

Exosome-Mediated Activation of the Prostasin-Matriptase Serine Protease Cascade in B Lymphoma Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Establishment of an HEK293T Subline Over-Expressing Prostasin for Exosome Production
2.3. Exosome Isolation and Cell Treatment
2.4. Reverse Transcription and Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
2.5. SDS-Polyacrylamide Gel Electrophoresis (PAGE), Western Blot Analysis, Gelatin-Gel Zymography
2.6. Protease Activity Assay
2.7. Flow Cytometry
2.8. Migration and Invasion
2.9. Statistical Analysis
3. Results
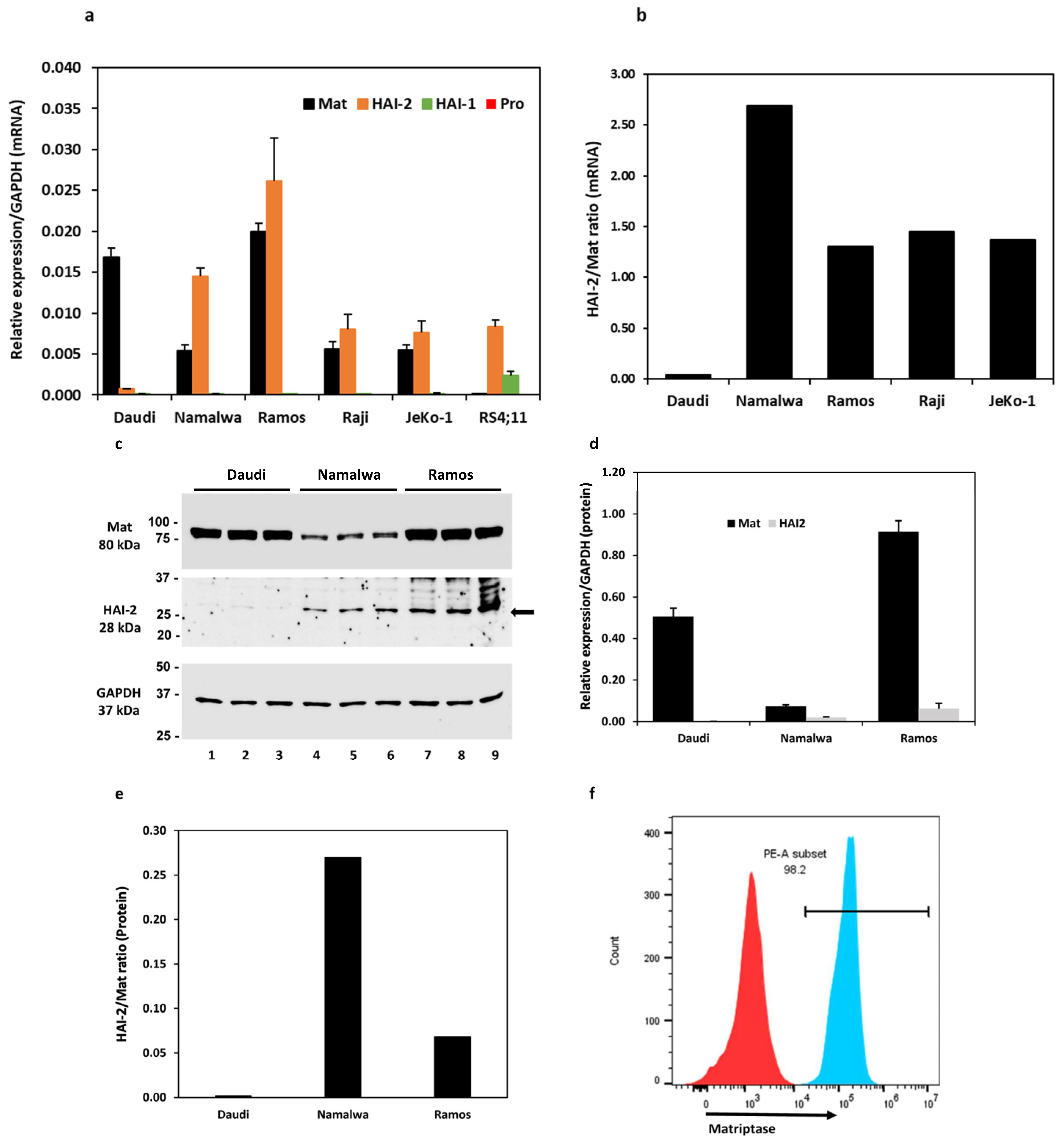
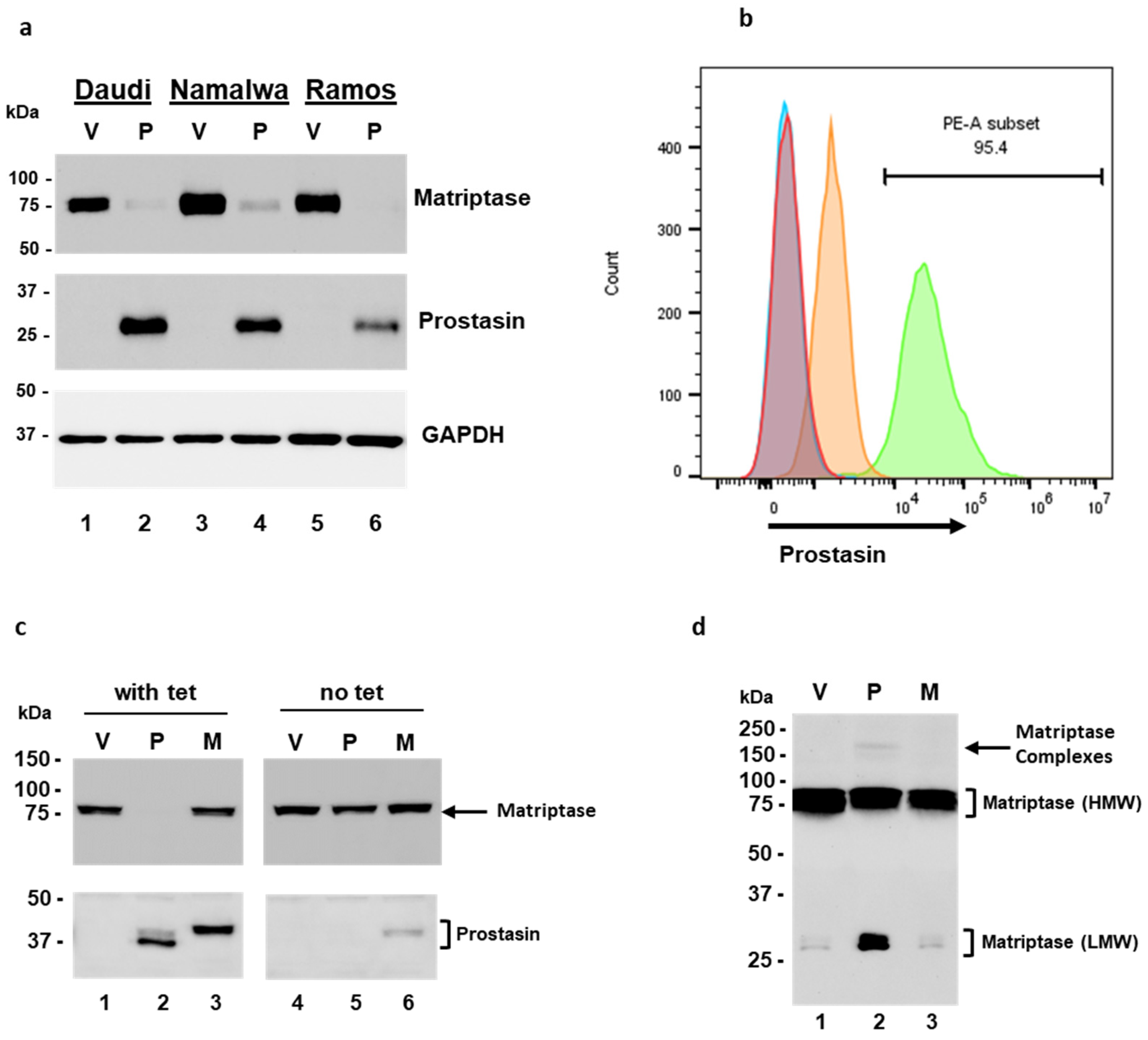
3.1. Expression of Matriptase and Hepatocyte Growth Factor Activator Inhibitors (HAIs) in B Lymphoma Cell Lines
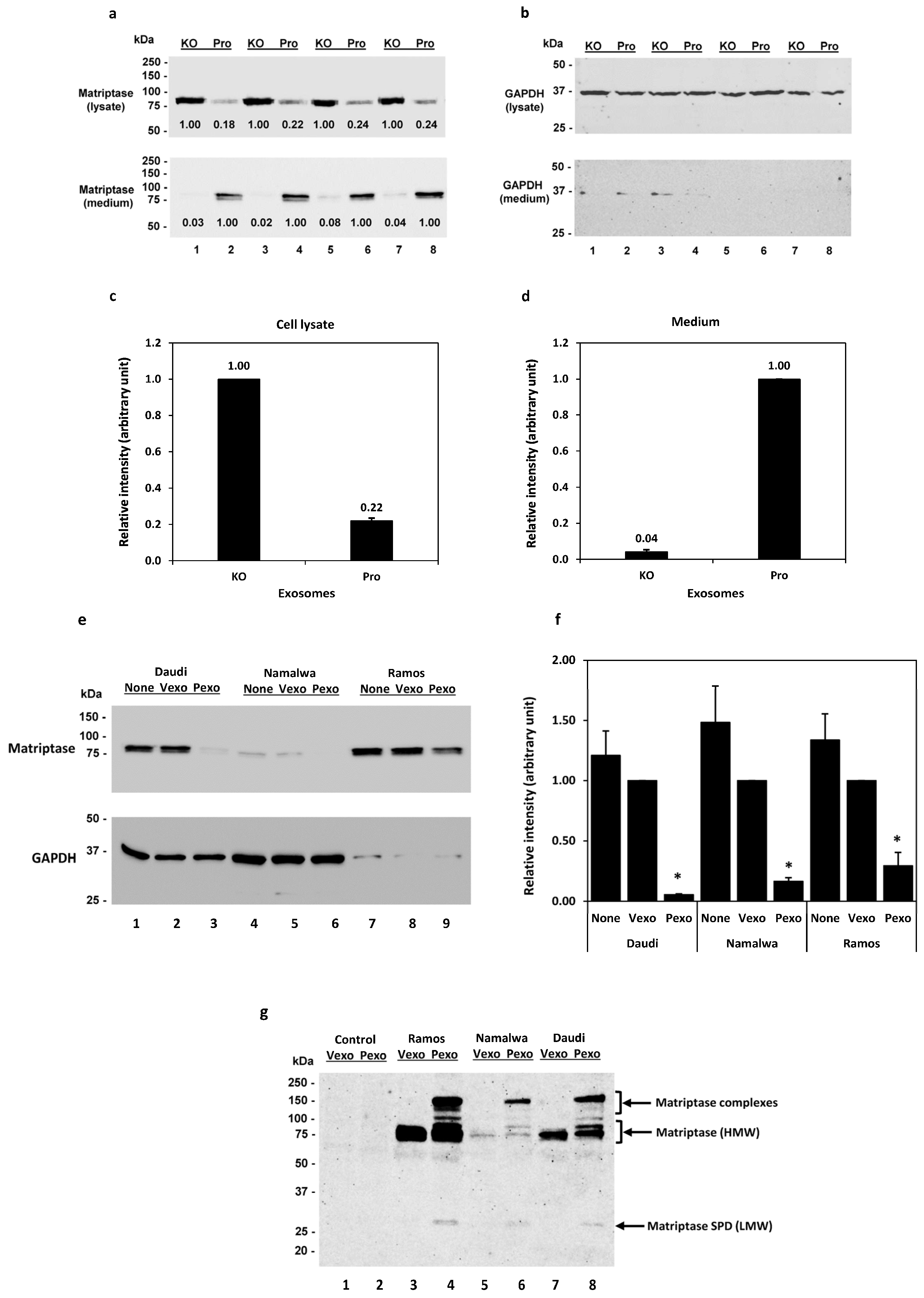
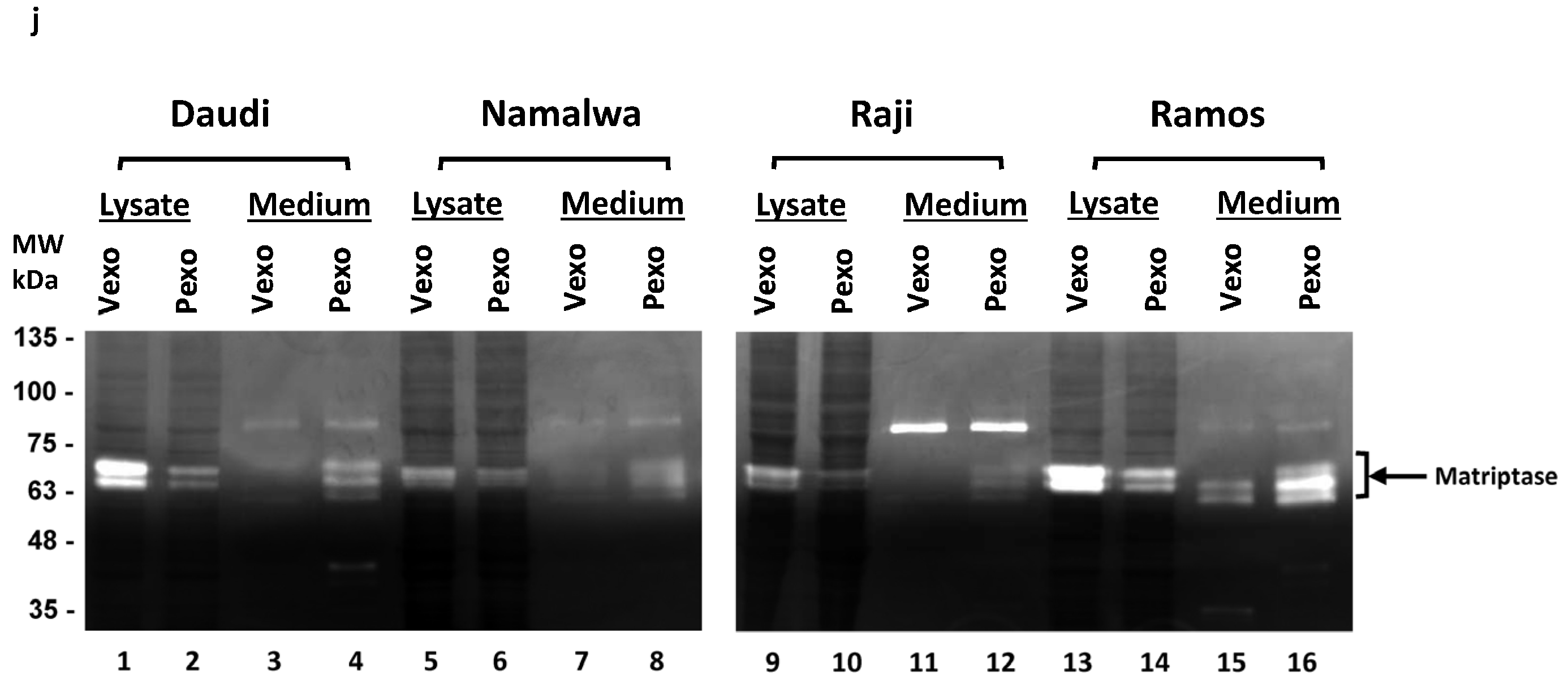
3.2. Prostasin Exosomes Reduce the Matriptase Protein Level in B Lymphoma Cells
3.3. The GPI Anchor and the Serine Active Site of Prostasin Play Roles in the Reduction of Matriptase Quantity in B Lymphoma Cells
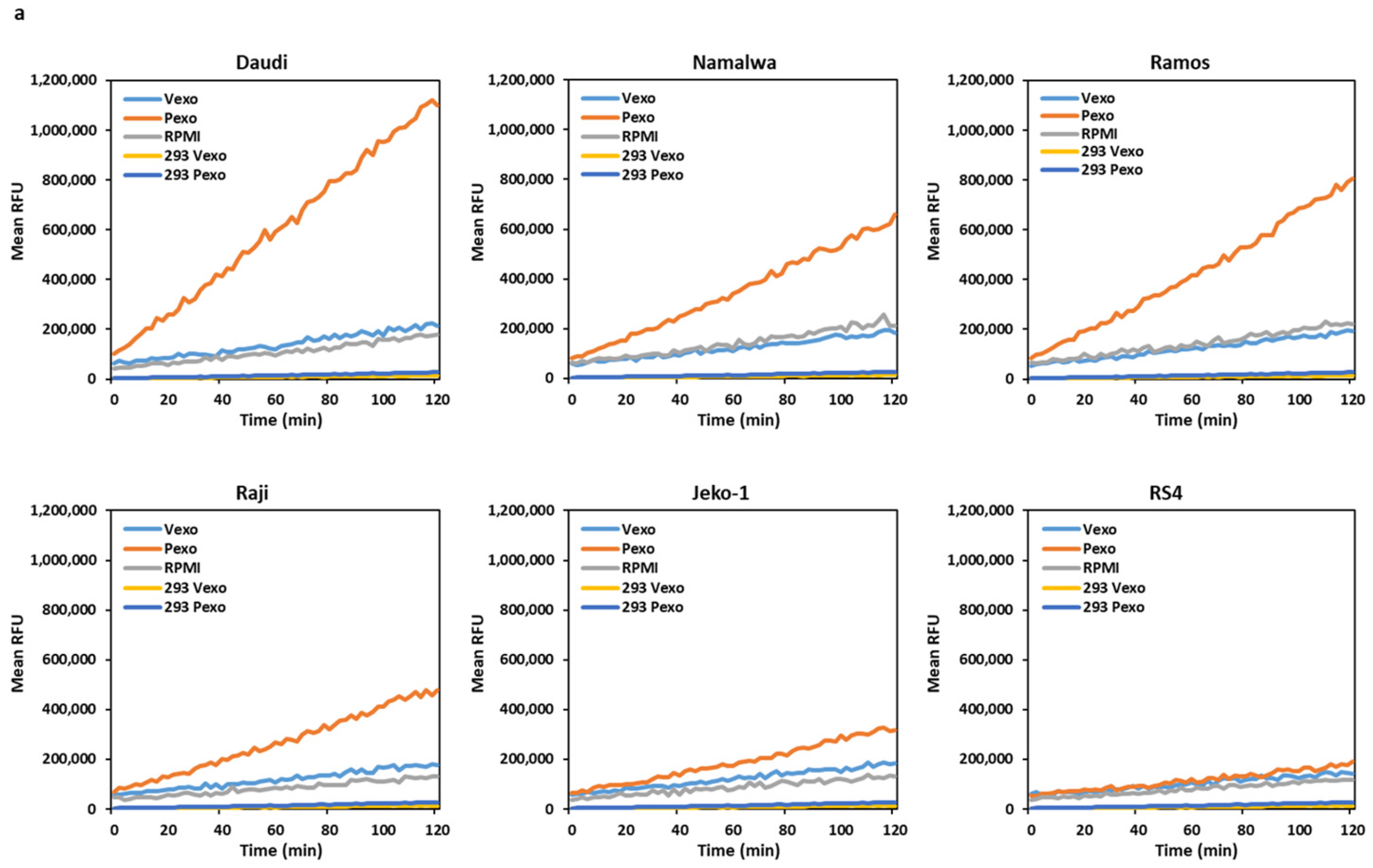
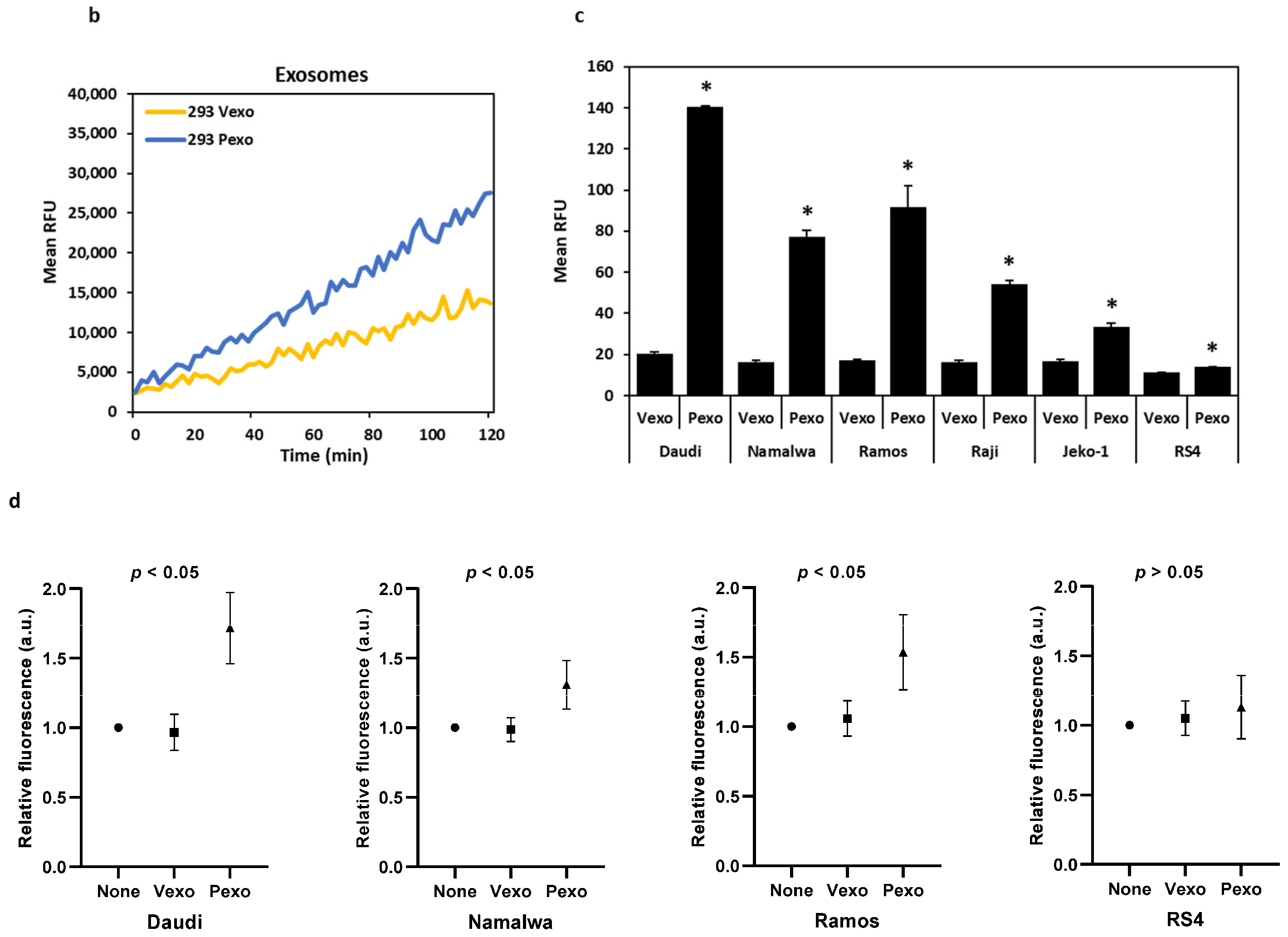
3.4. Trypsin-like Serine Protease Activity Is Increased in the Conditioned Media of Cells Treated with Prostasin Exosomes
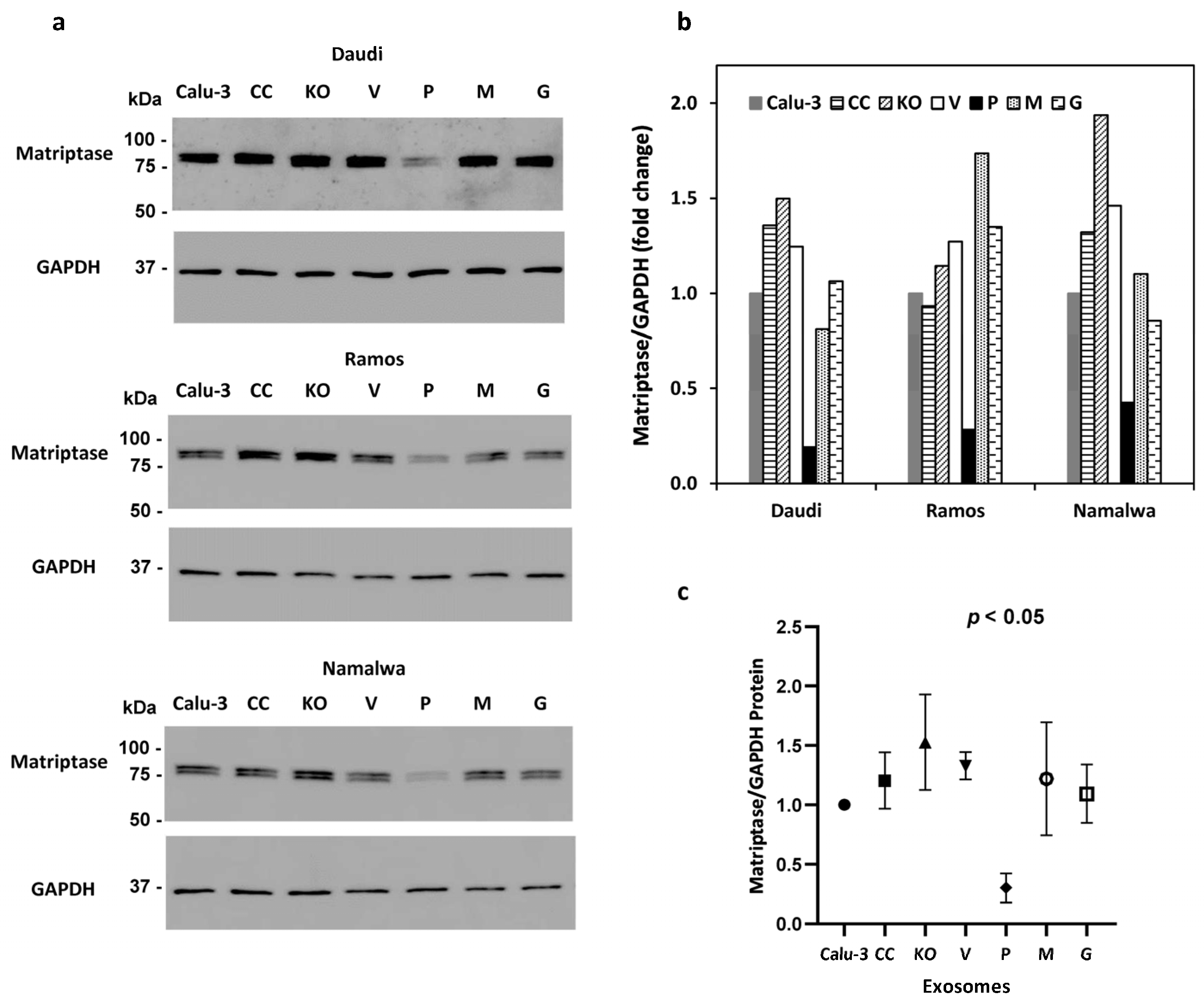
3.5. Over-Expression of Prostasin Reduces Matriptase Quantity in B Cells
3.6. Proliferation, Migration, Invasion, and Gelatinase Activity Changes Associated with Prostasin
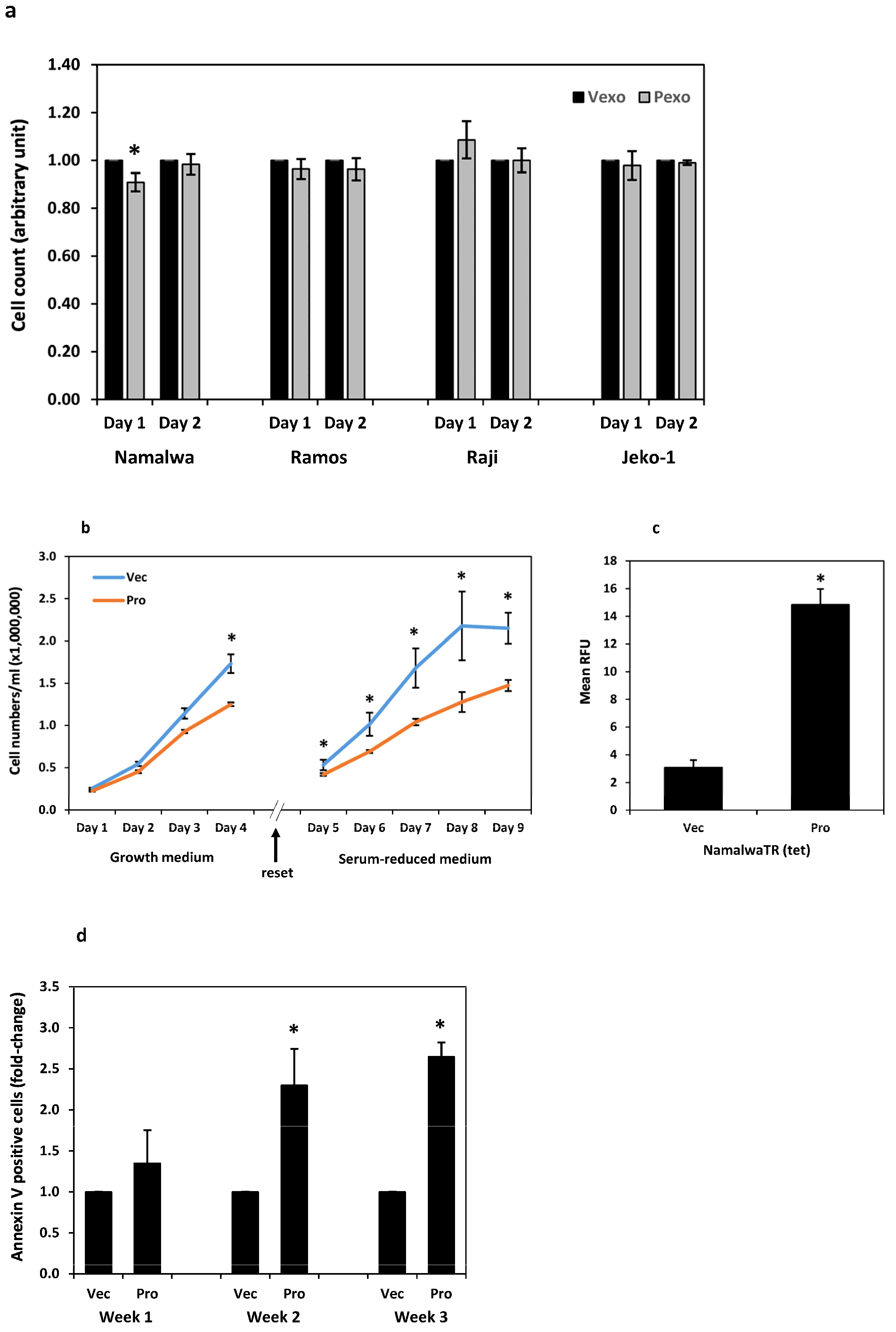
3.6.1. Proliferation and Apoptosis
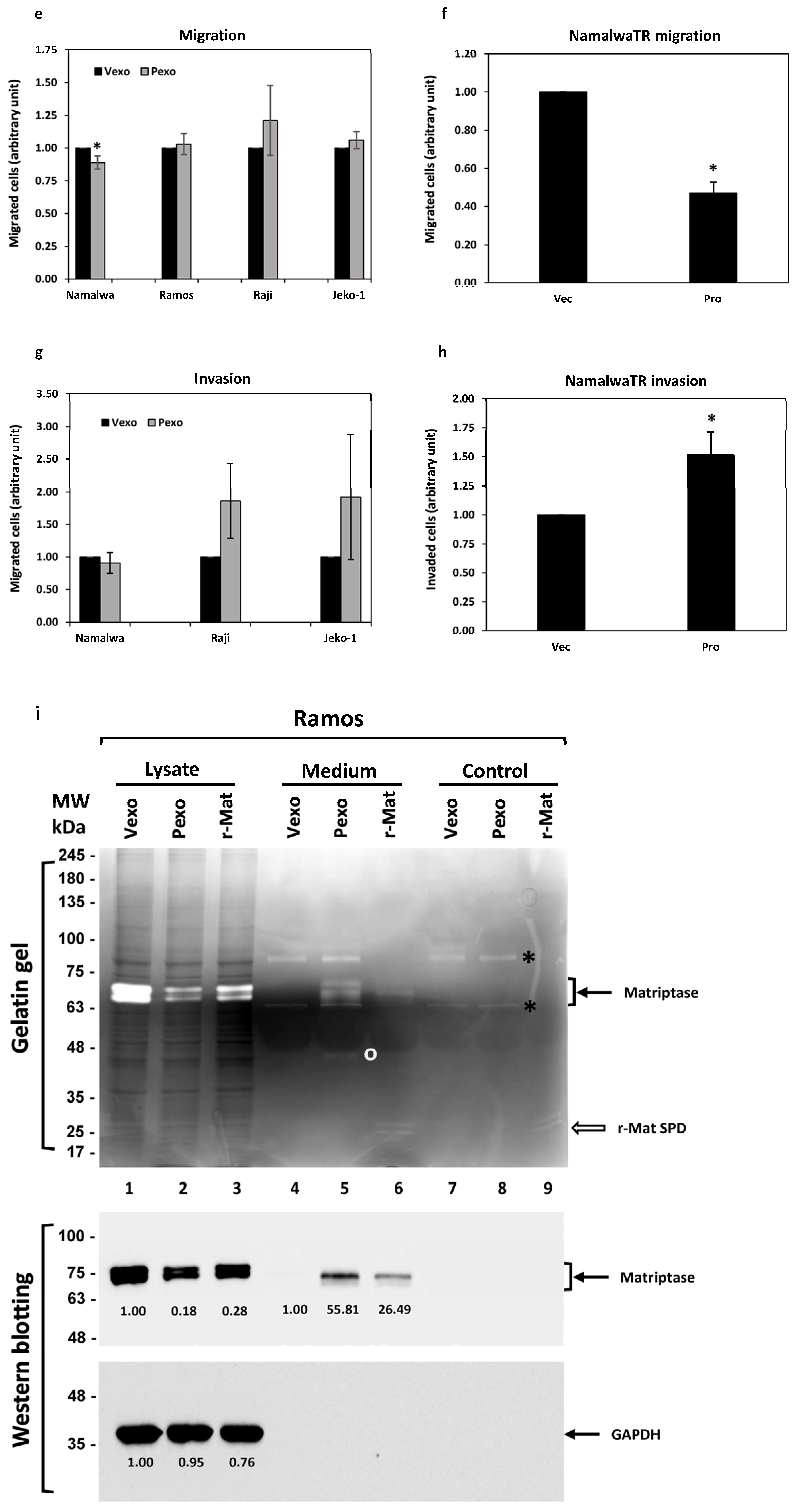
3.6.2. Migration and Invasion
3.6.3. Gelatinase Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barrett, A.J.; Rawlings, N.D.; Salvesen, G.; Woessner, J.F. Introduction. In Handbook of Proteolytic Enzymes, 3rd ed.; Rawlings, N.D., Salvesen, G., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: London, UK, 2013; Volume 1, pp. li–liv. [Google Scholar] [CrossRef]
- Leytus, S.P.; Loeb, K.R.; Hagen, F.S.; Kurachi, K.; Davie, E.W. A novel trypsin-like serine protease (hepsin) with a putative transmembrane domain expressed by human liver and hepatoma cells. Biochemistry 1988, 27, 1067–1074. [Google Scholar] [CrossRef]
