
Phenolic Phytochemicals for Prevention and Treatment of Colorectal Cancer: A Critical Evaluation of In Vivo Studies
 , , , ,
, , , ,  ,
,  , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Risk Factors
3. Pathogenesis
4. Chemoprevention
5. Treatment
6. Literature Search Methodology
7. Phenolic Compounds with In Vivo Anti-CRC Activities
7.1. Flavonoids
7.1.1. Baicalin
7.1.2. Curcumin
7.1.3. Catechins
7.1.4. Fisetin
7.1.5. Genistein
7.1.6. Kaempferol
7.1.7. Luteolin
7.1.8. Myricetin
7.1.9. Naringenin
7.1.10. Quercetin
7.1.11. Rutin
7.1.12. Tangeretin
7.1.13. Wogonin
7.2. Phenolic Acids
7.2.1. Caffeic Acid
7.2.2. Gallic Acid
7.3. Stilbenes
Resveratrol
8. Phenolics in Clinical Trials for CRC Treatment
9. Conclusions and Future Perspectives
- 1.
- Apparently, the functions of many phytochemicals are limited by their poor solubility, absorption, and bioavailability. Encapsulation by nano-formulation as well as chemical derivatization of the compound could resolve this issue.
- 2.
- Some cases reproducing the activity observed in preclinical animal models into the clinic/human could be challenging due to several factors. Success in this endeavor requires careful optimization in administered doses to assess functional synergy, if any, with anti-CRC regimens used in the clinic. Once positive results are obtained in the preclinical settings, testing the validity of the finding, such as safety and efficacy, in clinical trials with appropriate controls will be important to move further.
- 3.
- It is reasonable to think that a phenolic compound showing very weak and toxic activity can yield desirable effect when combined with another phytochemical. Therefore, a careful combination of selected polyphenols can yield unique anti-CRC activity. It is important to clearly determine the maximum tolerable dose of a phytochemical to better understand its therapeutic efficacy alone or in combination with another phytochemical or drug.
- 4.
- Once a phenolic compound with unique anti-CRC activity is identified, it would be important to develop strategies to synthesize the compound in the laboratory, given the very low abundance of a secondary metabolite in the plants. A detailed understanding of the pharmacophore responsible for the observed function should be helpful for chemical synthesis or semi-synthesis, and cellular target identification of the compound. Given the structural complexity of the plant secondary metabolites, it is often a major challenge for natural product chemists and medicinal chemists to solve. Ideally, the simultaneous engagement of experts from interdisciplinary areas, such as ethnopharmacology, molecular biology, biochemistry, natural product chemistry, medicinal chemistry, bioinformatics, and pharmacology, will be necessary to achieve progress in real-time in harvesting the full potential of natural products as the source of novel drug leads.
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 5-FU | 5-fluorouracil |
| 5′ND | 5-nucleotidase |
| 6CEPN | 6-C-(E-phenylethyl)-naringenin |
| ACF | aberrant crypt foci |
| AOM | azoxymethane |
| APC | adenomatous polyposis coli |
| ATST | atorvastatin |
| BAX | B-cell lymphoma 2-associated x protein |
| BCL-2 | B-cell lymphoma 2 |
| BID | BH3 interacting-domain death agonist |
| BRAF-B | rapidly accelerated fibrosarcoma/murine sarcoma viral oncogene homolog B |
| CAC | colitis-associated colorectal cancer |
| CACNA14 | voltage-dependent P/Q type calcium channel subunit alpha1A |
| CAP | capecitabine |
| CAPE, | caffeic acid phenethyl ester |
| CAPPE | caffeic acid phenylpropyl ester |
| CAPE-pNO2 | caffeic acid p-nitro-phenylethyl ester |
| CAPIRI | capecitabine and irinotecan |
| CAPOX | capecitabine and oxaliplatin |
| CEA | carcinoembryonic antigen |
| CIMP | CpG island methylation phenotype |
| CIN | chromosomal instability |
| CLXC12 | C–X–C chemokine 12 |
| COX-2 | cyclooxygenase-2 |
| CRC | colorectal cancer |
| CSC | cancer stem cell |
| cyt. c | cytochrome c |
| DII | dietary inflammatory index |
| Dkk | Dickkopf |
| DMH | dimethyl hydrazine |
| DNMT | DNA methyltransferase |
| DSS | dextran sulphate sodium |
| Dvl | Discevelled |
| EGF-β | epidermal growth factor-β |
| EGFR | epidermal growth factor receptor |
| ER | endoplasmic reticulum |
| ERK | extracellular signal-regulated kinase |
| FAP | familial adenomatous polyposis syndrome |
| FDA | Federal Drug Administration |
| FOLFOX | 5-FU and oxaliplatin |
| FOXFIRI | 5-FU and irinotecan |
| FZD | Frizzled receptor |
| G6PD | glucose-6-phosphate dehydrogenase |
| GEPIA | gene expression profiling interactive analysis |
| GGT | γ-glutamyl transferase |
| GSH | glutathione |
| GSK-3β | glycogen synthase kinase-3β |
| GTP | green tea polyphenol |
| HMACF | high multiplicity aberrant crypt foci |
| hMLH1 | human MutL homolog 1 |
| HPP | hyperplastic polyposis |
| IDEA | International Duration Evaluation of Adjuvant Chemotherapy |
| IGF-2 | insulin like growth factor-2 |
| IGFBP3 | insulin like growth factor-binding protein 3 |
| iNOS | inducible nitric oxide synthase |
| IRI | irinotecan |
| KRAS | Kirsten rat sarcoma viral oncogene homolog |
| LC3B | light chain 3B of microtubule-associated proteins 1A/1B |
| LPO | lipid peroxide |
| MAP | MUTYG- associated polyposis |
| MAPK | mitogen-activated protein kinase |
| MMP | matrix metalloproteinase |
| MSI | microsatellite instability |
| mTOR | mammalian target of rapamycin |
| NF-κβ | nuclear factor-κβ |
| NO | nitric oxide |
| NSAID | nonsteroidal anti-inflammatory drug |
| oncomiRs | oncogenic miRNAs |
| OPE | orange peel extract |
| OX | oxaliplatin |
| PDTX | patient-derived tumor xenograft |
| PI3K | phosphoinositide 3-kinase |
| PPE | polyphenol E |
| PTEN | phosphatase and tensin homolog deleted on chromosome 10 |
| SEER | surveillance epidemiology and end results |
| SGK1 | serum glucocorticoid kinase 1 |
| Skip | Ski-interacting protein |
| SOD | superoxide dismutase |
| SPI | soya protein isolate |
| TAGE | tangeretin |
| TIMP | tissue inhibitor metalloproteinase |
| TNM | tumor/node/metastasis |
| TRPV1 | transient receptor potential vanilloid 1 |
| VEGF | vascular endothelial growth factor |
| VEGFR | vascular endothelial growth factor receptor |
| YAP | yes-associated protein |
| yCRC | young-onset colorectal cancer |
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.; Piñeros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.; Sierra, M.S.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global patterns and trends in colorectal cancer incidence and mortality. Gut 2017, 66, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Haque, A.; Brazeau, D.; Amin, A.R. Perspectives on natural compounds in chemoprevention and treatment of cancer: An update with new promising compounds. Eur. J. Cancer 2021, 149, 165–183. [Google Scholar] [CrossRef]
- Swetha, M.; Keerthana, C.; Rayginia, T.P.; Anto, R.J. Cancer chemoprevention: A strategic approach using phytochemicals. Front. Pharmacol. 2021, 12, 809308. [Google Scholar]
- Dhillon, P.K.; Mathur, P.; Nandakumar, A.; Fitzmaurice, C.; Kumar, G.A.; Mehrotra, R.; Shukla, D.; Rath, G.; Gupta, P.C.; Swaminathan, R. The burden of cancers and their variations across the states of India: The Global Burden of Disease Study 1990–2016. Lancet Oncol. 2018, 19, 1289–1306. [Google Scholar] [CrossRef]
- Choudhari, A.S.; Mandave, P.C.; Deshpande, M.; Ranjekar, P.; Prakash, O. Phytochemicals in cancer treatment: From preclinical studies to clinical practice. Front. Pharmacol. 2020, 10, 1614. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Goding Sauer, A.; Fedewa, S.A.; Butterly, L.F.; Anderson, J.C.; Cercek, A.; Smith, R.A.; Jemal, A. Colorectal cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 145–164. [Google Scholar] [CrossRef]
- Ranjan, A.; Ramachandran, S.; Gupta, N.; Kaushik, I.; Wright, S.; Srivastava, S.; Das, H.; Srivastava, S.; Prasad, S.; Srivastava, S.K. Role of phytochemicals in cancer prevention. Int. J. Mol. Sci. 2019, 20, 4981. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fedewa, S.A.; Ahnen, D.J.; Meester, R.G.; Barzi, A.; Jemal, A. Colorectal cancer statistics, 2017. CA Cancer J. Clin. 2017, 67, 177–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauri, G.; Sartore-Bianchi, A.; Russo, A.G.; Marsoni, S.; Bardelli, A.; Siena, S. Early-onset colorectal cancer in young individuals. Mol. Oncol. 2019, 13, 109–131. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.C.; Lund, J.L.; Sandler, R.S. Young-onset colorectal cancer: Earlier diagnoses or increasing disease burden? Gastroenterology 2017, 152, 1809–1812.e1803. [Google Scholar] [CrossRef] [PubMed]
- You, Y.N.; Xing, Y.; Feig, B.W.; Chang, G.J.; Cormier, J.N. Young-onset colorectal cancer: Is it time to pay attention? Arch. Intern. Med. 2012, 172, 287–289. [Google Scholar] [CrossRef] [PubMed]
- Holowatyj, A.N.; Ruterbusch, J.J.; Rozek, L.S.; Cote, M.L.; Stoffel, E.M. Racial/ethnic disparities in survival among patients with young-onset colorectal cancer. J. Clin. Oncol. 2016, 34, 2148. [Google Scholar] [CrossRef] [PubMed]
- Thanikachalam, K.; Khan, G. Colorectal cancer and nutrition. Nutrients 2019, 11, 164. [Google Scholar] [CrossRef]
- Tanaka, S.; Oka, S.; Chayama, K. Colorectal endoscopic submucosal dissection: Present status and future perspective, including its differentiation from endoscopic mucosal resection. J. Gastroenterol. 2008, 43, 641–651. [Google Scholar] [CrossRef]
- Chakedis, J.; Schmidt, C.R. Surgical treatment of metastatic colorectal cancer. Surg. Oncol. Clin. 2018, 27, 377–399. [Google Scholar] [CrossRef]
- Mojtahedi, Z.; Koo, J.S.; Yoo, J.; Kim, P.; Kang, H.-T.; Hwang, J.; Joo, M.K.; Shen, J.J. Palliative care and life-sustaining/local procedures in colorectal Cancer in the United States hospitals: A ten-year perspective. Cancer Manag. Res. 2021, 13, 7569–7577. [Google Scholar] [CrossRef]
- Townsend, A.; Price, T.; Karapetis, C. Selective internal radiation therapy for liver metastases from colorectal cancer. Cochrane Database Syst. Rev. 2009, CD007045. [Google Scholar] [CrossRef]
- Kanani, A.; Veen, T.; Søreide, K. Neoadjuvant immunotherapy in primary and metastatic colorectal cancer. Br. J. Surg. 2021, 108, 1417–1425. [Google Scholar] [CrossRef]
- Loughrey, M.B. Neoadjuvant immunotherapy and colorectal cancer treatment: Implications for the primary role of surgery. Color. Dis. Off. J. Assoc. Coloproctol. Great Br. Irel. 2022, 24, 1460–1461. [Google Scholar] [CrossRef]
- Aiello, P.; Sharghi, M.; Mansourkhani, S.M.; Ardekan, A.P.; Jouybari, L.; Daraei, N.; Peiro, K.; Mohamadian, S.; Rezaei, M.; Heidari, M. Medicinal plants in the prevention and treatment of colon cancer. Oxidative Med. Cell. Longev. 2019, 2019, 2075614. [Google Scholar] [CrossRef] [PubMed]
- Muppala, S. Phytochemicals Targeting Colorectal Cancer Growth and Metastasis. Crit. Rev. Oncog. 2020, 25, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Sain, A.; Sahu, S.; Naskar, D. Potential of Olive oil and its phenolic compounds as therapeutic intervention against colorectal cancer: A comprehensive review. Br. J. Nutr. 2021, 128, 1257–1273. [Google Scholar] [CrossRef] [PubMed]
- Cueva, C.; Silva, M.; Pinillos, I.; Bartolomé, B.; Moreno-Arribas, M.V. Interplay between dietary polyphenols and oral and gut microbiota in the development of colorectal cancer. Nutrients 2020, 12, 625. [Google Scholar] [CrossRef]
- Andrews, L. Dietary flavonoids for the prevention of colorectal cancer. Clin. J. Oncol. Nurs. 2013, 17, 671–673. [Google Scholar] [CrossRef]
- Woo, H.D.; Kim, J. Dietary flavonoid intake and risk of stomach and colorectal cancer. World J. Gastroenterol. 2013, 19, 1011. [Google Scholar] [CrossRef]
- Kocic, B.; Kitic, D.; Brankovic, S. Dietary flavonoid intake and colorectal cancer risk: Evidence from human population studies. J. BUON 2013, 18, 34–43. [Google Scholar]
- Jin, H.; Leng, Q.; Li, C. Dietary flavonoid for preventing colorectal neoplasms. Cochrane Database Syst. Rev. 2012, CD009350. [Google Scholar] [CrossRef]
- Han, S.; Cao, Y.; Guo, T.; Lin, Q.; Luo, F. Targeting lncRNA/Wnt axis by flavonoids: A promising therapeutic approach for colorectal cancer. Phytother. Res. 2022, 36, 4024–4040. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Wilson, C. Can dietary flavonoids be useful in the personalized treatment of colorectal cancer? World J. Gastrointest. Oncol. 2022, 14, 1115. [Google Scholar] [CrossRef] [PubMed]
- Fernández, J.; Silván, B.; Entrialgo-Cadierno, R.; Villar, C.J.; Capasso, R.; Uranga, J.A.; Lombó, F.; Abalo, R. Antiproliferative and palliative activity of flavonoids in colorectal cancer. Biomed. Pharmacother. 2021, 143, 112241. [Google Scholar] [CrossRef] [PubMed]
