
Long Non-Coding RNAs and Metabolic Rewiring in Pancreatic Cancer


{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Epidemiology and Treatment
1.2. Metabolic Rewiring in Pancreatic Cancer
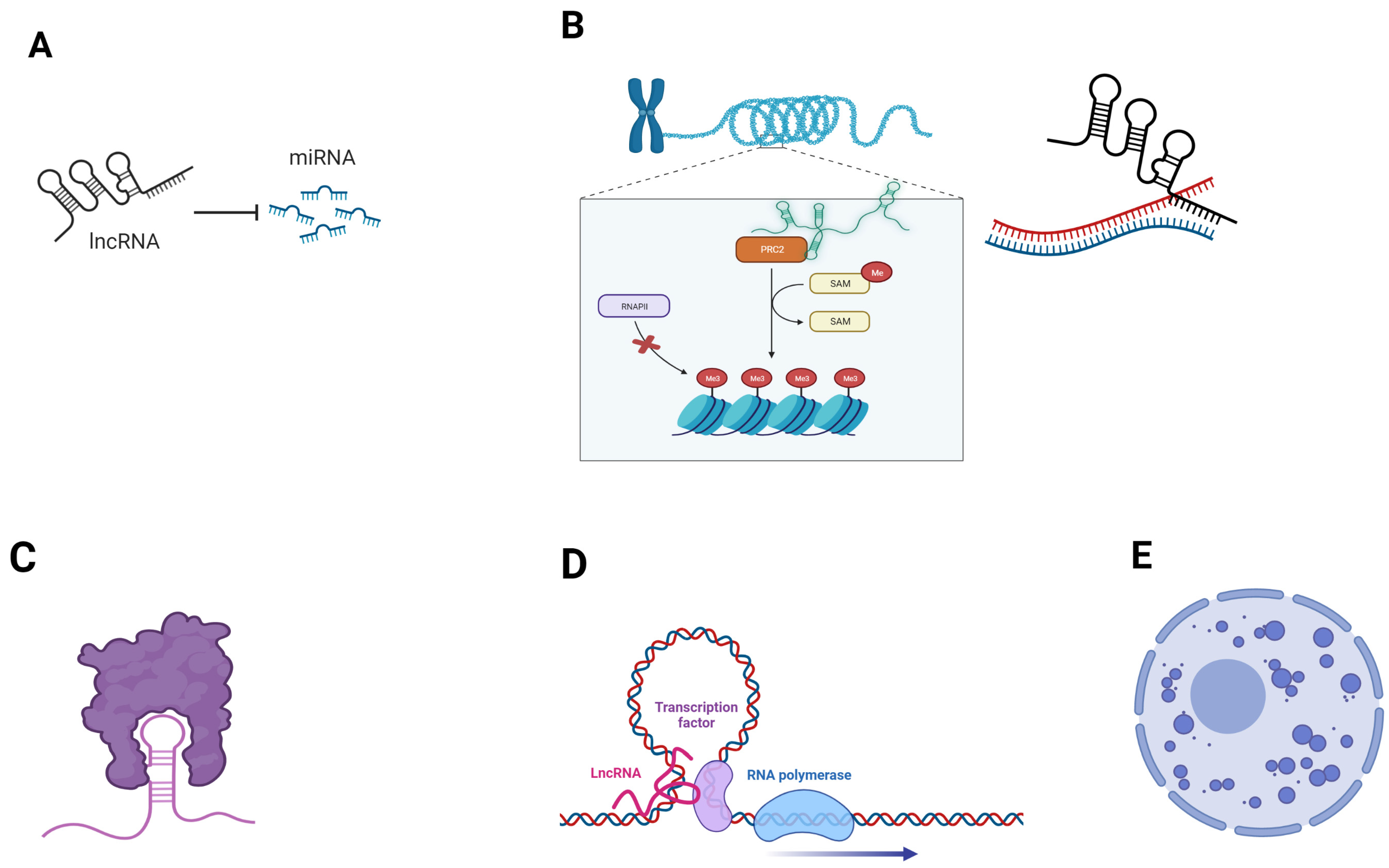
1.3. Long Non-Coding RNA Functions and Their Dysregulation in Cancer
1.4. Diagnostic, Prognostic and Therapeutic Potential of LncRNAs—State of the Art, Advances and Caveats
1.5. Search Strategy and Selection Criteria
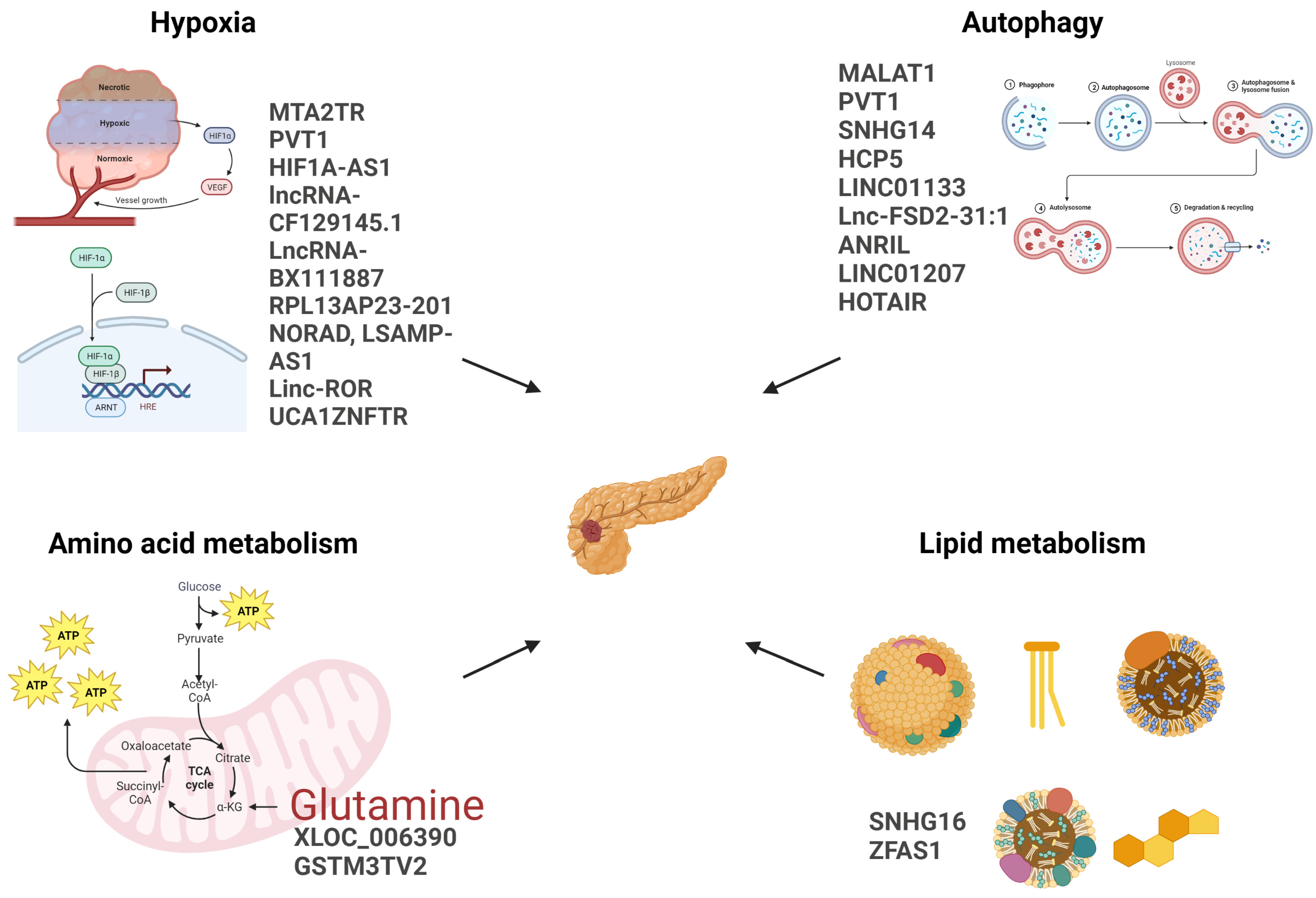
2. Hypoxia-Responsive lncRNA
2.1. MTA2TR
2.2. PVT1
2.3. HIF1A-AS1
2.4. LncRNA-CF129145.1
2.5. LncRNA-BX111887, ZEB-1AS1 and NR2F1-AS1
2.6. RPL13AP23-201
2.7. FEZF1-AS1
2.8. NORAD, LSAMP-AS1
2.9. Linc-ROR
2.10. UCA1
3. Autophagy-Related LncRNA
3.1. MALAT1/NEAT2
3.2. PVT1
3.3. SNHG14 and HCP5
3.4. Lnc-FSD2-31:1
3.5. ANRIL/CDKN2B-AS1
3.6. LINC01207 and LINC01133
3.7. HOTAIR
3.8. LncRNA Signatures
4. LncRNAs Implicated in Other Metabolic Pathways in PDAC
4.1. SNHG16
4.2. ZFAS1
4.3. XLOC_006390
4.4. GSTM3TV2
5. Conclusions and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Rahib, L.; Wehner, M.R.; Matrisian, L.M.; Nead, K.T. Estimated Projection of US Cancer Incidence and Death to 2040. JAMA Netw. Open 2021, 4, e214708. [Google Scholar] [CrossRef] [PubMed]
- Mizrahi, J.D.; Surana, R.; Valle, J.W.; Shroff, R.T. Pancreatic Cancer. Lancet 2020, 395, 2008–2020. [Google Scholar] [CrossRef]
