
P38 MAPK and Radiotherapy: Foes or Friends?
 , , , and
, , , and 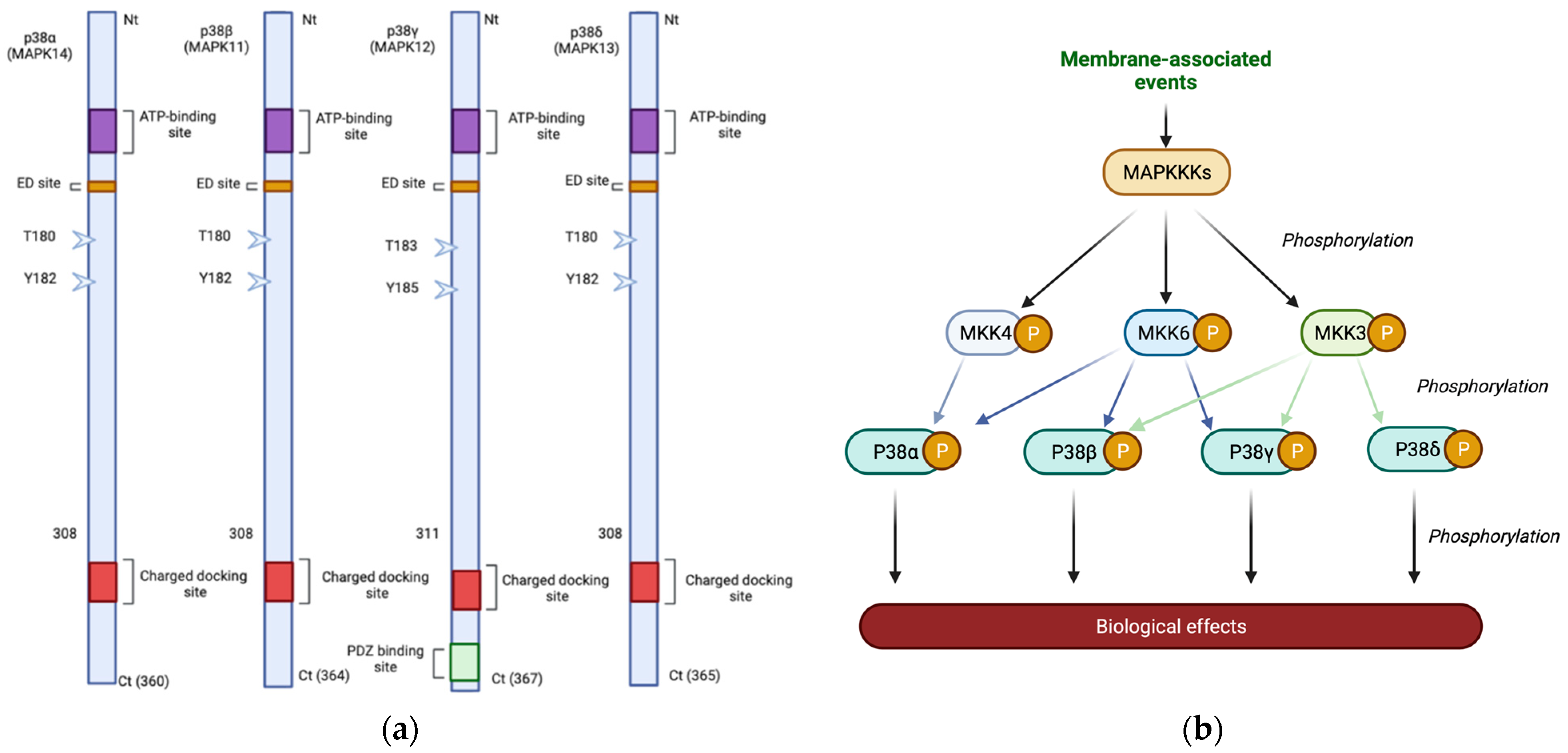{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Knowing the p38 MAPK Protein Family
2. The p38 MAPK Pathway
3. p38 MAPK and Cancer
4. p38 and UV Radiation
5. The P38 MAPK Pathway and Ionizing Radiation
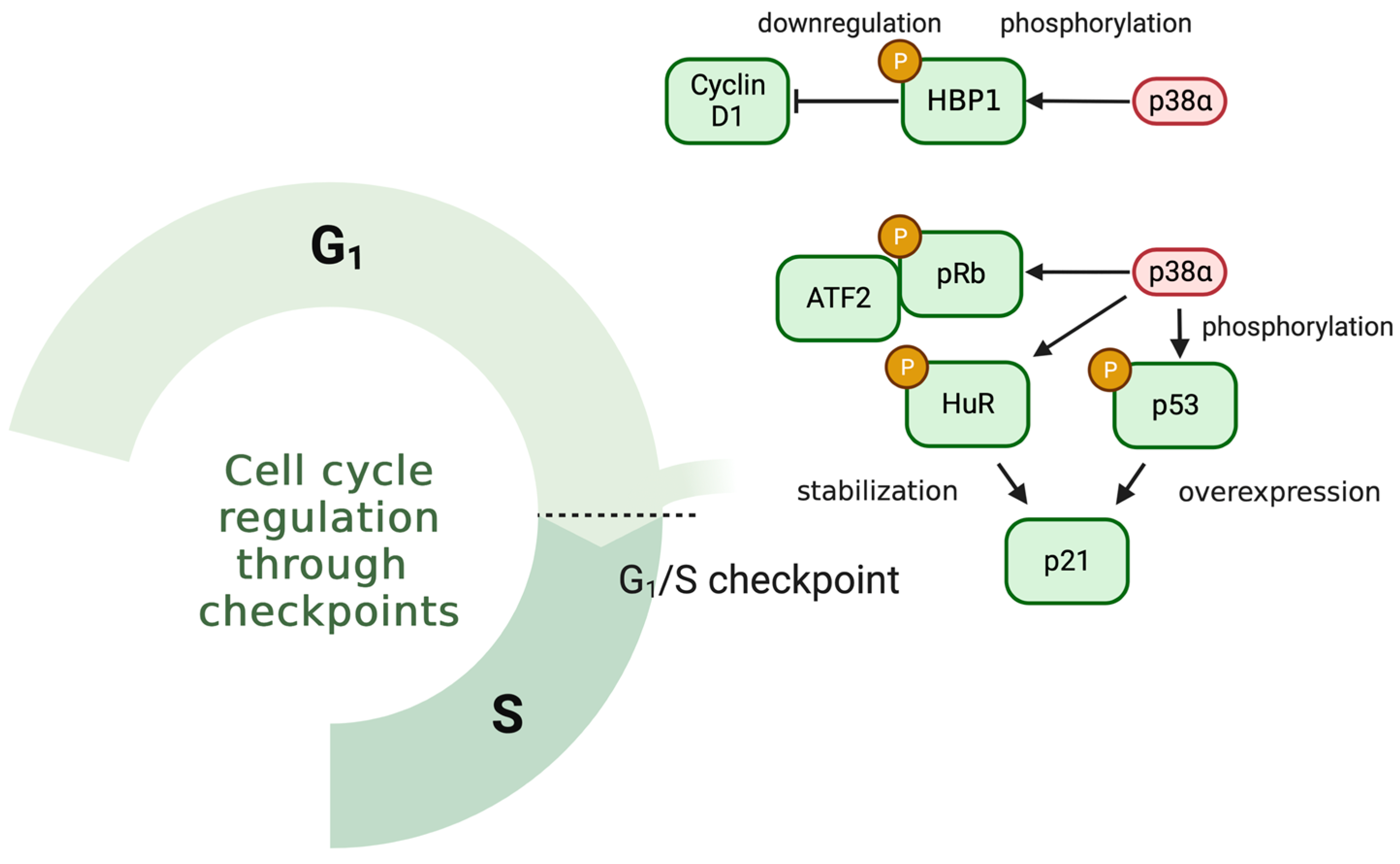
5.1. p38 MAPK and Cell Cycle in Response to IR
5.2. p38 MAPK, ROS and Cell Fate after IR Exposure
5.3. P38 MAPK and Epithelial–Mesenchymal Transition in Response to IR
5.4. Regulation of p38 MAPK Signaling in Response to IR
6. Role of p38 MAPK in the Irradiation of Healthy Tissue
7. p38 MAPK and Radiosensitivity
8. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol. Mol. Biol. Rev. MMBR 2011, 75, 50–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coulombe, P.; Meloche, S. Atypical mitogen-activated protein kinases: Structure, regulation and functions. Biochim. Biophys. Acta 2007, 1773, 1376–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulton, T.G.; Yancopoulos, G.D.; Gregory, J.S.; Slaughter, C.; Moomaw, C.; Hsu, J.; Cobb, M.H. An Insulin-Stimulated Protein Kinase Similar to Yeast Kinases Involved in Cell Cycle Control. Science 1990, 249, 64–67. [Google Scholar] [CrossRef] [PubMed]
- Bogoyevitch, M.A.; Ngoei, K.R.; Zhao, T.T.; Yeap, Y.Y.; Ng, D.C. c-Jun N-terminal kinase (JNK) signaling: Recent advances and challenges. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2010, 1804, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Ezquerro, J.J.; Cuenda, A. p38 Signalling Pathway. Int. J. Mol. Sci. 2021, 22, 1003. [Google Scholar] [CrossRef]
- Paudel, R.; Fusi, L.; Schmidt, M. The MEK5/ERK5 Pathway in Health and Disease. Int. J. Mol. Sci. 2021, 22, 7594. [Google Scholar] [CrossRef]
- Cuenda, A.; Rousseau, S. p38 MAP-kinases pathway regulation, function and role in human diseases. Biochim. Biophys. Acta 2007, 1773, 1358–1375. [Google Scholar] [CrossRef] [Green Version]
- Risco, A.; Cuenda, A. New Insights into the p38γ and p38δ MAPK Pathways. J. Signal Transduct. 2011, 2012, 520289. [Google Scholar] [CrossRef] [Green Version]
- Cuenda, A.; Rouse, J.; Doza, Y.N.; Meier, R.; Cohen, P.; Gallagher, T.F.; Young, P.R.; Lee, J.C. SB 203580 is a specific inhibitor of a MAP kinase homologue which is stimulated by cellular stresses and interleukin-1. FEBS Lett. 1995, 364, 229–233. [Google Scholar]
- Uddin, S.; Ah-Kang, J.; Ulaszek, J.; Mahmud, D.; Wickrema, A. Differentiation stage-specific activation of p38 mitogen-activated protein kinase isoforms in primary human erythroid cells. Proc. Natl. Acad. Sci. USA 2004, 101, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Pogozelski, A.R.; Geng, T.; Li, P.; Yin, X.; Lira, V.A.; Zhang, M.; Chi, J.-T.; Yan, Z. p38gamma mitogen-activated protein kinase is a key regulator in skeletal muscle metabolic adaptation in mice. PLoS ONE 2009, 4, e7934. [Google Scholar] [CrossRef] [PubMed]
- Goedert, M.; Cuenda, A.; Craxton, M.; Jakes, R.; Cohen, P. Activation of the novel stress-activated protein kinase SAPK4 by cytokines and cellular stresses is mediated by SKK3 (MKK6); comparison of its substrate specificity with that of other SAP kinases. EMBO J. 1997, 16, 3563–3571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raingeaud, J.; Gupta, S.; Rogers, J.S.; Dickens, M.; Han, J.; Ulevitch, R.J.; Davis, R.J. Pro-inflammatory Cytokines and Environmental Stress Cause p38 Mitogen-activated Protein Kinase Activation by Dual Phosphorylation on Tyrosine and Threonine. J. Biol. Chem. 1995, 270, 7420–7426. [Google Scholar] [CrossRef] [Green Version]
- Ben-Levy, R.; Hooper, S.; Wilson, R.; Paterson, H.F.; Marshall, C.J. Nuclear export of the stress-activated protein kinase p38 mediated by its substrate MAPKAP kinase-2. Curr. Biol. 1998, 8, 1049–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, C.D.; Thornton, T.M.; Sabio, G.; Davis, R.A.; Rincon, M. Nuclear Localization of p38 MAPK in Response to DNA Damage. Int. J. Biol. Sci. 2009, 5, 428–437. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Lee, J.D.; Bibbs, L.; Ulevitch, R.J. A MAP Kinase Targeted by Endotoxin and Hyperosmolarity in Mammalian Cells. Science 1994, 265, 808–811. [Google Scholar] [CrossRef]
- Rouse, J.; Cohen, P.; Trigon, S.; Morange, M.; Alonso-Llamazares, A.; Zamanillo, D.; Hunt, T.; Nebreda, A.R. A novel kinase cascade triggered by stress and heat shock that stimulates MAPKAP kinase-2 and phosphorylation of the small heat shock proteins. Cell 1994, 78, 1027–1037. [Google Scholar] [CrossRef]
- Canovas, B.; Nebreda, Á.R. Diversity and versatility of p38 kinase signalling in health and disease. Nat. Rev. Mol. Cell Biol. 2021, 22, 346–366. [Google Scholar] [CrossRef]
- del Barco Barrantes, I.; Coya, J.M.; Maina, F.; Arthur, J.S.C.; Nebreda, Á.R. Genetic analysis of specific and redundant roles for p38α and p38β MAPKs during mouse development. Proc. Natl. Acad. Sci. USA 2011, 108, 12764–12769. [Google Scholar] [CrossRef] [Green Version]
- O’Keefe, S.J.; Mudgett, J.S.; Cupo, S.; Parsons, J.N.; Chartrain, N.A.; Fitzgerald, C.; Chen, S.-L.; Lowitz, K.; Rasa, C.; Visco, D.; et al. Chemical Genetics Define the Roles of p38α and p38β in Acute and Chronic Inflammation. J. Biol. Chem. 2007, 282, 34663–34671. [Google Scholar] [CrossRef] [Green Version]
