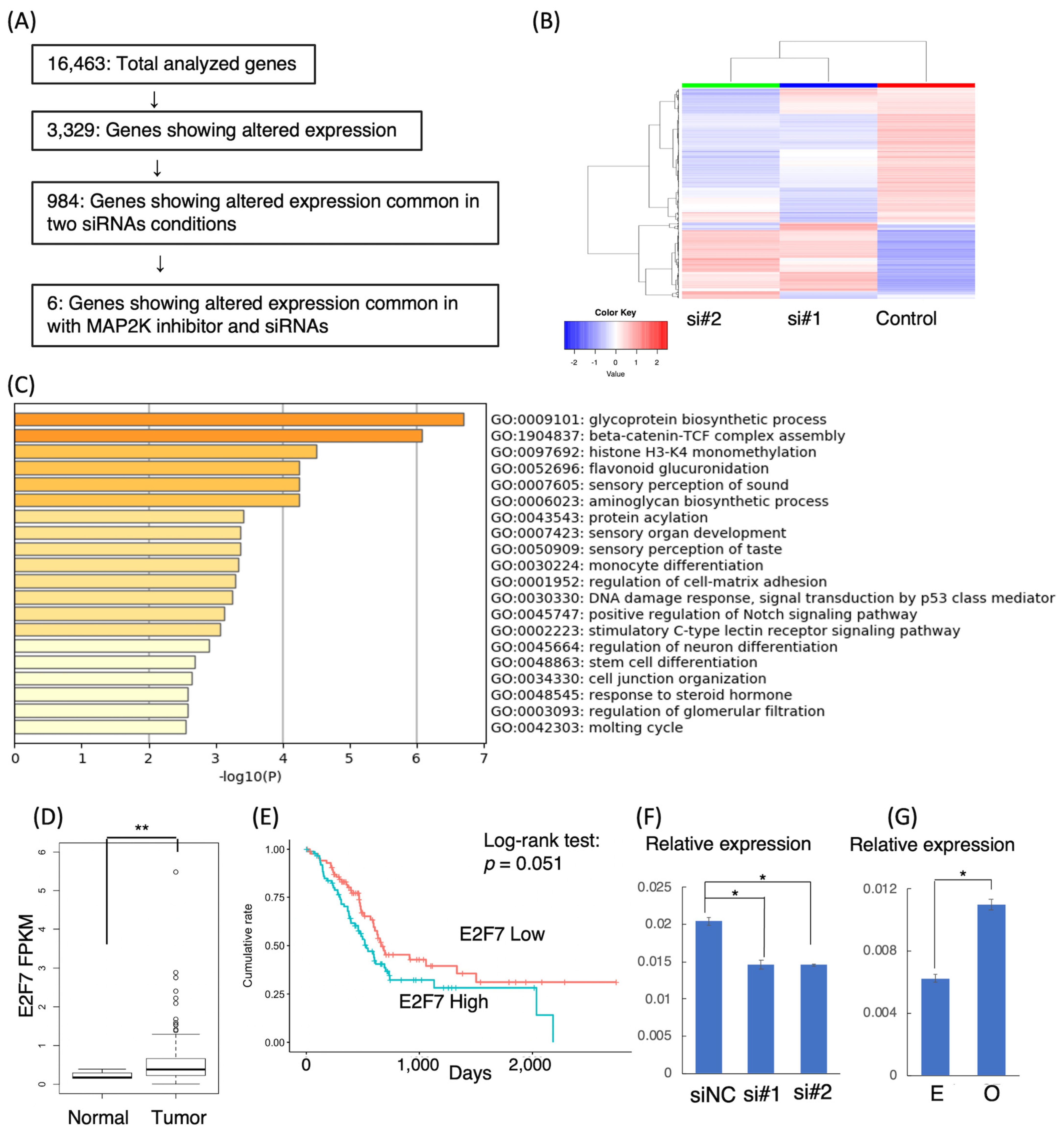
2.10. RNA-Seq Data Analysis
2.11. Statistical Analysis
2.12. Data Availability
3. Results
3.1. LncRNAs Associated with MAPK
3.2. Prognostic Impacts of lncRNAs Associated with MAPK
3.3. LINC00941 Promoter Activity Associated with MAPK
3.4. Effect of LINC00941 on Cell Proliferation
3.5. Target Genes of LINC00941
4. Discussions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matsuda, T.; Ajiki, W.; Marugame, T.; Ioka, A.; Tsukuma, H.; Sobue, T. Population-based survival of cancer patients diagnosed between 1993 and 1999 in Japan: A chronological and international comparative study. Jpn. J. Clin. Oncol. 2011, 41, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, T. Impacts of activation of the mitogen-activated protein kinase pathway in pancreatic cancer. Front. Oncol. 2015, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Cox, A.D.; Fesik, S.W.; Kimmelman, A.C.; Luo, J.; Der, C.J. Drugging the undruggable RAS: Mission possible? Nat. Rev. Drug Discov. 2014, 13, 828–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jänne, P.A.; van den Heuvel, M.M.; Barlesi, F.; Cobo, M.; Mazieres, J.; Crinò, L.; Orlov, S.; Blackhall, F.; Wolf, J.; Garrido, P.; et al. Selumetinib Plus Docetaxel Compared With Docetaxel Alone and Progression-Free Survival in Patients With KRAS-Mutant Advanced Non-Small Cell Lung Cancer: The SELECT-1 Randomized Clinical Trial. JAMA 2017, 317, 1844–1853. [Google Scholar] [CrossRef] [Green Version]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef]
- Sharma, G.G.; Okada, Y.; Von Hoff, D.; Goel, A. Non-coding RNA biomarkers in pancreatic ductal adenocarcinoma. Semin. Cancer Biol. 2021, 75, 153–168. [Google Scholar] [CrossRef]
- Ikeda, Y.; Tanji, E.; Makino, N.; Kawata, S.; Furukawa, T. MicroRNAs associated with mitogen-activated protein kinase in human pancreatic cancer. Mol. Cancer Res. 2012, 10, 259–269. [Google Scholar] [CrossRef] [Green Version]
- Akamatsu, M.; Makino, N.; Ikeda, Y.; Matsuda, A.; Ito, M.; Kakizaki, Y.; Saito, Y.; Ishizawa, T.; Kobayashi, T.; Furukawa, T.; et al. Specific MAPK-Associated MicroRNAs in Serum Differentiate Pancreatic Cancer from Autoimmune Pancreatitis. PLoS ONE 2016, 11, e0158669. [Google Scholar] [CrossRef] [Green Version]
- Iwai, K.; Ishikura, H.; Kaji, M.; Sugiura, H.; Ishizu, A.; Takahashi, C.; Kato, H.; Tanabe, T.; Yoshiki, T. Importance of E-selectin (ELAM-1) and sialyl Lewis(a) in the adhesion of pancreatic carcinoma cells to activated endothelium. Int. J. Cancer 1993, 54, 972–977. [Google Scholar] [CrossRef]
- Mansour, S.J.; Candia, J.M.; Matsuura, J.E.; Manning, M.C.; Ahn, N.G. Interdependent domains controlling the enzymatic activity of mitogen-activated protein kinase kinase 1. Biochemistry 1996, 35, 15529–15536. [Google Scholar] [CrossRef] [PubMed]
