
Targeted Alpha Therapy (TAT) with Single-Domain Antibodies (Nanobodies)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Antibody Vehicles for TαT
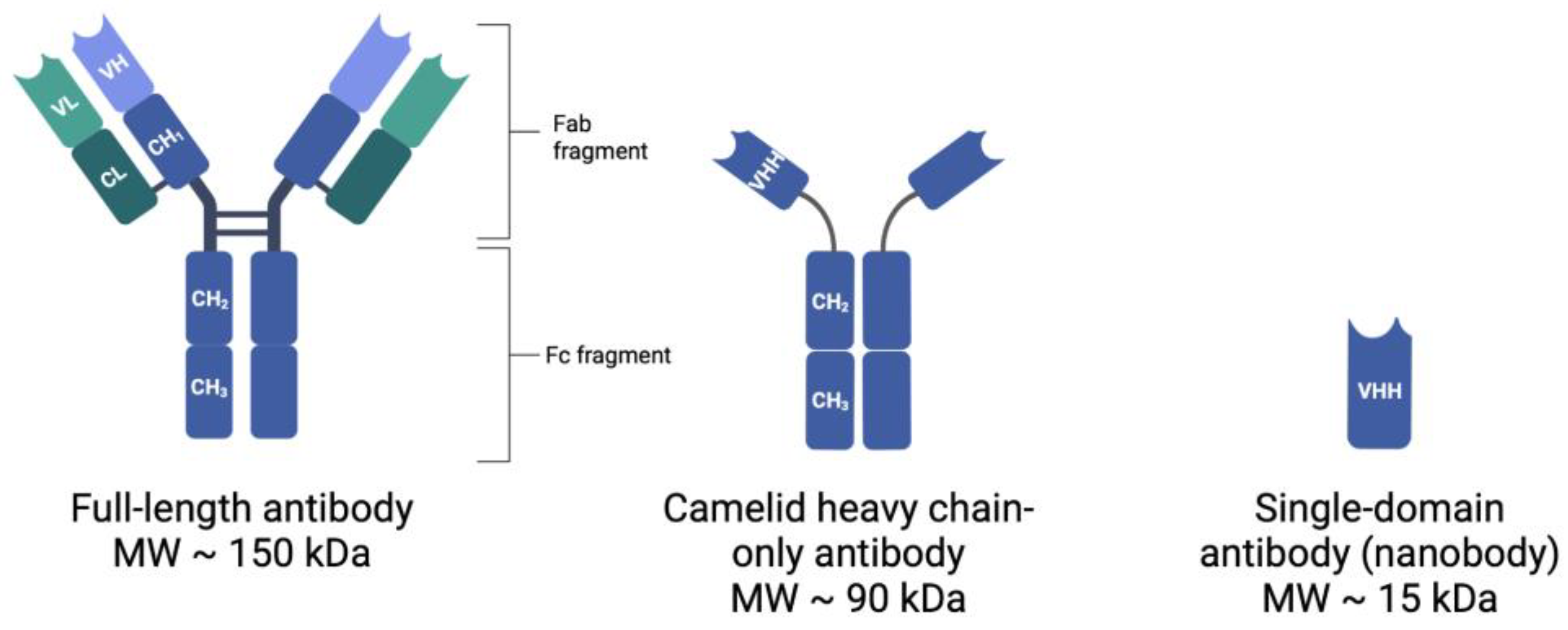
2.1. Full-Length Antibodies
2.2. Nanobodies
3. Nanobody Characteristics
3.1. High Stability
3.2. Improved Antigen Access and Binding
3.3. Low ‘Off-Target’ Effects and Immunogenicity
4. Nanobody Production
4.1. Immune or Naïve Production
4.2. Synthetic Production
4.3. Nanobody Library Selection Methods
4.4. Advantages of Nanobody Production
5. Radiolabeling Nanobodies
5.1. Direct Labeling
5.2. Indirect Labeling with a Chelator Group
5.3. Indirect Labeling with a Prosthetic Group
6. Use of Radiolabeled Nanobodies
6.1. Immunoscintigraphy
6.2. Therapeutic Purposes
Radiation Types for Targeted Radionuclide Therapy
7. Targeted Alpha Therapy
Nanobody-Based Targeted Alpha Therapy
8. Theranostics
9. Considerations for Nanobody-Based Targeted Alpha Therapy
10. Future Perspectives
11. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Global Cancer Observatory: Cancer Tomorrow. Available online: https://gco.iarc.fr/tomorrow/en/dataviz/isotype (accessed on 19 June 2023).
- Adler, M.J.; Dimitrov, D.S. Therapeutic Antibodies Against Cancer. Hematol. Oncol. Clin. N. Am. 2012, 26, 447–481. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Pandit, A. Therapeutic Monoclonal Antibodies Approved by FDA in 2022. J. Clin. Exp. Immunol. 2023, 8, 533–535. [Google Scholar]
- Kang, W.; Ding, C.; Zheng, D.; Ma, X.; Yi, L.; Tong, X.; Wu, C.; Xue, C.; Yu, Y.; Zhou, Q. Nanobody Conjugates for Targeted Cancer Therapy and Imaging. Technol. Cancer Res. Treat. 2021, 20, 153303382110101. [Google Scholar] [CrossRef]
- Brechbiel, M.W. Targeted α-Therapy: Past, Present, Future? Dalton Trans. 2007, 43, 4918. [Google Scholar] [CrossRef] [Green Version]
- Koenig, P.-A.; Das, H.; Liu, H.; Kümmerer, B.M.; Gohr, F.N.; Jenster, L.-M.; Schiffelers, L.D.J.; Tesfamariam, Y.M.; Uchima, M.; Wuerth, J.D.; et al. Structure-Guided Multivalent Nanobodies Block SARS-CoV-2 Infection and Suppress Mutational Escape. Science 2021, 371, eabe6230. [Google Scholar] [CrossRef] [PubMed]
- Güttler, T.; Aksu, M.; Dickmanns, A.; Stegmann, K.M.; Gregor, K.; Rees, R.; Taxer, W.; Rymarenko, O.; Schünemann, J.; Dienemann, C.; et al. Neutralization of SARS-CoV-2 by Highly Potent, Hyperthermostable, and Mutation-tolerant Nanobodies. EMBO J. 2021, 40, e107985. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Nambulli, S.; Xiao, Z.; Liu, H.; Sang, Z.; Duprex, W.P.; Schneidman-Duhovny, D.; Zhang, C.; Shi, Y. Versatile and Multivalent Nanobodies Efficiently Neutralize SARS-CoV-2. Science 2020, 370, 1479–1484. [Google Scholar] [CrossRef]
- Schoof, M.; Faust, B.; Saunders, R.A.; Sangwan, S.; Rezelj, V.; Hoppe, N.; Boone, M.; Billesbølle, C.B.; Puchades, C.; Azumaya, C.M.; et al. An Ultrapotent Synthetic Nanobody Neutralizes SARS-CoV-2 by Stabilizing Inactive Spike. Science 2020, 370, 1473–1479. [Google Scholar] [CrossRef]
- Nelson, B.J.B.; Andersson, J.D.; Wuest, F. Targeted Alpha Therapy: Progress in Radionuclide Production, Radiochemistry, and Applications. Pharmaceutics 2020, 13, 49. [Google Scholar] [CrossRef]
- Institute of Medicine. Advancing Nuclear Medicine through Innovation; National Academies Press: Washington, DC, USA, 2007; p. 11985. ISBN 978-0-309-11067-9. [Google Scholar]
- Targeted Alpha Therapy Working Group; Parker, C.; Lewington, V.; Shore, N.; Kratochwil, C.; Levy, M.; Lindén, O.; Noordzij, W.; Park, J.; Saad, F. Targeted Alpha Therapy, an Emerging Class of Cancer Agents: A Review. JAMA Oncol. 2018, 4, 1765. [Google Scholar] [CrossRef]
- Friedberg, J.W.; Fisher, R.I. Iodine-131 Tositumomab (Bexxar®): Radioimmunoconjugate Therapy for Indolent and Transformed B-Cell Non-Hodgkin’s Lymphoma. Expert Rev. Anticancer Ther. 2004, 4, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Jurcic, J.G. Targeted Alpha-Particle Therapy for Hematologic Malignancies. Semin. Nucl. Med. 2020, 50, 152–161. [Google Scholar] [CrossRef] [PubMed]
- McGann, S.; Horton, E.R. Radium-223 Dichloride: A Novel Treatment Option for Castration-Resistant Prostate Cancer Patients With Symptomatic Bone Metastases. Ann. Pharmacother. 2015, 49, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Gasser, M.; Waaga-Gasser, A.M. Therapeutic Antibodies in Cancer Therapy. In Protein Targeting Compounds; Böldicke, T., Ed.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2016; Volume 917, pp. 95–120. ISBN 978-3-319-32804-1. [Google Scholar]
- Muyldermans, S.; Smider, V.V. Distinct Antibody Species: Structural Differences Creating Therapeutic Opportunities. Curr. Opin. Immunol. 2016, 40, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Dekempeneer, Y.; Keyaerts, M.; Krasniqi, A.; Puttemans, J.; Muyldermans, S.; Lahoutte, T.; D’huyvetter, M.; Devoogdt, N. Targeted Alpha Therapy Using Short-Lived Alpha-Particles and the Promise of Nanobodies as Targeting Vehicle. Expert Opin. Biol. Ther. 2016, 16, 1035–1047. [Google Scholar] [CrossRef] [Green Version]
- Jurcic, J.G.; Larson, S.M.; Sgouros, G.; McDevitt, M.R.; Finn, R.D.; Divgi, C.R.; Ballangrud, A.M.; Hamacher, K.A.; Ma, D.; Humm, J.L.; et al. Targeted Alpha Particle Immunotherapy for Myeloid Leukemia. Blood 2002, 100, 1233–1239. [Google Scholar] [CrossRef] [Green Version]
- Rosenblat, T.L.; McDevitt, M.R.; Mulford, D.A.; Pandit-Taskar, N.; Divgi, C.R.; Panageas, K.S.; Heaney, M.L.; Chanel, S.; Morgenstern, A.; Sgouros, G.; et al. Sequential Cytarabine and α-Particle Immunotherapy with Bismuth-213–Lintuzumab (HuM195) for Acute Myeloid Leukemia. Clin. Cancer Res. 2010, 16, 5303–5311. [Google Scholar] [CrossRef] [Green Version]