- Afshari, K.; Haddadi, N.S.; Haj-Mirzaian, A.; Farzaei, M.H.; Rohani, M.M.; Akramian, F.; Naseri, R.; Sureda, A.; Ghanaatian, N.; Abdolghaffari, A.H. Natural flavonoids for the prevention of colon cancer: A comprehensive review of preclinical and clinical studies. J. Cell. Physiol. 2019, 234, 21519–21546. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, T.; Chen, G.Y. Flavonoids and colorectal cancer prevention. Antioxidants 2018, 7, 187. [Google Scholar] [CrossRef]
- Koosha, S.; Alshawsh, M.A.; Looi, C.Y.; Seyedan, A.; Mohamed, Z. An association map on the effect of flavonoids on the signaling pathways in colorectal cancer. Int. J. Med. Sci. 2016, 13, 374. [Google Scholar] [CrossRef]
- Potter, J.D. Nutrition and colorectal cancer. Cancer Causes Control 1996, 7, 127–146. [Google Scholar] [CrossRef]
- Martinez, M.E.; Willett, W.C. Calcium, vitamin D, and colorectal cancer: A review of the epidemiologic evidence. Cancer Epidemiol. Biomark. Prev. 1998, 7, 163–168. [Google Scholar]
- Terry, P.; Giovannucci, E.; Michels, K.B.; Bergkvist, L.; Hansen, H.; Holmberg, L.; Wolk, A. Fruit, vegetables, dietary fiber, and risk of colorectal cancer. J. Natl. Cancer Inst. 2001, 93, 525–533. [Google Scholar] [CrossRef]
- Santarelli, R.L.; Pierre, F.; Corpet, D.E. Processed meat and colorectal cancer: A review of epidemiologic and experimental evidence. Nutr. Cancer 2008, 60, 131–144. [Google Scholar]
- Benarba, B.; Pandiella, A. Colorectal cancer and medicinal plants: Principle findings from recent studies. Biomed. Pharmacother. 2018, 107, 408–423. [Google Scholar] [CrossRef] [PubMed]
- La Vecchia, S.; Sebastián, C. Metabolic pathways regulating colorectal cancer initiation and progression. Semin. Cell Dev.Biol. 2020, 98, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Keum, N.; Giovannucci, E. Global burden of colorectal cancer: Emerging trends, risk factors and prevention strategies. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 713–732. [Google Scholar] [CrossRef] [PubMed]
- Lynch, H.T.; Smyrk, T.C.; Watson, P.; Lanspa, S.J.; Lynch, J.F.; Lynch, P.M.; Cavalieri, R.J.; Boland, C.R. Genetics, natural history, tumor spectrum, and pathology of hereditary nonpolyposis colorectal cancer: An updated review. Gastroenterology 1993, 104, 1535–1549. [Google Scholar] [CrossRef] [PubMed]
- Niessen, R.C.; Berends, M.J.; Wu, Y.; Sijmons, R.H.; Hollema, H.; Ligtenberg, M.J.; de Walle, H.E.; de Vries, E.G.; Karrenbeld, A.; Buys, C.H. Identification of mismatch repair gene mutations in young patients with colorectal cancer and in patients with multiple tumours associated with hereditary non-polyposis colorectal cancer. Gut 2006, 55, 1781–1788. [Google Scholar] [CrossRef] [PubMed]
- Seppälä, T.; Latchford, A.; Negoi, I.; Sampaio Soares, A.; Jimenez-Rodriguez, R.; Sánchez-Guillén, L.; Evans, D.; Ryan, N.; Crosbie, E.; Dominguez-Valentin, M. European guidelines from the EHTG and ESCP for Lynch syndrome: An updated third edition of the Mallorca guidelines based on gene and gender. Br. J. Surg. 2021, 108, 484–498. [Google Scholar] [CrossRef]
- Møller, P.; Seppälä, T.T.; Bernstein, I.; Holinski-Feder, E.; Sala, P.; Evans, D.G.; Lindblom, A.; Macrae, F.; Blanco, I.; Sijmons, R.H. Cancer risk and survival in path_MMR carriers by gene and gender up to 75 years of age: A report from the Prospective Lynch Syndrome Database. Gut 2018, 67, 1306–1316. [Google Scholar] [CrossRef]
- Ponti, G.; Manfredini, M.; Tomasi, A.; Pellacani, G. Muir–Torre Syndrome and founder mismatch repair gene mutations: A long gone historical genetic challenge. Gene 2016, 589, 127–132. [Google Scholar] [CrossRef]
- Grzybowski, A.; Jablonska, S. Muir–Torre Syndrome—Is It Really a New Syndrome? Am. J. Dermatopathol. 2009, 31, 799–802. [Google Scholar] [CrossRef]
- Kidambi, T.D.; Kohli, D.R.; Samadder, N.J.; Singh, A. Hereditary polyposis syndromes. Curr. Treat. Options Gastroenterol. 2019, 17, 650–665. [Google Scholar] [CrossRef]
- Hamilton, S.R.; Liu, B.; Parsons, R.E.; Papadopoulos, N.; Jen, J.; Powell, S.M.; Krush, A.J.; Berk, T.; Cohen, Z.; Tetu, B. The molecular basis of Turcot’s syndrome. N. Engl. J. Med. 1995, 332, 839–847. [Google Scholar] [CrossRef]
- Rutz, H.P.; de Tribolet, N.; Calmes, J.M.; Chapuis, G. Long-time survival of a patient with glioblastoma and Turcot’s syndrome: Case report. J. Neurosurg. 1991, 74, 813–815. [Google Scholar] [CrossRef] [PubMed]
- Haggar, F.A.; Boushey, R.P. Colorectal cancer epidemiology: Incidence, mortality, survival, and risk factors. Clin. Colon Rectal Surg. 2009, 22, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Galiatsatos, P.; Foulkes, W.D. Familial adenomatous polyposis. Off. J. Am. Coll. Gastroenterol. ACG 2006, 101, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Coffin, C.M.; Hornick, J.L.; Zhou, H.; Fletcher, C.D. Gardner fibroma: A clinicopathologic and immunohistochemical analysis of 45 patients with 57 fibromas. Am. J. Surg. Pathol. 2007, 31, 410–416. [Google Scholar] [CrossRef]
- Kiessling, P.; Dowling, E.; Huang, Y.; Ho, M.L.; Balakrishnan, K.; Weigel, B.J.; Highsmith, W.E.; Niu, Z.; Schimmenti, L.A. Identification of aggressive Gardner syndrome phenotype associated with a de novo APC variant, c. 4666dup. Mol. Case Stud. 2019, 5, a003640. [Google Scholar] [CrossRef] [PubMed]
- Sutcliffe, E.G.; Thompson, A.B.; Stettner, A.R.; Marshall, M.L.; Roberts, M.E.; Susswein, L.R.; Wang, Y.; Klein, R.T.; Hruska, K.S.; Solomon, B.D. Multi-gene panel testing confirms phenotypic variability in MUTYH-Associated Polyposis. Fam. Cancer 2019, 18, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Curia, M.C.; Catalano, T.; Aceto, G.M. MUTYH: Not just polyposis. World J. Clin. Oncol. 2020, 11, 428. [Google Scholar] [CrossRef]
- Vogt, S.; Jones, N.; Christian, D.; Engel, C.; Nielsen, M.; Kaufmann, A.; Steinke, V.; Vasen, H.F.; Propping, P.; Sampson, J.R. Expanded extracolonic tumor spectrum in MUTYH-associated polyposis. Gastroenterology 2009, 137, 1976–1985.e1910. [Google Scholar] [CrossRef]
- Calva-Cerqueira, D.; Chinnathambi, S.; Pechman, B.; Bair, J.; Larsen-Haidle, J.; Howe, J. The rate of germline mutations and large deletions of SMAD4 and BMPR1A in juvenile polyposis. Clin. Genet. 2009, 75, 79–85. [Google Scholar] [CrossRef]
- Gallione, C.J.; Repetto, G.M.; Legius, E.; Rustgi, A.K.; Schelley, S.L.; Tejpar, S.; Mitchell, G.; Drouin, É.; Westermann, C.J.; Marchuk, D.A. A combined syndrome of juvenile polyposis and hereditary haemorrhagic telangiectasia associated with mutations in MADH4 (SMAD4). Lancet 2004, 363, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Brosens, L.A.; Van Hattem, A.; Hylind, L.M.; Iacobuzio-Donahue, C.; Romans, K.E.; Axilbund, J.; Cruz-Correa, M.; Tersmette, A.C.; Offerhaus, G.J.A.; Giardiello, F.M. Risk of colorectal cancer in juvenile polyposis. Gut 2007, 56, 965–967. [Google Scholar] [CrossRef]
- Kopacova, M.; Tacheci, I.; Rejchrt, S.; Bures, J. Peutz-Jeghers syndrome: Diagnostic and therapeutic approach. World J. Gastroenterol. 2009, 15, 5397. [Google Scholar] [CrossRef] [PubMed]
- Westerman, A.M.; Entius, M.M.; De Baar, E.; Boor, P.P.; Koole, R.; Van Velthuysen, M.L.F.; Offerhaus, G.J.A.; Lindhout, D.; De Rooij, F.W.; Wilson, J.P. Peutz-Jeghers syndrome: 78-year follow-up of the original family. Lancet 1999, 353, 1211–1215. [Google Scholar] [CrossRef]
- Smerdel, M.P.; Skytte, A.-B.; Jelsig, A.M.; Ebbehøj, E.; Stochholm, K. Revised Danish guidelines for the cancer surveillance of patients with Cowden Syndrome. Eur. J. Med. Genet. 2020, 63, 103873. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, K.M.; Dennis, M. Cowden’s disease: A possible new symptom complex with multiple system involvement. Ann. Intern. Med. 1963, 58, 136–142. [Google Scholar] [CrossRef]
- Miyaki, M.; Konishi, M.; Kikuchi-Yanoshita, R.; Enomoto, M.; Igari, T.; Tanaka, K.; Muraoka, M.; Takahashi, H.; Amada, Y.; Fukayama, M. Characteristics of somatic mutation of the adenomatous polyposis coli gene in colorectal tumors. Cancer Res. 1994, 54, 3011–3020. [Google Scholar]
- Lamlum, H.; Ilyas, M.; Rowan, A.; Clark, S.; Johnson, V.; Bell, J.; Frayling, I.; Efstathiou, J.; Pack, K.; Payne, S. The type of somatic mutation at APC in familial adenomatous polyposis is determined by the site of the germline mutation: A new facet to Knudson’s’ two-hit’hypothesis. Nat. Med. 1999, 5, 1071–1075. [Google Scholar] [CrossRef]
- Fodde, R.; Smits, R.; Clevers, H. APC, signal transduction and genetic instability in colorectal cancer. Nat. Rev. Cancer 2001, 1, 55–67. [Google Scholar] [CrossRef]
- Kanth, P.; Grimmett, J.; Champine, M.; Burt, R.; Samadder, J.N. Hereditary colorectal polyposis and cancer syndromes: A primer on diagnosis and management. Off. J. Am. Coll. Gastroenterol. ACG 2017, 112, 1509–1525. [Google Scholar] [CrossRef]
- Mokarram, P.; Albokashy, M.; Zarghooni, M.; Moosavi, M.A.; Sepehri, Z.; Chen, Q.M.; Hudecki, A.; Sargazi, A.; Alizadeh, J.; Moghadam, A.R. New frontiers in the treatment of colorectal cancer: Autophagy and the unfolded protein response as promising targets. Autophagy 2017, 13, 781–819. [Google Scholar] [CrossRef] [PubMed]
- Jasperson, K.W.; Tuohy, T.M.; Neklason, D.W.; Burt, R.W. Hereditary and familial colon cancer. Gastroenterology 2010, 138, 2044–2058. [Google Scholar] [CrossRef]
- Potack, J.; Itzkowitz, S.H. Colorectal cancer in inflammatory bowel disease. Gut Liver 2008, 2, 61. [Google Scholar] [CrossRef] [PubMed]
- Armenian, S.H.; Robison, L.L. Childhood cancer survivorship: An update on evolving paradigms for understanding pathogenesis and screening for therapy-related late effects. Curr. Opin. Pediatr. 2013, 25, 16. [Google Scholar] [CrossRef] [PubMed]
- Baxter, N.N.; Goldwasser, M.A.; Paszat, L.F.; Saskin, R.; Urbach, D.R.; Rabeneck, L. Association of colonoscopy and death from colorectal cancer. Ann. Intern. Med. 2009, 150, 1–8. [Google Scholar] [CrossRef]
- Hadjiliadis, D.; Khoruts, A.; Zauber, A.G.; Hempstead, S.E.; Maisonneuve, P.; Lowenfels, A.B.; Braid, A.L.; Cullina, J.; Daggett, A.; Fink, A. Cystic fibrosis colorectal cancer screening consensus recommendations. Gastroenterology 2018, 154, 736–745.e714. [Google Scholar] [CrossRef]
- Botteri, E.; Iodice, S.; Bagnardi, V.; Raimondi, S.; Lowenfels, A.B.; Maisonneuve, P. Smoking and colorectal cancer: A meta-analysis. JAMA 2008, 300, 2765–2778. [Google Scholar] [CrossRef]
- Cai, S.; Li, Y.; Ding, Y.; Chen, K.; Jin, M. Alcohol drinking and the risk of colorectal cancer death: A meta-analysis. Eur. J. Cancer Prev. 2014, 23, 532–539. [Google Scholar] [CrossRef]
- Kyrgiou, M.; Kalliala, I.; Markozannes, G.; Gunter, M.J.; Paraskevaidis, E.; Gabra, H.; Martin-Hirsch, P.; Tsilidis, K.K. Adiposity and cancer at major anatomical sites: Umbrella review of the literature. BMJ 2017, 356, j477. [Google Scholar] [CrossRef]
- Wong, M.C.; Ding, H.; Wang, J.; Chan, P.S.; Huang, J. Prevalence and risk factors of colorectal cancer in Asia. Intest. Res. 2019, 17, 317. [Google Scholar] [CrossRef]
- Shivappa, N.; Godos, J.; Hébert, J.R.; Wirth, M.D.; Piuri, G.; Speciani, A.F.; Grosso, G. Dietary inflammatory index and colorectal cancer risk—A meta-analysis. Nutrients 2017, 9, 1043. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Sun, J.; Yu, J.; Wang, C.; Su, J. Dietary intakes of calcium, iron, magnesium, and potassium elements and the risk of colorectal cancer: A meta-analysis. Biol. Trace Elem. Res. 2019, 189, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Parra-Soto, S.; Ahumada, D.; Petermann-Rocha, F.; Boonpoor, J.; Gallegos, J.L.; Anderson, J.; Sharp, L.; Malcomson, F.C.; Livingstone, K.M.; Mathers, J.C. Association of meat, vegetarian, pescatarian and fish-poultry diets with risk of 19 cancer sites and all cancer: Findings from the UK Biobank prospective cohort study and meta-analysis. BMC Med. 2022, 20, 1–16. [Google Scholar] [CrossRef]
- Zhong, Y.; Zhu, Y.; Li, Q.; Wang, F.; Ge, X.; Zhou, G.; Miao, L. Association between Mediterranean diet adherence and colorectal cancer: A dose-response meta-analysis. Am. J. Clin. Nutr. 2020, 111, 1214–1225. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.; Kim, H.; Kim, J. Dietary intake in association with all-cause mortality and colorectal cancer mortality among colorectal cancer survivors: A systematic review and meta-analysis of prospective studies. Cancers 2020, 12, 3391. [Google Scholar] [CrossRef]
- Cho, K.R.; Vogelstein, B. Genetic alterations in the adenoma–carcinoma sequence. Cancer 1992, 70, 1727–1731. [Google Scholar] [CrossRef]
- Cho, K.R.; Vogelstein, B. Suppressor gene alterations in the colorectal adenoma-carcinoma sequence. J. Cell. Biochem. 1992, 50, 137–141. [Google Scholar] [CrossRef]
- Tariq, K.; Ghias, K. Colorectal cancer carcinogenesis: A review of mechanisms. Cancer Biol. Med. 2016, 13, 120. [Google Scholar] [CrossRef]