- Tomás-Loba, A.; Manieri, E.; González-Terán, B.; Mora, A.; Leiva-Vega, L.; Santamans, A.M.; Romero-Becerra, R.; Rodríguez, E.; Pintor-Chocano, A.; Feixas, F.; et al. p38γ is essential for cell cycle progression and liver tumorigenesis. Nature 2019, 568, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Alsina-Beauchamp, D.; Escós, A.; Fajardo, P.; González-Romero, D.; Díaz-Mora, E.; Risco, A.; Martín-Serrano, M.A.; Del Fresno, C.; Dominguez-Andrés, J.; Aparicio, N.; et al. Myeloid cell deficiency of p38γ/p38δ protects against candidiasis and regulates antifungal immunity. EMBO Mol. Med. 2018, 10, e8485. [Google Scholar] [CrossRef] [PubMed]
- Cuenda, A.; Sanz-Ezquerro, J.J. p38γ and p38δ: From Spectators to Key Physiological Players. Trends Biochem. Sci. 2017, 42, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Ichijo, H.; Nishida, E.; Irie, K.; ten Dijke, P.; Saitoh, M.; Moriguchi, T.; Takagi, M.; Matsumoto, K.; Miyazono, K.; Gotoh, Y. Induction of apoptosis by ASK1, a mammalian MAPKKK that activates SAPK/JNK and p38 signaling pathways. Science 1997, 275, 90–94. [Google Scholar] [CrossRef]
- Dérijard, B.; Raingeaud, J.; Barrett, T.; Wu, I.-H.; Han, J.; Ulevitch, R.J.; Davis, R.J. Independent Human MAP-Kinase Signal Transduction Pathways Defined by MEK and MKK Isoforms. Science 1995, 267, 682–685. [Google Scholar] [CrossRef]
- Remy, G.; Risco, A.M.; Iñesta-Vaquera, F.A.; González-Terán, B.; Sabio, G.; Davis, R.J.; Cuenda, A. Differential activation of p38MAPK isoforms by MKK6 and MKK3. Cell. Signal. 2010, 22, 660–667. [Google Scholar] [CrossRef]
- Salvador, J.M.; Mittelstadt, P.R.; Guszczynski, T.; Copeland, T.D.; Yamaguchi, H.; Appella, E.; Fornace, A.J.; Ashwell, J.D. Alternative p38 activation pathway mediated by T cell receptor–proximal tyrosine kinases. Nat. Immunol. 2005, 6, 390–395. [Google Scholar] [CrossRef]
- Im, J.-S.; Lee, J.-K. ATR-dependent activation of p38 MAP kinase is responsible for apoptotic cell death in cells depleted of Cdc7. J. Biol. Chem. 2008, 283, 25171–25177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanoue, T.; Adachi, M.; Moriguchi, T.; Nishida, E. A conserved docking motif in MAP kinases common to substrates, activators and regulators. Nat. Cell Biol. 2000, 2, 110–116. [Google Scholar] [CrossRef]
- Tanoue, T.; Maeda, R.; Adachi, M.; Nishida, E. Identification of a docking groove on ERK and p38 MAP kinases that regulates the specificity of docking interactions. EMBO J. 2001, 20, 466–479. [Google Scholar] [CrossRef]
- Han, J.; Wu, J.; Silke, J. An overview of mammalian p38 mitogen-activated protein kinases, central regulators of cell stress and receptor signaling. F1000Research 2020, 9, 653. [Google Scholar] [CrossRef] [PubMed]
- Lang, R.; Raffi, F.A.M. Dual-Specificity Phosphatases in Immunity and Infection: An Update. Int. J. Mol. Sci. 2019, 20, 2710. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Zhang, Y.; Yu, H.; Shen, B.; Liang, Y.; Jin, R.; Liu, X.; Shi, L.; Cai, X. Role of DUSP1/MKP1 in tumorigenesis, tumor progression and therapy. Cancer Med. 2016, 5, 2061–2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahta, R.; Castellino, R.C. Phosphatase magnesium-dependent 1 δ (PPM1D), serine/threonine protein phosphatase and novel pharmacological target in cancer. Biochem. Pharmacol. 2021, 184, 114362. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xia, Y.; Kuang, D.; Duan, Y.; Wang, G. PP2A regulates SCF-induced cardiac stem cell migration through interaction with p38 MAPK. Life Sci. 2017, 191, 59–67. [Google Scholar] [CrossRef]
- Shillingford, S.; Zhang, L.; Surovtseva, Y.; Dorry, S.; Lolis, E.; Bennett, A.M. A novel site on dual-specificity phosphatase MKP7/DUSP16 is required for catalysis and MAPK binding. J. Biol. Chem. 2022, 298, 102617. [Google Scholar] [CrossRef]
- García-Hernández, L.; García-Ortega, M.B.; Ruiz-Alcalá, G.; Carrillo, E.; Marchal, J.A.; García, M.Á. The p38 MAPK Components and Modulators as Biomarkers and Molecular Targets in Cancer. Int. J. Mol. Sci. 2021, 23, 370. [Google Scholar] [CrossRef]
- Ventura, J.J.; Tenbaum, S.; Perdiguero, E.; Huth, M.; Guerra, C.; Barbacid, M.; Pasparakis, M.; Nebreda, Á.R. p38α MAP kinase is essential in lung stem and progenitor cell proliferation and differentiation. Nat. Genet. 2007, 39, 750–758. [Google Scholar] [CrossRef]
- Hui, L.; Bakiri, L.; Mairhorfer, A.; Schweifer, N.; Haslinger, C.; Kenner, L.; Komnenovic, V.; Scheuch, H.; Beug, H.; Wagner, E.F. p38α suppresses normal and cancer cell proliferation by antagonizing the JNK–c-Jun pathway. Nat. Genet. 2007, 39, 741–749. [Google Scholar] [CrossRef]
- Wang, W.; Chen, J.X.; Liao, R.; Deng, Q.; Zhou, J.J.; Huang, S.; Sun, P. Sequential Activation of the MEK-Extracellular Signal-Regulated Kinase and MKK3/6-p38 Mitogen-Activated Protein Kinase Pathways Mediates Oncogenic ras-Induced Premature Senescence. Mol. Cell. Biol. 2002, 22, 3389–3403. [Google Scholar] [CrossRef] [Green Version]
- Gupta, J.; Igea, A.; Papaioannou, M.; Lopez-Casas, P.P.; Llonch, E.; Hidalgo, M.; Gorgoulis, V.G.; Nebreda, Á.R. Pharmacological inhibition of p38 MAPK reduces tumor growth in patient-derived xenografts from colon tumors. Oncotarget 2015, 6, 8539–8551. [Google Scholar] [CrossRef] [PubMed]
- Gupta, J.; del Barco Barrantes, I.; Igea, A.; Sakellariou, S.; Pateras, I.S.; Gorgoulis, V.G.; Nebreda, Á.R. Dual function of p38α MAPK in colon cancer: Suppression of colitis-associated tumor initiation but requirement for cancer cell survival. Cancer Cell 2014, 25, 484–500. [Google Scholar] [CrossRef] [Green Version]