- Holliger, P.; Hudson, P.J. Engineered Antibody Fragments and the Rise of Single Domains. Nat. Biotechnol. 2005, 23, 1126–1136. [Google Scholar] [CrossRef]
- Adams, G.P.; Shaller, C.C.; Chappell, L.L.; Wu, C.; Horak, E.M.; Simmons, H.H.; Litwin, S.; Marks, J.D.; Weiner, L.M.; Brechbiel, M.W. Delivery of the α-Emitting Radioisotope Bismuth-213 to Solid Tumors via Single-Chain Fv and Diabody Molecules. Nucl. Med. Biol. 2000, 27, 339–346. [Google Scholar] [CrossRef]
- Robinson, M.K.; Shaller, C.; Garmestani, K.; Plascjak, P.S.; Hodge, K.M.; Yuan, Q.-A.; Marks, J.D.; Waldmann, T.A.; Brechbiel, M.W.; Adams, G.P. Effective Treatment of Established Human Breast Tumor Xenografts in Immunodeficient Mice with a Single Dose of the α-Emitting Radioisotope Astatine-211 Conjugated to Anti-HER2/ Neu Diabodies. Clin. Cancer Res. 2008, 14, 875–882. [Google Scholar] [CrossRef] [Green Version]
- Hamers-Casterman, C.; Atarhouch, T.; Muyldermans, S.; Robinson, G.; Hammers, C.; Songa, E.B.; Bendahman, N.; Hammers, R. Naturally Occurring Antibodies Devoid of Light Chains. Nature 1993, 363, 446–448. [Google Scholar] [CrossRef] [PubMed]
- Arbabi-Ghahroudi, M. Camelid Single-Domain Antibodies: Historical Perspective and Future Outlook. Front. Immunol. 2017, 8, 1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, C.-Y.; Secombes, C.J.; Porter, A.J. Rapid Isolation of IgNAR Variable Single-Domain Antibody Fragments from a Shark Synthetic Library. Mol. Immunol. 2007, 44, 656–665. [Google Scholar] [CrossRef]
- Muyldermans, S. Single Domain Camel Antibodies: Current Status. Rev. Mol. Biotechnol. 2001, 74, 277–302. [Google Scholar] [CrossRef]
- Muyldermans, S. Nanobodies: Natural Single-Domain Antibodies. Annu. Rev. Biochem. 2013, 82, 775–797. [Google Scholar] [CrossRef] [Green Version]
- NANOBODY Technology Platform. Available online: https://www.sanofi.com/en/science-and-innovation/research-and-development/technology-platforms/nanobody-technology-platform (accessed on 24 August 2022).
- Hosseindokht, M.; Bakherad, H.; Zare, H. Nanobodies: A Tool to Open New Horizons in Diagnosis and Treatment of Prostate Cancer. Cancer Cell Int. 2021, 21, 580. [Google Scholar] [CrossRef]
- Nguyen, V.K.; Muyldermans, S.; Hamers, R. The Specific Variable Domain of Camel Heavy-Chain Antibodies Is Encoded in the Germline. J. Mol. Biol. 1998, 275, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Muyldermans, S.; Atarhouch, T.; Saldanha, J.; Barbosa, J.A.R.G.; Hamers, R. Sequence and Structure of VH Domain from Naturally Occurring Camel Heavy Chain Immunoglobulins Lacking Light Chains. Protein Eng. Des. Sel. 1994, 7, 1129–1135. [Google Scholar] [CrossRef]
- Hoey, R.J.; Eom, H.; Horn, J.R. Structure and Development of Single Domain Antibodies as Modules for Therapeutics and Diagnostics. Exp. Biol. Med. 2019, 244, 1568–1576. [Google Scholar] [CrossRef]
- van der Linden, R.H.J.; Frenken, L.G.J.; de Geus, B.; Harmsen, M.M.; Ruuls, R.C.; Stok, W.; de Ron, L.; Wilson, S.; Davis, P.; Verrips, C.T. Comparison of Physical Chemical Properties of Llama VHH Antibody Fragments and Mouse Monoclonal Antibodies. Biochim. Biophys. Acta BBA Protein Struct. Mol. Enzymol. 1999, 1431, 37–46. [Google Scholar] [CrossRef]
- Ewert, S.; Cambillau, C.; Conrath, K.; Plückthun, A. Biophysical Properties of Camelid V HH Domains Compared to Those of Human V H 3 Domains. Biochemistry 2002, 41, 3628–3636. [Google Scholar] [CrossRef]
- Pérez, J.M.J.; Renisio, J.G.; Prompers, J.J.; van Platerink, C.J.; Cambillau, C.; Darbon, H.; Frenken, L.G.J. Thermal Unfolding of a Llama Antibody Fragment: A Two-State Reversible Process. Biochemistry 2001, 40, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Kunz, P.; Zinner, K.; Mücke, N.; Bartoschik, T.; Muyldermans, S.; Hoheisel, J.D. The Structural Basis of Nanobody Unfolding Reversibility and Thermoresistance. Sci. Rep. 2018, 8, 7934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmsen, M.M.; van Solt, C.B.; van Zijderveld-van Bemmel, A.M.; Niewold, T.A.; van Zijderveld, F.G. Selection and Optimization of Proteolytically Stable Llama Single-Domain Antibody Fragments for Oral Immunotherapy. Appl. Microbiol. Biotechnol. 2006, 72, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Bakherad, H.; Mousavi Gargari, S.L.; Rasooli, I.; RajabiBazl, M.; Mohammadi, M.; Ebrahimizadeh, W.; Safaee Ardakani, L.; Zare, H. In Vivo Neutralization of Botulinum Neurotoxins Serotype E with Heavy-Chain Camelid Antibodies (VHH). Mol. Biotechnol. 2013, 55, 159–167. [Google Scholar] [CrossRef]
- Hussack, G.; Hirama, T.; Ding, W.; MacKenzie, R.; Tanha, J. Engineered Single-Domain Antibodies with High Protease Resistance and Thermal Stability. PLoS ONE 2011, 6, e28218. [Google Scholar] [CrossRef] [Green Version]
- Pillay, T.S.; Muyldermans, S. Application of Single-Domain Antibodies (“Nanobodies”) to Laboratory Diagnosis. Ann. Lab. Med. 2021, 41, 549–558. [Google Scholar] [CrossRef]
- Mitchell, L.S.; Colwell, L.J. Comparative Analysis of Nanobody Sequence and Structure Data. Proteins Struct. Funct. Bioinforma. 2018, 86, 697–706. [Google Scholar] [CrossRef]
- Desmyter, A.; Transue, T.R.; Ghahroudi, M.A.; Dao Thi, M.-H.; Poortmans, F.; Hamers, R.; Muyldermans, S.; Wyns, L. Crystal Structure of a Camel Single-Domain VH Antibody Fragment in Complex with Lysozyme. Nat. Struct. Mol. Biol. 1996, 3, 803–811. [Google Scholar] [CrossRef]
- Transue, T.R.; De Genst, E.; Ghahroudi, M.A.; Wyns, L.; Muyldermans, S. Camel Single-Domain Antibody Inhibits Enzyme by Mimicking Carbohydrate Substrate. Proteins Struct. Funct. Genet. 1998, 32, 515–522. [Google Scholar] [CrossRef]
