

Prognostic Value of SGK1 and Bcl-2 in Invasive Breast Cancer
 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population and Tissue Microarray Construction
2.2. Immunohistochemical Staining
2.3. Immunohistochemical Scoring and Molecular Breast Tissue Subtyping
2.4. Statistical Analysis
3. Results
3.1. Baseline Characteristics of the BCCC Subcohort in the Tissue Microarray Study
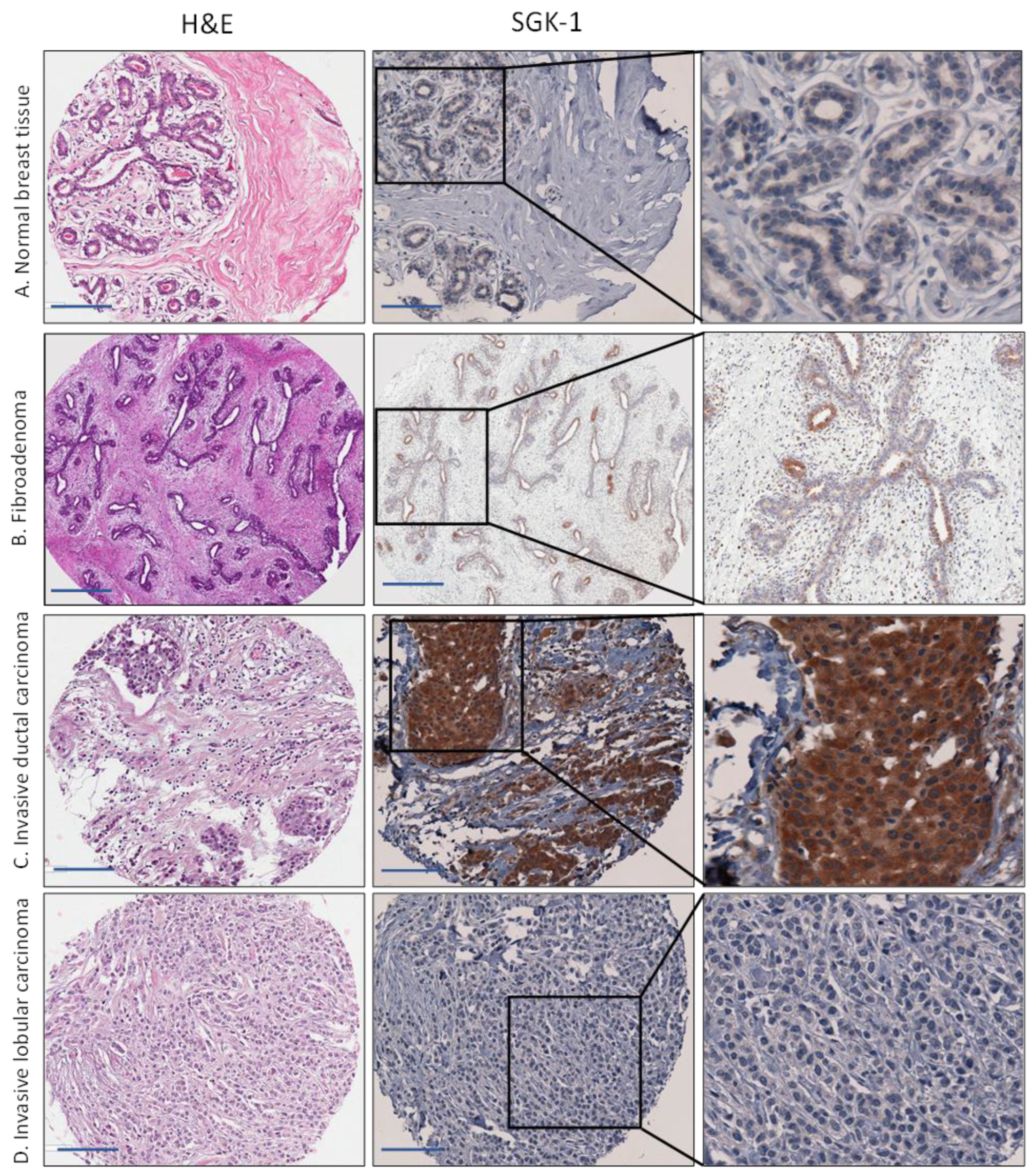
3.2. Increased Expression of SGK1 in Breast Cancer Tissue
3.3. Decreased Expression of Bcl-2 Expression in Breast Cancer
3.4. SGK1, Bcl-2 Expression, and Clinicopathological Characteristics of Breast Cancer
3.5. Expression of SGK1 and Bcl-2 and Breast Cancer Survival
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- AACR Cancer Disparities Progress Report. 2022. Available online: https://cancerprogressreport.aacr.org/disparities/ (accessed on 20 November 2022).