- Lord, C.J.; Ashworth, A. PARP Inhibitors: Synthetic Lethality in the Clinic. Science 2017, 355, 1152–1158. [Google Scholar] [CrossRef]
- Dalmasso, B.; Puccini, A.; Catalano, F.; Borea, R.; Iaia, M.L.; Bruno, W.; Fornarini, G.; Sciallero, S.; Rebuzzi, S.E.; Ghiorzo, P. Beyond BRCA: The Emerging Significance of DNA Damage Response and Personalized Treatment in Pancreatic and Prostate Cancer Patients. Int. J. Mol. Sci. 2022, 23, 4709. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogunleye, A.O.; Nimmakayala, R.K.; Batra, S.K.; Ponnusamy, M.P. Metabolic Rewiring and Stemness: A Critical Attribute of Pancreatic Cancer Progression. Stem. Cells 2023, 41, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Li, Y.; Ma, M.; Wang, L.; Wang, H.; Liu, M.; Jiang, C. Adipocytes Promote Pancreatic Cancer Migration and Invasion through Fatty Acid Metabolic Reprogramming. Oncol. Rep. 2023, 50, 141. [Google Scholar] [CrossRef]
- Hu, T.; Shukla, S.K.; Vernucci, E.; He, C.; Wang, D.; King, R.J.; Jha, K.; Siddhanta, K.; Mullen, N.J.; Attri, K.S.; et al. Metabolic Rewiring by Loss of Sirt5 Promotes Kras-Induced Pancreatic Cancer Progression. Gastroenterology 2021, 161, 1584–1600. [Google Scholar] [CrossRef]
- Hypoxic and Ras-Transformed Cells Support Growth by Scavenging Unsaturated Fatty Acids from Lysophospholipids. Available online: https://www.pnas.org/doi/10.1073/pnas.1307237110?url_ver=Z39.88-2003&rfr_id=ori:rid:crossref.org&rfr_dat=cr_pub%20%200pubmed (accessed on 25 May 2023).