- Hermsen, M.; Postma, C.; Baak, J.; Weiss, M.; Rapallo, A.; Sciutto, A.; Roemen, G.; Arends, J.W.; Williams, R.; Giaretti, W. Colorectal adenoma to carcinoma progression follows multiple pathways of chromosomal instability. Gastroenterology 2002, 123, 1109–1119. [Google Scholar] [CrossRef]
- Baran, B.; Ozupek, N.M.; Tetik, N.Y.; Acar, E.; Bekcioglu, O.; Baskin, Y. Difference between left-sided and right-sided colorectal cancer: A focused review of literature. Gastroenterol. Res. 2018, 11, 264. [Google Scholar] [CrossRef]
- Gryfe, R.; Bapat, B.; Gallinger, S.; Swallow, C.; Redston, M.; Couture, J. Molecular biology of colorectal cancer. Curr. Probl. Cancer 1997, 21, 233–299. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.B.; Smith, K.J.; Beazer-Barclay, Y.; Hamilton, S.R.; Vogelstein, B.; Kinzler, K.W. Inactivation of both APC alleles in human and mouse tumors. Cancer Res. 1994, 54, 5953–5958. [Google Scholar] [PubMed]
- Dow, L.E.; O’Rourke, K.P.; Simon, J.; Tschaharganeh, D.F.; van Es, J.H.; Clevers, H.; Lowe, S.W. Apc restoration promotes cellular differentiation and reestablishes crypt homeostasis in colorectal cancer. Cell 2015, 161, 1539–1552. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Network. Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330. [Google Scholar] [CrossRef]
- Fischer, M.M.; Yeung, V.P.; Cattaruzza, F.; Hussein, R.; Yen, W.-C.; Murriel, C.; Evans, J.W.; O’Young, G.; Brunner, A.L.; Wang, M. RSPO3 antagonism inhibits growth and tumorigenicity in colorectal tumors harboring common Wnt pathway mutations. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; He, X.; Jia, M.; Liu, Y.; Qu, D.; Wu, D.; Wu, P.; Ni, C.; Zhang, Z.; Ye, J. β-catenin overexpression in the nucleus predicts progress disease and unfavourable survival in colorectal cancer: A meta-analysis. PLoS ONE 2013, 8, e63854. [Google Scholar] [CrossRef]
- Grady, W.M.; Markowitz, S.D. Genetic and epigenetic alterations in colon cancer. Annu. Rev. Genom. Hum. Genet. 2002, 3, 101–128. [Google Scholar] [CrossRef]
- Armaghany, T.; Wilson, J.D.; Chu, Q.; Mills, G. Genetic alterations in colorectal cancer. Gastrointest. Cancer Res. 2012, 5, 19. [Google Scholar]
- Lynch, H.T.; Boland, C.R.; Gong, G.; Shaw, T.G.; Lynch, P.M.; Fodde, R.; Lynch, J.F.; de la Chapelle, A. Phenotypic and genotypic heterogeneity in the Lynch syndrome: Diagnostic, surveillance and management implications. Eur. J. Hum. Genet. 2006, 14, 390–402. [Google Scholar] [CrossRef]
- Toyota, M.; Ahuja, N.; Ohe-Toyota, M.; Herman, J.G.; Baylin, S.B.; Issa, J.-P.J. CpG island methylator phenotype in colorectal cancer. Proc. Natl. Acad. Sci. USA 1999, 96, 8681–8686. [Google Scholar] [CrossRef]
- Weisenberger, D.J.; Siegmund, K.D.; Campan, M.; Young, J.; Long, T.I.; Faasse, M.A.; Kang, G.H.; Widschwendter, M.; Weener, D.; Buchanan, D. CpG island methylator phenotype underlies sporadic microsatellite instability and is tightly associated with BRAF mutation in colorectal cancer. Nat. Genet. 2006, 38, 787–793. [Google Scholar] [CrossRef]
- Advani, S.M.; Advani, P.; DeSantis, S.M.; Brown, D.; VonVille, H.M.; Lam, M.; Loree, J.M.; Sarshekeh, A.M.; Bressler, J.; Lopez, D.S. Clinical, pathological, and molecular characteristics of CpG island methylator phenotype in colorectal cancer: A systematic review and meta-analysis. Transl. Oncol. 2018, 11, 1188–1201. [Google Scholar] [CrossRef]
- Bosetti, C.; Santucci, C.; Gallus, S.; Martinetti, M.; La Vecchia, C. Aspirin and the risk of colorectal and other digestive tract cancers: An updated meta-analysis through 2019. Ann. Oncol. 2020, 31, 558–568. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Nishihara, R.; Qian, Z.R.; Song, M.; Mima, K.; Inamura, K.; Nowak, J.A.; Drew, D.A.; Lochhead, P.; Nosho, K. Regular aspirin use associates with lower risk of colorectal cancers with low numbers of tumor-infiltrating lymphocytes. Gastroenterology 2016, 151, 879–892.e874. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.T.; Ogino, S.; Fuchs, C.S. Aspirin and the risk of colorectal cancer in relation to the expression of COX-2. N. Engl. J. Med. 2007, 356, 2131–2142. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, P.M.; Wilson, M.; Elwin, C.-E.; Norrving, B.; Algra, A.; Warlow, C.P.; Meade, T.W. Long-term effect of aspirin on colorectal cancer incidence and mortality: 20-year follow-up of five randomised trials. Lancet 2010, 376, 1741–1750. [Google Scholar] [CrossRef] [PubMed]
- Tomić, T.; Domínguez-López, S.; Barrios-Rodríguez, R. Non-aspirin non-steroidal anti-inflammatory drugs in prevention of colorectal cancer in people aged 40 or older: A systematic review and meta-analysis. Cancer Epidemiol. 2019, 58, 52–62. [Google Scholar] [CrossRef]
- Meyskens, F.L.; McLaren, C.E.; Pelot, D.; Fujikawa-Brooks, S.; Carpenter, P.M.; Hawk, E.; Kelloff, G.; Lawson, M.J.; Kidao, J.; McCracken, J. Difluoromethylornithine plus sulindac for the prevention of sporadic colorectal adenomas: A randomized placebo-controlled, double-blind trial. Cancer Prev. Res. 2008, 1, 32–38. [Google Scholar] [CrossRef]
- Thomasset, S.; Berry, D.P.; Cai, H.; West, K.; Marczylo, T.H.; Marsden, D.; Brown, K.; Dennison, A.; Garcea, G.; Miller, A. Pilot study of oral anthocyanins for colorectal cancer chemoprevention. Cancer Prev. Res. 2009, 2, 625–633. [Google Scholar] [CrossRef]
- Sinicrope, F.A.; Velamala, P.R.; Song, L.M.W.K.; Viggiano, T.R.; Bruining, D.H.; Rajan, E.; Gostout, C.J.; Kraichely, R.E.; Buttar, N.S.; Schroeder, K.W. Efficacy of difluoromethylornithine and aspirin for treatment of adenomas and aberrant crypt foci in patients with prior advanced colorectal neoplasms. Cancer Prev. Res. 2019, 12, 821–830. [Google Scholar] [CrossRef]
- Samadder, N.J.; Kuwada, S.K.; Boucher, K.M.; Byrne, K.; Kanth, P.; Samowitz, W.; Jones, D.; Tavtigian, S.V.; Westover, M.; Berry, T. Association of sulindac and erlotinib vs placebo with colorectal neoplasia in familial adenomatous polyposis: Secondary analysis of a randomized clinical trial. JAMA Oncol. 2018, 4, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Veettil, S.K.; Nathisuwan, S.; Ching, S.M.; Jinatongthai, P.; Lim, K.G.; Kew, S.T.; Chaiyakunapruk, N. Efficacy and safety of celecoxib on the incidence of recurrent colorectal adenomas: A systematic review and meta-analysis. Cancer Manag. Res. 2019, 11, 561. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Miguel, A.; García-Rodríguez, L.A.; Gil, M.; Montoya, H.; Rodríguez-Martín, S.; de Abajo, F.J. Clopidogrel and low-dose aspirin, alone or together, reduce risk of colorectal cancer. Clin. Gastroenterol. Hepatol. 2019, 17, 2024–2033.e2022. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.-A.W.; Jiang, A.A.; Toh, E.M.S.; Ng, C.H.; Ong, Z.H.; Peng, S.; Tham, H.Y.; Sundar, R.; Chong, C.S.; Khoo, C.M. Metformin and colorectal cancer: A systematic review, meta-analysis and meta-regression. Int. J. Color. Dis. 2020, 35, 1501–1512. [Google Scholar] [CrossRef]
- Huang, W.-K.; Hsu, H.-C.; Liu, J.-R.; Yang, T.-S.; Chen, J.-S.; Chang, J.W.-C.; Lin, Y.-C.; Yu, K.-H.; Kuo, C.-F.; See, L.-C. The association of ursodeoxycholic acid use with colorectal cancer risk: A nationwide cohort study. Medicine 2016, 95, e2980. [Google Scholar] [CrossRef]
- Li, Y.; He, X.; Ding, Y.e.; Chen, H.; Sun, L. Statin uses and mortality in colorectal cancer patients: An updated systematic review and meta-analysis. Cancer Med. 2019, 8, 3305–3313. [Google Scholar] [CrossRef] [Green Version]
- Barbalata, C.I.; Tefas, L.R.; Achim, M.; Tomuta, I.; Porfire, A.S. Statins in risk-reduction and treatment of cancer. World J. Clin. Oncol. 2020, 11, 573. [Google Scholar] [CrossRef]
- Botteri, E.; Støer, N.C.; Sakshaug, S.; Graff-Iversen, S.; Vangen, S.; Hofvind, S.; De Lange, T.; Bagnardi, V.; Ursin, G.; Weiderpass, E. Menopausal hormone therapy and colorectal cancer: A linkage between nationwide registries in Norway. BMJ Open 2017, 7, e017639. [Google Scholar] [CrossRef]
- Li, Y.-Y.; Gao, L.-J.; Zhang, Y.-X.; Liu, S.-J.; Cheng, S.; Liu, Y.-P.; Jia, C.-X. Bisphosphonates and risk of cancers: A systematic review and meta-analysis. Br. J. Cancer 2020, 123, 1570–1581. [Google Scholar] [CrossRef]
- Eberhart, C.E.; Coffey, R.J.; Radhika, A.; Giardiello, F.M.; Ferrenbach, S.; Dubois, R.N. Up-regulation of cyclooxygenase 2 gene expression in human colorectal adenomas and adenocarcinomas. Gastroenterology 1994, 107, 1183–1188. [Google Scholar] [CrossRef]
- Oshima, M.; Dinchuk, J.E.; Kargman, S.L.; Oshima, H.; Hancock, B.; Kwong, E.; Trzaskos, J.M.; Evans, J.F.; Taketo, M.M. Suppression of intestinal polyposis in ApcΔ716 knockout mice by inhibition of cyclooxygenase 2 (COX-2). Cell 1996, 87, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Castellone, M.D.; Teramoto, H.; Williams, B.O.; Druey, K.M.; Gutkind, J.S. Prostaglandin E2 promotes colon cancer cell growth through a Gs-axin-ß-catenin signaling axis. Science 2005, 310, 1504–1510. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; DuBois, R.N. The role of COX-2 in intestinal inflammation and colorectal cancer. Oncogene 2010, 29, 781–788. [Google Scholar] [CrossRef]
- Masferrer, J.L.; Leahy, K.M.; Koki, A.T.; Zweifel, B.S.; Settle, S.L.; Woerner, B.M.; Edwards, D.A.; Flickinger, A.G.; Moore, R.J.; Seibert, K. Antiangiogenic and antitumor activities of cyclooxygenase-2 inhibitors. Cancer Res. 2000, 60, 1306–1311. [Google Scholar] [PubMed]
- Seno, H.; Oshima, M.; Ishikawa, T.-O.; Oshima, H.; Takaku, K.; Chiba, T.; Narumiya, S.; Taketo, M.M. Cyclooxygenase 2-and prostaglandin E2 receptor EP2-dependent angiogenesis in ApcΔ716 mouse intestinal polyps. Cancer Res. 2002, 62, 506–511. [Google Scholar] [PubMed]
- Nan, H.; Hutter, C.M.; Lin, Y.; Jacobs, E.J.; Ulrich, C.M.; White, E.; Baron, J.A.; Berndt, S.I.; Brenner, H.; Butterbach, K. Association of aspirin and NSAID use with risk of colorectal cancer according to genetic variants. JAMA 2015, 313, 1133–1142. [Google Scholar] [CrossRef] [Green Version]
- Arber, N.; Eagle, C.J.; Spicak, J.; Rácz, I.; Dite, P.; Hajer, J.; Zavoral, M.; Lechuga, M.J.; Gerletti, P.; Tang, J. Celecoxib for the prevention of colorectal adenomatous polyps. N. Engl. J. Med. 2006, 355, 885–895. [Google Scholar] [CrossRef]
- Dehmer, S.P.; Maciosek, M.V.; Flottemesch, T.J.; LaFrance, A.B.; Whitlock, E.P. Aspirin for the primary prevention of cardiovascular disease and colorectal cancer: A decision analysis for the US Preventive Services Task Force. Ann. Intern. Med. 2016, 164, 777–786. [Google Scholar] [CrossRef]
- Chubak, J.; Whitlock, E.P.; Williams, S.B.; Kamineni, A.; Burda, B.U.; Buist, D.S.; Anderson, M.L. Aspirin for the prevention of cancer incidence and mortality: Systematic evidence reviews for the US Preventive Services Task Force. Ann. Intern. Med. 2016, 164, 814–825. [Google Scholar] [CrossRef]
- Katona, B.W.; Weiss, J.M. Chemoprevention of colorectal cancer. Gastroenterology 2020, 158, 368–388. [Google Scholar] [CrossRef]
- Liu, F.; Yan, L.; Wang, Z.; Lu, Y.; Chu, Y.; Li, X.; Liu, Y.; Rui, D.; Nie, S.; Xiang, H. Metformin therapy and risk of colorectal adenomas and colorectal cancer in type 2 diabetes mellitus patients: A systematic review and meta-analysis. Oncotarget 2017, 8, 16017. [Google Scholar] [CrossRef] [PubMed]
- Petrera, M.; Paleari, L.; Clavarezza, M.; Puntoni, M.; Caviglia, S.; Briata, I.M.; Oppezzi, M.; Mislej, E.M.; Stabuc, B.; Gnant, M. The ASAMET trial: A randomized, phase II, double-blind, placebo-controlled, multicenter, 2× 2 factorial biomarker study of tertiary prevention with low-dose aspirin and metformin in stage I-III colorectal cancer patients. BMC Cancer 2018, 18, 1–9. [Google Scholar] [CrossRef]
- Chan, K.K.; Oza, A.M.; Siu, L.L. The statins as anticancer agents. Clin. Cancer Res. 2003, 9, 10–19. [Google Scholar] [PubMed]
- Teraoka, N.; Mutoh, M.; Takasu, S.; Ueno, T.; Yamamoto, M.; Sugimura, T.; Wakabayashi, K. Inhibition of intestinal polyp formation by pitavastatin, a HMG-CoA reductase inhibitor. Cancer Prev. Res. 2011, 4, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Suh, N.; Reddy, B.S.; DeCastro, A.; Paul, S.; Lee, H.J.; Smolarek, A.K.; So, J.Y.; Simi, B.; Wang, C.X.; Janakiram, N.B. Combination of atorvastatin with sulindac or naproxen profoundly inhibits colonic adenocarcinomas by suppressing the p65/β-catenin/cyclin D1 signaling pathway in rats. Cancer Prev. Res. 2011, 4, 1895–1902. [Google Scholar] [CrossRef]
- Malila, N.; Virtamo, J.; Virtanen, M.; Albanes, D.; Tangrea, J.A.; Huttunen, J.K. The effect of α-tocopherol and β-carotene supplementation on colorectal adenomas in middle-aged male smokers. Cancer Epidemiol. Prev. Biomark. 1999, 8, 489–493. [Google Scholar]
- Papaioannou, D.; Cooper, K.; Carroll, C.; Hind, D.; Squires, H.; Tappenden, P.; Logan, R. Antioxidants in the chemoprevention of colorectal cancer and colorectal adenomas in the general population: A systematic review and meta-analysis. Color. Dis. 2011, 13, 1085–1099. [Google Scholar] [CrossRef]