- Urosevic, J.; Garcia-Albéniz, X.; Planet, E.; Real, S.; Céspedes, M.V.; Guiu, M.; Fernandez, E.; Bellmunt, A.; Gawrzak, S.; Pavlovic, M.; et al. Colon cancer cells colonize the lung from established liver metastases through p38 MAPK signalling and PTHLH. Nat. Cell Biol. 2014, 16, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Thoms, H.C.; Dunlop, M.G.; Stark, L.A. p38-mediated inactivation of cyclin D1/cyclin-dependent kinase 4 stimulates nucleolar translocation of RelA and apoptosis in colorectal cancer cells. Cancer Res. 2007, 67, 1660–1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gubern, A.; Joaquin, M.; Marquès, M.; Maseres, P.; Garcia-Garcia, J.; Amat, R.; González-Nuñez, D.; Oliva, B.; Real, F.X.; de Nadal, E.; et al. The N-Terminal Phosphorylation of RB by p38 Bypasses Its Inactivation by CDKs and Prevents Proliferation in Cancer Cells. Mol. Cell 2016, 64, 25–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Prieto, R.; Rojas, J.M.; Taya, Y.; Gutkind, J.S. A role for the p38 mitogen-acitvated protein kinase pathway in the transcriptional activation of p53 on genotoxic stress by chemotherapeutic agents. Cancer Res. 2000, 60, 2464–2472. [Google Scholar]
- Bulavin, D.V.; Higashimoto, Y.; Popoff, I.J.; Gaarde, W.A.; Basrur, V.; Potapova, O.; Appella, E.; Fornace, A.J. Initiation of a G2/M checkpoint after ultraviolet radiation requires p38 kinase. Nature 2001, 411, 102–107. [Google Scholar] [CrossRef]
- Reinhardt, H.C.; Aslanian, A.S.; Lees, J.A.; Yaffe, M.B. p53-Deficient Cells Rely on ATM- and ATR-Mediated Checkpoint Signaling through the p38MAPK/MK2 Pathway for Survival after DNA Damage. CCELL 2007, 11, 175–189. [Google Scholar] [CrossRef] [Green Version]
- Brichkina, A.; Bertero, T.; Loh, H.M.; Nguyen, N.T.M.; Emelyanov, A.; Rigade, S.; Ilie, M.; Hofman, P.; Gaggioli, C.; Bulavin, D.V. p38MAPK builds a hyaluronan cancer niche to drive lung tumorigenesis. Genes Dev. 2016, 30, 2623–2636. [Google Scholar] [CrossRef] [Green Version]
- Wen, S.; Hou, Y.; Fu, L.; Xi, L.; Yang, D.; Zhao, M.; Qin, Y.; Sun, K.; Teng, Y.; Liu, M. Cancer-associated fibroblast (CAF)-derived IL32 promotes breast cancer cell invasion and metastasis via integrin β3–p38 MAPK signalling. Cancer Lett. 2019, 442, 320–332. [Google Scholar] [CrossRef]
- Li, Z.; Liu, F.-Y.; Kirkwood, K.L. The p38/MKP-1 signaling axis in oral cancer: Impact of tumor-associated macrophages. Oral Oncol. 2020, 103, 104591. [Google Scholar] [CrossRef] [PubMed]
- Neamatallah, T. Mitogen-Activated Protein Kinase Pathway: A Critical Regulator in Tumor-associated Macrophage Polarization. J. Microsc. Ultrastruct. 2019, 7, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, A.; Rottoli, E.; Mazza, E.M.C.; Rezzonico-Jost, T.; De Ponte Conti, B.; Proietti, M.; Perotti, M.; Civanelli, E.; Perruzza, L.; Catapano, A.L.; et al. P2X7 Receptor Activity Limits Accumulation of T Cells within Tumors. Cancer Res. 2020, 80, 3906–3919. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Liu, R.; Dai, Y.; Hong, S.; Dong, H.; Wang, H. The Role of p38γ in Cancer: From review to outlook. Int. J. Biol. Sci. 2021, 17, 4036–4046. [Google Scholar] [CrossRef]
- Roche, O.; Fernández-Aroca, D.M.; Arconada-Luque, E.; García-Flores, N.; Mellor, L.F.; Ruiz-Hidalgo, M.J.; Sánchez-Prieto, R. p38β and Cancer: The Beginning of the Road. Int. J. Biol. Sci. 2020, 21, 7524. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.-Z.; Xin, H.; Qi, X.-M.; Chen, G. Isoform-specific and cell/tissue-dependent effects of p38 MAPKs in regulating inflammation and inflammation-associated oncogenesis. Front. Biosci. (Landmark Ed.) 2022, 27, 31. [Google Scholar] [CrossRef] [PubMed]
- Barrio, L.; Román-García, S.; Díaz-Mora, E.; Risco, A.; Jiménez-Saiz, R.; Carrasco, Y.R.; Cuenda, A. B Cell Development and T-Dependent Antibody Response Are Regulated by p38γ and p38δ. Front. Cell Dev. Biol. 2020, 8, 189. [Google Scholar] [CrossRef]
- Wang, F.; Qi, X.-M.; Wertz, R.; Mortensen, M.; Hagen, C.; Evans, J.; Sheinin, Y.; James, M.; Liu, P.; Tsai, S.; et al. p38γ MAPK Is Essential for Aerobic Glycolysis and Pancreatic Tumorigenesis. Cancer Res. 2020, 80, 3251–3264. [Google Scholar] [CrossRef]
- Fajardo, P.; Taskova, M.; Martín-Serrano, M.A.; Hansen, J.; Slott, S.; Jakobsen, A.K.; Wibom, M.-L.; Salegi, B.; Muñoz, A.; Barbachano, A.; et al. p38γ and p38δ as biomarkers in the interplay of colon cancer and inflammatory bowel diseases. Cancer Commun. 2022, 42, 897–901. [Google Scholar] [CrossRef]
- Zheng, S.; Yang, C.; Liu, T.; Liu, Q.; Dai, F.; Sheyhidin, I.; Lu, X. Clinicopathological significance of p38β, p38γ, and p38δ and its biological roles in esophageal squamous cell carcinoma. Tumour Biol. 2016, 37, 7255–7266. [Google Scholar] [CrossRef]
- Katopodis, P.; Kerslake, R.; Zikopoulos, A.; Beri, N.; Anikin, V. p38β-MAPK11 and its role in female cancers. J. Ovarian Res. 2021, 14, 84. [Google Scholar] [CrossRef]
- López-Camarillo, C.; Ocampo, E.A.; Casamichana, M.L.; Pérez-Plasencia, C.; Alvarez-Sánchez, E.; Marchat, L.A. Protein kinases and transcription factors activation in response to UV-radiation of skin: Implications for carcinogenesis. Int. J. Biol. Sci. 2012, 13, 142–172. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Guyton, K.Z.; Gorospe, M.; Xu, Q.; Lee, J.C.; Holbrook, N.J. Differential activation of ERK, JNK/SAPK and P38/CSBP/RK map kinase family members during the cellular response to arsenite. Free. Radic. Biol. Med. 1996, 21, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Manke, I.A.; Nguyen, A.; Lim, D.; Stewart, M.Q.; Elia, A.E.H.; Yaffe, M.B. MAPKAP kinase-2 is a cell cycle checkpoint kinase that regulates the G2/M transition and S phase progression in response to UV irradiation. Mol. Cell 2005, 17, 37–48. [Google Scholar] [CrossRef]