- Hata, T.; Furukawa, T.; Sunamura, M.; Egawa, S.; Motoi, F.; Ohmura, N.; Marumoto, T.; Saya, H.; Horii, A. RNA interference targeting aurora kinase a suppresses tumor growth and enhances the taxane chemosensitivity in human pancreatic cancer cells. Cancer Res. 2005, 65, 2899–2905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, S.X.; Son, E.W.; Yao, R. iDEP: An integrated web application for differential expression and pathway analysis of RNA-Seq data. BMC Bioinform. 2018, 19, 534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Matsui, M.; Corey, D.R. Non-coding RNAs as drug targets. Nat. Rev. Drug Discov. 2017, 16, 167–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Cui, R.; Ye, L.; Wang, Y.; Wang, X.; Zhang, Q.; Wang, K.; Dong, C.; Le, W.; Chen, B. LINC00941 promotes glycolysis in pancreatic cancer by modulating the Hippo pathway. Mol. Ther. Nucleic Acids. 2021, 26, 280–294. [Google Scholar] [CrossRef]
- Wang, J.; He, Z.; Xu, J.; Chen, P.; Jiang, J. Long noncoding RNA LINC00941 promotes pancreatic cancer progression by competitively binding miR-335-5p to regulate ROCK1-mediated LIMK1/Cofilin-1 signaling. Cell Death Dis. 2021, 12, 36. [Google Scholar] [CrossRef]
- Yan, X.; Zhang, D.; Wu, W.; Wu, S.; Qian, J.; Hao, Y.; Yan, F.; Zhu, P.; Wu, J.; Huang, G.; et al. Mesenchymal Stem Cells Promote Hepatocarcinogenesis via lncRNA-MUF Interaction with ANXA2 and miR-34a. Cancer Res. 2017, 77, 6704–6716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhao, H.; Xu, Y.; Li, J.; Deng, C.; Deng, Y.; Bai, J.; Li, X.; Xiao, Y.; Zhang, Y. Systematic identification of lincRNA-based prognostic biomarkers by integrating lincRNA expression and copy number variation in lung adenocarcinoma. Int. J. Cancer 2019, 144, 1723–1734. [Google Scholar] [CrossRef]
- Gugnoni, M.; Manicardi, V.; Torricelli, F.; Sauta, E.; Bellazzi, R.; Manzotti, G.; Vitale, E.; de Biase, D.; Piana, S.; Ciarrocchi, A. Linc00941 Is a Novel Transforming Growth Factor β Target That Primes Papillary Thyroid Cancer Metastatic Behavior by Regulating the Expression of Cadherin 6. Thyroid 2021, 31, 247–263. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Tao, Y.; Zhang, Y.; Zhu, Y.; Minyao, D.N.; Haleem, M.; Dong, C.; Zhang, L.; Zhang, X.; Zhao, J.; et al. Regulatory network analysis of high expressed long non-coding RNA LINC00941 in gastric cancer. Gene 2018, 662, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Jiang, M.; Liu, H.; Chu, Y.; Wang, D.; Cao, J.; Wang, Z.; Xie, X.; Han, Y.; Xu, B. LINC00941 promotes CRC metastasis through preventing SMAD4 protein degradation and activating the TGF-beta/SMAD2/3 signaling pathway. Cell Death Differ. 2021, 28, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Ai, Y.; Wu, S.; Zou, C.; Wei, H. LINC00941 promotes oral squamous cell carcinoma progression via activating CAPRIN2 and canonical WNT/beta-catenin signaling pathway. J. Cell Mol. Med. 2020, 24, 10512–10524. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Guo, G.; Li, J.; Chen, J.; Tan, P. Screening key lncRNAs with diagnostic and prognostic value for head and neck squamous cell carcinoma based on machine learning and mRNA-lncRNA co-expression network analysis. Cancer Biomark 2020, 27, 195–206. [Google Scholar] [CrossRef]