- Vilasco, M.; Communal, L.; Mourra, N.; Courtin, A.; Forgez, P.; Gompel, A. Glucocorticoid receptor and breast cancer. Breast Cancer Res. Treat. 2011, 130, 4446. [Google Scholar] [CrossRef] [PubMed]
- Reichardt, H.M.; Horsch, K.; Grone, H.J.; Kolbus, A.; Beug, H.; Hynes, N.; Schutz, G. Mammary gland development and lactation are controlled by different glucocorticoid receptor activities. Eur. J. Endocrinol. 2001, 145, 519–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaidya, J.S.; Baldassarre, G.; Thorat, M.A.; Massarut, S. Role of glucocorticoids in breast cancer. Curr. Pharm. Des. 2010, 16, 3593–3600. [Google Scholar] [CrossRef] [PubMed]
- Rosmond, R. The glucocorticoid receptor gene and its association to metabolic syndrome. Obes. Res. 2002, 10, 1078–1086. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.R. Glucocorticoids and cardiovascular disease. Eur. J. Endocrinol. 2007, 157, 545–559. [Google Scholar] [CrossRef] [Green Version]
- Donn, R.; Payne, D.; Ray, D. Glucocorticoid receptor gene polymorphisms and susceptibility to rheumatoid arthritis. Clin. Endocrinol. 2007, 67, 342–345. [Google Scholar] [CrossRef] [Green Version]
- Spijker, A.T.; van Rossum, E.F. Glucocorticoid receptor polymorphisms in major depression. Focus on glucocorticoid sensitivity and neurocognitive functioning. Ann. N. Y. Acad. Sci. 2009, 1179, 199–215. [Google Scholar] [CrossRef]
- Curran, J.E.; Lea, R.A.; Rutherford, S.; Weinstein, S.R.; Griffiths, L.R. Association of estrogen receptor and glucocorticoid receptor gene polymorphisms with sporadic breast cancer. Int. J. Cancer 2001, 95, 271–275. [Google Scholar] [CrossRef]
- Schmidt, S.; Rainer, J.; Ploner, C.; Presul, E.; Riml, S.; Kofler, R. Glucocorticoid-induced apoptosis and glucocorticoid resistance: Molecular mechanisms and clinical relevance. Cell Death Differ. 2004, 11, S45–S55. [Google Scholar] [CrossRef] [Green Version]
- Gruver-Yates, A.L.; Cidlowski, J.A. Tissue-specific actions of glucocorticoids on apoptosis: A double-edged sword. Cells 2013, 2, 202–223. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.J.; Pramyothin, P.; Karastergiou, K.; Fried, S.K. Deconstructing the roles of glucocorticoids in adipose tissue biology and the development of central obesity. Biochim. Biophys. Acta 2014, 1842, 473–481. [Google Scholar] [CrossRef] [Green Version]
- Al-Alem, U.; Mahmoud, A.M.; Batai, K.; Shah-Williams, E.; Gann, P.H.; Kittles, R.; Rauscher, G.H. Genetic Variation and Immunohistochemical Localization of the Glucocorticoid Receptor in Breast Cancer Cases from the Breast Cancer Care in Chicago Cohort. Cancers 2021, 13, 2261. [Google Scholar] [CrossRef]
- Tessier, M.; Woodgett, J.R. Serum and glucocorticoid-regulated protein kinases: Variations on a theme. J. Cell. Biochem. 2006, 98, 1391–1407. [Google Scholar] [CrossRef]