- Ulsamer, A.; Martínez-Limón, A.; Bader, S.; Rodríguez-Acebes, S.; Freire, R.; Méndez, J.; de Nadal, E.; Posas, F. Regulation of Claspin by the p38 stress-activated protein kinase protects cells from DNA damage. Cell Rep. 2022, 40, 111375. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Barakat, B.M.; Qin, S.; Ray, A.; El-Mahdy, M.A.; Wani, G.; Arafa, E.-S.; Mir, S.N.; Wang, Q.-E.; Wani, A.A. The p38 mitogen-activated protein kinase augments nucleotide excision repair by mediating DDB2 degradation and chromatin relaxation. J. Biol. Chem. 2008, 283, 32553–32561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jagirdar, K.; Yin, K.; Harrison, M.; Lim, W.; Muscat, G.E.O.; Sturm, R.A.; Smith, A.G. The NR4A2 nuclear receptor is recruited to novel nuclear foci in response to UV irradiation and participates in nucleotide excision repair. PLoS ONE 2013, 8, e78075. [Google Scholar] [CrossRef] [Green Version]
- Bulavin, D.V.; Saito, S.; Hollander, M.C.; Sakaguchi, K.; Anderson, C.W.; Appella, E.; Fornace, A.J. Phosphorylation of human p53 by p38 kinase coordinates N-terminal phosphorylation and apoptosis in response to UV radiation. EMBO J. 1999, 18, 6845–6854. [Google Scholar] [CrossRef]
- Kim, M.-A.; Kim, H.-J.; Jee, H.J.; Kim, A.J.; Bae, Y.-S.; Bae, S.S.; Yun, J. Akt2, but not Akt1, is required for cell survival by inhibiting activation of JNK and p38 after UV irradiation. Oncogene 2009, 28, 1241–1247. [Google Scholar] [CrossRef] [Green Version]
- Munshi, A.; Ramesh, R. Mitogen-activated protein kinases and their role in radiation response. Genes Cancer 2013, 4, 401–408. [Google Scholar] [CrossRef]
- Broustas, C.G.; Duval, A.J.; Chaudhary, K.R.; Friedman, R.A.; Virk, R.K.; Lieberman, H.B. Targeting MEK5 impairs nonhomologous end-joining repair and sensitizes prostate cancer to DNA damaging agents. Oncogene 2020, 39, 2467–2477. [Google Scholar] [CrossRef]
- Jiang, W.; Jin, G.; Cai, F.; Chen, X.; Cao, N.; Zhang, X.; Liu, J.; Chen, F.; Wang, F.; Dong, W.; et al. Extracellular signal-regulated kinase 5 increases radioresistance of lung cancer cells by enhancing the DNA damage response. Exp. Mol. Med. 2019, 51, 1–20. [Google Scholar] [CrossRef]
- Dent, P.; Yacoub, A.; Fisher, P.B.; Hagan, M.P.; Grant, S. MAPK pathways in radiation responses. Oncogene 2003, 22, 5885–5896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, E.-H.; Lee, S.-J.; Kim, J.-S.; Lee, K.-H.; Um, H.-D.; Kim, J.-H.; Kim, S.-J.; Kim, J.-I.; Hwang, S.-G. Ionizing Radiation Induces Cellular Senescence of Articular Chondrocytes via Negative Regulation of SIRT1 by p38 Kinase. J. Biol. Chem. 2010, 285, 1283–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, P.; Raingeaud, J.; Kaneki, M.; Weichselbaum, R.; Davis, R.J.; Kufe, D.; Kharbanda, S. Activation of p38 mitogen-activated protein kinase by c-Abl-dependent and -independent mechanisms. J. Biol. Chem. 1996, 271, 23775–23779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pejchal, J.; Novotný, J.; Mařák, V.; Osterreicher, J.; Tichy, A.; Vávrová, J.; Sinkorová, Z.; Zárybnická, L.; Novotná, E.; Chládek, J.; et al. Activation of p38 MAPK and expression of TGF-β1 in rat colon enterocytes after whole body γ-irradiation. Int. J. Radiat. Biol. 2012, 88, 348–358. [Google Scholar] [CrossRef]
- Segreto, H.R.C.; Oshima, C.T.F.; Franco, M.F.; Silva, M.R.R.; Egami, M.I.; Teixeira, V.P.C.; Segreto, R.A. Phosphorylation and cytoplasmic localization of MAPK p38 during apoptosis signaling in bone marrow granulocytes of mice irradiated in vivo and the role of amifostine in reducing these effects. Acta Histochem. 2011, 113, 300–307. [Google Scholar] [CrossRef]
- Raman, M.; Earnest, S.; Zhang, K.; Zhao, Y.; Cobb, M.H. TAO kinases mediate activation of p38 in response to DNA damage. EMBO J. 2007, 26, 2005–2014. [Google Scholar] [CrossRef]
- Sharma, P.K.; Dwarakanath, B.S.; Varshney, R. Radiosensitization by 2-deoxy-D-glucose and 6-aminonicotinamide involves activation of redox sensitive ASK1-JNK/p38MAPK signaling in head and neck cancer cells. Free Radic. Biol. Med. 2012, 53, 1500–1513. [Google Scholar] [CrossRef]
- Bulavin, D.V.; Amundson, S.A.; Fornace, A.J. p38 and Chk1 kinases: Different conductors for the G(2)/M checkpoint symphony. Curr. Opin. Genet. Dev. 2002, 12, 92–97. [Google Scholar] [CrossRef]
- He, H.; Lin, K.; Zou, C.; Pan, J.; Fu, W.; Zhou, Y.; Lin, H.; Chen, C.; Su, Y. Knockdown of Annexin A2 Enhances Radiosensitivity by Increasing G2/M-Phase Arrest, Apoptosis and Activating the p38 MAPK-HSP27 Pathway in Nasopharyngeal Carcinoma. Front. Oncol. 2022, 12, 769544. [Google Scholar] [CrossRef] [PubMed]