- Chatalic, K.L.S.; Veldhoven-Zweistra, J.; Bolkestein, M.; Hoeben, S.; Koning, G.A.; Boerman, O.C.; de Jong, M.; van Weerden, W.M. A Novel 111 In-Labeled Anti–Prostate-Specific Membrane Antigen Nanobody for Targeted SPECT/CT Imaging of Prostate Cancer. J. Nucl. Med. 2015, 56, 1094–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoefman, S.; Ottevaere, I.; Baumeister, J.; Sargentini-Maier, M. Pre-Clinical Intravenous Serum Pharmacokinetics of Albumin Binding and Non-Half-Life Extended Nanobodies®. Antibodies 2015, 4, 141–156. [Google Scholar] [CrossRef] [Green Version]
- Ackaert, C.; Smiejkowska, N.; Xavier, C.; Sterckx, Y.G.J.; Denies, S.; Stijlemans, B.; Elkrim, Y.; Devoogdt, N.; Caveliers, V.; Lahoutte, T.; et al. Immunogenicity Risk Profile of Nanobodies. Front. Immunol. 2021, 12, 632687. [Google Scholar] [CrossRef]
- Vincke, C.; Loris, R.; Saerens, D.; Martinez-Rodriguez, S.; Muyldermans, S.; Conrath, K. General Strategy to Humanize a Camelid Single-Domain Antibody and Identification of a Universal Humanized Nanobody Scaffold. J. Biol. Chem. 2009, 284, 3273–3284. [Google Scholar] [CrossRef] [Green Version]
- Drabek, D.; Janssens, R.; de Boer, E.; Rademaker, R.; Kloess, J.; Skehel, J.; Grosveld, F. Expression Cloning and Production of Human Heavy-Chain-Only Antibodies from Murine Transgenic Plasma Cells. Front. Immunol. 2016, 7, 619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muyldermans, S. A Guide to: Generation and Design of Nanobodies. FEBS J. 2021, 288, 2084–2102. [Google Scholar] [CrossRef] [PubMed]
- Verheesen, P.; Roussis, A.; de Haard, H.J.; Groot, A.J.; Stam, J.C.; den Dunnen, J.T.; Frants, R.R.; Verkleij, A.J.; Theo Verrips, C.; van der Maarel, S.M. Reliable and Controllable Antibody Fragment Selections from Camelid Non-Immune Libraries for Target Validation. Biochim. Biophys. Acta BBA Proteins Proteom. 2006, 1764, 1307–1319. [Google Scholar] [CrossRef]
- Zimmermann, I.; Egloff, P.; Hutter, C.A.; Arnold, F.M.; Stohler, P.; Bocquet, N.; Hug, M.N.; Huber, S.; Siegrist, M.; Hetemann, L.; et al. Synthetic Single Domain Antibodies for the Conformational Trapping of Membrane Proteins. eLife 2018, 7, e34317. [Google Scholar] [CrossRef]
- Sabir, J.S.M.; Atef, A.; El-Domyati, F.M.; Edris, S.; Hajrah, N.; Alzohairy, A.M.; Bahieldin, A. Construction of Naïve Camelids VHH Repertoire in Phage Display-Based Library. C. R. Biol. 2014, 337, 244–249. [Google Scholar] [CrossRef]
- Zimmermann, I.; Egloff, P.; Hutter, C.A.J.; Kuhn, B.T.; Bräuer, P.; Newstead, S.; Dawson, R.J.P.; Geertsma, E.R.; Seeger, M.A. Generation of Synthetic Nanobodies against Delicate Proteins. Nat. Protoc. 2020, 15, 1707–1741. [Google Scholar] [CrossRef]
- Liu, B.; Yang, D. Easily Established and Multifunctional Synthetic Nanobody Libraries as Research Tools. Int. J. Mol. Sci. 2022, 23, 1482. [Google Scholar] [CrossRef] [PubMed]
- Saerens, D.; Pellis, M.; Loris, R.; Pardon, E.; Dumoulin, M.; Matagne, A.; Wyns, L.; Muyldermans, S.; Conrath, K. Identification of a Universal VHH Framework to Graft Non-Canonical Antigen-Binding Loops of Camel Single-Domain Antibodies. J. Mol. Biol. 2005, 352, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Moutel, S.; Bery, N.; Bernard, V.; Keller, L.; Lemesre, E.; de Marco, A.; Ligat, L.; Rain, J.-C.; Favre, G.; Olichon, A.; et al. NaLi-H1: A Universal Synthetic Library of Humanized Nanobodies Providing Highly Functional Antibodies and Intrabodies. eLife 2016, 5, e16228. [Google Scholar] [CrossRef] [Green Version]
- Uchański, T.; Zögg, T.; Yin, J.; Yuan, D.; Wohlkönig, A.; Fischer, B.; Rosenbaum, D.M.; Kobilka, B.K.; Pardon, E.; Steyaert, J. An Improved Yeast Surface Display Platform for the Screening of Nanobody Immune Libraries. Sci. Rep. 2019, 9, 382. [Google Scholar] [CrossRef] [Green Version]
- Egloff, P.; Zimmermann, I.; Arnold, F.M.; Hutter, C.A.J.; Morger, D.; Opitz, L.; Poveda, L.; Keserue, H.-A.; Panse, C.; Roschitzki, B.; et al. Engineered Peptide Barcodes for In-Depth Analyses of Binding Protein Libraries. Nat. Methods 2019, 16, 421–428. [Google Scholar] [CrossRef]
- Cawez, F.; Duray, E.; Hu, Y.; Vandenameele, J.; Romão, E.; Vincke, C.; Dumoulin, M.; Galleni, M.; Muyldermans, S.; Vandevenne, M. Combinatorial Design of a Nanobody That Specifically Targets Structured RNAs. J. Mol. Biol. 2018, 430, 1652–1670. [Google Scholar] [CrossRef]
- Pardon, E.; Laeremans, T.; Triest, S.; Rasmussen, S.G.F.; Wohlkönig, A.; Ruf, A.; Muyldermans, S.; Hol, W.G.J.; Kobilka, B.K.; Steyaert, J. A General Protocol for the Generation of Nanobodies for Structural Biology. Nat. Protoc. 2014, 9, 674–693. [Google Scholar] [CrossRef]
- André, A.S.; Moutinho, I.; Dias, J.N.R.; Aires-da-Silva, F. In Vivo Phage Display: A Promising Selection Strategy for the Improvement of Antibody Targeting and Drug Delivery Properties. Front. Microbiol. 2022, 13, 962124. [Google Scholar] [CrossRef] [PubMed]
- Lambin, P. Les gènes des immunoglobulines et l’origine de la diversité des anticorps. Rev. Fr. Transfus. Hémobiologie 1992, 35, 47–65. [Google Scholar] [CrossRef]
- Piramoon, M.; Khodadust, F.; Hosseinimehr, S.J. Radiolabeled Nanobodies for Tumor Targeting: From Bioengineering to Imaging and Therapy. Biochim. Biophys. Acta BBA Rev. Cancer 2021, 1875, 188529. [Google Scholar] [CrossRef]
- Hrynchak, I.; Santos, L.; Falcão, A.; Gomes, C.M.; Abrunhosa, A.J. Nanobody-Based Theranostic Agents for HER2-Positive Breast Cancer: Radiolabeling Strategies. Int. J. Mol. Sci. 2021, 22, 10745. [Google Scholar] [CrossRef] [PubMed]