- Karasinska, J.M.; Topham, J.T.; Kalloger, S.E.; Jang, G.H.; Denroche, R.E.; Culibrk, L.; Williamson, L.M.; Wong, H.-L.; Lee, M.K.C.; O’Kane, G.M.; et al. Altered Gene Expression along the Glycolysis–Cholesterol Synthesis Axis Is Associated with Outcome in Pancreatic Cancer. Clin. Cancer Res. 2020, 26, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Mehla, K.; Singh, P.K. Metabolic Subtyping for Novel Personalized Therapies Against Pancreatic Cancer. Clin. Cancer Res. 2020, 26, 6–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, K.; Iwadate, D.; Kato, H.; Nakai, Y.; Tateishi, K.; Fujishiro, M. Targeting the Metabolic Rewiring in Pancreatic Cancer and Its Tumor Microenvironment. Cancers 2022, 14, 4351. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Yang, G.; Zhou, W.; Qiu, J.; Chen, G.; Luo, W.; Zhao, F.; You, L.; Zheng, L.; Zhang, T.; et al. Targeting Hypoxic Tumor Microenvironment in Pancreatic Cancer. J. Hematol. Oncol. 2021, 14, 14. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, C.; Liu, Y. The Role of Metabolic Reprogramming in Pancreatic Cancer Chemoresistance. Front. Pharmacol. 2023, 13, 1108776. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, Y.; Ikenaga, N.; Ohuchida, K.; Setoyama, D.; Irie, M.; Miura, D.; Wariishi, H.; Murata, M.; Mizumoto, K.; Hashizume, M.; et al. Mass Spectrometry-Based Metabolic Profiling of Gemcitabine-Sensitive and Gemcitabine-Resistant Pancreatic Cancer Cells. Pancreas 2014, 43, 311. [Google Scholar] [CrossRef]
- Halbrook, C.J.; Lyssiotis, C.A. Employing Metabolism to Improve the Diagnosis and Treatment of Pancreatic Cancer. Cancer Cell 2017, 31, 5–19. [Google Scholar] [CrossRef] [Green Version]
- Qin, C.; Yang, G.; Yang, J.; Ren, B.; Wang, H.; Chen, G.; Zhao, F.; You, L.; Wang, W.; Zhao, Y. Metabolism of Pancreatic Cancer: Paving the Way to Better Anticancer Strategies. Mol. Cancer 2020, 19, 50. [Google Scholar] [CrossRef] [Green Version]
- Gonzàlez-Porta, M.; Frankish, A.; Rung, J.; Harrow, J.; Brazma, A. Transcriptome Analysis of Human Tissues and Cell Lines Reveals One Dominant Transcript per Gene. Genome Biol. 2013, 14, R70. [Google Scholar] [CrossRef] [Green Version]
- Uszczynska-Ratajczak, B.; Lagarde, J.; Frankish, A.; Guigó, R.; Johnson, R. Towards a Complete Map of the Human Long Non-Coding RNA Transcriptome. Nat. Rev. Genet. 2018, 19, 535–548. [Google Scholar] [CrossRef]
- Deveson, I.W.; Brunck, M.E.; Blackburn, J.; Tseng, E.; Hon, T.; Clark, T.A.; Clark, M.B.; Crawford, J.; Dinger, M.E.; Nielsen, L.K.; et al. Universal Alternative Splicing of Noncoding Exons. Cell Syst. 2018, 6, 245–255.e5. [Google Scholar] [CrossRef] [Green Version]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.-L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long Non-Coding RNAs: Definitions, Functions, Challenges and Recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Z.; Jiang, J.; Xu, C.; Kang, J.; Xiao, L.; Wu, M.; Xiong, J.; Guo, X.; Liu, H. Endogenous MiRNA Sponge LincRNA-RoR Regulates Oct4, Nanog, and Sox2 in Human Embryonic Stem Cell Self-Renewal. Dev. Cell 2013, 25, 69–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, R.V.; Feretzaki, M.; Lingner, J. The Makings of TERRA R-Loops at Chromosome Ends. Cell Cycle 2021, 20, 1745–1759. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Norseen, J.; Wiedmer, A.; Riethman, H.; Lieberman, P.M. TERRA RNA Binding to TRF2 Facilitates Heterochromatin Formation and ORC Recruitment at Telomeres. Mol. Cell 2009, 35, 403–413. [Google Scholar] [CrossRef] [Green Version]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene Regulation by Long Non-Coding RNAs and Its Biological Functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Schmitz, S.U.; Grote, P.; Herrmann, B.G. Mechanisms of Long Noncoding RNA Function in Development and Disease. Cell. Mol. Life Sci. 2016, 73, 2491–2509. [Google Scholar] [CrossRef] [Green Version]
- Morriss, G.R.; Cooper, T.A. Protein Sequestration as a Normal Function of Long Noncoding RNAs and a Pathogenic Mechanism of RNAs Containing Nucleotide Repeat Expansions. Hum. Genet. 2017, 136, 1247–1263. [Google Scholar] [CrossRef]
- Bridges, M.C.; Daulagala, A.C.; Kourtidis, A. LNCcation: LncRNA Localization and Function. J. Cell Biol. 2021, 220, e202009045. [Google Scholar] [CrossRef]
- Peng, W.-X.; Koirala, P.; Mo, Y.-Y. LncRNA-Mediated Regulation of Cell Signaling in Cancer. Oncogene 2017, 36, 5661–5667. [Google Scholar] [CrossRef]