- Furukawa, T.; Kanai, N.; Shiwaku, H.O.; Soga, N.; Uehara, A.; Horii, A. AURKA is one of the downstream targets of MAPK1/ERK2 in pancreatic cancer. Oncogene 2006, 25, 4831–4839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foulds, C.E.; Nelson, M.L.; Blaszczak, A.G.; Graves, B.J. Ras/mitogen-activated protein kinase signaling activates Ets-1 and Ets-2 by CBP/p300 recruitment. Mol. Cell Biol. 2004, 24, 10954–10964. [Google Scholar] [CrossRef] [Green Version]
- Kobberup, S.; Nyeng, P.; Juhl, K.; Hutton, J.; Jensen, J. ETS-family genes in pancreatic development. Dev. Dyn. 2007, 236, 3100–3110. [Google Scholar] [CrossRef]
- Furukawa, T.; Tanji, E.; Xu, S.; Horii, A. Feedback regulation of DUSP6 transcription responding to MAPK1 via ETS2 in human cells. Biochem. Biophys. Res. Commun. 2008, 377, 317–320. [Google Scholar] [CrossRef]
- Huang, B.; Chang, C.; Wang, B.L.; Li, H. ELK1-induced upregulation of lncRNA TRPM2-AS promotes tumor progression in gastric cancer by regulating miR-195/ HMGA1 axis. J. Cell Biochem. 2019, 120, 16921–16933. [Google Scholar] [CrossRef]
- Su, X.; Teng, J.; Jin, G.; Li, J.; Zhao, Z.; Cao, X.; Guo, Y.; Guo, M.; Li, X.; Wu, J.; et al. ELK1-induced upregulation of long non-coding RNA MIR100HG predicts poor prognosis and promotes the progression of osteosarcoma by epigenetically silencing LATS1 and LATS2. Biomed. Pharmacother. 2019, 109, 788–797. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Gao, L.; Cui, G.; Cao, Y. LncRNA NEAT1 facilitates pancreatic cancer growth and metastasis through stabilizing ELF3 mRNA. Am. J. Cancer Res. 2020, 10, 237–248. [Google Scholar] [PubMed]
- Luo, Y.; Liu, F.; Yan, C.; Qu, W.; Zhu, L.; Guo, Z.; Zhou, F.; Zhang, W. Long Non-Coding RNA CASC19 Sponges microRNA-532 and Promotes Oncogenicity of Clear Cell Renal Cell Carcinoma by Increasing ETS1 Expression. Cancer Manag. Res. 2020, 12, 2195–2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Stefano, L.; Jensen, M.R.; Helin, K. E2F7, a novel E2F featuring DP-independent repression of a subset of E2F-regulated genes. EMBO J. 2003, 22, 6289–6298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Jo, Y.H.; Jang, M.; Nguyen, N.N.Y.; Yun, H.R.; Ko, S.H.; Shin, Y.; Lee, J.S.; Kang, I.; Ha, J.; et al. PAC-5 Gene Expression Signature for Predicting Prognosis of Patients with Pancreatic Adenocarcinoma. Cancers 2019, 11, 1749. [Google Scholar] [CrossRef] [Green Version]
- Raman, P.; Maddipati, R.; Lim, K.H.; Tozeren, A. Pancreatic cancer survival analysis defines a signature that predicts outcome. PLoS ONE 2018, 13, e0201751. [Google Scholar] [CrossRef]
- Lu, T.; Wei, G.H.; Wang, J.; Shen, J. LncRNA CASC19 contributed to the progression of pancreatic cancer through modulating miR-148b/E2F7 axis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 10462–10471. [Google Scholar] [CrossRef]
- Xu, C.; Qi, X. MiR-10b inhibits migration and invasion of pancreatic ductal adenocarcinoma via regulating E2F7. J. Clin. Lab. Anal. 2020, 34, e23442. [Google Scholar] [CrossRef]