- Pruszynski, M.; Koumarianou, E.; Vaidyanathan, G.; Revets, H.; Devoogdt, N.; Lahoutte, T.; Zalutsky, M.R. Targeting Breast Carcinoma with Radioiodinated Anti-HER2 Nanobody. Nucl. Med. Biol. 2013, 40, 52–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruszynski, M.; Koumarianou, E.; Vaidyanathan, G.; Revets, H.; Devoogdt, N.; Lahoutte, T.; Lyerly, H.K.; Zalutsky, M.R. Improved Tumor Targeting of Anti-HER2 Nanobody Through N -Succinimidyl 4-Guanidinomethyl-3-Iodobenzoate Radiolabeling. J. Nucl. Med. 2014, 55, 650–656. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.M. (Ed.) . The Protein Protocols Handbook; Humana Press: Towota, NJ, USA, 1996; ISBN 978-0-89603-338-2. [Google Scholar]
- Küppers, J.; Kürpig, S.; Bundschuh, R.A.; Essler, M.; Lütje, S. Radiolabeling Strategies of Nanobodies for Imaging Applications. Diagnostics 2021, 11, 1530. [Google Scholar] [CrossRef]
- D’Huyvetter, M.; Aerts, A.; Xavier, C.; Vaneycken, I.; Devoogdt, N.; Gijs, M.; Impens, N.; Baatout, S.; Ponsard, B.; Muyldermans, S.; et al. Development of 177 Lu-Nanobodies for Radioimmunotherapy of HER2-Positive Breast Cancer: Evaluation of Different Bifunctional Chelators: 177 LU-NANOBODIES FOR RADIOIMMUNOTHERAPY. Contrast Media Mol. Imaging 2012, 7, 254–264. [Google Scholar] [CrossRef]
- D’Huyvetter, M.; Xavier, C.; Caveliers, V.; Lahoutte, T.; Muyldermans, S.; Devoogdt, N. Radiolabeled Nanobodies as Theranostic Tools in Targeted Radionuclide Therapy of Cancer. Expert Opin. Drug Deliv. 2014, 11, 1939–1954. [Google Scholar] [CrossRef]
- Massa, S.; Xavier, C.; De Vos, J.; Caveliers, V.; Lahoutte, T.; Muyldermans, S.; Devoogdt, N. Site-Specific Labeling of Cysteine-Tagged Camelid Single-Domain Antibody-Fragments for Use in Molecular Imaging. Bioconjug. Chem. 2014, 25, 979–988. [Google Scholar] [CrossRef]
- Sugiura, G.; Kühn, H.; Sauter, M.; Haberkorn, U.; Mier, W. Radiolabeling Strategies for Tumor-Targeting Proteinaceous Drugs. Molecules 2014, 19, 2135–2165. [Google Scholar] [CrossRef]
- Zhou, Z.; Chitneni, S.K.; Devoogdt, N.; Zalutsky, M.R.; Vaidyanathan, G. Fluorine-18 Labeling of an Anti-HER2 VHH Using a Residualizing Prosthetic Group via a Strain-Promoted Click Reaction: Chemistry and Preliminary Evaluation. Bioorg. Med. Chem. 2018, 26, 1939–1949. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Vaidyanathan, G.; Koumarianou, E.; Kang, C.M.; Zalutsky, M.R. Astatine-211 Labeled Anti-HER2 5F7 Single Domain Antibody Fragment Conjugates: Radiolabeling and Preliminary Evaluation. Nucl. Med. Biol. 2018, 56, 10–20. [Google Scholar] [CrossRef] [Green Version]
- D’Huyvetter, M.; De Vos, J.; Xavier, C.; Pruszynski, M.; Sterckx, Y.G.J.; Massa, S.; Raes, G.; Caveliers, V.; Zalutsky, M.R.; Lahoutte, T.; et al. 131I-Labeled Anti-HER2 Camelid SdAb as a Theranostic Tool in Cancer Treatment. Clin. Cancer Res. 2017, 23, 6616–6628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaidyanathan, G.; McDougald, D.; Choi, J.; Pruszynski, M.; Koumarianou, E.; Zhou, Z.; Zalutsky, M.R. N-Succinimidyl 3-((4-(4-[ 18 F]Fluorobutyl)-1H-1,2,3-Triazol-1-Yl)Methyl)-5-(Guanidinomethyl)Benzoate ([ 18 F]SFBTMGMB): A Residualizing Label for 18 F-Labeling of Internalizing Biomolecules. Org. Biomol. Chem. 2016, 14, 1261–1271. [Google Scholar] [CrossRef] [Green Version]
- Vaidyanathan, G.; McDougald, D.; Choi, J.; Koumarianou, E.; Weitzel, D.; Osada, T.; Lyerly, H.K.; Zalutsky, M.R. Preclinical Evaluation of 18 F-Labeled Anti-HER2 Nanobody Conjugates for Imaging HER2 Receptor Expression by Immuno-PET. J. Nucl. Med. 2016, 57, 967–973. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; McDougald, D.; Devoogdt, N.; Zalutsky, M.R.; Vaidyanathan, G. Labeling Single Domain Antibody Fragments with Fluorine-18 Using 2,3,5,6-Tetrafluorophenyl 6-[ 18 F]Fluoronicotinate Resulting in High Tumor-to-Kidney Ratios. Mol. Pharm. 2019, 16, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Vuillez, J.P. Les Immunoscintigraphies En Oncologie. Rev. Médecine Interne 1995, 16, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Artiko, V. Monoclonal Immunoscintigraphy for Detection of Metastasis and Recurrence of Colorectal Cancer. World J. Gastroenterol. 2011, 17, 2424. [Google Scholar] [CrossRef]
- Yao, Y.-F. Immunoscintigraphy of Local Recurrent Rectal Cancer with 99m Tc-Labeled Anti-CEA Monoclonal Antibody CL58. World J. Gastroenterol. 2007, 13, 1841. [Google Scholar] [CrossRef] [Green Version]
- Vaneycken, I.; D’huyvetter, M.; Hernot, S.; De Vos, J.; Xavier, C.; Devoogdt, N.; Caveliers, V.; Lahoutte, T. Immuno-Imaging Using Nanobodies. Curr. Opin. Biotechnol. 2011, 22, 877–881. [Google Scholar] [CrossRef]
- Shaw, R.J. Glucose Metabolism and Cancer. Curr. Opin. Cell Biol. 2006, 18, 598–608. [Google Scholar] [CrossRef]
- Vaneycken, I.; Devoogdt, N.; Van Gassen, N.; Vincke, C.; Xavier, C.; Wernery, U.; Muyldermans, S.; Lahoutte, T.; Caveliers, V. Preclinical Screening of Anti-HER2 Nanobodies for Molecular Imaging of Breast Cancer. FASEB J. 2011, 25, 2433–2446. [Google Scholar] [CrossRef] [Green Version]
- Lecocq, Q.; De Vlaeminck, Y.; Hanssens, H.; D’Huyvetter, M.; Raes, G.; Goyvaerts, C.; Keyaerts, M.; Devoogdt, N.; Breckpot, K. Theranostics in Immuno-Oncology Using Nanobody Derivatives. Theranostics 2019, 9, 7772–7791. [Google Scholar] [CrossRef]
- Berland, L.; Kim, L.; Abousaway, O.; Mines, A.; Mishra, S.; Clark, L.; Hofman, P.; Rashidian, M. Nanobodies for Medical Imaging: About Ready for Prime Time? Biomolecules 2021, 11, 637. [Google Scholar] [CrossRef] [PubMed]
- Thurber, G.M.; Schmidt, M.M.; Wittrup, K.D. Antibody Tumor Penetration: Transport Opposed by Systemic and Antigen-Mediated Clearance. Adv. Drug Deliv. Rev. 2008, 60, 1421–1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampath, S. Treatment: Radiation Therapy. In Lung Cancer; Reckamp, K.L., Ed.; Cancer Treatment and Research; Springer International Publishing: Cham, Switzerland, 2016; Volume 170, pp. 105–118. ISBN 978-3-319-40387-8. [Google Scholar]