- Lafarga, V.; Cuadrado, A.; Lopez de Silanes, I.; Bengoechea, R.; Fernandez-Capetillo, O.; Nebreda, Á.R. p38 Mitogen-activated protein kinase- and HuR-dependent stabilization of p21(Cip1) mRNA mediates the G(1)/S checkpoint. Mol. Cell. Biol. 2009, 29, 4341–4351. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; McGowan, C.H.; Zhao, M.; He, L.; Downey, J.S.; Fearns, C.; Wang, Y.; Huang, S.; Han, J. Involvement of the MKK6-p38gamma cascade in gamma-radiation-induced cell cycle arrest. Mol. Cell. Biol. 2000, 20, 4543–4552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trempolec, N.; Muñoz, J.P.; Slobodnyuk, K.; Marin, S.; Cascante, M.; Zorzano, A.; Nebreda, Á.R. Induction of oxidative metabolism by the p38α/MK2 pathway. Sci. Rep. 2017, 7, 11367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tormos, A.M.; Taléns-Visconti, R.; Nebreda, A.R.; Sastre, J. p38 MAPK: A dual role in hepatocyte proliferation through reactive oxygen species. Free Radic. Res. 2013, 47, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Luo, J.; Gao, W.; Wu, J.; Shao, Z.; Wang, Z.; Meng, J.; Ou, Z.; Yang, G. COX-2 promotes breast cancer cell radioresistance via p38/MAPK-mediated cellular anti-apoptosis and invasiveness. Tumour Biol. 2013, 34, 2817–2826. [Google Scholar] [CrossRef]
- Tang, F.; Zhang, R.; Wang, J. Cyclooxygenase-2-Mediated Up-Regulation of Mitochondrial Transcription Factor A Mitigates the Radio-Sensitivity of Cancer Cells. Int. J. Mol. Sci. 2019, 20, 1218. [Google Scholar] [CrossRef] [Green Version]
- Hur, J.-M.; Yun, H.-J.; Yang, S.-H.; Lee, W.-Y.; Joe, M.-H.; Kim, D. Gliotoxin enhances radiotherapy via inhibition of radiation-induced GADD45a, p38, and NFkappaB activation. J. Cell. Biochem. 2008, 104, 2174–2184. [Google Scholar] [CrossRef]
- Choi, C.-H.; Xu, H.; Bark, H.; Lee, T.-B.; Yun, J.; Kang, S.-I.; Oh, Y.-K. Balance of NF-kappaB and p38 MAPK is a determinant of radiosensitivity of the AML-2 and its doxorubicin-resistant cell lines. Leuk. Res. 2007, 31, 1267–1276. [Google Scholar] [CrossRef]
- Viktorsson, K.; Ekedahl, J.; Lindebro, M.C.; Lewensohn, R.; Zhivotovsky, B.; Linder, S.; Shoshan, M.C. Defective stress kinase and Bak activation in response to ionizing radiation but not cisplatin in a non-small cell lung carcinoma cell line. Exp. Cell Res. 2003, 289, 256–264. [Google Scholar] [CrossRef]
- Koturbash, I.; Merrifield, M.; Kovalchuk, O. Fractionated exposure to low doses of ionizing radiation results in accumulation of DNA damage in mouse spleen tissue and activation of apoptosis in a p53/Atm-independent manner. Int. J. Radiat. Biol. 2017, 93, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Miller, A.I.; Polverini, P.J. p38 MAPK mediates gamma-irradiation-induced endothelial cell apoptosis, and vascular endothelial growth factor protects endothelial cells through the phosphoinositide 3-kinase-Akt-Bcl-2 pathway. J. Biol. Chem. 2004, 279, 43352–43360. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-Y.; Kim, M.-J.; Kang, C.-M.; Bae, S.; Cho, C.-K.; Soh, J.-W.; Kim, J.-H.; Kang, S.; Chung, H.Y.; Lee, Y.-S.; et al. Activation of Bak and Bax through c-abl-protein kinase Cdelta-p38 MAPK signaling in response to ionizing radiation in human non-small cell lung cancer cells. J. Biol. Chem. 2006, 281, 7049–7059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, W.; Lu, Y.; Zhou, A.; Chen, Y.; Wang, Z.; Wang, L.; Tian, Y. Mast1 mediates radiation-induced gastric injury via the P38 MAPK pathway. Exp. Cell Res. 2021, 409, 112913. [Google Scholar] [CrossRef]
- Salim, H.; Akbar, N.S.; Zong, D.; Vaculova, A.H.; Lewensohn, R.; Moshfegh, A.; Viktorsson, K.; Zhivotovsky, B. miRNA-214 modulates radiotherapy response of non-small cell lung cancer cells through regulation of p38MAPK, apoptosis and senescence. Br. J. Cancer 2012, 107, 1361–1373. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.-J.; Choi, S.-Y.; Park, I.-C.; Hwang, S.-G.; Kim, C.; Choi, Y.-H.; Kim, H.; Lee, K.-H.; Lee, S.-J. Opposing roles of c-Jun NH2-terminal kinase and p38 mitogen-activated protein kinase in the cellular response to ionizing radiation in human cervical cancer cells. Mol. Cancer Res. 2008, 6, 1718–1731. [Google Scholar] [CrossRef] [Green Version]
- Ståhl, S.; Fung, E.; Adams, C.; Lengqvist, J.; Mörk, B.; Stenerlöw, B.; Lewensohn, R.; Lehtiö, J.; Zubarev, R.; Viktorsson, K. Proteomics and pathway analysis identifies JNK signaling as critical for high linear energy transfer radiation-induced apoptosis in non-small lung cancer cells. Mol. Cell. Proteom. MCP 2009, 8, 1117–1129. [Google Scholar] [CrossRef] [Green Version]
- Whitaker, R.H.; Cook, J.G. Stress Relief Techniques: p38 MAPK Determines the Balance of Cell Cycle and Apoptosis Pathways. Biomolecules 2021, 11, 1444. [Google Scholar] [CrossRef]
- Al-Jumayli, M.; Brown, S.L.; Chetty, I.J.; Extermann, M.; Movsas, B. The Biological Process of Aging and the Impact of Ionizing Radiation. Semin. Radiat. Oncol. 2022, 32, 172–178. [Google Scholar] [CrossRef]
- Han, J.; Sun, P. The pathways to tumor suppression via route p38. Trends Biochem. Sci. 2007, 32, 364–371. [Google Scholar] [CrossRef]
- Wang, Y.; Scheiber, M.N.; Neumann, C.; Calin, G.A.; Zhou, D. MicroRNA regulation of ionizing radiation-induced premature senescence. Int. J. Radiat. Oncol. Biol. Phys. 2011, 81, 839–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Aroca, D.M.; García-Flores, N.; Frost, S.; Jiménez-Suarez, J.; Rodríguez-González, A.; Fernández-Aroca, P.; Sabater, S.; Andrés, I.; Garnés-García, C.; Belandia, B.; et al. Genetic perturbation of MAPK11 (p38β) promotes radiosensitivity by enhancing IR-associated senescence. bioRxiv 2022. [Google Scholar] [CrossRef]
- Roy, A.; Bera, S.; Saso, L.; Dwarakanath, B.S. Role of autophagy in tumor response to radiation: Implications for improving radiotherapy. Front. Oncol. 2022, 12, 957373. [Google Scholar] [CrossRef] [PubMed]