- Chan, J.J.; Tay, Y. Noncoding RNA:RNA Regulatory Networks in Cancer. Int. J. Mol. Sci. 2018, 19, 1310. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, J.; Zhong, Y.; Zhang, Y.; Yang, L.; Wu, P.; Hou, X.; Xiong, F.; Li, X.; Zhang, S.; Gong, Z.; et al. Long Non-Coding RNAs Are Involved in Alternative Splicing and Promote Cancer Progression. Br. J. Cancer 2022, 126, 1113–1124. [Google Scholar] [CrossRef]
- Tan, Y.-T.; Lin, J.-F.; Li, T.; Li, J.-J.; Xu, R.-H.; Ju, H.-Q. LncRNA-Mediated Posttranslational Modifications and Reprogramming of Energy Metabolism in Cancer. Cancer Commun. 2021, 41, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Micha, A.E.; Hähnel, S.; Friess, H.; Büchler, M.W.; Adler, G.; Gress, T.M. Genomic Imprinting of IGF-II and H19 in Adult Human Pancreatic Tissues. Digestion 1999, 60, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Raveh, E.; Matouk, I.J.; Gilon, M.; Hochberg, A. The H19 Long Non-Coding RNA in Cancer Initiation, Progression and Metastasis—A Proposed Unifying Theory. Mol. Cancer 2015, 14, 184. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Nong, K.; Zhu, H.; Wang, W.; Huang, X.; Yuan, Z.; Ai, K. H19 Promotes Pancreatic Cancer Metastasis by Derepressing Let-7’s Suppression on Its Target HMGA2-Mediated EMT. Tumor Biol. 2014, 35, 9163–9169. [Google Scholar] [CrossRef] [PubMed]
- Devi, K.T.R.; Karthik, D.; Mahendran, T.; Jaganathan, M.K.; Hemdev, S.P. Long Noncoding RNAs: Role and Contribution in Pancreatic Cancer. Transcription 2021, 12, 12–27. [Google Scholar] [CrossRef]
- Tumor-Derived Exosomal Long Noncoding RNA LINC01133, Regulated by Periostin, Contributes to Pancreatic Ductal Adenocarcinoma Epithelial-Mesenchymal Transition through the Wnt/β-Catenin Pathway by Silencing AXIN2|Oncogene. Available online: https://www.nature.com/articles/s41388-021-01762-0 (accessed on 25 May 2023).
- Lin, J.; Zhai, S.; Zou, S.; Xu, Z.; Zhang, J.; Jiang, L.; Deng, X.; Chen, H.; Peng, C.; Zhang, J.; et al. Positive Feedback between LncRNA FLVCR1-AS1 and KLF10 May Inhibit Pancreatic Cancer Progression via the PTEN/AKT Pathway. J. Exp. Clin. Cancer Res. 2021, 40, 316. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zheng, X.; Liang, W.; Jiang, C.; Su, D.; Fu, B. Long Noncoding RNA MIR600HG Binds to MicroRNA-125a-5p to Prevent Pancreatic Cancer Progression Via Mitochondrial Tumor Suppressor 1–Dependent Suppression of Extracellular Regulated Protein Kinases Signaling Pathway. Pancreas 2022, 51, 1434. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yue, G.; Li, M.; Yang, D.; Yue, C.; Hu, W.; Lu, H. LncRNA GAS8-AS1 Is a Novel Prognostic and Diagnostic Biomarker for Pancreatic Cancer. Crit. Rev. Eukaryot. 2022, 32, 83–92. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Fathi, M.; Zhai, T.; Taheri, M.; Dong, P. LncRNAs: Novel Biomarkers for Pancreatic Cancer. Biomolecules 2021, 11, 1665. [Google Scholar] [CrossRef]
- Chen, M.; Lu, Y.; Qin, S.; Hu, Z.; Chen, H.; Lu, L.; Mo, C.; Zhang, X.; Huang, J.; Qin, X. Long Intergenic Non-Coding 00162 as Diagnostic Biomarker for Early-Stage Pancreatic Cancer. Ann. Clin. Lab. Sci. 2022, 52, 533–543. [Google Scholar]
- Liu, Y.; Feng, W.; Liu, W.; Kong, X.; Li, L.; He, J.; Wang, D.; Zhang, M.; Zhou, G.; Xu, W.; et al. Circulating LncRNA ABHD11-AS1 Serves as a Biomarker for Early Pancreatic Cancer Diagnosis. J. Cancer 2019, 10, 3746–3756. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Chu, M.; Song, G.; Zuo, Z.; Han, Z.; Chen, C.; Li, Y.; Wang, Z.-W. Emerging Roles of Long Noncoding RNAs in Chemoresistance of Pancreatic Cancer. Semin. Cancer Biol. 2022, 83, 303–318. [Google Scholar] [CrossRef]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA Therapeutics—Challenges and Potential Solutions. Nat. Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef] [PubMed]
- Arun, G.; Diermeier, S.D.; Spector, D.L. Therapeutic Targeting of Long Non-Coding RNAs in Cancer. Trends Mol. Med. 2018, 24, 257–277. [Google Scholar] [CrossRef] [PubMed]
- Dizaji, B.F. Strategies to Target Long Non-Coding RNAs in Cancer Treatment: Progress and Challenges. Egypt. J. Med. Hum. Genet. 2020, 21, 41. [Google Scholar] [CrossRef]
- Infantino, V.; Santarsiero, A.; Convertini, P.; Todisco, S.; Iacobazzi, V. Cancer Cell Metabolism in Hypoxia: Role of HIF-1 as Key Regulator and Therapeutic Target. Int. J. Mol. Sci. 2021, 22, 5703. [Google Scholar] [CrossRef]
- FOLFIRINOX With Digoxin in Patients with Resectable Pancreatic Cancer. Available online: https://clinicaltrials.gov/study/NCT04141995?term=NCT04141995&rank=1 (accessed on 23 June 2023).