- Yusof, A.S.; Isa, Z.M.; Shah, S.A. Dietary patterns and risk of colorectal cancer: A systematic review of cohort studies (2000–2011). Asian Pac. J. Cancer Prev. 2012, 13, 4713–4717. [Google Scholar] [CrossRef]
- Vanio, H.; Bianchini, F. Weight Control and Physical Activity. In IARC Handbooks of Cancer Prevention; IARC Press: Lyon, France, 2002. [Google Scholar]
- He, X.; Wu, K.; Zhang, X.; Nishihara, R.; Cao, Y.; Fuchs, C.S.; Giovannucci, E.L.; Ogino, S.; Chan, A.T.; Song, M. Dietary intake of fiber, whole grains and risk of colorectal cancer: An updated analysis according to food sources, tumor location and molecular subtypes in two large US cohorts. Int. J. Cancer 2019, 145, 3040–3051. [Google Scholar] [CrossRef]
- Brenner, H.; Chang–Claude, J.; Jansen, L.; Knebel, P.; Stock, C.; Hoffmeister, M. Reduced risk of colorectal cancer up to 10 years after screening, surveillance, or diagnostic colonoscopy. Gastroenterology 2014, 146, 709–717. [Google Scholar] [CrossRef]
- Wells, K.O.; Hawkins, A.T.; Krishnamurthy, D.M.; Dharmarajan, S.; Glasgow, S.C.; Hunt, S.R.; Mutch, M.G.; Wise, P.; Silviera, M.L. Omission of adjuvant chemotherapy is associated with increased mortality in patients with T3N0 colon cancer with inadequate lymph node harvest. Dis. Colon Rectum 2017, 60, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Ribic, C.M.; Sargent, D.J.; Moore, M.J.; Thibodeau, S.N.; French, A.J.; Goldberg, R.M.; Hamilton, S.R.; Laurent-Puig, P.; Gryfe, R.; Shepherd, L.E. Tumor microsatellite-instability status as a predictor of benefit from fluorouracil-based adjuvant chemotherapy for colon cancer. N. Engl. J. Med. 2003, 349, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Sargent, D.J.; Marsoni, S.; Monges, G.; Thibodeau, S.N.; Labianca, R.; Hamilton, S.R.; French, A.J.; Kabat, B.; Foster, N.R.; Torri, V. Defective mismatch repair as a predictive marker for lack of efficacy of fluorouracil-based adjuvant therapy in colon cancer. J. Clin. Oncol. 2010, 28, 3219. [Google Scholar] [CrossRef]
- Argilés, G.; Tabernero, J.; Labianca, R.; Hochhauser, D.; Salazar, R.; Iveson, T.; Laurent-Puig, P.; Quirke, P.; Yoshino, T.; Taieb, J. Localised colon cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2020, 31, 1291–1305. [Google Scholar] [CrossRef] [PubMed]
- Angell, H.K.; Bruni, D.; Barrett, J.C.; Herbst, R.; Galon, J. The immunoscore: Colon cancer and beyond. Clin. Cancer Res. 2020, 26, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.-H.; Chen, Y.-X.; Fang, J.-Y. Comprehensive review of targeted therapy for colorectal cancer. Signal Transduct. Target. Ther. 2020, 5, 1–30. [Google Scholar]
- Falcone, A.; Ricci, S.; Brunetti, I.; Pfanner, E.; Allegrini, G.; Barbara, C.; Crinò, L.; Benedetti, G.; Evangelista, W.; Fanchini, L. Phase III trial of infusional fluorouracil, leucovorin, oxaliplatin, and irinotecan (FOLFOXIRI) compared with infusional fluorouracil, leucovorin, and irinotecan (FOLFIRI) as first-line treatment for metastatic colorectal cancer: The Gruppo Oncologico Nord Ovest. J. Clin. Oncol. 2007, 25, 1670–1676. [Google Scholar]
- Souglakos, J.; Androulakis, N.; Syrigos, K.; Polyzos, A.; Ziras, N.; Athanasiadis, A.; Kakolyris, S.; Tsousis, S.; Kouroussis, C.; Vamvakas, L. FOLFOXIRI (folinic acid, 5-fluorouracil, oxaliplatin and irinotecan) vs FOLFIRI (folinic acid, 5-fluorouracil and irinotecan) as first-line treatment in metastatic colorectal cancer (MCC): A multicentre randomised phase III trial from the Hellenic Oncology Research Group (HORG). Br. J. Cancer 2006, 94, 798–805. [Google Scholar]
- Cunningham, D.; Humblet, Y.; Siena, S.; Khayat, D.; Bleiberg, H.; Santoro, A.; Bets, D.; Mueser, M.; Harstrick, A.; Verslype, C. Cetuximab monotherapy and cetuximab plus irinotecan in irinotecan-refractory metastatic colorectal cancer. N. Engl. J. Med. 2004, 351, 337–345. [Google Scholar] [CrossRef]
- Mendelsohn, J.; Prewett, M.; Rockwell, P.; Goldstein, N.I. CCR 20th anniversary commentary: A chimeric antibody, C225, inhibits EGFR activation and tumor growth. Clin. Cancer Res. 2015, 21, 227–229. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, A.F.; Bretes, L.; Furtado, I. Review of PD-1/PD-L1 inhibitors in metastatic dMMR/MSI-H colorectal cancer. Front. Oncol. 2019, 9, 396. [Google Scholar] [CrossRef] [PubMed]
- Jonker, D.J.; O’Callaghan, C.J.; Karapetis, C.S.; Zalcberg, J.R.; Tu, D.; Au, H.-J.; Berry, S.R.; Krahn, M.; Price, T.; Simes, R.J. Cetuximab for the treatment of colorectal cancer. N. Engl. J. Med. 2007, 357, 2040–2048. [Google Scholar] [CrossRef] [PubMed]
- Roskoski, R., Jr. The ErbB/HER family of protein-tyrosine kinases and cancer. Pharmacol. Res. 2014, 79, 34–74. [Google Scholar] [CrossRef]
- Shibuya, M. Vascular endothelial growth factor (VEGF) and its receptor (VEGFR) signaling in angiogenesis: A crucial target for anti-and pro-angiogenic therapies. Genes Cancer 2011, 2, 1097–1105. [Google Scholar] [CrossRef]
- Guba, M.; Seeliger, H.; Kleespies, A.; Jauch, K.-W.; Bruns, C. Vascular endothelial growth factor in colorectal cancer. Int. J. Color. Dis. 2004, 19, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Syst. Rev. 2021, 88, 105906. [Google Scholar]
- De Paulo Farias, D.; de Araujo, F.F.; Neri-Numa, I.A.; Pastore, G.M. Antidiabetic potential of dietary polyphenols: A mechanistic review. Food Res. Int. 2021, 145, 110383. [Google Scholar] [CrossRef]
- Tsimogiannis, D.; Oreopoulou, V. Classification of phenolic compounds in plants. Polyphen. Plants (Second Ed.) 2019, 263–284. [Google Scholar] [CrossRef]
- Vuolo, M.M.; Lima, V.S.; Junior, M.R.M. Phenolic compounds: Structure, classification, and antioxidant power. Bioact. Compd. 2019, 33–50. [Google Scholar] [CrossRef]
- Kiruthiga, C.; Devi, K.P.; Nabavi, S.M.; Bishayee, A. Autophagy: A potential therapeutic target of polyphenols in hepatocellular carcinoma. Cancers 2020, 12, 562. [Google Scholar] [CrossRef]
- Darvesh, A.S.; Carroll, R.T.; Bishayee, A.; Geldenhuys, W.J.; Van der Schyf, C.J. Oxidative stress and Alzheimer’s disease: Dietary polyphenols as potential therapeutic agents. Expert Rev. Neurother. 2010, 10, 729–745. [Google Scholar] [CrossRef]
- Karimi, A.; Majlesi, M.; Rafieian-Kopaei, M. Herbal versus synthetic drugs; beliefs and facts. J. Nephropharmacol. 2015, 4, 27. [Google Scholar] [PubMed]
- Samec, M.; Liskova, A.; Koklesova, L.; Samuel, S.M.; Zhai, K.; Buhrmann, C.; Varghese, E.; Abotaleb, M.; Qaradakhi, T.; Zulli, A. Flavonoids against the Warburg phenotype—Concepts of predictive, preventive and personalised medicine to cut the Gordian knot of cancer cell metabolism. Epma J. 2020, 11, 377–398. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Madu, C.O.; Lu, Y. Phytochemicals in chemoprevention: A cost-effective complementary approach. J. Cancer 2021, 12, 3686. [Google Scholar] [CrossRef] [PubMed]
- Rajamanickam, S.; Agarwal, R. Natural products and colon cancer: Current status and future prospects. Drug Dev. Res. 2008, 69, 460–471. [Google Scholar] [CrossRef]
- Bishayee, A.; Sethi, G. Bioactive natural products in cancer prevention and therapy: Progress and promise. Semin. Cancer Biol. 2016, 40, 1–3. [Google Scholar] [CrossRef]
- Huang, X.-M.; Yang, Z.-J.; Xie, Q.; Zhang, Z.-K.; Zhang, H.; Ma, J.-Y. Natural products for treating colorectal cancer: A mechanistic review. Biomed. Pharmacother. 2019, 117, 109142. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Lin, C.-L.; Jeng, J.-H.; Wu, C.-C.; Hsieh, S.-L.; Huang, G.-C.; Leung, W.; Lee, C.-T.; Chen, C.-Y.; Lee, C.-H. Chemopreventive potential of 2, 3, 5, 4′-tetrahydroxystilbene-2-O-β-D-glucoside on The formation of aberrant crypt foci in azoxymethane-induced colorectal cancer in rats. BioMed Res. Int. 2017, 2017, 3634915. [Google Scholar] [CrossRef]
- Chen, Q.; Lei, J.; Zhou, J.; Ma, S.; Huang, Q.; Ge, B. Chemopreventive effect of 4′-hydroxychalcone on intestinal tumorigenesis in Apc Min mice Corrigendum in/10.3892/ol. 2021.12741. Oncol. Lett. 2021, 21, 213. [Google Scholar] [CrossRef]
- Lai, C.-Y.; Tsai, A.-C.; Chen, M.-C.; Chang, L.-H.; Sun, H.-L.; Chang, Y.-L.; Chen, C.-C.; Teng, C.-M.; Pan, S.-L. Aciculatin induces p53-dependent apoptosis via MDM2 depletion in human cancer cells in vitro and in vivo. PLoS ONE 2012, 7, e42192. [Google Scholar] [CrossRef] [Green Version]
- Au, A.; Li, B.; Wang, W.; Roy, H.; Koehler, K.; Birt, D. Effect of dietary apigenin on colonic ornithine decarboxylase activity, aberrant crypt foci formation, and tumorigenesis in different experimental models. Nutr. Cancer 2006, 54, 243–251. [Google Scholar] [CrossRef]
- Wang, Q.R.; Yao, X.Q.; Wen, G.; Fan, Q.; Li, Y.-J.; Fu, X.Q.; Li, C.K.; Sun, X.G. Apigenin suppresses the growth of colorectal cancer xenografts via phosphorylation and up-regulated FADD expression. Oncol. Lett. 2011, 2, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Chunhua, L.; Donglan, L.; Xiuqiong, F.; Lihua, Z.; Qin, F.; Yawei, L.; Liang, Z.; Ge, W.; Linlin, J.; Ping, Z. Apigenin up-regulates transgelin and inhibits invasion and migration of colorectal cancer through decreased phosphorylation of AKT. J. Nutr. Biochem. 2013, 24, 1766–1775. [Google Scholar] [CrossRef]
- Zhong, Y.; Krisanapun, C.; Lee, S.-H.; Nualsanit, T.; Sams, C.; Peungvicha, P.; Baek, S.J. Molecular targets of apigenin in colorectal cancer cells: Involvement of p21, NAG-1 and p53. Eur. J. Cancer 2010, 46, 3365–3374. [Google Scholar] [CrossRef]
- Chen, X.; Xu, H.; Yu, X.; Wang, X.; Zhu, X.; Xu, X. Apigenin inhibits in vitro and in vivo tumorigenesis in cisplatin-resistant colon cancer cells by inducing autophagy, programmed cell death and targeting m-TOR/PI3K/Akt signalling pathway. J. Buon 2019, 24, 488–493. [Google Scholar] [PubMed]
- Shao, H.; Jing, K.; Mahmoud, E.; Huang, H.; Fang, X.; Yu, C. Apigenin sensitizes colon cancer cells to antitumor activity of ABT-263. Mol. Cancer Ther. 2013, 12, 2640–2650. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Van Wie, P.G.; Fai, L.Y.; Kim, D.; Wang, L.; Poyil, P.; Luo, J.; Zhang, Z. Downregulation of NEDD9 by apigenin suppresses migration, invasion, and metastasis of colorectal cancer cells. Toxicol. Appl. Pharmacol. 2016, 311, 106–112. [Google Scholar] [CrossRef]
- Kim, D.H.; Hossain, M.A.; Kang, Y.J.; Jang, J.Y.; Lee, Y.J.; Im, E.; Yoon, J.-H.; Kim, H.S.; Chung, H.Y.; Kim, N.D. Baicalein, an active component of Scutellaria baicalensis Georgi, induces apoptosis in human colon cancer cells and prevents AOM/DSS-induced colon cancer in mice. Int. J. Oncol. 2013, 43, 1652–1658. [Google Scholar] [CrossRef]
- Chen, W.-C.; Kuo, T.-H.; Tzeng, Y.-S.; Tsai, Y.-C. Baicalin induces apoptosis in SW620 human colorectal carcinoma cells in vitro and suppresses tumor growth in vivo. Molecules 2012, 17, 3844–3857. [Google Scholar] [CrossRef]
- Song, L.; Zhu, S.; Liu, C.; Zhang, Q.; Liang, X. Baicalin triggers apoptosis, inhibits migration, and enhances anti-tumor immunity in colorectal cancer via TLR4/NF-κB signaling pathway. J. Food Biochem. 2022, 46, e13703. [Google Scholar] [CrossRef]
- Kim, S.-J.; Kim, H.-J.; Kim, H.-R.; Lee, S.-H.; Cho, S.-D.; Choi, C.-S.; Nam, J.-S.; Jung, J.-Y. Antitumor actions of baicalein and wogonin in HT-29 human colorectal cancer cells. Mol. Med. Rep. 2012, 6, 1443–1449. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.; Xu, J.; Yan, B. The anti-metastatic effect of baicalein on colorectal cancer. Oncol. Rep. 2017, 37, 2317–2323. [Google Scholar] [CrossRef] [PubMed]
- Dou, J.; Wang, Z.; Ma, L.; Peng, B.; Mao, K.; Li, C.; Su, M.; Zhou, C.; Peng, G. Baicalein and baicalin inhibit colon cancer using two distinct fashions of apoptosis and senescence. Oncotarget 2018, 9, 20089. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ma, L.; Su, M.; Zhou, Y.; Mao, K.; Li, C.; Peng, G.; Zhou, C.; Shen, B.; Dou, J. Baicalin induces cellular senescence in human colon cancer cells via upregulation of DEPP and the activation of Ras/Raf/MEK/ERK signaling. Cell Death Dis. 2018, 9, 217. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Zhan, S.; Wang, Y.; Zhou, G.; Liang, H.; Chen, X.; Shen, H. Baicalin, the major component of traditional Chinese medicine Scutellaria baicalensis induces colon cancer cell apoptosis through inhibition of oncomiRNAs. Sci. Rep. 2018, 8, 14477. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Bai, H.; Sa, Y.; Zhu, P.; Liu, P. Inhibiting EMT, stemness and cell cycle involved in baicalin-induced growth inhibition and apoptosis in colorectal cancer cells. J. Cancer 2020, 11, 2303. [Google Scholar] [CrossRef]