- Bao, G.; Tang, M.; Zhao, J.; Zhu, X. Nanobody: A Promising Toolkit for Molecular Imaging and Disease Therapy. EJNMMI Res. 2021, 11, 6. [Google Scholar] [CrossRef]
- Jadvar, H. Targeted Radionuclide Therapy: An Evolution Toward Precision Cancer Treatment. Am. J. Roentgenol. 2017, 209, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Ramogida, C.F.; Orvig, C. Tumour Targeting with Radiometals for Diagnosis and Therapy. Chem. Commun. 2013, 49, 4720. [Google Scholar] [CrossRef] [PubMed]
- Artigas, C.; Mileva, M.; Flamen, P.; Karfis, I. Targeted Radionuclide Therapy: An Emerging Field in Solid Tumours. Curr. Opin. Oncol. 2021, 33, 493–499. [Google Scholar] [CrossRef]
- ClinicalTrials Database. Available online: https://clinicaltrials.gov/ (accessed on 20 June 2023).
- Randhawa, P.; Olson, A.P.; Chen, S.; Gower-Fry, K.L.; Hoehr, C.; Engle, J.W.; Ramogida, C.F.; Radchenko, V. Meitner-Auger Electron Emitters for Targeted Radionuclide Therapy: Mercury-197m/g and Antimony-119. Curr. Radiopharm. 2021, 14, 394–419. [Google Scholar] [CrossRef]
- Boswell, C.A.; Brechbiel, M.W. Development of Radioimmunotherapeutic and Diagnostic Antibodies: An inside-out View. Nucl. Med. Biol. 2007, 34, 757–778. [Google Scholar] [CrossRef] [Green Version]
- Silindir-Gunay, M.; Karpuz, M.; Ozer, A.Y. Targeted Alpha Therapy and Nanocarrier Approach. Cancer Biother. Radiopharm. 2020, 35, 446–458. [Google Scholar] [CrossRef]
- de Kruijff, R.; Wolterbeek, H.; Denkova, A. A Critical Review of Alpha Radionuclide Therapy—How to Deal with Recoiling Daughters? Pharmaceuticals 2015, 8, 321–336. [Google Scholar] [CrossRef] [Green Version]
- Dash, A.; Knapp, F.F.; Pillai, M. Targeted Radionuclide Therapy—An Overview. Curr. Radiopharm. 2013, 6, 152–180. [Google Scholar] [CrossRef]
- Read, E.D.; Eu, P.; Little, P.J.; Piva, T.J. The Status of Radioimmunotherapy in CD20+ Non-Hodgkin’s Lymphoma. Target. Oncol. 2015, 10, 15–26. [Google Scholar] [CrossRef]
- Trujillo-Nolasco, M.; Morales-Avila, E.; Cruz-Nova, P.; Katti, K.V.; Ocampo-García, B. Nanoradiopharmaceuticals Based on Alpha Emitters: Recent Developments for Medical Applications. Pharmaceutics 2021, 13, 1123. [Google Scholar] [CrossRef] [PubMed]
- Jurcic, J.G.; Rosenblat, T.L. Targeted Alpha-Particle Immunotherapy for Acute Myeloid Leukemia. Am. Soc. Clin. Oncol. Educ. Book 2014, 34, e126–e131. [Google Scholar] [CrossRef] [PubMed]
- Jurcic, J.G.; Rosenblat, T.L.; McDevitt, M.R.; Pandit-Taskar, N.; Carrasquillo, J.A.; Chanel, S.M.; Ryan, C.; Frattini, M.G.; Cicic, D.; Larson, S.M.; et al. Phase I Trial of the Targeted Alpha-Particle Nano-Generator Actinium-225 (225 Ac-Lintuzumab) (Anti-CD33; HuM195) in Acute Myeloid Leukemia (AML). J. Clin. Oncol. 2011, 29, 6516. [Google Scholar] [CrossRef]
- Zalutsky, M.R.; Reardon, D.A.; Akabani, G.; Coleman, R.E.; Friedman, A.H.; Friedman, H.S.; McLendon, R.E.; Wong, T.Z.; Bigner, D.D. Clinical Experience with α-Particle–Emitting 211 At: Treatment of Recurrent Brain Tumor Patients with 211 At-Labeled Chimeric Antitenascin Monoclonal Antibody 81C6. J. Nucl. Med. 2008, 49, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, S.; Larsen, R.H.; Fosså, S.D.; Balteskard, L.; Borch, K.W.; Westlin, J.-E.; Salberg, G.; Bruland, Ø.S. First Clinical Experience with α-Emitting Radium-223 in the Treatment of Skeletal Metastases. Clin. Cancer Res. 2005, 11, 4451–4459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krasniqi, A.; D’Huyvetter, M.; Xavier, C.; Van der Jeught, K.; Muyldermans, S.; Van Der Heyden, J.; Lahoutte, T.; Tavernier, J.; Devoogdt, N. Theranostic Radiolabeled Anti-CD20 SdAb for Targeted Radionuclide Therapy of Non-Hodgkin Lymphoma. Mol. Cancer Ther. 2017, 16, 2828–2839. [Google Scholar] [CrossRef] [Green Version]
- D’Huyvetter, M.; Vincke, C.; Xavier, C.; Aerts, A.; Impens, N.; Baatout, S.; De Raeve, H.; Muyldermans, S.; Caveliers, V.; Devoogdt, N.; et al. Targeted Radionuclide Therapy with A 177 Lu-Labeled Anti-HER2 Nanobody. Theranostics 2014, 4, 708–720. [Google Scholar] [CrossRef] [Green Version]
- Nonnekens, J.; Chatalic, K.L.S.; Molkenboer-Kuenen, J.D.M.; Beerens, C.E.M.T.; Bruchertseifer, F.; Morgenstern, A.; Veldhoven-Zweistra, J.; Schottelius, M.; Wester, H.-J.; van Gent, D.C.; et al. 213 Bi-Labeled Prostate-Specific Membrane Antigen-Targeting Agents Induce DNA Double-Strand Breaks in Prostate Cancer Xenografts. Cancer Biother. Radiopharm. 2017, 32, 67–73. [Google Scholar] [CrossRef]
- Ballangrud, A.M.; Yang, W.H.; Charlton, D.E.; McDevitt, M.R.; Hamacher, K.A.; Panageas, K.S.; Ma, D.; Bander, N.H.; Scheinberg, D.A.; Sgouros, G. Response of LNCaP Spheroids after Treatment with an Alpha-Particle Emitter (213Bi)-Labeled Anti-Prostate-Specific Membrane Antigen Antibody (J591). Cancer Res. 2001, 61, 2008–2014. [Google Scholar] [PubMed]
- Li, Y.; Tian, Z.; Rizvi, S.M.A.; Bander, N.H.; Allen, B.J. In Vitro and Preclinical Targeted Alpha Therapy of Human Prostate Cancer with Bi-213 Labeled J591 Antibody against the Prostate Specific Membrane Antigen. Prostate Cancer Prostatic Dis. 2002, 5, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Agus, D.B.; Bunn, P.A.; Franklin, W.; Garcia, M.; Ozols, R.F. HER-2/Neu as a Therapeutic Target in Non-Small Cell Lung Cancer, Prostate Cancer, and Ovarian Cancer. Semin. Oncol. 2000, 27, 53–63, discussion 92–100. [Google Scholar] [PubMed]
- Dekempeneer, Y.; Bäck, T.; Aneheim, E.; Jensen, H.; Puttemans, J.; Xavier, C.; Keyaerts, M.; Palm, S.; Albertsson, P.; Lahoutte, T.; et al. Labeling of Anti-HER2 Nanobodies with Astatine-211: Optimization and the Effect of Different Coupling Reagents on Their in Vivo Behavior. Mol. Pharm. 2019, 16, 3524–3533. [Google Scholar] [CrossRef]