- Jiang, H.; Zhao, H.; Zhang, M.; He, Y.; Li, X.; Xu, Y.; Liu, X. Hypoxia Induced Changes of Exosome Cargo and Subsequent Biological Effects. Front. Immunol. 2022, 13, 824188. [Google Scholar] [CrossRef]
- Zeng, Z.; Xu, F.; Zheng, H.; Cheng, P.; Chen, Q.; Ye, Z.; Zhong, J.; Deng, S.; Liu, M.; Huang, K.; et al. LncRNA-MTA2TR Functions as a Promoter in Pancreatic Cancer via Driving Deacetylation-Dependent Accumulation of HIF-1α. Theranostics 2019, 9, 5298–5314. [Google Scholar] [CrossRef] [PubMed]
- Increased Expression of the lncRNA PVT1 Is Associated with Poor Prognosis in Pancreatic Cancer Patients. Minerva Med. 2015, 106, 143–149. Available online: https://www.minervamedica.it/it/riviste/minerva-medica/articolo.php?cod=R10Y2015N03A0143 (accessed on 25 May 2023).
- Zhu, Y.; Wu, F.; Gui, W.; Zhang, N.; Matro, E.; Zhu, L.; Eserberg, D.T.; Lin, X. A Positive Feedback Regulatory Loop Involving the LncRNA PVT1 and HIF-1α in Pancreatic Cancer. J. Mol. Cell Biol. 2021, 13, 676–689. [Google Scholar] [CrossRef] [PubMed]
- Leisegang, M.S.; Bains, J.K.; Seredinski, S.; Oo, J.A.; Krause, N.M.; Kuo, C.-C.; Günther, S.; Cetin, N.S.; Warwick, T.; Cao, C.; et al. HIF1α-AS1 Is a DNA:DNA:RNA Triplex-Forming LncRNA Interacting with the HUSH Complex. Nat. Commun. 2022, 13, 6563. [Google Scholar] [CrossRef]
- Xu, F.; Huang, M.; Chen, Q.; Niu, Y.; Hu, Y.; Hu, P.; Chen, D.; He, C.; Huang, K.; Zeng, Z.; et al. LncRNA HIF1A-AS1 Promotes Gemcitabine Resistance of Pancreatic Cancer by Enhancing Glycolysis through Modulating the AKT/YB1/HIF1α Pathway. Cancer Res. 2021, 81, 5678–5691. [Google Scholar] [CrossRef]
- Liu, M.; Zhong, J.; Zeng, Z.; Huang, K.; Ye, Z.; Deng, S.; Chen, H.; Xu, F.; Li, Q.; Zhao, G. Hypoxia-Induced Feedback of HIF-1α and LncRNA-CF129 Contributes to Pancreatic Cancer Progression through Stabilization of P53 Protein. Theranostics 2019, 9, 4795–4810. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Chen, H.; Ye, Z.; Deng, S.; Zhu, S.; Zeng, Z.; He, C.; Liu, M.; Huang, K.; Zhong, J.; et al. Hypoxia-Induced LncRNA-BX111 Promotes Metastasis and Progression of Pancreatic Cancer through Regulating ZEB1 Transcription. Oncogene 2018, 37, 5811–5828. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zhang, Z.; Yu, Q.; Zeng, Z.; Song, H.; Huang, X.; Kong, Q.; Hu, H.; Xia, Y. Positive Reciprocal Feedback of LncRNA ZEB1-AS1 and HIF-1α Contributes to Hypoxia-Promoted Tumorigenesis and Metastasis of Pancreatic Cancer. Front. Oncol. 2021, 11, 4897. [Google Scholar]
- Liu, Y.; Chen, S.; Cai, K.; Zheng, D.; Zhu, C.; Li, L.; Wang, F.; He, Z.; Yu, C.; Sun, C. Hypoxia-Induced Long Noncoding RNA NR2F1-AS1 Maintains Pancreatic Cancer Proliferation, Migration, and Invasion by Activating the NR2F1/AKT/MTOR Axis. Cell Death Dis. 2022, 13, 232. [Google Scholar] [CrossRef]
- Transcript: ENST00000480739.2 (RPL13AP23-201)-Summary-Homo_Sapiens-Ensembl Genome Browser 109. Available online: https://www.ensembl.org/Homo_sapiens/Transcript/Summary?db=core;g=ENSG00000242990;r=12:57674665-57675250;t=ENST00000480739 (accessed on 25 May 2023).