- Wang, C.-Z.; Zhang, C.-F.; Luo, Y.; Yao, H.; Yu, C.; Chen, L.; Yuan, J.; Huang, W.-H.; Wan, J.-Y.; Zeng, J. Baicalein, an enteric microbial metabolite, suppresses gut inflammation and cancer progression in Apc Min/+ mice. Clin. Transl. Oncol. 2020, 22, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, Q.; Luo, L.; Song, J.; Han, K.; Liu, R.; Gong, Y.; Guo, X. Use Chou’s 5-steps rule to study how Baicalin suppresses the malignant phenotypes and induces the apoptosis of colorectal cancer cells. Arch. Biochem. Biophys. 2021, 705, 108919. [Google Scholar] [CrossRef]
- Zhou, C.; Ou, W.; Xu, Q.; Lin, L.; Xu, F.; Chen, R.; Miao, H. Chemoprotective effect of boeravinone B against 1, 2-dimethyl hydrazine induced colorectal cancer in rats via suppression of oxidative stress and inflammatory reaction. J. Gastrointest. Oncol. 2022, 13, 1832–1841. [Google Scholar] [CrossRef]
- Miyamoto, S.; Kohno, H.; Suzuki, R.; Sugie, S.; Murakami, A.; Ohigashi, H.; Tanaka, T. Preventive effects of chrysin on the development of azoxymethane-induced colonic aberrant crypt foci in rats. Oncol. Rep. 2006, 15, 1169–1173. [Google Scholar] [CrossRef] [Green Version]
- Salama, A.A.; Allam, R.M. Promising targets of chrysin and daidzein in colorectal cancer: Amphiregulin, CXCL1, and MMP-9. Eur. J. Pharmacol. 2021, 892, 173763. [Google Scholar] [CrossRef] [PubMed]
- Villegas, I.; Sánchez-Fidalgo, S.; de la Lastra, C.A. Chemopreventive effect of dietary curcumin on inflammation-induced colorectal carcinogenesis in mice. Mol. Nutr. Food Res. 2011, 55, 259–267. [Google Scholar] [CrossRef]
- Milacic, V.; Banerjee, S.; Landis-Piwowar, K.R.; Sarkar, F.H.; Majumdar, A.P.; Dou, Q.P. Curcumin inhibits the proteasome activity in human colon cancer cells in vitro and in vivo. Cancer Res. 2008, 68, 7283–7292. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Dai, X.; Gao, J.; Li, Y.; Hou, Z.; Chang, Z.; Wang, Y. Curcumin suppresses colorectal tumorigenesis via the Wnt/β-catenin signaling pathway by downregulating Axin2. Oncol. Lett. 2021, 21, 186. [Google Scholar] [CrossRef]
- Kubota, M.; Shimizu, M.; Sakai, H.; Yasuda, Y.; Terakura, D.; Baba, A.; Ohno, T.; Tsurumi, H.; Tanaka, T.; Moriwaki, H. Preventive effects of curcumin on the development of azoxymethane-induced colonic preneoplastic lesions in male C57BL/KsJ-db/db obese mice. Nutr. Cancer 2012, 64, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Kunnumakkara, A.B.; Diagaradjane, P.; Guha, S.; Deorukhkar, A.; Shentu, S.; Aggarwal, B.B.; Krishnan, S. Curcumin sensitizes human colorectal cancer xenografts in nude mice to γ-radiation by targeting nuclear factor-κB–regulated gene products. Clin. Cancer Res. 2008, 14, 2128–2136. [Google Scholar] [CrossRef]
- Li, L.; Ahmed, B.; Mehta, K.; Kurzrock, R. Liposomal curcumin with and without oxaliplatin: Effects on cell growth, apoptosis, and angiogenesis in colorectal cancer. Mol. Cancer Ther. 2007, 6, 1276–1282. [Google Scholar] [CrossRef]
- McFadden, R.-M.T.; Larmonier, C.B.; Shehab, K.W.; Midura-Kiela, M.; Ramalingam, R.; Harrison, C.A.; Besselsen, D.G.; Chase, J.H.; Caporaso, J.G.; Jobin, C. The role of curcumin in modulating colonic microbiota during colitis and colon cancer prevention. Inflamm. Bowel Dis. 2015, 21, 2483–2494. [Google Scholar] [CrossRef]
- Marjaneh, R.M.; Rahmani, F.; Hassanian, S.M.; Rezaei, N.; Hashemzehi, M.; Bahrami, A.; Ariakia, F.; Fiuji, H.; Sahebkar, A.; Avan, A. Phytosomal curcumin inhibits tumor growth in colitis-associated colorectal cancer. J. Cell. Physiol. 2018, 233, 6785–6798. [Google Scholar] [CrossRef]
- Cooke, D.; Schwarz, M.; Boocock, D.; Winterhalter, P.; Steward, W.P.; Gescher, A.J.; Marczylo, T.H. Effect of cyanidin-3-glucoside and an anthocyanin mixture from bilberry on adenoma development in the ApcMin mouse model of intestinal carcinogenesis—Relationship with tissue anthocyanin levels. Int. J. Cancer 2006, 119, 2213–2220. [Google Scholar] [CrossRef]
- Huang, C.-C.; Hung, C.-H.; Hung, T.-W.; Lin, Y.-C.; Wang, C.-J.; Kao, S.-H. Dietary delphinidin inhibits human colorectal cancer metastasis associating with upregulation of miR-204-3p and suppression of the integrin/FAK axis. Sci. Rep. 2019, 9, 18954. [Google Scholar] [CrossRef] [PubMed]
- Koosha, S.; Mohamed, Z.; Sinniah, A.; Alshawsh, M.A. Evaluation of anti-tumorigenic effects of diosmetin against human colon cancer xenografts in athymic nude mice. Molecules 2019, 24, 2522. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Shirakami, Y.; Sakai, H.; Yasuda, Y.; Kubota, M.; Adachi, S.; Tsurumi, H.; Hara, Y.; Moriwaki, H. (−)-Epigallocatechin gallate inhibits growth and activation of the VEGF/VEGFR axis in human colorectal cancer cells. Chem.-Biol. Interact. 2010, 185, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Shirakami, Y.; Sakai, H.; Adachi, S.; Hata, K.; Hirose, Y.; Tsurumi, H.; Tanaka, T.; Moriwaki, H. (−)-Epigallocatechin gallate suppresses azoxymethane-induced colonic premalignant lesions in male C57BL/KsJ-db/db mice. Cancer Prev. Res. 2008, 1, 298–304. [Google Scholar] [CrossRef]
- Zhong, Y.; Chiou, Y.-S.; Pan, M.-H.; Ho, C.-T.; Shahidi, F. Protective effects of epigallocatechin gallate (EGCG) derivatives on azoxymethane-induced colonic carcinogenesis in mice. J. Funct. Foods 2012, 4, 323–330. [Google Scholar] [CrossRef]
- Toden, S.; Tran, H.-M.; Tovar-Camargo, O.A.; Okugawa, Y.; Goel, A. Epigallocatechin-3-gallate targets cancer stem-like cells and enhances 5-fluorouracil chemosensitivity in colorectal cancer. Oncotarget 2016, 7, 16158. [Google Scholar] [CrossRef]
- Xu, G.; Ren, G.; Xu, X.; Yuan, H.; Wang, Z.; Kang, L.; Yu, W.; Tian, K. Combination of curcumin and green tea catechins prevents dimethylhydrazine-induced colon carcinogenesis. Food Chem. Toxicol. 2010, 48, 390–395. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, H.-Y.; Fang, M.-Z.; Wang, X.-F.; Chen, H.; Huang, S.-L.; Kong, D.-S.; Li, M.; Zhang, X.; Sun, Y. Epigallocatechin gallate inhibits dimethylhydrazine-induced colorectal cancer in rats. World J. Gastroenterol. 2020, 26, 2064. [Google Scholar] [CrossRef]
- Mariyappan, P.; Kalaiyarasu, T.; Manju, V. Effect of eriodictyol on preneoplastic lesions, oxidative stress and bacterial enzymes in 1, 2-dimethyl hydrazine-induced colon carcinogenesis. Toxicol. Res. 2017, 6, 678–692. [Google Scholar] [CrossRef]
- Wang, N.; Zhou, F.; Guo, J.; Zhu, H.; Luo, S.; Cao, J. Euxanthone suppresses tumor growth and metastasis in colorectal cancer via targeting CIP2A/PP2A pathway. Life Sci. 2018, 209, 498–506. [Google Scholar] [CrossRef]
- Kunchari Kalaimathi, S.; Sudhandiran, G. Fisetin ameolirates the azoxymethane and dextran sodium sulfate induced colitis associated colorectal cancer. Int. J. Pharm. Clin. Res. 2016, 8, 551–560. [Google Scholar]
- Khan, N.; Jajeh, F.; Eberhardt, E.L.; Miller, D.D.; Albrecht, D.M.; Van Doorn, R.; Hruby, M.D.; Maresh, M.E.; Clipson, L.; Mukhtar, H. Fisetin and 5-fluorouracil: Effective combination for PIK3CA-mutant colorectal cancer. Int. J. Cancer 2019, 145, 3022–3032. [Google Scholar] [CrossRef]
- Jeng, L.B.; Kumar Velmurugan, B.; Chen, M.C.; Hsu, H.H.; Ho, T.J.; Day, C.H.; Lin, Y.M.; Padma, V.V.; Tu, C.C.; Huang, C.Y. Fisetin mediated apoptotic cell death in parental and Oxaliplatin/irinotecan resistant colorectal cancer cells in vitro and in vivo. J. Cell. Physiol. 2018, 233, 7134–7142. [Google Scholar] [CrossRef] [PubMed]
- Leu, J.D.; Wang, B.S.; Chiu, S.J.; Chan, C.Y.; Chen, C.C.; Chen, F.D.; Avirmed, S.; Lee, Y.J. Combining fisetin and ionizing radiation suppresses the growth of mammalian colorectal cancers in xenograft tumor models. Oncol. Lett. 2016, 12, 4975–4982. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, Q.; Song, L.; He, T.; Li, Y.; Li, L.; Su, W.; Liu, L.; Qian, Z.; Gong, C. Polymeric micelles encapsulating fisetin improve the therapeutic effect in colon cancer. ACS Appl. Mater. Interfaces 2015, 7, 534–542. [Google Scholar] [CrossRef]
- Li, L.; Wang, M.; Yang, H.; Li, Y.; Huang, X.; Guo, J.; Liu, Z. Fisetin Inhibits Trypsin Activity and Suppresses the Growth of Colorectal Cancer in Vitro and in Vivo. Nat. Prod. Commun. 2022, 17, 1934578X221115511. [Google Scholar] [CrossRef]
- Winkelmann, I.; Diehl, D.; Oesterle, D.; Daniel, H.; Wenzel, U. The suppression of aberrant crypt multiplicity in colonic tissue of 1, 2-dimethylhydrazine-treated C57BL/6J mice by dietary flavone is associated with an increased expression of Krebs cycle enzymes. Carcinogenesis 2007, 28, 1446–1454. [Google Scholar] [CrossRef]
- Auyeung, K.K.-W.; Law, P.-C.; Ko, J.K.-S. Novel anti-angiogenic effects of formononetin in human colon cancer cells and tumor xenograft. Oncol. Rep. 2012, 28, 2188–2194. [Google Scholar] [CrossRef]
- Ma, Z.; Bao, X.; Gu, J. Furowanin A-induced autophagy alleviates apoptosis and promotes cell cycle arrest via inactivation STAT3/Mcl-1 axis in colorectal cancer. Life Sci. 2019, 218, 47–57. [Google Scholar] [CrossRef]
- Sekar, V.; Anandasadagopan, S.K.; Ganapasam, S. Genistein regulates tumor microenvironment and exhibits anticancer effect in dimethyl hydrazine-induced experimental colon carcinogenesis. Biofactors 2016, 42, 623–637. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Q.; Zhou, D.; Chen, H. Genistein, a soya isoflavone, prevents azoxymethane-induced up-regulation of WNT/β-catenin signalling and reduces colon pre-neoplasia in rats. Br. J. Nutr. 2013, 109, 33–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, X.; Liu, Z.; Wang, R.; Wang, J.; Zhang, S.; Cai, X.; Wu, K.; Bergan, R.C.; Xu, L.; Fan, D. Genistein suppresses FLT4 and inhibits human colorectal cancer metastasis. Oncotarget 2015, 6, 3225. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Song, Z.-J.; He, X.; Zhang, R.-Q.; Zhang, C.-F.; Li, F.; Wang, C.-Z.; Yuan, C.-S. Antitumor and immunomodulatory activity of genkwanin on colorectal cancer in the APCMin/+ mice. Int. Immunopharmacol. 2015, 29, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.-F.; Yin, C.-M.; Ouyang, T.; Sun, S.-D.; Chen, W.-G.; Yang, X.-L.; He, X.; Zhang, C.-F. Self-Nanoemulsifying Drug Delivery System of Genkwanin: A Novel Approach for Anti-Colitis-Associated Colorectal Cancer. Drug Des. Dev. Ther. 2021, 15, 557. [Google Scholar] [CrossRef] [PubMed]
- Saiprasad, G.; Chitra, P.; Manikandan, R.; Sudhandiran, G. Hesperidin alleviates oxidative stress and downregulates the expressions of proliferative and inflammatory markers in azoxymethane-induced experimental colon carcinogenesis in mice. Inflamm. Res. 2013, 62, 425–440. [Google Scholar] [CrossRef]
- Saiprasad, G.; Chitra, P.; Manikandan, R.; Sudhandiran, G. Hesperidin induces apoptosis and triggers autophagic markers through inhibition of Aurora-A mediated phosphoinositide-3-kinase/Akt/mammalian target of rapamycin and glycogen synthase kinase-3 beta signalling cascades in experimental colon carcinogenesis. Eur. J. Cancer 2014, 50, 2489–2507. [Google Scholar] [CrossRef]
- Tanaka, T.; Makita, H.; Kawabata, K.; Mori, H.; Kakumoto, M.; Satoh, K.; Hara, A.; Sumida, T.; Tanaka, T.; Ogawa, H. Chemoprevention of azoxymethane-induced rat colon carcinogenesis by the naturally occurring flavonoids, diosmin and hesperidin. Carcinogenesis 1997, 18, 957–965. [Google Scholar] [CrossRef]
- El-Deek, S.E.; Abd-Elghaffar, S.K.; Hna, R.S.; Mohamed, H.G.; El-Deek, H.E. Effect of hesperidin against induced colon cancer in rats: Impact of Smad4 and activin a signaling pathway. Nutr. Cancer 2022, 74, 697–714. [Google Scholar] [CrossRef]
- Zhou, J.; Zhao, R.; Ye, T.; Yang, S.; Li, Y.; Yang, F.; Wang, G.; Xie, Y.; Li, Q. Antitumor activity in colorectal cancer induced by hinokiflavone. J. Gastroenterol. Hepatol. 2019, 34, 1571–1580. [Google Scholar] [CrossRef]
- Shi, C.-J.; Li, S.-Y.; Shen, C.-H.; Pan, F.-F.; Deng, L.-Q.; Fu, W.-M.; Wang, J.-Y.; Zhang, J.-F. Icariside II suppressed tumorigenesis by epigenetically regulating the circβ-catenin-Wnt/β-catenin axis in colorectal cancer. Bioorganic Chem. 2022, 124, 105800. [Google Scholar] [CrossRef]
- Zhou, C.; Gu, J.; Zhang, G.; Dong, D.; Yang, Q.; Chen, M.-B.; Xu, D. AMPK-autophagy inhibition sensitizes icaritin-induced anti-colorectal cancer cell activity. Oncotarget 2017, 8, 14736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, S.; Cai, S.; Ji, S.; Yan, X.; Zhang, W.; Qiao, X.; Zhang, H.; Ye, M.; Yu, S. Isoangustone A induces autophagic cell death in colorectal cancer cells by activating AMPK signaling. Fitoterapia 2021, 152, 104935. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhang, X.; Chen, X.; Li, Y.; Ke, Z.; Tang, T.; Chai, H.; Guo, A.M.; Chen, H.; Yang, J. Isoliquiritigenin, a flavonoid from licorice, blocks M2 macrophage polarization in colitis-associated tumorigenesis through downregulating PGE2 and IL-6. Toxicol. Appl. Pharmacol. 2014, 279, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Takasuka, N.; Iigo, M.; Baba, M.; Nishino, H.; Tsuda, H.; Okuyama, T. Isoliquiritigenin, a flavonoid from licorice, reduces prostaglandin E2 and nitric oxide, causes apoptosis, and suppresses aberrant crypt foci development. Cancer Sci. 2004, 95, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Antunes-Ricardo, M.; Guardado-Félix, D.; Rocha-Pizaña, M.; Garza-Martínez, J.; Acevedo-Pacheco, L.; Gutiérrez-Uribe, J.; Villela-Castrejón, J.; López-Pacheco, F.; Serna-Saldívar, S. Opuntia ficus-indica Extract and Isorhamnetin-3-O-Glucosyl-Rhamnoside Diminish Tumor Growth of Colon Cancer Cells Xenografted in Immune-Suppressed Mice through the Activation of Apoptosis Intrinsic Pathway. Plant Foods Hum. Nutr. 2021, 76, 434–441. [Google Scholar] [CrossRef]