- Almawi, W.Y.; Melemedjian, O.K.; Jaoude, M.M. On the link between Bcl-2 family proteins and glucocorticoid-induced apoptosis. J. Leukoc. Biol. 2004, 76, 7–14. [Google Scholar] [CrossRef]
- Jang, H.; Park, Y.; Jang, J. Serum and glucocorticoid-regulated kinase 1: Structure, biological functions, and its inhibitors. Front. Pharmacol. 2022, 13, 1036844. [Google Scholar] [CrossRef]
- Ghani, M.J. SGK1, autophagy and cancer: An overview. Mol. Biol. Rep. 2022, 49, 675–685. [Google Scholar] [CrossRef]
- Wu, W.; Zou, M.; Brickley, D.R.; Pew, T.; Conzen, S.D. Glucocorticoid receptor activation signals through forkhead transcription factor 3a in breast cancer cells. Mol. Endocrinol. 2006, 20, 2304–2314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Chaudhuri, S.; Brickley, D.R.; Pang, D.; Karrison, T.; Conzen, S.D. Microarray analysis reveals glucocorticoid-regulated survival genes that are associated with inhibition of apoptosis in breast epithelial cells. Cancer Res. 2004, 64, 1757–1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callagy, G.M.; Pharoah, P.D.; Pinder, S.E.; Hsu, F.D.; Nielsen, T.O.; Ragaz, J.; Ellis, I.O.; Huntsman, D.; Caldas, C. Bcl-2 is a prognostic marker in breast cancer independently of the Nottingham Prognostic Index. Clin. Cancer Res. 2006, 12, 2468–2475. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, A.M.; Macias, V.; Al-Alem, U.; Deaton, R.J.; Kadjaksy-Balla, A.; Gann, P.H.; Rauscher, G.H. BRCA1 protein expression and subcellular localization in primary breast cancer: Automated digital microscopy analysis of tissue microarrays. PLoS ONE 2017, 12, e0184385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, F.; Shumilina, E. Regulation of ion channels by the serum- and glucocorticoid-inducible kinase SGK1. FASEB J. 2013, 27, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Li, J.; Sun, F.; Zhou, H.; Yang, J.; Yang, C. The functional duality of SGK1 in the regulation of hyperglycemia. Endocr. Connect. 2020, 9, R187–R194. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, S.; Brickley, D.R.; Kocherginsky, M.; Conzen, S.D. Coordinate expression of the PI3-kinase downstream effectors serum and glucocorticoid-induced kinase (SGK-1) and Akt-1 in human breast cancer. Eur. J. Cancer 2005, 41, 2754–2759. [Google Scholar] [CrossRef]
- Fagerli, U.M.; Ullrich, K.; Stuhmer, T.; Holien, T.; Kochert, K.; Holt, R.U.; Bruland, O.; Chatterjee, M.; Nogai, H.; Lenz, G.; et al. Serum/glucocorticoid-regulated kinase 1 (SGK1) is a prominent target gene of the transcriptional response to cytokines in multiple myeloma and supports the growth of myeloma cells. Oncogene 2011, 30, 3198–3206. [Google Scholar] [CrossRef] [Green Version]