- Sun, Y.-W.; Chen, Y.-F.; Li, J.; Huo, Y.-M.; Liu, D.-J.; Hua, R.; Zhang, J.-F.; Liu, W.; Yang, J.-Y.; Fu, X.-L.; et al. A Novel Long Non-Coding RNA ENST00000480739 Suppresses Tumour Cell Invasion by Regulating OS-9 and HIF-1α in Pancreatic Ductal Adenocarcinoma. Br. J. Cancer 2014, 111, 2131–2141. [Google Scholar] [CrossRef] [Green Version]
- Ou, Z.-L.; Zhang, M.; Ji, L.-D.; Luo, Z.; Han, T.; Lu, Y.-B.; Li, Y.-X. Long Noncoding RNA FEZF1-AS1 Predicts Poor Prognosis and Modulates Pancreatic Cancer Cell Proliferation and Invasion through MiR-142/HIF-1α and MiR-133a/EGFR upon Hypoxia/Normoxia. J. Cell. Physiol. 2019, 234, 15407–15419. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, H.; Chen, C. Long Non-Coding RNA PCED1B-AS1 Promotes Pancreatic Ductal Adenocarcinoma Progression by Regulating the MiR-411-3p/HIF-1α Axis. Oncol. Rep. 2021, 46, 134. [Google Scholar] [CrossRef]
- Li, H.; Wang, X.; Wen, C.; Huo, Z.; Wang, W.; Zhan, Q.; Cheng, D.; Chen, H.; Deng, X.; Peng, C.; et al. Long Noncoding RNA NORAD, a Novel Competing Endogenous RNA, Enhances the Hypoxia-Induced Epithelial-Mesenchymal Transition to Promote Metastasis in Pancreatic Cancer. Mol. Cancer 2017, 16, 169. [Google Scholar] [CrossRef]
- Li, L.; Zou, W.; Xiao, Z.; Deng, Z.; Liu, R. Hypoxia-Induced Long Non-Coding RNA LSAMP-AS1 Regulates CeRNA Network to Predict Prognosis for Pancreatic Cancer. Comb. Chem. High Throughput Screen. 2023, 26, 2358–2371. [Google Scholar] [CrossRef]
- Sun, Z.; Sun, D.; Feng, Y.; Zhang, B.; Sun, P.; Zhou, B.; Du, L.; Wang, Y.; Fan, Z.; Yang, J.; et al. Exosomal Linc-ROR Mediates Crosstalk between Cancer Cells and Adipocytes to Promote Tumor Growth in Pancreatic Cancer. Mol. Ther. Nucleic Acids 2021, 26, 253–268. [Google Scholar] [CrossRef]
- Guo, Z.; Wang, X.; Yang, Y.; Chen, W.; Zhang, K.; Teng, B.; Huang, C.; Zhao, Q.; Qiu, Z. Hypoxic Tumor-Derived Exosomal Long Noncoding RNA UCA1 Promotes Angiogenesis via MiR-96-5p/AMOTL2 in Pancreatic Cancer. Mol. Ther. Nucleic Acids 2020, 22, 179–195. [Google Scholar] [CrossRef]
- Chi, Y.; Xin, H.; Liu, Z. Exosomal LncRNA UCA1 Derived From Pancreatic Stellate Cells Promotes Gemcitabine Resistance in Pancreatic Cancer via the SOCS3/EZH2 Axis. Front. Oncol. 2021, 11, 3724. [Google Scholar] [CrossRef]
- Li, W.; Han, S.; Hu, P.; Chen, D.; Zeng, Z.; Hu, Y.; Xu, F.; Tang, J.; Wang, F.; Zhao, Y.; et al. LncRNA ZNFTR Functions as an Inhibitor in Pancreatic Cancer by Modulating ATF3/ZNF24/VEGFA Pathway. Cell Death Dis. 2021, 12, 830. [Google Scholar] [CrossRef]
- Levy, J.M.M.; Towers, C.G.; Thorburn, A. Targeting Autophagy in Cancer. Nat. Rev. Cancer 2017, 17, 528–542. [Google Scholar] [CrossRef]
- White, E. Deconvoluting the Context-Dependent Role for Autophagy in Cancer. Nat. Rev. Cancer 2012, 12, 401–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onorati, A.V.; Dyczynski, M.; Ojha, R.; Amaravadi, R.K. Targeting Autophagy in Cancer. Cancer 2018, 124, 3307–3318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piffoux, M.; Eriau, E.; Cassier, P.A. Autophagy as a Therapeutic Target in Pancreatic Cancer. Br. J. Cancer 2021, 124, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Zeh, H.J.; Bahary, N.; Boone, B.A.; Singhi, A.D.; Miller-Ocuin, J.L.; Normolle, D.P.; Zureikat, A.H.; Hogg, M.E.; Bartlett, D.L.; Lee, K.K.; et al. A Randomized Phase II Preoperative Study of Autophagy Inhibition with High-Dose Hydroxychloroquine and Gemcitabine/Nab-Paclitaxel in Pancreatic Cancer Patients. Clin. Cancer Res. 2020, 26, 3126–3134. [Google Scholar] [CrossRef] [Green Version]
- Jahangiri, L.; Ishola, T.; Pucci, P.; Trigg, R.M.; Pereira, J.; Williams, J.A.; Cavanagh, M.L.; Gkoutos, G.V.; Tsaprouni, L.; Turner, S.D. The Role of Autophagy and LncRNAs in the Maintenance of Cancer Stem Cells. Cancers 2021, 13, 1239. [Google Scholar] [CrossRef]
- Goyal, B.; Yadav, S.R.M.; Awasthee, N.; Gupta, S.; Kunnumakkara, A.B.; Gupta, S.C. Diagnostic, Prognostic, and Therapeutic Significance of Long Non-Coding RNA MALAT1 in Cancer. Biochim. Biophys. Acta (BBA) Rev. Cancer 2021, 1875, 188502. [Google Scholar] [CrossRef]
- Pang, E.-J.; Yang, R.; Fu, X.; Liu, Y. Overexpression of Long Non-Coding RNA MALAT1 Is Correlated with Clinical Progression and Unfavorable Prognosis in Pancreatic Cancer. Tumor Biol. 2015, 36, 2403–2407. [Google Scholar] [CrossRef]