- de la Cruz-Morcillo, M.A.; Valero, M.L.L.; Callejas-Valera, J.L.; Arias-González, L.; Melgar-Rojas, P.; Galán-Moya, E.M.; García-Gil, E.; García-Cano, J.; Sánchez-Prieto, R. P38MAPK is a major determinant of the balance between apoptosis and autophagy triggered by 5-fluorouracil: Implication in resistance. Oncogene 2011, 31, 1073–1085. [Google Scholar] [CrossRef]
- Králová, V.; Benešová, S.; Cervinka, M.; Rudolf, E. Selenite-induced apoptosis and autophagy in colon cancer cells. Toxicol. Vitr. Int. J. Publ. Assoc. BIBRA 2012, 26, 258–268. [Google Scholar] [CrossRef]
- Zhu, Q.; Guo, Y.; Chen, S.; Fu, D.; Li, Y.; Li, Z.; Ni, C. Irinotecan Induces Autophagy-Dependent Apoptosis and Positively Regulates ROS-Related JNK- and P38-MAPK Pathways in Gastric Cancer Cells. OTT 2020, 13, 2807–2817. [Google Scholar] [CrossRef] [Green Version]
- Qiao, L.; Chen, Y.; Liang, N.; Xie, J.; Deng, G.; Chen, F.; Wang, X.; Liu, F.; Li, Y.; Zhang, J. Targeting Epithelial-to-Mesenchymal Transition in Radioresistance: Crosslinked Mechanisms and Strategies. Front. Oncol. 2022, 12, 775238. [Google Scholar] [CrossRef]
- Jung, J.-W.; Hwang, S.-Y.; Hwang, J.-S.; Oh, E.-S.; Park, S.; Han, I.-O. Ionising radiation induces changes associated with epithelial-mesenchymal transdifferentiation and increased cell motility of A549 lung epithelial cells. Eur. J. Cancer 2007, 43, 1214–1224. [Google Scholar] [CrossRef]
- Pan, Y.; Zhou, C.; Yuan, D.; Zhang, J.; Shao, C. Radiation Exposure Promotes Hepatocarcinoma Cell Invasion through Epithelial Mesenchymal Transition Mediated by H2S/CSE Pathway. Radiat. Res. 2016, 185, 96–105. [Google Scholar] [CrossRef]
- Berggren, K.L.; Restrepo Cruz, S.; Hixon, M.D.; Cowan, A.T.; Keysar, S.B.; Craig, S.; James, J.; Barry, M.; Ozbun, M.A.; Jimeno, A.; et al. MAPKAPK2 (MK2) inhibition mediates radiation-induced inflammatory cytokine production and tumor growth in head and neck squamous cell carcinoma. Oncogene 2019, 38, 7329–7341. [Google Scholar] [CrossRef]
- Chowdhury, P.; Dey, P.; Ghosh, S.; Sarma, A.; Ghosh, U. Reduction of metastatic potential by inhibiting EGFR/Akt/p38/ERK signaling pathway and epithelial-mesenchymal transition after carbon ion exposure is potentiated by PARP-1 inhibition in non-small-cell lung cancer. BMC Cancer 2019, 19, 829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, P.; Dey, P.; De, D.; Ghosh, U. Gamma ray-induced in vitro cell migration via EGFR/ERK/Akt/p38 activation is prevented by olaparib pretreatment. Int. J. Radiat. Biol. 2020, 96, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.K.; Khan, S.A.; Sharda, A.; Reddy, D.V.; Gupta, S. MKP1 phosphatase mediates G1-specific dephosphorylation of H3Serine10P in response to DNA damage. Mutat. Res. 2015, 778, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Candas, D.; Lu, C.-L.; Fan, M.; Chuang, F.Y.S.; Sweeney, C.; Borowsky, A.D.; Li, J.J. Mitochondrial MKP1 is a target for therapy-resistant HER2-positive breast cancer cells. Cancer Res. 2014, 74, 7498–7509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, R.M.; Sloane, V.M.; Wu, H.; Luo, L.; Kumar, A.; Kumar, M.V.; Gewirtz, A.T.; Neish, A.S. Flagellin administration protects gut mucosal tissue from irradiation-induced apoptosis via MKP-7 activity. Gut 2011, 60, 648–657. [Google Scholar] [CrossRef]
- Palanichamy, K.; Kanji, S.; Gordon, N.; Thirumoorthy, K.; Jacob, J.R.; Litzenberg, K.T.; Patel, D.; Chakravarti, A. NNMT Silencing Activates Tumor Suppressor PP2A, Inactivates Oncogenic STKs, and Inhibits Tumor Forming Ability. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2017, 23, 2325–2334. [Google Scholar] [CrossRef] [Green Version]
- Hong, C.S.; Ho, W.; Zhang, C.; Yang, C.; Elder, J.B.; Zhuang, Z. LB100, a small molecule inhibitor of PP2A with potent chemo- and radio-sensitizing potential. Cancer Biol. Ther. 2015, 16, 821–833. [Google Scholar] [CrossRef] [Green Version]
- Taher, M.M.; Hershey, C.M.; Oakley, J.D.; Valerie, K. Role of the p38 and MEK-1/2/p42/44 MAP kinase pathways in the differential activation of human immunodeficiency virus gene expression by ultraviolet and ionizing radiation. Photochem. Photobiol. 2000, 71, 455–459. [Google Scholar] [CrossRef]
- Choi, J.-A.; Park, M.-T.; Kang, C.-M.; Um, H.-D.; Bae, S.; Lee, K.-H.; Kim, T.-H.; Kim, J.-H.; Cho, C.-K.; Lee, Y.-S.; et al. Opposite effects of Ha-Ras and Ki-Ras on radiation-induced apoptosis via differential activation of PI3K/Akt and Rac/p38 mitogen-activated protein kinase signaling pathways. Oncogene 2004, 23, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Pal, S.; Yadav, P.; Sainis, K.B.; Shankar, B.S. TNF-α and IGF-1 differentially modulate ionizing radiation responses of lung cancer cell lines. Cytokine 2018, 101, 89–98. [Google Scholar] [CrossRef]
- Sproull, M.; Shankavaram, U.; Camphausen, K. Novel Murine Biomarkers of Radiation Exposure Using An Aptamer-Based Proteomic Technology. Front. Pharmacol. 2021, 12, 633131. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, A.N.; Hammad Khan, A.M.; Mansha, M.A.; Shaukat, F.; Tahseen, R. Abscopal Effect: An Old Concept with a New Horizon. Gulf J. Oncol. 2021, 1, 60–66. [Google Scholar]