- Szmulewitz, R.Z.; Chung, E.; Al-Ahmadie, H.; Daniel, S.; Kocherginsky, M.; Razmaria, A.; Zagaja, G.P.; Brendler, C.B.; Stadler, W.M.; Conzen, S.D. Serum/glucocorticoid-regulated kinase 1 expression in primary human prostate cancers. Prostate 2012, 72, 157–164. [Google Scholar] [CrossRef]
- Lang, F.; Cohen, P. Regulation and physiological roles of serum- and glucocorticoid-induced protein kinase isoforms. Sci. STKE 2001, 2001, re17. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Kong, P.; Zhang, S.; Zhang, L.; Cao, Y.; Duan, X.; Sun, T.; Tao, Z.; Liu, W. SGK1 in Human Cancer: Emerging Roles and Mechanisms. Front. Oncol. 2021, 10, 608722. [Google Scholar] [CrossRef]
- Zhang, L.; Cui, R.; Cheng, X.; Du, J. Antiapoptotic effect of serum and glucocorticoid-inducible protein kinase is mediated by novel mechanism activating I{kappa}B kinase. Cancer Res. 2005, 65, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Liu, L.; Zhang, C.; Zheng, T.; Wang, J.; Lin, M.; Zhao, Y.; Wang, X.; Levine, A.J.; Hu, W. Chronic restraint stress attenuates p53 function and promotes tumorigenesis. Proc. Natl. Acad. Sci. USA 2012, 109, 7013–7018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasir, O.; Wang, K.; Foller, M.; Gu, S.; Bhandaru, M.; Ackermann, T.F.; Boini, K.M.; Mack, A.; Klingel, K.; Amato, R.; et al. Relative resistance of SGK1 knockout mice against chemical carcinogenesis. IUBMB Life 2009, 61, 768–776. [Google Scholar] [CrossRef]
- Towhid, S.T.; Liu, G.-L.; Ackermann, T.F.; Beier, N.; Scholz, W.; Fuchß, T.; Toulany, M.; Rodemann, H.-P.; Lang, F. Inhibition of colonic tumor growth by the selective SGK inhibitor EMD638683. Cell. Physiol. Biochem. 2013, 32, 838–848. [Google Scholar] [CrossRef]
- Mason, J.; Davison-Versagli, C.; Leliaert, A.; Pape, D.; McCallister, C.; Zuo, J.; Durbin, S.; Buchheit, C.; Zhang, S.; Schafer, Z. Oncogenic Ras differentially regulates metabolism and anoikis in extracellular matrix-detached cells. Cell Death Differ. 2016, 23, 1271–1282. [Google Scholar] [CrossRef] [Green Version]
- Cicenas, J.; Meskinyte-Kausiliene, E.; Jukna, V.; Rimkus, A.; Simkus, J.; Soderholm, D. SGK1 in Cancer: Biomarker and Drug Target. Cancers 2022, 14, 2385. [Google Scholar] [CrossRef]
- Siddiqui, W.A.; Ahad, A.; Ahsan, H. The mystery of BCL2 family: Bcl-2 proteins and apoptosis: An update. Arch. Toxicol. 2015, 89, 289–317. [Google Scholar] [CrossRef]
- Lu, P.J.; Lu, Q.L.; Rughetti, A.; Taylor-Papadimitriou, J. Bcl-2 overexpression inhibits cell death and promotes the morphogenesis, but not tumorigenesis of human mammary epithelial cells. J. Cell Biol. 1995, 129, 1363–1378. [Google Scholar] [CrossRef] [Green Version]
- Callagy, G.M.; Webber, M.J.; Pharoah, P.D.; Caldas, C. Meta-analysis confirms BCL2 is an independent prognostic marker in breast cancer. BMC Cancer 2008, 8, 153. [Google Scholar] [CrossRef] [Green Version]