- Yu, J.X.; Chao, L.; Chao, J. Prostasin is a novel human serine proteinase from seminal fluid. Purification, tissue distribution, and localization in prostate gland. J. Biol. Chem. 1994, 269, 18843–18848. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.X.; Chao, L.; Chao, J. Molecular Cloning, Tissue-specific Expression, and Cellular Localization of Human Prostasin mRNA. J. Biol. Chem. 1995, 270, 13483–13489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.-Y.; Anders, J.; Johnson, M.; Sang, Q.A.; Dickson, R.B. Molecular Cloning of cDNA for Matriptase, a Matrix-degrading Serine Protease with Trypsin-like Activity. J. Biol. Chem. 1999, 274, 18231–18236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, T.; Shuman, M.A.; Craik, C.S. Reverse biochemistry: Use of macromolecular protease inhibitors to dissect complex biological processes and identify a membrane-type serine protease in epithelial cancer and normal tissue. Proc. Natl. Acad. Sci. USA 1999, 96, 11054–11061. [Google Scholar] [CrossRef]
- Chen, L.M.; Skinner, M.L.; Kauffman, S.W.; Chao, J.; Chao, L.; Thaler, C.D.; Chai, K.X. Prostasin is a glycosylphosphati-dylinositol-anchored active serine protease. J. Biol. Chem. 2001, 276, 21434–21442. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.M.; Chai, K.X. PRSS8 (protease, serine, 8). Atlas Genet. Cytogenet. Oncol. Haematol. 2012, 16, 658–664. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.; Guo, Y.; Yang, Y.; Wei, X.; Zhang, S.; Zhang, Y.; Li, K.; Yuan, M.; Guo, D.; Macias, V.; et al. PRSS8 suppresses colorectal carcinogenesis and metastasis. Oncogene 2019, 38, 497–517, Erratum in Oncogene 2021, 40, 1922–1924. [Google Scholar] [CrossRef]
- List, K.; Szabo, R.; Molinolo, A.; Sriuranpong, V.; Redeye, V.; Murdock, T.; Burke, B.; Nielsen, B.S.; Gutkind, J.S.; Bugge, T.H. Deregulated matriptase causes ras-independent multistage carcinogenesis and promotes ras-mediated malignant transformation. Genes Dev. 2005, 19, 1934–1950. [Google Scholar] [CrossRef] [Green Version]
- Netzel-Arnett, S.; Currie, B.M.; Szabo, R.; Lin, C.-Y.; Chen, L.-M.; Chai, K.X.; Antalis, T.M.; Bugge, T.H.; List, K. Evidence for a Matriptase-Prostasin Proteolytic Cascade Regulating Terminal Epidermal Differentiation. J. Biol. Chem. 2006, 281, 32941–32945. [Google Scholar] [CrossRef] [Green Version]