- Ghosh, S.; Kumar, A.; Tripathi, R.P.; Chandna, S. Connexin-43 regulates p38-mediated cell migration and invasion induced selectively in tumour cells by low doses of γ-radiation in an ERK-1/2-independent manner. Carcinogenesis 2014, 35, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Asur, R.; Balasubramaniam, M.; Marples, B.; Thomas, R.A.; Tucker, J.D. Involvement of MAPK proteins in bystander effects induced by chemicals and ionizing radiation. Mutat. Res. 2010, 686, 15–29. [Google Scholar] [CrossRef]
- Hu, S.; Zhu, L.; Song, Y.; Zhao, X.; Chen, Q.; Pan, Y.; Zhang, J.; Bai, Y.; Zhang, H.; Shao, C. Radiation-induced abscopal reproductive effect is driven by TNF-α/p38 MAPK/Rac1 axis in Sertoli cells. Theranostics 2021, 11, 5742–5758. [Google Scholar] [CrossRef]
- Fan, Z.; Han, Y.; Ye, Y.; Liu, C.; Cai, H. l-carnitine preserves cardiac function by activating p38 MAPK/Nrf2 signalling in hearts exposed to irradiation. Eur. J. Pharmacol. 2017, 804, 7–12. [Google Scholar] [CrossRef]
- Song, Y.-H.; Chai, Q.; Wang, N.-L.; Yang, F.-F.; Wang, G.-H.; Hu, J.-Y. X-rays induced IL-8 production in lung cancer cells via p38/MAPK and NF-κB pathway. Int. J. Radiat. Biol. 2020, 96, 1374–1381. [Google Scholar] [CrossRef]
- Sundaramoorthy, P.; Wang, Q.; Zheng, Z.; Jiao, Y.; Chen, B.J.; Doan, P.L.; Chao, N.J.; Kang, Y. Thioredoxin mitigates radiation-induced hematopoietic stem cell injury in mice. Stem Cell Res. Ther. 2017, 8, 263. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Wang, Y.-Y.; Zhang, J.-L.; Li, D.-G.; Meng, A.-M. p38 MAPK Inhibitor Insufficiently Attenuates HSC Senescence Administered Long-Term after 6 Gy Total Body Irradiation in Mice. Int. J. Mol. Sci. 2016, 17, 905. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Wang, Y.; Wu, H.; Lu, L.; Zhang, H.; Chang, J.; Zhai, Z.; Zhang, J.; Wang, Y.; Zhou, D.; et al. Mitigation of ionizing radiation-induced bone marrow suppression by p38 inhibition and G-CSF administration. J. Radiat. Res. 2011, 52, 712–716. [Google Scholar] [CrossRef] [Green Version]
- Konkol, M.; Śniatała, P.; Milecki, P. Radiation-induced lung injury-what do we know in the era of modern radiotherapy? Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2022, 27, 552–565. [Google Scholar] [CrossRef]
- Yahyapour, R.; Amini, P.; Rezapoor, S.; Rezaeyan, A.; Farhood, B.; Cheki, M.; Fallah, H.; Najafi, M. Targeting of Inflammation for Radiation Protection and Mitigation. Curr. Mol. Pharmacol. 2018, 11, 203–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, A.; Bhuria, V.; Singh, S.; Pathak, U.; Mathur, S.; Hazari, P.P.; Roy, B.G.; Sandhir, R.; Soni, R.; Dwarakanath, B.S.; et al. Amifostine analog, DRDE-30, alleviates radiation induced lung damage by attenuating inflammation and fibrosis. Life Sci. 2022, 298, 120518. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Li, W.; Chen, L.; Su, Q.; Wang, Y.; Guo, Z.; Lu, Y.; Liu, B.; Qin, S. Radiation-induced intestinal damage: Latest molecular and clinical developments. Future Oncol. 2019, 15, 4105–4118. [Google Scholar] [CrossRef] [PubMed]
- Meena, S.K.; Joriya, P.R.; Yadav, S.M.; Kumar, R.; Meena, P.; Patel, D.D. Modulation of radiation-induced intestinal injury by radioprotective agents: A cellular and molecular perspectives. Rev. Environ. Health 2022. [Google Scholar] [CrossRef] [PubMed]
- Tanoue, T.; Yamamoto, T.; Maeda, R.; Nishida, E. A Novel MAPK phosphatase MKP-7 acts preferentially on JNK/SAPK and p38 alpha and beta MAPKs. J. Biol. Chem. 2001, 276, 26629–26639. [Google Scholar] [CrossRef] [Green Version]
- Sha, H.; Gu, Y.; Shen, W.; Zhang, L.; Qian, F.; Zhao, Y.; Li, H.; Zhang, T.; Lu, W. Rheinic acid ameliorates radiation-induced acute enteritis in rats through PPAR-γ/NF-κB. Genes Genom. 2019, 41, 909–917. [Google Scholar] [CrossRef]
- Kiang, J.G.; Cannon, G.; Olson, M.G.; Smith, J.T.; Anderson, M.N.; Zhai, M.; Umali, M.V.; Ho, K.; Ho, C.; Cui, W.; et al. Female Mice are More Resistant to the Mixed-Field (67% Neutron + 33% Gamma) Radiation-Induced Injury in Bone Marrow and Small Intestine than Male Mice due to Sustained Increases in G-CSF and the Bcl-2/Bax Ratio and Lower miR-34a and MAPK Activation. Radiat. Res. 2022, 198, 120–133. [Google Scholar] [CrossRef]
- de la Cruz-Morcillo, M.A.; García-Cano, J.; Arias-González, L.; García-Gil, E.; Artacho-Cordón, F.; Ríos-Arrabal, S.; Valero, M.L.; Cimas, F.J.; Serrano-Oviedo, L.; Villas, M.V.; et al. Abrogation of the p38 MAPK α signaling pathway does not promote radioresistance but its activity is required for 5-Fluorouracil-associated radiosensitivity. Cancer Lett. 2013, 335, 66–74. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Yang, L.; Yin, D. Intermittent low dose irradiation enhances the effectiveness of radio- and chemo-therapy for human colorectal adenocarcinoma cell line HT-29. Oncol. Rep. 2017, 38, 591–597. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Useros, J.; Moreno, I.; Fernandez-Aceñero, M.J.; Rodriguez-Remirez, M.; Borrero-Palacios, A.; Cebrian, A.; Gomez del Pulgar, T.; del Puerto-Nevado, L.; Li, W.; Puime-Otin, A.; et al. The potential predictive value of DEK expression for neoadjuvant chemoradiotherapy response in locally advanced rectal cancer. BMC Cancer 2018, 18, 144. [Google Scholar] [CrossRef] [Green Version]
- Pascual-Serra, R.; Fernández-Aroca, D.M.; Sabater, S.; Roche, O.; Andrés, I.; Ortega-Muelas, M.; Arconada-Luque, E.; García-Flores, N.; Bossi, G.; Belandia, B.; et al. p38β (MAPK11) mediates gemcitabine-associated radiosensitivity in sarcoma experimental models. Radiother. Oncol. 2021, 156, 136–144. [Google Scholar] [CrossRef]