- Merino, D.; Lok, S.W.; Visvader, J.E.; Lindeman, G.J. Targeting BCL-2 to enhance vulnerability to therapy in estrogen receptor-positive breast cancer. Oncogene 2016, 35, 1877–1887. [Google Scholar] [CrossRef] [PubMed]
- Escórcio-Dourado, C.S.; Martins, L.M.; Simplício-Revoredo, C.M.; Sampaio, F.A.; Tavares, C.B.; da Silva-Sampaio, J.P.; Borges, U.S.; Alves-Ribeiro, F.A.; Lopes-Costa, P.V.; Lima-Dourado, J.C.; et al. Bcl-2 antigen expression in luminal A and triple-negative breast cancer. Med. Oncol. 2017, 34, 161. [Google Scholar] [CrossRef] [PubMed]
- Matias, A.T.; Jacinta-Fernandes, A.; Maia, A.-T.; Braga, S.; Jacinto, A.; Cabral, M.G.; Brito, P.H. Differential expression between African-ancestry and White patients diagnosed with Triple-Negative Breast Cancer: EGFR, Myc, Bcl2 and β-Catenin as ancestry-associated markers. bioRxiv 2020. [Google Scholar] [CrossRef]
- Martinez-Arribas, F.; Alvarez, T.; Del Val, G.; Martin-Garabato, E.; Nunez-Villar, M.J.; Lucas, R.; Sanchez, J.; Tejerina, A.; Schneider, J. Bcl-2 expression in breast cancer: A comparative study at the mRNA and protein level. Anticancer Res. 2007, 27, 219–222. [Google Scholar]
- Abdel-Fatah, T.M.A.; Perry, C.; Dickinson, P.; Ball, G.; Moseley, P.; Madhusudan, S.; Ellis, I.O.; Chan, S.Y.T. Bcl2 is an independent prognostic marker of triple negative breast cancer (TNBC) and predicts response to anthracycline combination (ATC) chemotherapy (CT) in adjuvant and neoadjuvant settings. Ann. Oncol. 2013, 24, 2801–2807. [Google Scholar] [CrossRef]
- Seong, M.K.; Lee, J.Y.; Byeon, J.; Sohn, Y.J.; Seol, H.; Lee, J.K.; Kim, E.K.; Kim, H.A.; Noh, W.C. Bcl-2 is a highly significant prognostic marker of hormone-receptor-positive, human epidermal growth factor receptor-2-negative breast cancer. Breast Cancer Res. Treat. 2015, 150, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Eom, Y.H.; Kim, H.S.; Lee, A.; Song, B.J.; Chae, B.J. BCL2 as a Subtype-Specific Prognostic Marker for Breast Cancer. J. Breast Cancer 2016, 19, 252–260. [Google Scholar] [CrossRef]
- Kawiak, A.; Kostecka, A. Regulation of Bcl-2 Family Proteins in Estrogen Receptor-Positive Breast Cancer and Their Implications in Endocrine Therapy. Cancers 2022, 14, 279. [Google Scholar] [CrossRef]
- Honma, N.; Horii, R.; Ito, Y.; Saji, S.; Younes, M.; Iwase, T.; Akiyama, F. Differences in clinical importance of Bcl-2 in breast cancer according to hormone receptors status or adjuvant endocrine therapy. BMC Cancer 2015, 15, 698. [Google Scholar] [CrossRef] [Green Version]
- Porporato, P.E.; Filigheddu, N.; Pedro, J.M.B.; Kroemer, G.; Galluzzi, L. Mitochondrial metabolism and cancer. Cell Res. 2018, 28, 265–280. [Google Scholar] [CrossRef] [PubMed]