- Saud, S.M.; Young, M.R.; Jones-Hall, Y.L.; Ileva, L.; Evbuomwan, M.O.; Wise, J.; Colburn, N.H.; Kim, Y.S.; Bobe, G. Chemopreventive Activity of Plant Flavonoid Isorhamnetin in Colorectal Cancer Is Mediated by Oncogenic Src and β-CateninIsorhamnetin-Induced CSK Inhibits Colorectal Cancer. Cancer Res. 2013, 73, 5473–5484. [Google Scholar] [CrossRef]
- Nirmala, P.; Ramanathan, M. Effect of kaempferol on lipid peroxidation and antioxidant status in 1, 2-dimethyl hydrazine induced colorectal carcinoma in rats. Eur. J. Pharmacol. 2011, 654, 75–79. [Google Scholar] [CrossRef]
- Hassan, E.S.; Hassanein, N.M.; Ahmed, H.M.S. Probing the chemoprevention potential of the antidepressant fluoxetine combined with epigallocatechin gallate or kaempferol in rats with induced early stage colon carcinogenesis. J. Pharmacol. Sci. 2021, 145, 29–41. [Google Scholar] [CrossRef]
- Hassanein, N.M.; Hassan, E.S.; Hegab, A.M.; Elahl, H.M.S. Chemopreventive effect of sulindac in combination with epigallocatechin gallate or kaempferol against 1, 2-dimethyl hydrazine-induced preneoplastic lesions in rats: A comparative study. J. Biochem. Mol. Toxicol. 2018, 32, e22198. [Google Scholar] [CrossRef]
- Osman, N.H.; Said, U.Z.; El-Waseef, A.M.; Ahmed, E.S. Luteolin supplementation adjacent to aspirin treatment reduced dimethylhydrazine-induced experimental colon carcinogenesis in rats. Tumor Biol. 2015, 36, 1179–1190. [Google Scholar] [CrossRef]
- Pandurangan, A.K.; Kumar, S.A.S.; Dharmalingam, P.; Ganapasam, S. Luteolin, a bioflavonoid inhibits azoxymethane-induced colon carcinogenesis: Involvement of iNOS and COX-2. Pharmacogn. Mag. 2014, 10, S306. [Google Scholar] [PubMed] [Green Version]
- Pandurangan, A.K.; Ananda Sadagopan, S.K.; Dharmalingam, P.; Ganapasam, S. Luteolin, a bioflavonoid inhibits Azoxymethane-induced colorectal cancer through activation of Nrf2 signaling. Toxicol. Mech. Methods 2014, 24, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Jung, S.K.; Byun, S.; Son, J.E.; Oh, M.H.; Lee, J.; Kang, M.J.; Heo, Y.S.; Lee, K.W.; Lee, H.J. Raf and PI3K are the molecular targets for the anti-metastatic effect of Luteolin. Phytother. Res. 2013, 27, 1481–1488. [Google Scholar] [CrossRef] [PubMed]
- Pandurangan, A.; Dharmalingam, P.; Sadagopan, S.; Ganapasam, S. Luteolin inhibits matrix metalloproteinase 9 and 2 in azoxymethane-induced colon carcinogenesis. Hum. Exp. Toxicol. 2014, 33, 1176–1185. [Google Scholar] [CrossRef]
- Yao, Y.; Rao, C.; Zheng, G.; Wang, S. Luteolin suppresses colorectal cancer cell metastasis via regulation of the miR-384/pleiotrophin axis. Oncol. Rep. 2019, 42, 131–141. [Google Scholar] [CrossRef]
- Xiao, B.; Qin, Y.; Ying, C.; Ma, B.; Wang, B.; Long, F.; Wang, R.; Fang, L.; Wang, Y. Combination of oncolytic adenovirus and luteolin exerts synergistic antitumor effects in colorectal cancer cells and a mouse model. Mol. Med. Rep. 2017, 16, 9375–9382. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Jiang, J.; Hou, L.; Ji, F. Lysionotin Induces Ferroptosis to Suppress Development of Colorectal Cancer via Promoting Nrf2 Degradation. Oxidative Med. Cell. Longev. 2022, 2022, 1366957. [Google Scholar] [CrossRef]
- Yu, H.; Yin, S.; Zhou, S.; Shao, Y.; Sun, J.; Pang, X.; Han, L.; Zhang, Y.; Gao, X.; Jin, C. Magnolin promotes autophagy and cell cycle arrest via blocking LIF/Stat3/Mcl-1 axis in human colorectal cancers. Cell Death Dis. 2018, 9, 702. [Google Scholar] [CrossRef]
- Chen, R.; Zhang, L. Morin inhibits colorectal tumor growth through inhibition of NF-κB signaling pathway. Immunopharmacol. Immunotoxicol. 2019, 41, 622–629. [Google Scholar] [CrossRef]
- Sharma, S.H.; Thulasingam, S.; Chellappan, D.R.; Chinnaswamy, P.; Nagarajan, S. Morin and Esculetin supplementation modulates c-myc induced energy metabolism and attenuates neoplastic changes in rats challenged with the procarcinogen 1, 2-dimethylhydrazine. Eur. J. Pharmacol. 2017, 796, 20–31. [Google Scholar] [CrossRef]
- Lori, G.; Paoli, P.; Femia, A.P.; Pranzini, E.; Caselli, A.; Tortora, K.; Romagnoli, A.; Raugei, G.; Caderni, G. Morin-dependent inhibition of low molecular weight protein tyrosine phosphatase (LMW-PTP) restores sensitivity to apoptosis during colon carcinogenesis: Studies in vitro and in vivo, in an Apc-driven model of colon cancer. Mol. Carcinog. 2019, 58, 686–698. [Google Scholar] [CrossRef] [PubMed]
- Vennila, S.; Nalini, N. Modifying effects of morin on the development of aberrant crypt foci and bacterial enzymes in experimental colon cancer. Food Chem. Toxicol. 2009, 47, 309–315. [Google Scholar]
- Sharma, S.H.; Kumar, J.S.; Chellappan, D.R.; Nagarajan, S. Molecular chemoprevention by morin–A plant flavonoid that targets nuclear factor kappa B in experimental colon cancer. Biomed. Pharmacother. 2018, 100, 367–373. [Google Scholar] [CrossRef]
- Sreedharan, V.; Venkatachalam, K.K.; Namasivayam, N. Effect of morin on tissue lipid peroxidation and antioxidant status in 1, 2-dimethylhydrazine induced experimental colon carcinogenesis. Investig. New Drugs 2009, 27, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Nirmala, P.; Ramanathan, M. Effect of myricetin on 1, 2 dimethylhydrazine induced rat colon carcinogenesis. J. Exp. Ther. Oncol. 2011, 9, 101–108. [Google Scholar]
- Li, Y.; Cui, S.-X.; Sun, S.-Y.; Shi, W.-N.; Song, Z.-Y.; Wang, S.-Q.; Yu, X.-F.; Gao, Z.-H.; Qu, X.-J. Chemoprevention of intestinal tumorigenesis by the natural dietary flavonoid myricetin in APCMin/+ mice. Oncotarget 2016, 7, 60446. [Google Scholar] [CrossRef]
- Zhang, M.-J.; Su, H.; Yan, J.-Y.; Li, N.; Song, Z.-Y.; Wang, H.-J.; Huo, L.-G.; Wang, F.; Ji, W.-S.; Qu, X.-J. Chemopreventive effect of Myricetin, a natural occurring compound, on colonic chronic inflammation and inflammation-driven tumorigenesis in mice. Biomed. Pharmacother. 2018, 97, 1131–1137. [Google Scholar] [CrossRef]
- Wang, F.; Song, Z.-Y.; Qu, X.-J.; Li, F.; Zhang, L.; Li, W.-B.; Cui, S.-X. M10, a novel derivative of Myricetin, prevents ulcerative colitis and colorectal tumor through attenuating robust endoplasmic reticulum stress. Carcinogenesis 2018, 39, 889–899. [Google Scholar] [CrossRef]
- Leonardi, T.; Vanamala, J.; Taddeo, S.S.; Davidson, L.A.; Murphy, M.E.; Patil, B.S.; Wang, N.; Carroll, R.J.; Chapkin, R.S.; Lupton, J.R. Apigenin and naringenin suppress colon carcinogenesis through the aberrant crypt stage in azoxymethane-treated rats. Exp. Biol. Med. 2010, 235, 710–717. [Google Scholar] [CrossRef]
- Dou, W.; Zhang, J.; Sun, A.; Zhang, E.; Ding, L.; Mukherjee, S.; Wei, X.; Chou, G.; Wang, Z.-T.; Mani, S. Protective effect of naringenin against experimental colitis via suppression of Toll-like receptor 4/NF-κB signalling. Br. J. Nutr. 2013, 110, 599–608. [Google Scholar] [CrossRef]
- Li, H.; Zhu, F.; Chen, H.; Cheng, K.W.; Zykova, T.; Oi, N.; Lubet, R.A.; Bode, A.M.; Wang, M.; Dong, Z. 6-C-(E-phenylethenyl)-Naringenin Suppresses Colorectal Cancer Growth by Inhibiting Cyclooxygenase-16CEPN Suppresses Colon Cancer by Inhibiting COX-1. Cancer Res. 2014, 74, 243–252. [Google Scholar] [CrossRef]
- Sequetto, P.L.; Oliveira, T.T.; Maldonado, I.R.; Augusto, L.E.F.; Mello, V.J.; Pizziolo, V.R.; Almeida, M.R.; Silva, M.E.; Novaes, R.D. Naringin accelerates the regression of pre-neoplastic lesions and the colorectal structural reorganization in a murine model of chemical carcinogenesis. Food Chem. Toxicol. 2014, 64, 200–209. [Google Scholar] [CrossRef]
- Zhang, Y.-S.; Wang, F.; Cui, S.-X.; Qu, X.-J. Natural dietary compound naringin prevents azoxymethane/dextran sodium sulfate-induced chronic colorectal inflammation and carcinogenesis in mice. Cancer Biol. Ther. 2018, 19, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Vanamala, J.; Leonardi, T.; Patil, B.S.; Taddeo, S.S.; Murphy, M.E.; Pike, L.M.; Chapkin, R.S.; Lupton, J.R.; Turner, N.D. Suppression of colon carcinogenesis by bioactive compounds in grapefruit. Carcinogenesis 2006, 27, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, S.; Yasui, Y.; Tanaka, T.; Ohigashi, H.; Murakami, A. Suppressive effects of nobiletin on hyperleptinemia and colitis-related colon carcinogenesis in male ICR mice. Carcinogenesis 2008, 29, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Thangaraj, K.; Vaiyapuri, M. Orientin, a C-glycosyl dietary flavone, suppresses colonic cell proliferation and mitigates NF-κB mediated inflammatory response in 1, 2-dimethylhydrazine induced colorectal carcinogenesis. Biomed. Pharmacother. 2017, 96, 1253–1266. [Google Scholar] [CrossRef]
- Thangaraj, K.; Natesan, K.; Palani, M.; Vaiyapuri, M. Orientin, a flavanoid, mitigates 1, 2 dimethylhydrazine-induced colorectal lesions in Wistar rats fed a high-fat diet. Toxicol. Rep. 2018, 5, 977–987. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, F.; Wang, Y.; Cai, M.; Wang, Q.; Guo, Q.; Li, Z.; Hu, R. Oroxylin A inhibits colitis-associated carcinogenesis through modulating the IL-6/STAT3 signaling pathway. Inflamm. Bowel Dis. 2013, 19, 1990–2000. [Google Scholar] [CrossRef]
- Ni, T.; He, Z.; Dai, Y.; Yao, J.; Guo, Q.; Wei, L. Oroxylin A suppresses the development and growth of colorectal cancer through reprogram of HIF1α-modulated fatty acid metabolism. Cell Death Dis. 2017, 8, e2865. [Google Scholar] [CrossRef]
- Gan, C.; Li, Y.; Yu, Y.; Yu, X.; Liu, H.; Zhang, Q.; Yin, W.; Yu, L.; Ye, T. Natural product pectolinarigenin exhibits potent anti-metastatic activity in colorectal carcinoma cells in vitro and in vivo. Bioorganic Med. Chem. 2019, 27, 115089. [Google Scholar] [CrossRef]
- Lim, S.; Xu, J.; Kim, J.; Chen, T.Y.; Su, X.; Standard, J.; Carey, E.; Griffin, J.; Herndon, B.; Katz, B. Role of anthocyanin-enriched purple-fleshed sweet potato p40 in colorectal cancer prevention. Mol. Nutr. Food Res. 2013, 57, 1908–1917. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Yan, Y.; Fan, M.; Gao, S.; Zhang, L.; Xiong, X.; Li, R.; Xiao, X.; Wang, X.; Ni, L. Pt3R5G inhibits colon cancer cell proliferation through inducing ferroptosis by down-regulating SLC7A11. Life Sci. 2022, 306, 120859. [Google Scholar] [CrossRef]
- Lin, S.-T.; Tu, S.-H.; Yang, P.-S.; Hsu, S.-P.; Lee, W.-H.; Ho, C.-T.; Wu, C.-H.; Lai, Y.-H.; Chen, M.-Y.; Chen, L.-C. Apple polyphenol phloretin inhibits colorectal cancer cell growth via inhibition of the type 2 glucose transporter and activation of p53-mediated signaling. J. Agric. Food Chem. 2016, 64, 6826–6837. [Google Scholar] [CrossRef]
- Hao, X.; Xiao, H.; Ju, J.; Lee, M.-J.; Lambert, J.D.; Yang, C.S. Green tea polyphenols inhibit colorectal tumorigenesis in azoxymethane-treated F344 rats. Nutr. Cancer 2017, 69, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Gossé, F.; Guyot, S.; Roussi, S.; Lobstein, A.; Fischer, B.; Seiler, N.; Raul, F. Chemopreventive properties of apple procyanidins on human colon cancer-derived metastatic SW620 cells and in a rat model of colon carcinogenesis. Carcinogenesis 2005, 26, 1291–1295. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Tian, X.; Yang, M.; Yu, Y.; Zhou, Y.; Gao, Y.; Zhang, L.; Li, Z.; Xiao, Y.; Moses, R.E. Procyanidin B2 promotes intestinal injury repair and attenuates colitis-associated tumorigenesis via suppression of oxidative stress in mice. Antioxid. Redox Signal. 2021, 35, 75–92. [Google Scholar] [CrossRef]
- Tutino, V.; De Nunzio, V.; Tafaro, A.; Bianco, G.; Gigante, I.; Scavo, M.P.; D’ALESSANDRO, R.; Refolo, M.G.; Messa, C.; Caruso, M.G. Cannabinoid receptor-1 up-regulation in azoxymethane (AOM)-treated mice after dietary treatment with quercetin. Anticancer Res. 2018, 38, 4485–4491. [Google Scholar] [CrossRef]
- Lin, C.; Yu, Y.; Zhao, H.-g.; Yang, A.; Yan, H.; Cui, Y. Combination of quercetin with radiotherapy enhances tumor radiosensitivity in vitro and in vivo. Radiother. Oncol. 2012, 104, 395–400. [Google Scholar] [CrossRef]
- Warren, C.A.; Paulhill, K.J.; Davidson, L.A.; Lupton, J.R.; Taddeo, S.S.; Hong, M.Y.; Carroll, R.J.; Chapkin, R.S.; Turner, N.D. Quercetin may suppress rat aberrant crypt foci formation by suppressing inflammatory mediators that influence proliferation and apoptosis. J. Nutr. 2009, 139, 101–105. [Google Scholar] [CrossRef]
- Lin, R.; Piao, M.; Song, Y.; Liu, C. Quercetin suppresses AOM/DSS-induced colon carcinogenesis through its anti-inflammation effects in mice. J. Immunol. Res. 2020, 2020, 9242601. [Google Scholar] [CrossRef]