- List, K.; Hobson, J.P.; Molinolo, A.; Bugge, T.H. Co-localization of the channel activating protease prostasin/(CAP1/PRSS8) with its candidate activator, matriptase. J. Cell Physiol. 2007, 213, 237–245. [Google Scholar] [CrossRef]
- Chen, M.; Chen, L.-M.; Lin, C.-Y.; Chai, K.X. The epidermal growth factor receptor (EGFR) is proteolytically modified by the Matriptase–Prostasin serine protease cascade in cultured epithelial cells. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2008, 1783, 896–903. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-W.; Wang, J.-K.; Chou, F.-P.; Chen, C.-Y.; Rorke, E.A.; Chen, L.-M.; Chai, K.X.; Eckert, R.L.; Johnson, M.D.; Lin, C.-Y. Regulation of the Matriptase-Prostasin Cell Surface Proteolytic Cascade by Hepatocyte Growth Factor Activator Inhibitor-1 during Epidermal Differentiation. J. Biol. Chem. 2010, 285, 31755–31762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Chen, Y.-W.; Battu, A.; Wilder, P.; Weber, D.; Yu, W.; MacKerell, A.D.; Chen, L.-M.; Chai, K.X.; Johnson, M.D.; et al. Targeting Zymogen Activation to Control the Matriptase-Prostasin Proteolytic Cascade. J. Med. Chem. 2011, 54, 7567–7578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, G.S.; List, K. The matriptase-prostasin proteolytic cascade in epithelial development and pathology. Cell Tissue Res. 2012, 351, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Friis, S.; Sales, K.U.; Godiksen, S.; Peters, D.E.; Lin, C.-Y.; Vogel, L.K.; Bugge, T.H. A Matriptase-Prostasin Reciprocal Zymogen Activation Complex with Unique Features: Prostasin as a non-enzymatic co-factor for matriptase activation. J. Biol. Chem. 2013, 288, 19028–19039. [Google Scholar] [CrossRef] [Green Version]
- Oberst, M.D.; Williams, C.A.; Dickson, R.B.; Johnson, M.D.; Lin, C.-Y. The Activation of Matriptase Requires Its Noncatalytic Domains, Serine Protease Domain, and Its Cognate Inhibitor. J. Biol. Chem. 2003, 278, 26773–26779. [Google Scholar] [CrossRef] [Green Version]
- Friis, S.; Sales, K.U.; Schafer, J.M.; Vogel, L.K.; Kataoka, H.; Bugge, T.H. The Protease Inhibitor HAI-2, but Not HAI-1, Regulates Matriptase Activation and Shedding through Prostasin. J. Biol. Chem. 2014, 289, 22319–22332. [Google Scholar] [CrossRef] [Green Version]