- Puttemans, J.; Dekempeneer, Y.; Eersels, J.L.; Hanssens, H.; Debie, P.; Keyaerts, M.; Windhorst, A.D.; van der Aa, F.; Lecocq, Q.; Breckpot, K.; et al. Preclinical Targeted α- and Β−-Radionuclide Therapy in HER2-Positive Brain Metastasis Using Camelid Single-Domain Antibodies. Cancers 2020, 12, 1017. [Google Scholar] [CrossRef] [Green Version]
- Dekempeneer, Y.; Caveliers, V.; Ooms, M.; Maertens, D.; Gysemans, M.; Lahoutte, T.; Xavier, C.; Lecocq, Q.; Maes, K.; Covens, P.; et al. Therapeutic Efficacy of 213 Bi-Labeled SdAbs in a Preclinical Model of Ovarian Cancer. Mol. Pharm. 2020, 17, 3553–3566. [Google Scholar] [CrossRef]
- Pruszynski, M.; D’Huyvetter, M.; Bruchertseifer, F.; Morgenstern, A.; Lahoutte, T. Evaluation of an Anti-HER2 Nanobody Labeled with 225 Ac for Targeted α-Particle Therapy of Cancer. Mol. Pharm. 2018, 15, 1457–1466. [Google Scholar] [CrossRef]
- De Veirman, K.; Puttemans, J.; Krasniqi, A.; Ertveldt, T.; Hanssens, H.; Romao, E.; Hose, D.; Goyvaert, C.; Vlummens, P.; Muyldermans, S.; et al. CS1-Specific Single-Domain Antibodies Labeled with Actinium-225 Prolong Survival and Increase CD8+ T Cells and PD-L1 Expression in Multiple Myeloma. OncoImmunology 2021, 10, 2000699. [Google Scholar] [CrossRef]
- Puttemans, J.; Stijlemans, B.; Keyaerts, M.; Vander Meeren, S.; Renmans, W.; Fostier, K.; Debie, P.; Hanssens, H.; Rodak, M.; Pruszynski, M.; et al. The Road to Personalized Myeloma Medicine: Patient-Specific Single-Domain Antibodies for Anti-Idiotypic Radionuclide Therapy. Mol. Cancer Ther. 2022, 21, 159–169. [Google Scholar] [CrossRef]
- Feng, Y.; Meshaw, R.; Zhao, X.-G.; Jannetti, S.; Vaidyanathan, G.; Zalutsky, M.R. Effective Treatment of Human Breast Carcinoma Xenografts with Single-Dose 211 At-Labeled Anti-HER2 Single-Domain Antibody Fragment. J. Nucl. Med. 2023, 64, 124–130. [Google Scholar] [CrossRef]
- Ertveldt, T.; Krasniqi, A.; Ceuppens, H.; Puttemans, J.; Dekempeneer, Y.; Jonghe, K.D.; Mey, W.D.; Lecocq, Q.; Vlaeminck, Y.D.; Awad, R.M.; et al. Targeted α-Therapy Using 225 Ac Radiolabeled Single-Domain Antibodies Induces Antigen-Specific Immune Responses and Instills Immunomodulation Both Systemically and at the Tumor Microenvironment. J. Nucl. Med. 2023, 64, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Gong, J.; Qi, Q.; Liu, C.; Su, H.; Xing, Y.; Zhao, J. 131I-Labeled Anti-HER2 Nanobody for Targeted Radionuclide Therapy of HER2-Positive Breast Cancer. Int. J. Nanomed. 2023, 18, 1915–1925. [Google Scholar] [CrossRef]
- Jiang, C.; Tian, Q.; Xu, X.; Li, P.; He, S.; Chen, J.; Yao, B.; Zhang, J.; Yang, Z.; Song, S. Enhanced Antitumor Immune Responses via a New Agent [131I]-Labeled Dual-Target Immunosuppressant. Eur. J. Nucl. Med. Mol. Imaging 2023, 50, 275–286. [Google Scholar] [CrossRef]
- Zhu, M.; Zhang, J.; Yang, M.; Zhang, H.; Xu, T.; Kan, F.; Zhang, X.; Zhang, S.; Yin, Y.; Yu, F. In Vitro and in Vivo Study on the Treatment of Non-Small Cell Lung Cancer with Radionuclide Labeled PD-L1 Nanobody. J. Cancer Res. Clin. Oncol. 2023, 1–14. [Google Scholar] [CrossRef]
- Lemaire, M.; D’Huyvetter, M.; Lahoutte, T.; Van Valckenborgh, E.; Menu, E.; De Bruyne, E.; Kronenberger, P.; Wernery, U.; Muyldermans, S.; Devoogdt, N.; et al. Imaging and Radioimmunotherapy of Multiple Myeloma with Anti-Idiotypic Nanobodies. Leukemia 2014, 28, 444–447. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhu, Y.; Liu, M.; Jin, D.; Zhang, L.; Cheng, J.; Liu, Y. Conjugation of Oxaliplatin with PEGylated-Nanobody for Enhancing Tumor Targeting and Prolonging Circulation. J. Inorg. Biochem. 2021, 223, 111553. [Google Scholar] [CrossRef] [PubMed]
- Tchouate Gainkam, L.O.; Caveliers, V.; Devoogdt, N.; Vanhove, C.; Xavier, C.; Boerman, O.; Muyldermans, S.; Bossuyt, A.; Lahoutte, T. Localization, Mechanism and Reduction of Renal Retention of Technetium-99m Labeled Epidermal Growth Factor Receptor-Specific Nanobody in Mice. Contrast Media Mol. Imaging 2011, 6, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Wållberg, H.; Orlova, A.; Altai, M.; Hosseinimehr, S.J.; Widström, C.; Malmberg, J.; Ståhl, S.; Tolmachev, V. Molecular Design and Optimization of 99m Tc-Labeled Recombinant Affibody Molecules Improves Their Biodistribution and Imaging Properties. J. Nucl. Med. 2011, 52, 461–469. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; de Kruijff, R.M.; Rol, A.; Thijssen, L.; Mendes, E.; Morgenstern, A.; Bruchertseifer, F.; Stuart, M.C.A.; Wolterbeek, H.T.; Denkova, A.G. Retention Studies of Recoiling Daughter Nuclides of 225Ac in Polymer Vesicles. Appl. Radiat. Isot. 2014, 85, 45–53. [Google Scholar] [CrossRef]
- McDevitt, M.R.; Ma, D.; Lai, L.T.; Simon, J.; Borchardt, P.; Frank, R.K.; Wu, K.; Pellegrini, V.; Curcio, M.J.; Miederer, M.; et al. Tumor Therapy with Targeted Atomic Nanogenerators. Science 2001, 294, 1537–1540. [Google Scholar] [CrossRef] [PubMed]
- Cordier, D.; Forrer, F.; Bruchertseifer, F.; Morgenstern, A.; Apostolidis, C.; Good, S.; Müller-Brand, J.; Mäcke, H.; Reubi, J.C.; Merlo, A. Targeted Alpha-Radionuclide Therapy of Functionally Critically Located Gliomas with 213Bi-DOTA-[Thi8,Met(O2)11]-Substance P: A Pilot Trial. Eur. J. Nucl. Med. Mol. Imaging 2010, 37, 1335–1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krolicki, L.; Bruchertseifer, F.; Kunikowska, J.; Koziara, H.; Królicki, B.; Jakuciński, M.; Pawlak, D.; Apostolidis, C.; Mirzadeh, S.; Rola, R.; et al. Prolonged Survival in Secondary Glioblastoma Following Local Injection of Targeted Alpha Therapy with 213Bi-Substance P Analogue. Eur. J. Nucl. Med. Mol. Imaging 2018, 45, 1636–1644. [Google Scholar] [CrossRef] [Green Version]