- Rottenberg, H.; Hoek, J.B. The Mitochondrial Permeability Transition: Nexus of Aging, Disease and Longevity. Cells 2021, 10, 79. [Google Scholar] [CrossRef] [PubMed]
- Sanson, M.; Vu Hong, A.; Massourides, E.; Bourg, N.; Suel, L.; Amor, F.; Corre, G.; Benit, P.; Barthelemy, I.; Blot, S.; et al. miR-379 links glucocorticoid treatment with mitochondrial response in Duchenne muscular dystrophy. Sci. Rep. 2020, 10, 9139. [Google Scholar] [CrossRef]
- Mani, S.; Swargiary, G.; Singh, K.K. Natural Agents Targeting Mitochondria in Cancer. Int. J. Mol. Sci. 2020, 21, 6992. [Google Scholar] [CrossRef]
- Ruvolo, P.P.; Deng, X.; May, W.S. Phosphorylation of Bcl2 and regulation of apoptosis. Leukemia 2001, 15, 515–522. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Manufacturer | Host | Clone Number | Dilution | Retrieval Method |
|---|---|---|---|---|---|
| GCR | Lecia/Novocastra | Mouse | 4H2 | 1:25 | HIER |
| SGK1 | Novus/Biologicals | Rabbit | NB100-92054 | 1:50 | CC1 Mild |
| Bcl-2 | Cell Marque | Mouse | 124 | Predilute | CC1 Mild |
| ER | Ventana | Rabbit | SP1 | Predilute | CC1 Mild |
| PR | Ventana | Rabbit | 1E2 | Predilute | CC1 Mild |
| Her-2 | Ventana | Mouse | 4B5 | Predilute | CC1 Mild |
| CK 5/6 | DAKO | Mouse | D5 and 16B4 | 1:50 | HIER |
| EGFR | Ventana | Mouse | 3C6 | Predilute | CC1 Mild |
| % Cases | |
|---|---|
| Self-reported race/ethnicity (n = 280) | |
| nH Blacks | 40 |
| nH Whites | 30 |
| Hispanics | 30 |
| Age at diagnosis (n = 280) | |
| Less than 50 years | 31 |
| Equal or greater than 50 years | 69 |
| CDC BMI categories of BMI (n = 278) | |
| Normal weight (18.5–24.9 kg/m2) | 21 |
| Overweight (≥25 kg/m2) | 79 |
| Menopausal (n = 279) | |
| No | 17 |
| Yes | 83 |
| Histological subtypes (n = 258) | |
| Ductal carcinoma | 76 |
| Lobular carcinoma | 11 |
| Mixed ductal/lobular carcinoma and Other | 13 |
| Grade (n = 272) | |
| Low/intermediate | 61 |
| High | 39 |
| Stage (n = 277) | |
| 0,1 (early stage) | 42 |
| 2,3,4 (late stage) | 58 |
| Hormone receptor status (n = 276) | |
| ER- and PR- | 23 |
| ER+ and/or PR+ | 77 |
| Breast Tissue | N | SGK1 H Score a Mean, p-Value d | High SGK1 b %, p-Value e | N | Bcl-2 H Score a Mean, p-Value d | High Bcl-2 c %, p-Value e |
|---|---|---|---|---|---|---|
| Reduction mammoplasty | 24 | 20 | 25 | 21 | 2.0 | 100 |
| Fibroadenoma | 6 | 30 | 50 | 6 | 2.0 | 100 |
| Breast cancer tissue | 272 | 46 | 53 | 264 | 1.5 | 77 |
| p < 0.0001 | p = 0.029 | p = 0.038 | p = 0.018 | |||
| Histological subtypes | ||||||
| Ductal carcinoma | 195 | 47 | 54 | 195 | 1.4 | 71 |
| Lobular carcinoma | 28 | 29 | 39 | 28 | 1.7 | 93 |
| Mixed & Other | 35 | 52 | 57 | 35 | 1.8 | 85 |
| p = 0.1044 | p = 0.287 | p = 0.038 | p = 0.017 | |||
| Molecular subtypes | ||||||
| Luminal A | 179 | 41 | 49 | 178 | 2.0 | 96 |
| Luminal B | 14 | 44 | 57 | 14 | 1.2 | 79 |