- Tseng, C.-C.; Jia, B.; Barndt, R.; Gu, Y.; Chen, C.-Y.; Tseng, I.-C.; Su, S.-F.; Wang, J.-K.; Johnson, M.D.; Lin, C.-Y. Matriptase shedding is closely coupled with matriptase zymogen activation and requires de novo proteolytic cleavage likely involving its own activity. PLoS ONE 2017, 12, e0183507. [Google Scholar] [CrossRef] [Green Version]
- Shimomura, T.; Denda, K.; Kitamura, A.; Kawaguchi, T.; Kito, M.; Kondo, J.; Kagaya, S.; Qin, L.; Takata, H.; Miyazawa, K.; et al. Hepatocyte Growth Factor Activator Inhibitor, a Novel Kunitz-type Serine Protease Inhibitor. J. Biol. Chem. 1997, 272, 6370–6376. [Google Scholar] [CrossRef] [Green Version]
- Marlor, C.W.; Delaria, K.A.; Davis, G.; Muller, D.K.; Greve, J.M.; Tamburini, P.P. Identification and Cloning of Human Placental Bikunin, a Novel Serine Protease Inhibitor Containing Two Kunitz Domains. J. Biol. Chem. 1997, 272, 12202–12208. [Google Scholar] [CrossRef] [Green Version]
- Oberst, M.D.; Chen, L.-Y.L.; Kiyomiya, K.-I.; Williams, C.A.; Lee, M.-S.; Johnson, M.D.; Dickson, R.B.; Lin, C.-Y. HAI-1 regulates activation and expression of matriptase, a membrane-bound serine protease. Am. J. Physiol. Physiol. 2005, 289, C462–C470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabo, R.; Sales, K.U.; Kosa, P.; Shylo, N.A.; Godiksen, S.; Hansen, K.K.; Friis, S.; Gutkind, J.S.; Vogel, L.K.; Hummler, E.; et al. Reduced Prostasin (CAP1/PRSS8) Activity Eliminates HAI-1 and HAI-2 Deficiency–Associated Developmental Defects by Preventing Matriptase Activation. PLOS Genet. 2012, 8, e1002937. [Google Scholar] [CrossRef] [PubMed]
- Nimishakavi, S.; Besprozvannaya, M.; Raymond, W.W.; Craik, C.S.; Gruenert, D.C.; Caughey, G.H.; Menou, A.; Duitman, J.; Flajolet, P.; Sallenave, J.-M.; et al. Activity and inhibition of prostasin and matriptase on apical and basolateral surfaces of human airway epithelial cells. Am. J. Physiol. Cell Mol. Physiol. 2012, 303, L97–L106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.-H.D.; Xu, Y.; Lai, H.; Yang, X.; Tseng, C.-C.; Lai, Y.-J.J.; Pan, Y.; Zhou, E.; Johnson, M.D.; Wang, J.-K.; et al. Differential Subcellular Localization Renders HAI-2 a Matriptase Inhibitor in Breast Cancer Cells but Not in Mammary Epithelial Cells. PLoS ONE 2015, 10, e0120489. [Google Scholar] [CrossRef]