- Li, L.; Chen, H.; Gao, Y.; Wang, Y.-W.; Zhang, G.-Q.; Pan, S.-H.; Ji, L.; Kong, R.; Wang, G.; Jia, Y.-H.; et al. Long Noncoding RNA MALAT1 Promotes Aggressive Pancreatic Cancer Proliferation and Metastasis via the Stimulation of Autophagy. Mol. Cancer Ther. 2016, 15, 2232–2243. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Yi, C.; Yi, Y.; Qin, W.; Yan, Y.; Dong, X.; Zhang, X.; Huang, Y.; Zhang, R.; Wei, J.; et al. LncRNA PVT1 Promotes Gemcitabine Resistance of Pancreatic Cancer via Activating Wnt/β-Catenin and Autophagy Pathway through Modulating the MiR-619-5p/Pygo2 and MiR-619-5p/ATG14 Axes. Mol. Cancer 2020, 19, 118. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-F.; Luo, D.; Li, X.; Li, Z.-Q.; Yu, X.; Zhu, H.-W. PVT1 Knockdown Inhibits Autophagy and Improves Gemcitabine Sensitivity by Regulating the MiR-143/HIF-1α/VMP1 Axis in Pancreatic Cancer. Pancreas 2021, 50, 227. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, P.; Wang, C.; Xin, B. SNHG14 Enhances Gemcitabine Resistance by Sponging MiR-101 to Stimulate Cell Autophagy in Pancreatic Cancer. Biochem. Biophys. Res. Commun. 2019, 510, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, J.; Dong, L.; Xia, L.; Zhu, H.; Li, Z.; Yu, X. Long Noncoding RNA HCP5 Regulates Pancreatic Cancer Gemcitabine (GEM) Resistance By Sponging Hsa-MiR-214-3p To Target HDGF. OncoTargets Ther. 2019, 12, 8207–8216. [Google Scholar] [CrossRef] [Green Version]
- Geng, X.; Li, L.; Luo, Y.; Yang, W.; Hu, J.; Zhao, Z.; Cheng, C.; Zhang, T.; Zhang, Y.; Liu, L.; et al. Tumor Cell Derived Lnc-FSD2-31:1 Contributes to Cancer-Associated Fibroblasts Activation in Pancreatic Ductal Adenocarcinoma Progression through Extracellular Vesicles Cargo MiR-4736. Adv. Sci. 2023, 10, 2203324. [Google Scholar] [CrossRef]
- Wang, L.; Bi, R.; Li, L.; Zhou, K.; Yin, H. LncRNA ANRIL Aggravates the Chemoresistance of Pancreatic Cancer Cells to Gemcitabine by Targeting Inhibition of MiR-181a and Targeting HMGB1-Induced Autophagy. Aging 2021, 13, 19272–19281. [Google Scholar] [CrossRef]
- Dumartin, L.; Whiteman, H.J.; Weeks, M.E.; Hariharan, D.; Dmitrovic, B.; Iacobuzio-Donahue, C.A.; Brentnall, T.A.; Bronner, M.P.; Feakins, R.M.; Timms, J.F.; et al. AGR2 Is a Novel Surface Antigen That Promotes the Dissemination of Pancreatic Cancer Cells through Regulation of Cathepsins B and D. Cancer Res. 2011, 71, 7091–7102. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Wang, J.-O.; Zhou, W.-Y.; Chang, X.-Y.; Zhang, M.-M.; Zhang, Y.; Yang, X.-H. Long Non-Coding RNA LINC01207 Silencing Suppresses AGR2 Expression to Facilitate Autophagy and Apoptosis of Pancreatic Cancer Cells by Sponging MiR-143-5p. Mol. Cell. Endocrinol. 2019, 493, 110424. [Google Scholar] [CrossRef]
- Zhang, J.; Gao, S.; Zhang, Y.; Yi, H.; Xu, M.; Xu, J.; Liu, H.; Ding, Z.; He, H.; Wang, H.; et al. MiR-216a-5p Inhibits Tumorigenesis in Pancreatic Cancer by Targeting TPT1/MTORC1 and Is Mediated by LINC01133. Int. J. Biol. Sci. 2020, 16, 2612–2627. [Google Scholar] [CrossRef]
- Bae, S.-Y.; Byun, S.; Bae, S.H.; Min, D.S.; Woo, H.A.; Lee, K. TPT1 (Tumor Protein, Translationally-Controlled 1) Negatively Regulates Autophagy through the BECN1 Interactome and an MTORC1-Mediated Pathway. Autophagy 2017, 13, 820–833. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Yang, L.; Qi, X.; Wang, T.; Li, M.; Xu, K. Inhibition of Long Non-Coding RNA HOTAIR Enhances Radiosensitivity via Regulating Autophagy in Pancreatic Cancer. Cancer Manag. Res. 2018, 10, 5261–5271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Chi, H.; Gou, S.; Guo, X.; Li, L.; Peng, G.; Zhang, J.; Xu, J.; Nian, S.; Yuan, Q. An Aggrephagy-Related LncRNA Signature for the Prognosis of Pancreatic Adenocarcinoma. Genes 2023, 14, 124. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Fu, C.; Zeng, X.; Fan, X.; Wu, Y. An Independent Prognostic Model Based on Ten Autophagy-Related Long Noncoding RNAs in Pancreatic Cancer Patients. Genet. Res. 2022, 2022, e3895396. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Yang, G.; Long, J.; Yang, J.; Qin, C.; Luo, W.; Qiu, J.; Zhao, F.; You, L.; Zhang, T.; et al. Comprehensive Analysis of Autophagy-Associated LncRNAs Reveal Potential Prognostic Prediction in Pancreatic Cancer. Front. Oncol. 2021, 11, 596573. [Google Scholar] [CrossRef]
- Wei, D.-M.; Jiang, M.-T.; Lin, P.; Yang, H.; Dang, Y.-W.; Yu, Q.; Liao, D.-Y.; Luo, D.-Z.; Chen, G. Potential CeRNA Networks Involved in Autophagy Suppression of Pancreatic Cancer Caused by Chloroquine Diphosphate: A Study Based on Differentially-expressed CircRNAs, LncRNAs, MiRNAs and MRNAs. Int. J. Oncol. 2019, 54, 600–626. [Google Scholar]
- Yoon, H.; Shaw, J.L.; Haigis, M.C.; Greka, A. Lipid Metabolism in Sickness and in Health: Emerging Regulators of Lipotoxicity. Mol. Cell 2021, 81, 3708–3730. [Google Scholar] [CrossRef] [PubMed]
- Snaebjornsson, M.T.; Janaki-Raman, S.; Schulze, A. Greasing the Wheels of the Cancer Machine: The Role of Lipid Metabolism in Cancer. Cell Metab. 2020, 31, 62–76. [Google Scholar] [CrossRef] [PubMed]
- Kuzu, O.F.; Noory, M.A.; Robertson, G.P. The Role of Cholesterol in Cancer. Cancer Res. 2016, 76, 2063–2070. [Google Scholar] [CrossRef] [Green Version]
- Cholesterol Disruption in Combination With FOLFIRINOX in Patients with Advanced Pancreatic Adenocarcinoma. Available online: https://www.clinicaltrials.gov/study/NCT04862260?term=NCT04862260&rank=1 (accessed on 23 June 2023).