- Holzwarth, U.; Ojea Jimenez, I.; Calzolai, L. A Random Walk Approach to Estimate the Confinement of α-Particle Emitters in Nanoparticles for Targeted Radionuclide Therapy. EJNMMI Radiopharm. Chem. 2018, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Majkowska-Pilip, A.; Gawęda, W.; Żelechowska-Matysiak, K.; Wawrowicz, K.; Bilewicz, A. Nanoparticles in Targeted Alpha Therapy. Nanomaterials 2020, 10, 1366. [Google Scholar] [CrossRef]
- Ostuni, E.; Taylor, M.R.G. Commercial and Business Aspects of Alpha Radioligand Therapeutics. Front. Med. 2023, 9, 1070497. [Google Scholar] [CrossRef]
- Li, R.G.; Stenberg, V.Y.; Larsen, R.H. An Experimental Generator for Production of High-Purity 212 Pb for Use in Radiopharmaceuticals. J. Nucl. Med. 2023, 64, 173–176. [Google Scholar] [CrossRef]
- Robertson, A.K.H.; Ramogida, C.F.; Schaffer, P.; Radchenko, V. Development of 225 Ac Radiopharmaceuticals: TRIUMF Perspectives and Experiences. Curr. Radiopharm. 2018, 11, 156–172. [Google Scholar] [CrossRef]
- Hassanzadeh Eskafi, A.; Oghalaei, A.; Mahboudi, F.; Ghaderi, H.; Behdani, M.; Shoari, A.; Kazemi-Lomedasht, F. Investigation of the Therapeutic Potential of Recombinant Bispecific Bivalent Anti-PD-L1/VEGF Nanobody in Inhibition of Angiogenesis. Immunopharmacol. Immunotoxicol. 2023, 45, 197–202. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Advantage | Molecular Reason |
|---|---|
| High stability | Ability to refold after denaturation; resistance to acidic and alkaline conditions |
| Improved antigen access | Small size and presence of large exposed loop to penetrate antigen cleft |
| Specific antigen binding | Greater structural variability |
| Low off-target toxicity | Small size and quick blood clearance |
| Rapid tumor penetration | Small size |
| Low immunogenicity | High degree of homology to human VH |
| Facile production | Only one mature domain required |
| Target | Nanobody | Disease | Primary Purpose | Clinical Trial | Phase |
|---|---|---|---|---|---|
| HER2 | 68GaNOTA-Anti-HER2 VHH1 | Breast Neoplasm | Diagnostic | NCT03331601 | 2 |
| Breast Carcinoma | |||||
| MMR | 68GaNOTA-Anti-MMR-VHH2 | Malignant Solid Tumor | Diagnostic | NCT04168528 | 1/2 |
| Breast Cancer | |||||
| Head and Neck Cancer | |||||
| Melanoma (Skin) | |||||
| MMR | 68GaNOTA-Anti-MMR-VHH2 | Squamous Cell Carcinoma of Head and Neck | Diagnostic | NCT04758650 | 2 |
| Cancer | |||||
| Carotid Stenosis | |||||
| Atherosclerosis of Artery | |||||
| Hodgkin Lymphoma | |||||
| Non-Hodgkin Lymphoma | |||||
| Hemophagocytic Lymphoistiocytosis (HLH) | |||||
| Cardiac Sarcaoidosis | |||||
| HER2 | 99mTc-MIRC208 | Cancer | Diagnostic | NCT04591652 | N/A |
| HER2 | 99mTc-NM-02 | Breast Cancer | Treatment | NCT04674722 | Early Phase I |
| 188Re-NM-02 | |||||
| PD-L1 | 99mTc-NM-01 | Non-Small Cell Lung Cancer | Diagnostic | NCT04992715 | 2 |
| Parent | α-Emitting Daughters | T1/2 | Energy of Emitted Particle (MeV) |
|---|---|---|---|
| 211At | 7.2 h | 6 | |
| 211Po | 516 ms | 7.5 | |
| 225Ac | 9.9 d | 6 | |
| 211Fr | 4.9 min | 6 | |
| 217At | 32.3 ms | 7 | |
| 213Bi | 45.6 min | 6 | |
| 213Po | 3.7 µs | 8 | |
| 227Th | 18.7 d | 6 | |
| 223Ra | 11.4 d | 6 | |
| 219Rn | 4 s | 7 | |
| 215Po | 1.8 ms | 7.5 | |
| 211Bi | 2.2 min | 7 |
| Target | Radionuclide | Labeling Strategy | Model Used | Main Findings | Reference |
|---|---|---|---|---|---|
| HER2 | 131I (β- and γ-emitter) | Prosthetic group SGMIB | In vitro: HER2+ cell lines: BT474/M1, JIMT-1, SKOV-3, and SKOV-3.IP1 | High tumor uptake in both mouse models, and low normal tissue uptake | [76] |
| In vivo: HER2+ tumor xenograft mouse models: (1) BT474/M1 and (2) SKOV-3.IP1 | [131I]SGMIB-2Rs15d alone, or in combination with trastuzumab, significantly extended tumor survival | ||||
| CD20 | 177Lu (β-emitter) | DTPA conjugation | In vitro: Daudi (hCD20pos), Reh (hCD20neg), and murine B16-F10 cell lines; hCD20pos B16 cell line generated for study | 177Lu-DTPA-sdAb 9079 showed much lower absorbed doses in non-target organs compared to 177Lu-DTPA-rituximab | [106] |
| In vivo: C57BL6 and CB17 SCID mice bearing hCD20+ B16 tumors | 177Lu-DTPA single-domain antibody (sdAb) 9079 resulted in significantly higher survival rates compared to control | ||||
| HER2 | 177Lu (β-emitter) | DTPA conjugation | In vitro: SKOV3 and SKOV3-LUC (in-house HER2pos/Luciferasepos) | Unwanted kidney retention of radiolabeled nanobodies was reduced when using untagged nanobodies and co-infusion with Gelofusin | [107] |
| In vivo: Female athymic mice bearing HER2+ (SKOV3) tumors | 177Lu-DTPA-2Rs15d efficiently inhibited tumor growth | ||||
| PSMA | 213Bi (α-emitter) | DOTA conjugation | In vitro: PSMA-expressing LNCaP cells | 213Bi-labeled nanobodies induced DNA double-strand breaks in both in vitro and in vivo models | [108] |
| In vivo: LNCaP xenograft BALB/C mice | |||||
| HER2 | 211At (α-emitter) | Prosthetic group SAGMB | In vitro: HER2+ BT474M1 breast carcinoma cells | Anti-HER2 sdAb 5F7 can be efficiently labeled with 211At with excellent affinity and immunoreactivity | [75] |
| In vivo: SCID mice with subcutaneous BT474M1 xenografts | [211At]SAGMB-5F7 had high and prolonged tumor targeting and rapid normal tissue clearance, with iso-[211At]SAGMB-5F7 demonstrating even more favorable results | ||||
| HER2 | 225Ac (α-emitter) | DOTA conjugation | In vitro: SKOV-3 and MDA-MB-231 (low HER2-expressing) cells | HER2 nanobody 2Rs15d can be effectively labeled with 225Ac with preserved affinity and immunoreactivity | [115] |
| In vivo: SKOV3 tumor-xenografted mice | 225Ac-DOTA-Nb was cytotoxic in vitro in a HER2-dependent manner and quickly accumulated in HER2+ tumors in vivo | ||||