- Kee, J.-Y.; Han, Y.-H.; Kim, D.-S.; Mun, J.-G.; Park, J.; Jeong, M.-Y.; Um, J.-Y.; Hong, S.-H. Inhibitory effect of quercetin on colorectal lung metastasis through inducing apoptosis, and suppression of metastatic ability. Phytomedicine 2016, 23, 1680–1690. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Z.; Jin, J.; Zhu, S.-X.; He, G.-Q.; Li, S.-H.; Wang, J.; Cai, Y. Quercetin pretreatment enhances the radiosensitivity of colon cancer cells by targeting Notch-1 pathway. Biochem. Biophys. Res. Commun. 2020, 523, 947–953. [Google Scholar] [CrossRef]
- Alonso-Castro, A.J.; Domínguez, F.; García-Carrancá, A. Rutin exerts antitumor effects on nude mice bearing SW480 tumor. Arch. Med. Res. 2013, 44, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Chen, L.; Sun, Q.; Zhao, H.; Yang, H.; Ren, S.; Liu, M.; Meng, X.; Xu, H. Scutellarin ameliorates colitis-associated colorectal cancer by suppressing Wnt/β-catenin signaling cascade. Eur. J. Pharmacol. 2021, 906, 174253. [Google Scholar] [CrossRef]
- Xiong, L.-L.; Du, R.-L.; Xue, L.-L.; Jiang, Y.; Huang, J.; Chen, L.; Liu, J.; Wang, T.-H. Anti-colorectal cancer effects of scutellarin revealed by genomic and proteomic analysis. Chin. Med. 2020, 15, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Tan, L.; Sun, Q.; Chen, L.; Zhao, H.; Liu, M.; Yang, H.; Ren, S.; Ming, T.; Tang, S. Suppression of colitis-associated colorectal cancer by scutellarin through inhibiting Hedgehog signaling pathway activity. Phytomedicine 2022, 98, 153972. [Google Scholar] [CrossRef]
- Yang, S.-H.; Lin, J.-K.; Huang, C.-J.; Chen, W.-S.; Li, S.-Y.; Chiu, J.-H. Silibinin inhibits angiogenesis via Flt-1, but not KDR, receptor up-regulation1. J. Surg. Res. 2005, 128, 140–146. [Google Scholar] [CrossRef]
- Kauntz, H.; Bousserouel, S.; Gosse, F.; Marescaux, J.; Raul, F. Silibinin, a natural flavonoid, modulates the early expression of chemoprevention biomarkers in a preclinical model of colon carcinogenesis. Int. J. Oncol. 2012, 41, 849–854. [Google Scholar] [CrossRef]
- Bao, H.; Zheng, N.; Li, Z.; Zhi, Y. Synergistic effect of tangeretin and atorvastatin for colon cancer combination therapy: Targeted delivery of these dual drugs using RGD Peptide decorated nanocarriers. Drug Des. Dev. Ther. 2020, 14, 3057. [Google Scholar] [CrossRef]
- Razak, S.; Afsar, T.; Ullah, A.; Almajwal, A.; Alkholief, M.; Alshamsan, A.; Jahan, S. Taxifolin, a natural flavonoid interacts with cell cycle regulators causes cell cycle arrest and causes tumor regression by activating Wnt/β-catenin signaling pathway. BMC Cancer 2018, 18, 1–18. [Google Scholar] [CrossRef]
- Yue, G.G.-L.; Gao, S.; Lee, J.K.-M.; Chan, Y.-Y.; Wong, E.C.-W.; Zheng, T.; Li, X.-X.; Shaw, P.-C.; Simmonds, M.S.; Lau, C.B.-S. A natural flavone tricin from grains can alleviate tumor growth and lung metastasis in colorectal tumor mice. Molecules 2020, 25, 3730. [Google Scholar] [CrossRef]
- Oyama, T.; Yasui, Y.; Sugie, S.; Koketsu, M.; Watanabe, K.; Tanaka, T. Dietary tricin suppresses inflammation-related colon carcinogenesis in male Crj: CD-1 mice. Cancer Prev. Res. 2009, 2, 1031–1038. [Google Scholar] [CrossRef]
- Vinothkumar, R.; Kumar, R.V.; Sudha, M.; Viswanathan, P.; Balasubramanian, T.; Nalini, N. Modulatory effect of troxerutin on biotransforming enzymes and preneoplasic lesions induced by 1, 2-dimethylhydrazine in rat colon carcinogenesis. Exp. Mol. Pathol. 2014, 96, 15–26. [Google Scholar] [CrossRef]
- Bhardwaj, M.; Paul, S.; Jakhar, R.; Khan, I.; Kang, J.I.; Kim, H.M.; Yun, J.W.; Lee, S.-J.; Cho, H.J.; Lee, H.G. Vitexin confers HSF-1 mediated autophagic cell death by activating JNK and ApoL1 in colorectal carcinoma cells. Oncotarget 2017, 8, 112426. [Google Scholar] [CrossRef]
- Bhardwaj, M.; Cho, H.J.; Paul, S.; Jakhar, R.; Khan, I.; Lee, S.-J.; Kim, B.-Y.; Krishnan, M.; Khaket, T.P.; Lee, H.G. Vitexin induces apoptosis by suppressing autophagy in multi-drug resistant colorectal cancer cells. Oncotarget 2018, 9, 3278. [Google Scholar] [CrossRef]
- Yao, J.; Zhao, L.; Zhao, Q.; Zhao, Y.; Sun, Y.; Zhang, Y.; Miao, H.; You, Q.; Hu, R.; Guo, Q. NF-κB and Nrf2 signaling pathways contribute to wogonin-mediated inhibition of inflammation-associated colorectal carcinogenesis. Cell Death Dis. 2014, 5, e1283. [Google Scholar] [CrossRef]
- Feng, Q.; Wang, H.; Pang, J.; Ji, L.; Han, J.; Wang, Y.; Qi, X.; Liu, Z.; Lu, L. Prevention of wogonin on colorectal cancer tumorigenesis by regulating p53 nuclear translocation. Front. Pharmacol. 2018, 9, 1356. [Google Scholar] [CrossRef]
- You, W.; Di, A.; Zhang, L.; Zhao, G. Effects of wogonin on the growth and metastasis of colon cancer through the Hippo signaling pathway. Bioengineered 2022, 13, 2586–2597. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, L.; Li, G.; Gao, Z. Xanthohumol protects against Azoxymethane-induced colorectal cancer in Sprague-Dawley rats. Environ. Toxicol. 2020, 35, 136–144. [Google Scholar] [CrossRef]
- Murillo, G.; Hirschelman, W.H.; Ito, A.; Moriarty, R.M.; Kinghorn, A.D.; Pezzuto, J.M.; Mehta, R.G. Zapotin, a phytochemical present in a Mexican fruit, prevents colon carcinogenesis. Nutr. Cancer 2007, 57, 28–37. [Google Scholar] [CrossRef]
- Kang, N.J.; Lee, K.W.; Kim, B.H.; Bode, A.M.; Lee, H.-J.; Heo, Y.-S.; Boardman, L.; Limburg, P.; Lee, H.J.; Dong, Z. Coffee phenolic phytochemicals suppress colon cancer metastasis by targeting MEK and TOPK. Carcinogenesis 2011, 32, 921–928. [Google Scholar] [CrossRef]
- Park, S.-R.; Kim, S.-R.; Hong, I.-S.; Lee, H.-Y. A novel therapeutic approach for colorectal cancer stem cells: Blocking the PI3K/Akt signaling axis with caffeic acid. Front. Cell Dev. Biol. 2020, 8, 585987. [Google Scholar] [CrossRef]
- Chiang, E.-P.I.; Tsai, S.-Y.; Kuo, Y.-H.; Pai, M.-H.; Chiu, H.-L.; Rodriguez, R.L.; Tang, F.-Y. Caffeic acid derivatives inhibit the growth of colon cancer: Involvement of the PI3-K/Akt and AMPK signaling pathways. PLoS ONE 2014, 9, e99631. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Yao, X.; Yao, C.; Zhao, X.; Zuo, H.; Li, Z. Anti-colon cancer effect of caffeic acid p-nitro-phenethyl ester in vitro and in vivo and detection of its metabolites. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Chen, C.; Kuo, Y.-H.; Lin, C.-C.; Chao, C.-Y.; Pai, M.-H.; Chiang, E.-P.I.; Tang, F.-Y. Decyl caffeic acid inhibits the proliferation of colorectal cancer cells in an autophagy-dependent manner in vitro and in vivo. PLoS ONE 2020, 15, e0232832. [Google Scholar] [CrossRef]
- Tanaka, T.; Nishikawa, A.; Shima, H.; Sugie, S.; Shinoda, T.; Yoshimi, N.; Iwata, H.; Mori, H. Inhibitory effects of chlorogenic acid, reserpine, polyprenoic acid (E-5166), or coffee on hepatocarcinogenesis in rats and hamsters. Basic Life Sci. 1990, 52, 429–440. [Google Scholar]
- Morishita, Y.; Yoshimi, N.; Kawabata, K.; Matsunaga, K.; Sugie, S.; Tanaka, T.; Mori, H. Regressive effects of various chemopreventive agents on azoxymethane-induced aberrant crypt foci in the rat colon. Jpn. J. Cancer Res. 1997, 88, 815–820. [Google Scholar] [CrossRef]
- Rao, C.V.; Tokumo, K.; Rigotty, J.; Zang, E.; Kelloff, G.; Reddy, B.S. Chemoprevention of colon carcinogenesis by dietary administration of piroxicam, α-difluoromethylornithine, 16α-fluoro-5-androsten-17-one, and ellagic acid individually and in combination. Cancer Res. 1991, 51, 4528–4534. [Google Scholar]
- Umesalma, S.; Sudhandiran, G. Chemomodulation of the antioxidative enzymes and peroxidative damage in the colon of 1, 2-dimethyl hydrazine-induced rats by ellagicacid. Phytother. Res. 2010, 24, S114–S119. [Google Scholar] [CrossRef]
- Umesalma, S.; Sudhandiran, G. Differential inhibitory effects of the polyphenol ellagic acid on inflammatory mediators NF-κB, iNOS, COX-2, TNF-α, and IL-6 in 1, 2-dimethylhydrazine-induced rat colon carcinogenesis. Basic Clin. Pharmacol. Toxicol. 2010, 107, 650–655. [Google Scholar] [CrossRef]
- Umesalma, S.; Sudhandiran, G. Ellagic acid prevents rat colon carcinogenesis induced by 1, 2 dimethyl hydrazine through inhibition of AKT-phosphoinositide-3 kinase pathway. Eur. J. Pharmacol. 2011, 660, 249–258. [Google Scholar] [CrossRef]
- Kumar, K.N.; Raja, S.B.; Vidhya, N.; Devaraj, S.N. Ellagic acid modulates antioxidant status, ornithine decarboxylase expression, and aberrant crypt foci progression in 1, 2-dimethylhydrazine-instigated colon preneoplastic lesions in rats. J. Agric. Food Chem. 2012, 60, 3665–3672. [Google Scholar] [CrossRef]
- Goyal, Y.; Koul, A.; Ranawat, P. Ellagic acid modulates cisplatin toxicity in DMH induced colorectal cancer: Studies on membrane alterations. Biochem. Biophys. Rep. 2022, 31, 101319. [Google Scholar] [CrossRef]
- Kawabata, K.; Yamamoto, T.; Hara, A.; Shimizu, M.; Yamada, Y.; Matsunaga, K.; Tanaka, T.; Mori, H. Modifying effects of ferulic acid on azoxymethane-induced colon carcinogenesis in F344 rats. Cancer Lett. 2000, 157, 15–21. [Google Scholar] [CrossRef]
- Han, B.S.; Park, C.B.; Takasuka, N.; Naito, A.; Sekine, K.; Nomura, E.; Taniguchi, H.; Tsuno, T.; Tsuda, H. A Ferulic Acid Derivative, Ethyl 3-(4′-Geranyloxy-3-methoxyphenyl)-2-propenoate, as a New Candidate Chemopreventive Agent for Colon Carcinogenesis in the Rat. Jpn. J. Cancer Res. 2001, 92, 404–409. [Google Scholar] [CrossRef]
- Giftson, J.S.; Jayanthi, S.; Nalini, N. Chemopreventive efficacy of gallic acid, an antioxidant and anticarcinogenic polyphenol, against 1, 2-dimethyl hydrazine induced rat colon carcinogenesis. Investig. New Drugs 2010, 28, 251–259. [Google Scholar] [CrossRef]
- Sanchez-Martin, V.; Plaza-Calonge, M.d.C.; Soriano-Lerma, A.; Ortiz-Gonzalez, M.; Linde-Rodriguez, A.; Perez-Carrasco, V.; Ramirez-Macias, I.; Cuadros, M.; Gutierrez-Fernandez, J.; Murciano-Calles, J. Gallic Acid: A Natural Phenolic Compound Exerting Antitumoral Activities in Colorectal Cancer via Interaction with G-Quadruplexes. Cancers 2022, 14, 2648. [Google Scholar] [CrossRef]
- Jeong, J.H.; Kim, E.Y.; Choi, H.J.; Chung, T.W.; Kim, K.J.; Kim, S.Y.; Ha, K.T. Gallic acid inhibits STAT3 phosphorylation and alleviates DDS-induced colitis via regulating cytokine production. J. Physiol. Pathol. Korean Med. 2016, 30, 338–346. [Google Scholar] [CrossRef]
- Lin, X.; Wang, G.; Liu, P.; Han, L.; Wang, T.; Chen, K.; Gao, Y. Gallic acid suppresses colon cancer proliferation by inhibiting SRC and EGFR phosphorylation. Exp. Ther. Med. 2021, 21, 1–11. [Google Scholar] [CrossRef]
- Hong, Z.; Tang, P.; Liu, B.; Ran, C.; Yuan, C.; Zhang, Y.; Lu, Y.; Duan, X.; Yang, Y.; Wu, H. Ferroptosis-related genes for overall survival prediction in patients with colorectal cancer can be inhibited by gallic acid. Int. J. Biol. Sci. 2021, 17, 942. [Google Scholar] [CrossRef]
- Senapathy, J.G.; Jayanthi, S.; Viswanathan, P.; Umadevi, P.; Nalini, N. Effect of gallic acid on xenobiotic metabolizing enzymes in 1, 2-dimethyl hydrazine induced colon carcinogenesis in Wistar rats–a chemopreventive approach. Food Chem. Toxicol. 2011, 49, 887–892. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.-A.; Liu, T.-B.; Lü, H.-N. Geraniin inhibits proliferation and induces apoptosis through inhibition of phosphatidylinositol 3-kinase/Akt pathway in human colorectal cancer in vitro and in vivo. Anti-Cancer Drugs 2020, 31, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.H.; Chellappan, D.R.; Chinnaswamy, P.; Nagarajan, S. Protective effect of p-coumaric acid against 1, 2 dimethylhydrazine induced colonic preneoplastic lesions in experimental rats. Biomed. Pharmacother. 2017, 94, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Zhu, S.; Niu, Z.; Yin, Y. The protective effect of syringic acid on dextran sulfate sodium-induced experimental colitis in BALB/c mice. Drug Dev. Res. 2019, 80, 731–740. [Google Scholar] [CrossRef]
- Mihanfar, A.; Darband, S.G.; Sadighparvar, S.; Kaviani, M.; Mirza-Aghazadeh-Attari, M.; Yousefi, B.; Majidinia, M. In vitro and in vivo anticancer effects of syringic acid on colorectal cancer: Possible mechanistic view. Chem.-Biol. Interact. 2021, 337, 109337. [Google Scholar] [CrossRef]
- Han, Y.-H.; Kee, J.-Y.; Kim, D.-S.; Mun, J.-g.; Jeong, M.-Y.; Park, S.-H.; Choi, B.-M.; Park, S.-J.; Kim, H.-J.; Um, J.-Y. Arctigenin inhibits lung metastasis of colorectal cancer by regulating cell viability and metastatic phenotypes. Molecules 2016, 21, 1135. [Google Scholar] [CrossRef]
- Kang, K.; Oh, S.H.; Yun, J.H.; Jho, E.H.; Kang, J.-H.; Batsuren, D.; Tunsag, J.; Park, K.H.; Kim, M.; Nho, C.W. A novel topoisomerase inhibitor, daurinol, suppresses growth of HCT116 cells with low hematological toxicity compared to etoposide. Neoplasia 2011, 13, 1043–1057. [Google Scholar] [CrossRef]