| Triple Negative | 48 | 56 | 60 | 45 | 0.4 | 31 |
| Her2 | 21 | 56 | 67 | 21 | 0.1 | 14 |
| p = 0.123 | p = 0.25 | p < 0.0001 | p < 0.0001 | |||
| Hormone receptor status | ||||||
| ER- and PR- | 62 | 58 | 66 | 59 | 0.3 | 27 |
| ER+ and/or PR+ | 202 | 43 | 51 | 201 | 0.9 | 92 |
| p = 0.028 | p = 0.031 | p < 0.000 | p = 0.031 | |||
| Glucocorticoid Receptor status | ||||||
| Low (<17 H score) | 148 | 44 | 51 | 147 | 1.4 | 73 |
| High (≥17 H score) | 140 | 43 | 51 | 135 | 1.7 | 84 |
| p = 0.81 | p = 0.99 | p = 0.008 | p = 0.024 |
| Breast Tissue | N | SGK1 H-Score a Mean, p-Value | High SGK1 b %, p-Value | N | Bcl-2 H-Score a Mean, p-Value | High Bcl-2 c %, p-Value |
|---|---|---|---|---|---|---|
| Self-reported race/ethnicity | ||||||
| nH Blacks | 118 | 45 | 55 | 112 | 1.4 | 70 |
| nH Whites | 90 | 40 | 51 | 87 | 1.7 | 89 |
| Hispanics | 94 | 45 | 46 | 92 | 1.5 | 80 |
| p = 0.63 | p = 0.52 | p = 0.116 d | p = 0.005 e | |||
| Age at diagnosis | ||||||
| Less than 50 years | 84 | 52 | 56 | 82 | 1.4 | 71 |
| Equal or greater than 50 years | 188 | 43 | 52 | 182 | 1.5 | 79 |
| p = 0.129 | p = 0.56 | p = 0.32 | p = 0.137 | |||
| CDC categories of BMI | ||||||
| Normal weight (18.5–24.9 kg/m2) | 56 | 43 | 52 | 56 | 1.3 | 66 |
| Overweight/obese (≥25.0 kg/m2) | 214 | 47 | 54 | 206 | 1.5 | 79 |
| p = 0.59 | p = 0.79 | p = 0.147 | p = 0.042 | |||
| Menopausal | ||||||
| No | 46 | 45 | 48 | 46 | 1.4 | 70 |
| Yes | 225 | 46 | 55 | 217 | 1.5 | 78 |
| p = 0.828 | p = 0.39 | p = 0.373 | p = 0.227 | |||
| Stage at diagnosis | ||||||
| 0,1 (early stage) | 111 | 43 | 53 | 106 | 2 | 90 |
| 2,3,4 (late stage) | 158 | 47 | 53 | 155 | 1 | 68 |
| p = 0.999 | p = 0.483 | p < 0.0001 | p < 0.0001 | |||
| Histologic grade | ||||||
| Low/intermediate | 161 | 43 | 52 | 99 | 2 | 90 |
| High | 103 | 51 | 56 | 258 | 1 | 54 |
| p = 0.483 | p = 0.511 | p < 0.0001 | p < 0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Alem, U.; Rauscher, G.H.; Alem, Q.A.; Kajdacsy-Balla, A.; Mahmoud, A.M. Prognostic Value of SGK1 and Bcl-2 in Invasive Breast Cancer. Cancers 2023, 15, 3151. https://doi.org/10.3390/cancers15123151
Al-Alem U, Rauscher GH, Alem QA, Kajdacsy-Balla A, Mahmoud AM. Prognostic Value of SGK1 and Bcl-2 in Invasive Breast Cancer. Cancers. 2023; 15(12):3151. https://doi.org/10.3390/cancers15123151
Chicago/Turabian StyleAl-Alem, Umaima, Garth H. Rauscher, Qais Al Alem, Andre Kajdacsy-Balla, and Abeer M. Mahmoud. 2023. "Prognostic Value of SGK1 and Bcl-2 in Invasive Breast Cancer" Cancers 15, no. 12: 3151. https://doi.org/10.3390/cancers15123151
APA StyleAl-Alem, U., Rauscher, G. H., Alem, Q. A., Kajdacsy-Balla, A., & Mahmoud, A. M. (2023). Prognostic Value of SGK1 and Bcl-2 in Invasive Breast Cancer. Cancers, 15(12), 3151. https://doi.org/10.3390/cancers15123151