- Larsen, B.R.; Steffensen, S.D.; Nielsen, N.V.; Friis, S.; Godiksen, S.; Bornholdt, J.; Soendergaard, C.; Nonboe, A.W.; Andersen, M.N.; Poulsen, S.S.; et al. Hepatocyte growth factor activator inhibitor-2 prevents shedding of matriptase. Exp. Cell Res. 2013, 319, 918–929. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-P.; Kao, C.-Y.; Chang, S.-C.; Chiu, Y.-L.; Chen, Y.-J.; Chen, M.-H.G.; Chang, C.-C.; Lin, Y.-W.; Chiang, C.-P.; Wang, J.-K.; et al. Tissue distribution and subcellular localizations determine in vivo functional relationship among prostasin, matriptase, HAI-1, and HAI-2 in human skin. PLoS ONE 2018, 13, e0192632. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Liu, M.; Dong, N.; Jiang, Y.; Lin, C.-Y.; Huang, M.; Wu, D.; Wu, Q. Matriptase is highly upregulated in chronic lymphocytic leukemia and promotes cancer cell invasion. Leukemia 2013, 27, 1191–1194. [Google Scholar] [CrossRef]
- Chou, F.-P.; Chen, Y.-W.; Zhao, X.F.; Xu-Monette, Z.Y.; Young, K.H.; Gartenhaus, R.B.; Wang, J.-K.; Kataoka, H.; Zuo, A.H.; Barndt, R.J.; et al. Imbalanced Matriptase Pericellular Proteolysis Contributes to the Pathogenesis of Malignant B-Cell Lymphomas. Am. J. Pathol. 2013, 183, 1306–1317. [Google Scholar] [CrossRef] [Green Version]
- Chiu, Y.-L.; Wu, Y.-Y.; Barndt, R.B.; Yeo, Y.H.; Lin, Y.-W.; Sytwo, H.-P.; Liu, H.-C.; Xu, Y.; Jia, B.; Wang, J.-K.; et al. Aberrant regulation favours matriptase proteolysis in neoplastic B-cells that co-express HAI-2. J. Enzym. Inhib. Med. Chem. 2019, 34, 692–702. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.; Smith, E.R.; Hanada, K.; Stevens, V.L.; Mayor, S. GPI anchoring leads to sphingolipid-dependent retention of endocytosed proteins in the recycling endosomal compartment. EMBO J. 2001, 20, 1583–1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, L.; Wang, M. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Lubbe, N.; Jansen, P.M.; Salih, M.; Fenton, R.A.; van den Meiracker, A.H.; Danser, A.H.; Zietse, R.; Hoorn, E.J. The Phosphorylated Sodium Chloride Cotransporter in Urinary Exosomes Is Superior to Prostasin as a Marker for Aldosteronism. Hypertension 2012, 60, 741–748. [Google Scholar] [CrossRef] [Green Version]