- Son, J.; Lyssiotis, C.A.; Ying, H.; Wang, X.; Hua, S.; Ligorio, M.; Perera, R.M.; Ferrone, C.R.; Mullarky, E.; Shyh-Chang, N.; et al. Glutamine Supports Pancreatic Cancer Growth through a KRAS-Regulated Metabolic Pathway. Nature 2013, 496, 101–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karner, C.M.; Esen, E.; Okunade, A.L.; Patterson, B.W.; Long, F. Increased Glutamine Catabolism Mediates Bone Anabolism in Response to WNT Signaling. J. Clin. Investig. 2015, 125, 551–562. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Luan, S.; Qi, C.; Tong, Q.; Yan, S.; Li, H.; Zhang, Y. Aspulvinone O, a Natural Inhibitor of GOT1 Suppresses Pancreatic Ductal Adenocarcinoma Cells Growth by Interfering Glutamine Metabolism. Cell Commun. Signal. 2019, 17, 111. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Yamasaki, S.; Kaneko, O.; Taoka, N.; Tomimoto, Y.; Namatame, I.; Yahata, T.; Kuromitsu, S.; Cantley, L.C.; Lyssiotis, C.A. A Covalent Small Molecule Inhibitor of Glutamate-Oxaloacetate Transaminase 1 Impairs Pancreatic Cancer Growth. Biochem. Biophys. Res. Commun. 2020, 522, 633–638. [Google Scholar] [CrossRef]
- Halama, A.; Suhre, K. Advancing Cancer Treatment by Targeting Glutamine Metabolism—A Roadmap. Cancers 2022, 14, 553. [Google Scholar] [CrossRef]
- Yu, Y.; Dong, J.-T.; He, B.; Zou, Y.-F.; Li, X.-S.; Xi, C.-H.; Yu, Y. LncRNA SNHG16 Induces the SREBP2 to Promote Lipogenesis and Enhance the Progression of Pancreatic Cancer. Future Oncol. 2019, 15, 3831–3844. [Google Scholar] [CrossRef]
- Madison, B.B. Srebp2: A Master Regulator of Sterol and Fatty Acid Synthesis. J. Lipid Res. 2016, 57, 333–335. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Ruan, Y.; Wu, X.; Zhou, X. LncRNA ZFAS1 Promotes HMGCR MRNA Stabilization via Binding U2AF2 to Modulate Pancreatic Carcinoma Lipometabolism. J. Immunol. Res. 2022, 2022, e4163198. [Google Scholar] [CrossRef]
- He, J.; Li, F.; Zhou, Y.; Hou, X.; Liu, S.; Li, X.; Zhang, Y.; Jing, X.; Yang, L. LncRNA XLOC_006390 Promotes Pancreatic Carcinogenesis and Glutamate Metabolism by Stabilizing C-Myc. Cancer Lett. 2020, 469, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Xiong, G.; Liu, C.; Yang, G.; Feng, M.; Xu, J.; Zhao, F.; You, L.; Zhou, L.; Zheng, L.; Hu, Y.; et al. Long Noncoding RNA GSTM3TV2 Upregulates LAT2 and OLR1 by Competitively Sponging Let-7 to Promote Gemcitabine Resistance in Pancreatic Cancer. J. Hematol. Oncol. 2019, 12, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Holst, J. L-Type Amino Acid Transport and Cancer: Targeting the MTORC1 Pathway to Inhibit Neoplasia. Am. J. Cancer Res. 2015, 5, 1281–1294. [Google Scholar]
- Murdocca, M.; De Masi, C.; Pucci, S.; Mango, R.; Novelli, G.; Di Natale, C.; Sangiuolo, F. LOX-1 and Cancer: An Indissoluble Liaison. Cancer Gene Ther. 2021, 28, 1088–1098. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dalmasso, B.; Ghiorzo, P. Long Non-Coding RNAs and Metabolic Rewiring in Pancreatic Cancer. Cancers 2023, 15, 3486. https://doi.org/10.3390/cancers15133486
Dalmasso B, Ghiorzo P. Long Non-Coding RNAs and Metabolic Rewiring in Pancreatic Cancer. Cancers. 2023; 15(13):3486. https://doi.org/10.3390/cancers15133486
Chicago/Turabian StyleDalmasso, Bruna, and Paola Ghiorzo. 2023. "Long Non-Coding RNAs and Metabolic Rewiring in Pancreatic Cancer" Cancers 15, no. 13: 3486. https://doi.org/10.3390/cancers15133486
APA StyleDalmasso, B., & Ghiorzo, P. (2023). Long Non-Coding RNAs and Metabolic Rewiring in Pancreatic Cancer. Cancers, 15(13), 3486. https://doi.org/10.3390/cancers15133486