- Ho, S.-Y.; Wu, W.-J.; Chiu, H.-W.; Chen, Y.-A.; Ho, Y.-S.; Guo, H.-R.; Wang, Y.-J. Arsenic trioxide and radiation enhance apoptotic effects in HL-60 cells through increased ROS generation and regulation of JNK and p38 MAPK signaling pathways. Chem.-Biol. Interact. 2011, 193, 162–171. [Google Scholar] [CrossRef]
- Kale, Ş.; Korcum, A.F.; Dündar, E.; Erin, N. HSP90 inhibitor PU-H71 increases radiosensitivity of breast cancer cells metastasized to visceral organs and alters the levels of inflammatory mediators. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2020, 393, 253–262. [Google Scholar] [CrossRef]
- Hur, J.-M.; Hyun, M.-S.; Lim, S.-Y.; Lee, W.-Y.; Kim, D. The combination of berberine and irradiation enhances anti-cancer effects via activation of p38 MAPK pathway and ROS generation in human hepatoma cells. J. Cell. Biochem. 2009, 107, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, P.S.; Guimarães, G.H.C.; De Faria, F.C.C.; Longo, G.M.D.C.; Lopes, G.P.D.F.; Netto, C.D.; Costa, P.R.R.; Maia, R.C. LQB-118 compound inhibits migration and induces cell death in glioblastoma cells. Oncol. Rep. 2020, 43, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.-J.; Ahn, K.-C.; Choi, J.Y.; Hwang, S.-G.; Kim, W.-J.; Um, H.-D.; Park, J.K. Luteolin acts as a radiosensitizer in non-small cell lung cancer cells by enhancing apoptotic cell death through activation of a p38/ROS/caspase cascade. Int. J. Oncol. 2015, 46, 1149–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, S.; Qiao, T.; Li, X.; Zhuang, X.; Chen, W.; Chen, X.; Zhang, Q. Toll-like receptor 9 activation by CpG oligodeoxynucleotide 7909 enhances the radiosensitivity of A549 lung cancer cells via the p53 signaling pathway. Oncol. Lett. 2018, 15, 5271–5279. [Google Scholar] [CrossRef] [Green Version]
- Cheol Son, J.; Woo Kang, D.; Mo Yang, K.; Choi, K.-Y.; Gen Son, T.; Min, D.S. Phospholipase D inhibitor enhances radiosensitivity of breast cancer cells. Exp. Mol. Med. 2013, 45, e38. [Google Scholar] [CrossRef]
- Camero, S.; Vitali, G.; Pontecorvi, P.; Ceccarelli, S.; Anastasiadou, E.; Cicchetti, F.; Flex, E.; Pomella, S.; Cassandri, M.; Rota, R.; et al. DNMT3A and DNMT3B Targeting as an Effective Radiosensitizing Strategy in Embryonal Rhabdomyosarcoma. Cells 2021, 10, 2956. [Google Scholar] [CrossRef]
- Wang, K.; Tang, J.; Liu, X.; Wang, Y.; Chen, W.; Zheng, R. UBR5 regulates proliferation and radiosensitivity in human laryngeal carcinoma via the p38/MAPK signaling pathway. Oncol. Rep. 2020, 44, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, W.; Dou, M.; Li, Z.; Liu, Z.; Li, J.; Tian, C.; Yao, Y.; Wang, C.; Li, Y.; et al. 125I seeds inhibit proliferation and promote apoptosis in cholangiocarcinoma cells by regulating the AGR2-mediated p38 MAPK pathway. Cancer Lett. 2022, 524, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Wang, L.-X.; Yang, M.-L.; Zhang, R.-M. lncRNA TPTEP1 inhibits stemness and radioresistance of glioma through miR-106a-5p-mediated P38 MAPK signaling. Mol. Med. Rep. 2020, 22, 4857–4867. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Shen, Y.; Wei, J.; Liu, F. MicroRNA-153/Nrf-2/GPx1 pathway regulates radiosensitivity and stemness of glioma stem cells via reactive oxygen species. Oncotarget 2015, 6, 22006–22027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biau, J.; Thivat, E.; Chautard, E.; Stefan, D.; Boone, M.; Chauffert, B.; Bourgne, C.; Richard, D.; Molnar, I.; Levesque, S.; et al. Phase 1 trial of ralimetinib (LY2228820) with radiotherapy plus concomitant temozolomide in the treatment of newly diagnosed glioblastoma. Radiother. Oncol. 2021, 154, 227–234. [Google Scholar] [CrossRef]
- Denny, W.A. Inhibitors and Activators of the p38 Mitogen-Activated MAP Kinase (MAPK) Family as Drugs to Treat Cancer and Inflammation. Curr. Cancer Drug Targets 2022, 22, 209–220. [Google Scholar] [CrossRef]
- Donoghue, C.; Cubillos-Rojas, M.; Gutierrez-Prat, N.; Sanchez-Zarzalejo, C.; Verdaguer, X.; Riera, A.; Nebreda, Á.R. Optimal linker length for small molecule PROTACs that selectively target p38α and p38β for degradation. Eur. J. Med. Chem. 2020, 201, 112451. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Flores, N.; Jiménez-Suárez, J.; Garnés-García, C.; Fernández-Aroca, D.M.; Sabater, S.; Andrés, I.; Fernández-Aramburo, A.; Ruiz-Hidalgo, M.J.; Belandia, B.; Sanchez-Prieto, R.; et al. P38 MAPK and Radiotherapy: Foes or Friends? Cancers 2023, 15, 861. https://doi.org/10.3390/cancers15030861
García-Flores N, Jiménez-Suárez J, Garnés-García C, Fernández-Aroca DM, Sabater S, Andrés I, Fernández-Aramburo A, Ruiz-Hidalgo MJ, Belandia B, Sanchez-Prieto R, et al. P38 MAPK and Radiotherapy: Foes or Friends? Cancers. 2023; 15(3):861. https://doi.org/10.3390/cancers15030861
Chicago/Turabian StyleGarcía-Flores, Natalia, Jaime Jiménez-Suárez, Cristina Garnés-García, Diego M. Fernández-Aroca, Sebastia Sabater, Ignacio Andrés, Antonio Fernández-Aramburo, María José Ruiz-Hidalgo, Borja Belandia, Ricardo Sanchez-Prieto, and et al. 2023. "P38 MAPK and Radiotherapy: Foes or Friends?" Cancers 15, no. 3: 861. https://doi.org/10.3390/cancers15030861
APA StyleGarcía-Flores, N., Jiménez-Suárez, J., Garnés-García, C., Fernández-Aroca, D. M., Sabater, S., Andrés, I., Fernández-Aramburo, A., Ruiz-Hidalgo, M. J., Belandia, B., Sanchez-Prieto, R., & Cimas, F. J. (2023). P38 MAPK and Radiotherapy: Foes or Friends? Cancers, 15(3), 861. https://doi.org/10.3390/cancers15030861