- Olivieri, O.; Chiecchi, L.; Pizzolo, F.; Castagna, A.; Raffaelli, R.; Gunasekaran, M.; Guarini, P.; Consoli, L.; Salvagno, G.; Kitamura, K. Urinary prostasin in normotensive individuals: Correlation with the aldosterone to renin ratio and urinary sodium. Hypertens. Res. 2013, 36, 528–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Y.; Wang, X.; Rose, K.L.; MacDonald, W.H.; Zhang, B.; Schey, K.L.; Luther, J.M. Activation of the Endogenous Renin-Angiotensin-Aldosterone System or Aldosterone Administration Increases Urinary Exosomal Sodium Channel Excretion. J. Am. Soc. Nephrol. 2016, 27, 646–656. [Google Scholar] [CrossRef] [Green Version]
- Zachar, R.; Jensen, B.L.; Svenningsen, P.; Takeda, R.; Stickford, A.S.; Best, S.A.; Yoo, J.-K.; Hissen, S.L.; Liu, Y.-L.; Fu, Q. Dietary Na+intake in healthy humans changes the urine extracellular vesicle prostasin abundance while the vesicle excretion rate, NCC, and ENaC are not altered. Am. J. Physiol. Physiol. 2019, 317, F1612–F1622. [Google Scholar] [CrossRef]
- Krishnamachary, B.; Cook, C.; Spikes, L.; Chalise, P.; Dhillon, N.K. The Potential Role of Extracellular Vesicles in COVID-19 Asso-ciated Endothelial injury and Pro-inflammation. medRxiv 2020. [Google Scholar] [CrossRef]
- Fontana, S.; Mauceri, R.; Novara, M.E.; Alessandro, R.; Campisi, G. Protein Cargo of Salivary Small Extracellular Vesicles as Potential Functional Signature of Oral Squamous Cell Carcinoma. Int. J. Mol. Sci. 2021, 22, 11160. [Google Scholar] [CrossRef]
- Chen, L.M. Prostasin in Human Health and Disease; World Scientific Publishing Co. Pte. Ltd.: Singapore, 2023; ISBN 978-981-126-814-4. [Google Scholar]
- Chen, L.-M.; Chai, J.C.; Liu, B.; Strutt, T.M.; McKinstry, K.K.; Chai, K.X. Prostasin regulates PD-L1 expression in human lung cancer cells. Biosci. Rep. 2021, 41, BSR20211370. [Google Scholar] [CrossRef]
- Chai, A.C.; Robinson, A.L.; Chai, K.X.; Chen, L.-M. Ibuprofen regulates the expression and function of membrane-associated serine proteases prostasin and matriptase. BMC Cancer 2015, 15, 1025. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.-Y.; Gao, W.-L.; Chen, M.; Chai, K.X.; Wang, Y.-L.; Chen, L.