| Renal accumulation of 225Ac-DOTA-Nb was effectively reduced with co-infusion of Gelofusin | |||||
| HER2 | 211At (α-emitter) | Conjugation with three different coupling reagents: SAGMB, SAB, MSB) | In vitro: SKOV3 cells | Nanobody labeled via SAGMB ([211At]SAGMB-2Rs15d) was deemed the preferred agent as the biological properties best matched the physical characteristics of 211At | [112] |
| In vivo: SKOV3 tumor-xenografted female nude BALB/C mice | [211At]SAGMB-2Rs15d showed fast and high accumulation in a HER2+ tumor mouse model together with a low non-target organ uptake | ||||
| HER2 | 225Ac (α-emitter) and 131I (β-emitter) | 225Ac: DOTA-based conjugation | In vitro: HER2+ cell lines SKOV3.IP1 and MDA-MB-231Br | [131I]-2Rs15d and [225Ac]-2Rs15d both showed high and specific tumor uptake in HER2+ brain lesions | [113] |
| 131I: prosthetic group SGMIB | In vivo: female athymic nude mice (Crl:NU(NCr)-Foxn1nu) with SVOV3.IP1or MDA-MB-231Br tumor xenografts | Administration of radiolabeled nanobodies alone and in combination with trastuzumab significantly increased median survival in tumor models (that were unresponsive to trastuzumab alone) | |||
| HER2 | 213Bi (α-emitter) | DTPA conjugation | In vitro: SKOV-3 (HER2+) and CHO (HER2−) cell lines | [213Bi]-DTPA-2Rs15d demonstrated a high tumor uptake, but low uptake in normal tissue (co-infusion of gelofusine also led to 2-fold reduction in kidney uptake) | [114] |
| In vivo: athymic nude mice (Crl/NU(NCr)-Foxn1nu) with SKOV3 tumor xenografts | [213Bi]-DTPA-2Rs15d alone and in combination with trastuzumab significantly increased median survival in in vivo model | ||||
| CS1 | 225Ac (α-emitter) | DOTA conjugation | In vitro: 5T3MMvt and 5TGM1 GFP+ cells | Administration of anti-CS1 sdAbs radiolabeled with 225Ac resulted in significantly increased survival of mice, an increase in CD8+ T-cells, and more PD-L1 expression on immune and non-immune cells | [116] |
| In vivo: C57BL6 mice injected with 5T3MM or 5TGM1 cells | |||||
| 5T33 idiotype | 177Lu (β-emitter) and 225Ac (α emitter) | DTPA conjugation for 177Lu, DOTA conjugation with 225Ac | In vitro: 5T3MM cells | Radiolabeled anti-idiotype sdAbs significantly delayed tumor progression in mice with low 5T33 myeloma lesion load | [117] |
| In vivo: C57BL/KalwRij mice intravenously injected with 5T3MM cells and C57BL/6 mice | Membrane expression of paraprotein was confirmed in five out of seven patients with newly diagnosed myeloma, and two anti-idiotype sdAbs were successfully generated from serum-isolated paraprotein | ||||
| HER2 | 211At (α-emitter) | Prosthetic group SAGMB | In vitro: BT474 cells | Clonogenic survival of BT474 cells exposed to iso-211At-SAGMB-5F7 was reduced | [118] |
| In vivo: NOD-scid-IL2Rgammanull and athymic mice with subcutaneous BT474 xenografts | Dose-dependent tumor growth inhibition was observed with 211At-labeled anti-HER2-specific nanobodies 5F7 and VHH_1028; prolongation in median survival was over 400% for both nanobodies. | ||||
| CD20 | 225Ac (α-emitter) | DOTA conjugation | In vivo: C57BL/6 mice subcutaneously transplanted with B16 melanoma cells expressing human CD20 | 225Ac-DOTA-9079 (nanobody targeting huCD20) resulted in delayed tumor growth and increased blood levels of various cytokines | [119] |
| 225Ac-DOTA-9079 also promoted an environment for antitumoral immune cells and increased the percentage of programmed death-ligand 1 (PD-L1)-positive immune cells in the tumor microenvironment | |||||
| HER2 | 131I (β- and γ-emitter) | Iodogen method | In vivo: BALB/c mice subcutaneously injected with MDA-MB-231 (HER2−) or SKBR3 cells (HER2+) | 131I-NM-02 was effectively taken up by HER2+ tumors with rapid blood clearance and favourable biodistribution | [120] |
| 131I-NM-02 significantly inhibited tumor growth and extended survival | |||||
| PD-L1 and cytotoxic T-lymphocyte-associated protein 4 (CTLA-4) | 131I (β- and γ-emitter) | Iodogen method | In vitro: B16F10 and MCF-7 cells | 131I-KN046 demonstrated high affinity and specificity for PD-L1/CTLA-4 immune targets and strong intratumoral retention capability | [121] |
| In vivo: female BALB/c mice injected with B16F10 or MCF-7 cells | 131I-KN046 enhanced the immune response, leading to upregulated expression of MHC-1 and Fas surface molecules, increases in T-cell activation, and a greater number of tumor-infiltrating immunocytes | ||||
| PD-L1 | 131I (β- and γ-emitter) | Chloramine-T method | In vitro: H460 (PD-L1+) and A549 (PD-L1−) cell lines | H460 cells demonstrated high 131I-Nb109 uptake | [122] |
| In vivo: female BALB/c nude mice inoculated with H460 cells | 131I-Nb109 showed accumulation in H460 tumors, successfully inhibited tumor growth without toxic side effects, and induced H460 cells to release DAMPs |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hurley, K.; Cao, M.; Huang, H.; Wang, Y. Targeted Alpha Therapy (TAT) with Single-Domain Antibodies (Nanobodies). Cancers 2023, 15, 3493. https://doi.org/10.3390/cancers15133493
Hurley K, Cao M, Huang H, Wang Y. Targeted Alpha Therapy (TAT) with Single-Domain Antibodies (Nanobodies). Cancers. 2023; 15(13):3493. https://doi.org/10.3390/cancers15133493
Chicago/Turabian StyleHurley, Kate, Meiyun Cao, Haiming Huang, and Yi Wang. 2023. "Targeted Alpha Therapy (TAT) with Single-Domain Antibodies (Nanobodies)" Cancers 15, no. 13: 3493. https://doi.org/10.3390/cancers15133493
APA StyleHurley, K., Cao, M., Huang, H., & Wang, Y. (2023). Targeted Alpha Therapy (TAT) with Single-Domain Antibodies (Nanobodies). Cancers, 15(13), 3493. https://doi.org/10.3390/cancers15133493