- Zhou, X.; Gao, W.; Hua, H.; Ji, Z. LncRNA-BLACAT1 Facilitates Proliferation, Migration and Aerobic Glycolysis of Pancreatic Cancer Cells by Repressing CDKN1C via EZH2-Induced H3K27me3. Front. Oncol. 2020, 10, 539805. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, Y.; Wang, Y.; Li, X.; Xiao, Y.; Wang, W. High Cancer Susceptibility Candidate 8 Expression Is Associated With Poor Prognosis of Pancreatic Adenocarcinoma: Validated Analysis Based on Four Cancer Databases. Front. Cell Dev. Biol. 2020, 8, 392. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, J.; Dong, L.; Xia, L.; Zhu, H.; Li, Z.; Yu, X. Long Noncoding RNA HCP5 Regulates Pancreatic Cancer Gemcitabine (GEM) Resistance By Sponging Hsa-miR-214-3p To Target HDGF. Onco. Targets Ther. 2019, 12, 8207–8216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Zhang, G.Q.; Chen, H.; Zhao, Z.J.; Chen, H.Z.; Liu, H.; Wang, G.; Jia, Y.H.; Pan, S.H.; Kong, R.; et al. Plasma and tumor levels of Linc-pint are diagnostic and prognostic biomarkers for pancreatic cancer. Oncotarget 2016, 7, 71773–71781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, L.; Dong, Y.; Wang, X.; Zhao, X.; Kong, C.; Liu, Y.; Jiang, X.; Zhang, X. Downregulation of long noncoding RNA SNHG1 inhibits cell proliferation, metastasis, and invasion by suppressing the Notch-1 signaling pathway in pancreatic cancer. J. Cell Biochem. 2019, 120, 6106–6112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Li, C.; Sun, Z. Long non-coding RNA LINC00346, LINC00578, LINC00673, LINC00671, LINC00261, and SNHG9 are novel prognostic markers for pancreatic cancer. Am. J. Transl. Res. 2018, 10, 2648–2658. [Google Scholar]
- Cao, W.; Zhou, G. LncRNA SNHG12 contributes proliferation, invasion and epithelial-mesenchymal transition of pancreatic cancer cells by absorbing miRNA-320b. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef]
- Guo, W.; Zhong, K.; Wei, H.; Nie, C.; Yuan, Z. Long non-coding RNA SPRY4-IT1 promotes cell proliferation and invasion by regulation of Cdc20 in pancreatic cancer cells. PLoS ONE 2018, 13, e0193483. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Zhang, Y.; Yang, J.; He, S.; Li, M.; Yan, S.; Chen, Y.; Qu, C.; Xu, L. NEAT1 regulates pancreatic cancer cell growth, invasion and migration though mircroRNA-335-5p/c-met axis. Am. J. Cancer Res. 2016, 6, 2361–2374. [Google Scholar] [PubMed]
- Zhang, H.; Zhu, C.; He, Z.; Chen, S.; Li, L.; Sun, C. LncRNA PSMB8-AS1 contributes to pancreatic cancer progression via modulating miR-382-3p/STAT1/PD-L1 axis. J. Exp. Clin. Cancer Res. 2020, 39, 179. [Google Scholar] [CrossRef]
- Li, N.; Yang, G.; Luo, L.; Ling, L.; Wang, X.; Shi, L.; Lan, J.; Jia, X.; Zhang, Q.; Long, Z.; et al. lncRNA THAP9-AS1 Promotes Pancreatic Ductal Adenocarcinoma Growth and Leads to a Poor Clinical Outcome via Sponging miR-484 and Interacting with YAP. Clin. Cancer Res. 2020, 26, 1736–1748. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yang, H.; Du, Y.; Liu, P.; Zhang, J.; Li, Y.; Shen, H.; Xing, L.; Xue, X.; Chen, J.; et al. Long noncoding RNA TP53TG1 promotes pancreatic ductal adenocarcinoma development by acting as a molecular sponge of microRNA-96. Cancer Sci. 2019, 110, 2760–2772. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Yang, D.; Zhang, L.; Lu, S.; Ye, J.; Li, M.; Hu, W. Linc-pint inhibits early stage pancreatic ductal adenocarcinoma growth through TGF-β pathway activation. Oncol. Lett. 2019, 17, 4633–4639. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Transcript ID | Effect of MAPK | p-Value for the Difference of Expression in Tumor (T) vs. Normal (N) in TCGA | p-Value (Log-Rank) for Overall Survival in TCGA | Expression Level Associated with Poor Prognosis |
|---|---|---|---|---|---|
| CCDC18-AS1 | NR_034089 | Downregulation | 0.61 (N > T) | 0.009 | Low |
| LINC00941 | NR_040245 | Upregulation | 3 × 10−6 (T > N) | 0.0008 | High |
| LINC-PINT | NR_015431 | Upregulation | 0.55 (N > T) | 0.01 | Low |
| NEAT1 | NR_028272 | Downregulation | 0.86 (N > T) | 0.8 | - |
| PRR7-AS1 | NR_038915 | Upregulation | 0.65 (N > T) | 0.07 | - |
| SNAI3-AS1 | NR_024399 | Downregulation | 0.29 (N > T) | 0.00001 | Low |
| SNHG4 | NR_003141 | Upregulation | 0.99 (T > N) | 0.7 | - |
| SPRY4-IT1 | NR_131221 | Upregulation | 0.22 (T > N) | 0.2 | - |
| Gene Symbol | Transcript ID | Effect of MAPK and LINC00941 | p-Value for the Difference of Expression in Tumor (T) vs. Normal (N) in TCGA | p-Value (Log-Rank) for Overall Survivalin TCGA | Expression Level Associated with Poor Prognosis |
|---|---|---|---|---|---|
| CD82 | NM_001024844 | Downregulation | 0.01 (T > N) | 0.04 | High |
| E2F7 | NM_203394 | Upregulation | 0.001 (T > N) | 0.05 | High |
| F3 | NM_001178096 | Downregulation | 0.36 (T > N) | 0.0001 | High |
| LOC100507291 | NR_121608 | Downregulation | 0.10 (T > N) | 0.3 | - |
| SPRY4-IT1 | NR_131221 | Upregulation | 0.22 (T > N) | 0.2 | - |
| TNNC1 | NM_003280 | Downregulation | 7 × 10−6 (T > N) | 0.5 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishikawa, T.; Fukushige, S.; Saiki, Y.; Hirose, K.; Hiyoshi, T.; Ogawa, T.; Katori, Y.; Furukawa, T. Long Non-Coding RNAs Associated with Mitogen-Activated Protein Kinase in Human Pancreatic Cancer. Cancers 2023, 15, 303. https://doi.org/10.3390/cancers15010303
Ishikawa T, Fukushige S, Saiki Y, Hirose K, Hiyoshi T, Ogawa T, Katori Y, Furukawa T. Long Non-Coding RNAs Associated with Mitogen-Activated Protein Kinase in Human Pancreatic Cancer. Cancers. 2023; 15(1):303. https://doi.org/10.3390/cancers15010303
Chicago/Turabian StyleIshikawa, Tomohiko, Shinichi Fukushige, Yuriko Saiki, Katsuya Hirose, Takako Hiyoshi, Takenori Ogawa, Yukio Katori, and Toru Furukawa. 2023. "Long Non-Coding RNAs Associated with Mitogen-Activated Protein Kinase in Human Pancreatic Cancer" Cancers 15, no. 1: 303. https://doi.org/10.3390/cancers15010303
APA StyleIshikawa, T., Fukushige, S., Saiki, Y., Hirose, K., Hiyoshi, T., Ogawa, T., Katori, Y., & Furukawa, T. (2023). Long Non-Coding RNAs Associated with Mitogen-Activated Protein Kinase in Human Pancreatic Cancer. Cancers, 15(1), 303. https://doi.org/10.3390/cancers15010303