-M. Prostasin regulates human placental trophoblast cell proliferation via the epidermal growth factor receptor signaling pathway. Hum. Reprod. 2010, 25, 623–632. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.E.; Torri, J.; Yieh, L.; Wellstein, A.; Lippman, M.E.; Dickson, R.B. Identification and characterization of a novel ma-trix-degrading protease from hormone-dependent human breast cancer cells. Cancer Res. 1993, 53, 1409–1415. [Google Scholar] [PubMed]
- Trentin, L.; Cabrelle, A.; Facco, M.; Carollo, D.; Miorin, M.; Tosoni, A.; Pizzo, P.; Binotto, G.; Nicolardi, L.; Zambello, R.; et al. Homeostatic chemokines drive migration of malignant B cells in patients with non-Hodgkin lymphomas. Blood 2004, 104, 502–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camerer, E.; Barker, A.; Duong, D.N.; Ganesan, R.; Kataoka, H.; Cornelissen, I.; Darragh, M.R.; Hussain, A.; Zheng, Y.-W.; Srinivasan, Y.; et al. Local Protease Signaling Contributes to Neural Tube Closure in the Mouse Embryo. Dev. Cell 2010, 18, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Buzza, M.S.; Martin, E.W.; Driesbaugh, K.H.; Désilets, A.; Leduc, R.; Antalis, T.M. Prostasin Is Required for Matriptase Activation in Intestinal Epithelial Cells to Regulate Closure of the Paracellular Pathway. J. Biol. Chem. 2013, 288, 10328–10337. [Google Scholar] [CrossRef] [Green Version]
- List, K.; Bugge, T.H.; Szabo, R. Matriptase: Potent Proteolysis on the Cell Surface. Mol. Med. 2006, 12, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Scott, C.A.; Ho, Y.N.; Mahabaleshwar, H.; Marsay, K.S.; Zhang, C.; Teow, C.K.; Ng, S.S.; Zhang, W.; Tergaonkar, V.; et al. Matriptase activation of Gq drives epithelial disruption and inflammation via RSK and DUOX. eLife 2021, 10, e66596. [Google Scholar] [CrossRef]
- Gaymon, D.O.; Barndt, R.; Stires, H.; Riggins, R.B.; Johnson, M.D. ROS is a master regulator of in vitro matriptase activation. PLoS ONE 2023, 18, e0267492. [Google Scholar] [CrossRef]
- Balasubramanian, K.; Schroit, A.J. Aminophospholipid Asymmetry: A Matter of Life and Death. Annu. Rev. Physiol. 2003, 65, 701–734. [Google Scholar] [CrossRef]
- Demchenko, A.P. Beyond annexin V: Fluorescence response of cellular membranes to apoptosis. Cytotechnology 2012, 65, 157–172. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-Y.; Wang, J.-K.; Torri, J.; Dou, L.; Sang, Q.A.; Dickson, R.B. Characterization of a Novel, Membrane-bound, 80-kDa Matrix-degrading Protease from Human Breast Cancer Cells. Monoclonal antibody production, isolation, and localization. J. Biol. Chem. 1997, 272, 9147–9152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blüml, S.; McKeever, K.; Ettinger, R.; Smolen, J.; Herbst, R. B-cell targeted therapeutics in clinical development. Arthritis Res. Ther. 2013, 15 (Suppl. S1), S4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paju, A.; Sorsa, T.; Tervahartiala, T.; Koivunen, E.; Haglund, C.; Leminen, A.; Wahlström, T.; Salo, T.; Stenman, U.-H. The levels of trypsinogen isoenzymes in ovarian tumour cyst fluids are associated with promatrix metalloproteinase-9 but not promatrix metalloproteinase-2 activation. Br. J. Cancer 2001, 84, 1363–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuoka, H.; Sisson, T.H.; Nishiuma, T.; Simon, R.H. Plasminogen-Mediated Activation and Release of Hepatocyte Growth Factor from Extracellular Matrix. Am. J. Respir. Cell Mol. Biol. 2006, 35, 705–713. [Google Scholar] [CrossRef] [Green Version]
- Jedeszko, C.; Victor, B.C.; Podgorski, I.; Sloane, B.F. Fibroblast Hepatocyte Growth Factor Promotes Invasion of Human Mammary Ductal Carcinoma in situ. Cancer Res. 2009, 69, 9148–9155. [Google Scholar] [CrossRef] [Green Version]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.-M.; Chai, K.X. Exosome-Mediated Activation of the Prostasin-Matriptase Serine Protease Cascade in B Lymphoma Cells. Cancers 2023, 15, 3848. https://doi.org/10.3390/cancers15153848
Chen L-M, Chai KX. Exosome-Mediated Activation of the Prostasin-Matriptase Serine Protease Cascade in B Lymphoma Cells. Cancers. 2023; 15(15):3848. https://doi.org/10.3390/cancers15153848
Chicago/Turabian StyleChen, Li-Mei, and Karl X. Chai. 2023. "Exosome-Mediated Activation of the Prostasin-Matriptase Serine Protease Cascade in B Lymphoma Cells" Cancers 15, no. 15: 3848. https://doi.org/10.3390/cancers15153848
APA StyleChen, L. -M., & Chai, K. X. (2023). Exosome-Mediated Activation of the Prostasin-Matriptase Serine Protease Cascade in B Lymphoma Cells. Cancers, 15(15), 3848. https://doi.org/10.3390/cancers15153848