- Li, C.; Zhang, K.; Pan, G.; Ji, H.; Li, C.; Wang, X.; Hu, X.; Liu, R.; Deng, L.; Wang, Y. Dehydrodiisoeugenol inhibits colorectal cancer growth by endoplasmic reticulum stress-induced autophagic pathways. J. Exp. Clin. Cancer Res. 2021, 40, 1–15. [Google Scholar] [CrossRef]
- Kee, J.-Y.; Han, Y.-H.; Mun, J.-G.; Park, S.-H.; Jeon, H.D.; Hong, S.-H. Gomisin a suppresses colorectal lung metastasis by inducing AMPK/P38-mediated apoptosis and decreasing metastatic abilities of colorectal cancer cells. Front. Pharmacol. 2018, 9, 986. [Google Scholar] [CrossRef]
- Li, Q.; Ma, Y.; Liu, X.L.; Mu, L.; He, B.C.; Wu, K.; Sun, W.J. Anti-proliferative effect of honokiol on SW620 cells through upregulating BMP7 expression via the TGF-β1/p53 signaling pathway. Oncol. Rep. 2020, 44, 2093–2107. [Google Scholar] [CrossRef]
- Won, S.J.; Yen, C.H.; Liu, H.S.; Wu, S.Y.; Lan, S.H.; Jiang-Shieh, Y.F.; Lin, C.N.; Su, C.L. Justicidin A-induced autophagy flux enhances apoptosis of human colorectal cancer cells via class III PI3K and Atg5 pathway. J. Cell. Physiol. 2015, 230, 930–946. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-M.; Weng, Y.-S.; Kuan, L.-Y.; Chen, J.-H.; Hsu, F.-T. Suppression of PKCδ/NF-κB Signaling and Apoptosis Induction through Extrinsic/Intrinsic Pathways Are Associated with Magnolol-Inhibited Tumor Progression in Colorectal Cancer In Vitro and In Vivo. Int. J. Mol. Sci. 2020, 21, 3527. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.-J.; Park, H.J.; Chung, H.-J.; Min, H.-Y.; Park, E.J.; Lee, M.A.; Shin, Y.; Lee, S.K. Wnt/β-catenin signaling mediates the antitumor activity of magnolol in colorectal cancer cells. Mol. Pharmacol. 2012, 82, 168–177. [Google Scholar] [CrossRef]
- Pu, Z.; Zhang, W.; Wang, M.; Xu, M.; Xie, H.; Zhao, J. Schisandrin B Attenuates colitis-associated colorectal cancer through SIRT1 linked SMURF2 signaling. Am. J. Chin. Med. 2021, 49, 1773–1789. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Wang, Z.; Zhong, J.; Zhang, L.; Zhang, H.; Zhang, D.; Xu, X.; Zhong, X.; Wang, J.; Li, H. Secoisolariciresinol diglucoside induces pyroptosis by activating caspase-1 to cleave GSDMD in colorectal cancer cells. Drug Dev. Res. 2022, 83, 1152–1166. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, T.; Yang, C.; Bao, T.; Yang, X.; He, F.; Zhang, Y.; Zhu, L.; Chen, H.; Rong, S. Secoisolariciresinol diglucoside suppresses Dextran sulfate sodium salt-induced colitis through inhibiting NLRP1 inflammasome. Int. Immunopharmacol. 2020, 78, 105931. [Google Scholar] [CrossRef]
- Ohira, H.; Oikawa, D.; Kurokawa, Y.; Aoki, Y.; Omura, A.; Kiyomoto, K.; Nakagawa, W.; Mamoto, R.; Fujioka, Y.; Nakayama, T. Suppression of colonic oxidative stress caused by chronic ethanol administration and attenuation of ethanol-induced colitis and gut leakiness by oral administration of sesaminol in mice. Food Funct. 2022, 13, 9285–9298. [Google Scholar] [CrossRef]
- Shin, M.-K.; Jeon, Y.-D.; Hong, S.-H.; Kang, S.-H.; Kee, J.-Y.; Jin, J.-S. In vivo and In vitro effects of Tracheloside on colorectal cancer cell proliferation and metastasis. Antioxidants 2021, 10, 513. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhong, J.; Liu, Y.; Huang, Y.; Luo, F.; Zhou, Y.; Pan, X.; Cao, S.; Zhang, L.; Zhang, Y. Purified vitexin compound 1, a new neolignan isolated compound, promotes PUMA-dependent apoptosis in colorectal cancer. Cancer Med. 2018, 7, 6158–6169. [Google Scholar] [CrossRef]
- Kimura, Y. Long-term oral administration of piceatannol (3, 5, 3′, 4′-tetrahydroxystilbene) attenuates colon tumor growth induced by azoxymethane plus dextran sulfate sodium in C57BL/6J mice. Nutr. Cancer 2022, 74, 2184–2195. [Google Scholar] [CrossRef]
- Jin, Y.; Zhan, X.; Zhang, B.; Chen, Y.; Liu, C.; Yu, L. Polydatin exerts an antitumor effect through regulating the miR-382/PD-L1 axis in colorectal cancer. Cancer Biother. Radiopharm. 2020, 35, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Chiou, Y.-S.; Tsai, M.-L.; Wang, Y.-J.; Cheng, A.-C.; Lai, W.-M.; Badmaev, V.; Ho, C.-T.; Pan, M.-H. Pterostilbene inhibits colorectal aberrant crypt foci (ACF) and colon carcinogenesis via suppression of multiple signal transduction pathways in azoxymethane-treated mice. J. Agric. Food Chem. 2010, 58, 8833–8841. [Google Scholar] [CrossRef]
- Paul, S.; DeCastro, A.J.; Lee, H.J.; Smolarek, A.K.; So, J.Y.; Simi, B.; Wang, C.X.; Zhou, R.; Rimando, A.M.; Suh, N. Dietary intake of pterostilbene, a constituent of blueberries, inhibits the β-catenin/p65 downstream signaling pathway and colon carcinogenesis in rats. Carcinogenesis 2010, 31, 1272–1278. [Google Scholar] [CrossRef] [PubMed]
- Chiou, Y.-S.; Tsai, M.-L.; Nagabhushanam, K.; Wang, Y.-J.; Wu, C.-H.; Ho, C.-T.; Pan, M.-H. Pterostilbene is more potent than resveratrol in preventing azoxymethane (AOM)-induced colon tumorigenesis via activation of the NF-E2-related factor 2 (Nrf2)-mediated antioxidant signaling pathway. J. Agric. Food Chem. 2011, 59, 2725–2733. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Y.; Sun, C.; Chen, X.; Han, L.; Wang, T.; Liu, J.; Chen, X.; Zhao, D. Effect of pterostilbene, a natural derivative of resveratrol, in the treatment of colorectal cancer through Top1/Tdp1-mediated DNA repair pathway. Cancers 2021, 13, 4002. [Google Scholar] [CrossRef]
- Suh, N.; Paul, S.; Hao, X.; Simi, B.; Xiao, H.; Rimando, A.M.; Reddy, B.S. Pterostilbene, an active constituent of blueberries, suppresses aberrant crypt foci formation in the azoxymethane-induced colon carcinogenesis model in rats. Clin. Cancer Res. 2007, 13, 350–355. [Google Scholar] [CrossRef]
- Ji, Q.; Liu, X.; Han, Z.; Zhou, L.; Sui, H.; Yan, L.; Jiang, H.; Ren, J.; Cai, J.; Li, Q. Resveratrol suppresses epithelial-to-mesenchymal transition in colorectal cancer through TGF-β1/Smads signaling pathway mediated Snail/E-cadherin expression. BMC Cancer 2015, 15, 1–12. [Google Scholar] [CrossRef]
- Saud, S.M.; Li, W.; Morris, N.L.; Matter, M.S.; Colburn, N.H.; Kim, Y.S.; Young, M.R. Resveratrol prevents tumorigenesis in mouse model of Kras activated sporadic colorectal cancer by suppressing oncogenic Kras expression. Carcinogenesis 2014, 35, 2778–2786. [Google Scholar] [CrossRef]
- Alfaras, I.; Juan, M.E.; Planas, J.M. trans-Resveratrol reduces precancerous colonic lesions in dimethylhydrazine-treated rats. J. Agric. Food Chem. 2010, 58, 8104–8110. [Google Scholar] [CrossRef]
- Majumdar, A.P.; Banerjee, S.; Nautiyal, J.; Patel, B.B.; Patel, V.; Du, J.; Yu, Y.; Elliott, A.A.; Levi, E.; Sarkar, F.H. Curcumin synergizes with resveratrol to inhibit colon cancer. Nutr. Cancer 2009, 61, 544–553. [Google Scholar] [CrossRef]
- Sudha, T.; El-Far, A.H.; Mousa, D.S.; Mousa, S.A. Resveratrol and its nanoformulation attenuate growth and the angiogenesis of xenograft and orthotopic colon cancer models. Molecules 2020, 25, 1412. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.-H.; Chan, G.K.-L.; Duan, R.; Wang, H.-Y.; Kong, X.-P.; Dong, T.T.-X.; Tsim, K.W.-K. Synergy of ginkgetin and resveratrol in suppressing VEGF-induced angiogenesis: A therapy in treating colorectal cancer. Cancers 2019, 11, 1828. [Google Scholar] [CrossRef] [PubMed]
- Sepporta, M.V.; Fuccelli, R.; Rosignoli, P.; Ricci, G.; Servili, M.; Fabiani, R. Oleuropein prevents Azoxymethane-induced Colon crypt dysplasia and leukocytes DNA damage in a/J mice. J. Med. Food 2016, 19, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Che, Y.; Zhang, Y.; Chen, M.; Guo, Q.; Zhang, W. Thymol Isolated from Thymus vulgaris L. inhibits colorectal cancer cell growth and metastasis by suppressing the Wnt/β-catenin pathway. Drug Des. Dev. Ther. 2020, 14, 2535. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Feng, Y.; Jin, Y.; Liu, X.; Sui, H.; Chai, N.; Chen, X.; Liu, N.; Ji, Q.; Wang, Y. Verbascoside promotes apoptosis by regulating HIPK2–p53 signaling in human colorectal cancer. BMC Cancer 2014, 14, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Meena, A.; Luqman, S. Baicalin mediated regulation of key signaling pathways in cancer. Pharmacol. Res. 2021, 164, 105387. [Google Scholar] [CrossRef]
- Patel, V.B.; Misra, S.; Patel, B.B.; Majumdar, A.P. Colorectal cancer: Chemopreventive role of curcumin and resveratrol. Nutr. Cancer 2010, 62, 958–967. [Google Scholar] [CrossRef]
- Prasad, S.; Aggarwal, B.B. Turmeric, the golden spice. In Herbal Medicine: Biomolecular and Clinical Aspects, 2nd ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2011; pp. 263–288. [Google Scholar]
- Wang, Y.; Bu, C.; Wu, K.; Wang, R.; Wang, J. Curcumin protects the pancreas from acute pancreatitis via the mitogen-activated protein kinase signaling pathway. Mol. Med. Rep. 2019, 20, 3027–3034. [Google Scholar] [CrossRef]
- Mishra, S.; Palanivelu, K. Thread Rating. Ann. Indian Acad. Neurol. 2008, 11, 13–19. [Google Scholar] [CrossRef]
- Zorena, K.; Jachimowicz-Duda, O.; Ślęzak, D.; Robakowska, M.; Mrugacz, M. Adipokines and obesity. Potential link to metabolic disorders and chronic complications. Int. J. Mol. Sci. 2020, 21, 3570. [Google Scholar] [CrossRef]
- Achari, A.E.; Jain, S.K. Adiponectin, a therapeutic target for obesity, diabetes, and endothelial dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef] [PubMed]
- Weidinger, C.; Ziegler, J.F.; Letizia, M.; Schmidt, F.; Siegmund, B. Adipokines and their role in intestinal inflammation. Front. Immunol. 2018, 9, 1974. [Google Scholar] [CrossRef] [PubMed]
- Kelesidis, T.; Kelesidis, I.; Chou, S.; Mantzoros, C.S. Narrative review: The role of leptin in human physiology: Emerging clinical applications. Ann. Intern. Med. 2010, 152, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Iikuni, N.; Kwan Lam, Q.L.; Lu, L.; Matarese, G.; Cava, A.L. Leptin and inflammation. Curr. Immunol. Rev. 2008, 4, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, V.; Tuli, H.S.; Tania, M.; Srivastava, S.; Ritzer, E.E.; Pandey, A.; Aggarwal, D.; Barwal, T.S.; Jain, A.; Kaur, G. Molecular mechanisms of action of epigallocatechin gallate in cancer: Recent trends and advancement. Semin. Cancer Biol. 2022, 80, 256–275. [Google Scholar] [CrossRef]
- Musial, C.; Kuban-Jankowska, A.; Gorska-Ponikowska, M. Beneficial properties of green tea catechins. Int. J. Mol. Sci. 2020, 21, 1744. [Google Scholar] [CrossRef]
- Kim, J.S.; Kim, J.-M.; Jeong-Ja, O.; Jeon, B.S. Inhibition of inducible nitric oxide synthase expression and cell death by (−)-epigallocatechin-3-gallate, a green tea catechin, in the 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine mouse model of Parkinson’s disease. J. Clin. Neurosci. 2010, 17, 1165–1168. [Google Scholar] [CrossRef]
- Kim, M.; Murakami, A.; Miyamoto, S.; Tanaka, T.; Ohigashi, H. The modifying effects of green tea polyphenols on acute colitis and inflammation-associated colon carcinogenesis in male ICR mice. Biofactors 2010, 36, 43–51. [Google Scholar] [CrossRef]
- De Oliveira, G.A.; Cheng, R.Y.; Ridnour, L.A.; Basudhar, D.; Somasundaram, V.; McVicar, D.W.; Monteiro, H.P.; Wink, D.A. Inducible nitric oxide synthase in the carcinogenesis of gastrointestinal cancers. Antioxid. Redox Signal. 2017, 26, 1059–1077. [Google Scholar] [CrossRef]
- Cianchi, F.; Cortesini, C.; Fantappiè, O.; Messerini, L.; Schiavone, N.; Vannacci, A.; Nistri, S.; Sardi, I.; Baroni, G.; Marzocca, C. Inducible nitric oxide synthase expression in human colorectal cancer: Correlation with tumor angiogenesis. Am. J. Pathol. 2003, 162, 793–801. [Google Scholar] [CrossRef]
- Mandal, P. Molecular signature of nitric oxide on major cancer hallmarks of colorectal carcinoma. Inflammopharmacology 2018, 26, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.; Sun, H.; Yu, F.-B.; Li, B.; Zhang, Y.; Zhu, Y.-T. The role of cyclooxygenase-2 in colorectal cancer. Int. J. Med. Sci. 2020, 17, 1095. [Google Scholar] [CrossRef] [PubMed]
- Greenhough, A.; Smartt, H.J.; Moore, A.E.; Roberts, H.R.; Williams, A.C.; Paraskeva, C.; Kaidi, A. The COX-2/PGE 2 pathway: Key roles in the hallmarks of cancer and adaptation to the tumour microenvironment. Carcinogenesis 2009, 30, 377–386. [Google Scholar] [CrossRef]
- Tuli, H.S.; Tuorkey, M.J.; Thakral, F.; Sak, K.; Kumar, M.; Sharma, A.K.; Sharma, U.; Jain, A.; Aggarwal, V.; Bishayee, A. Molecular mechanisms of action of genistein in cancer: Recent advances. Front. Pharmacol. 2019, 10, 1336. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Gu, J.; Wu, Y.; Liang, P.; Shen, M.; Xi, J.; Qin, J. Clinical characteristics of colorectal cancer patients and anti-neoplasm activity of genistein. Biomed. Pharmacother. 2020, 124, 109835. [Google Scholar] [CrossRef]
- Pintova, S.; Dharmupari, S.; Moshier, E.; Zubizarreta, N.; Ang, C.; Holcombe, R.F. Genistein combined with FOLFOX or FOLFOX–Bevacizumab for the treatment of metastatic colorectal cancer: Phase I/II pilot study. Cancer Chemother. Pharmacol. 2019, 84, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Felice, M.R.; Maugeri, A.; De Sarro, G.; Navarra, M.; Barreca, D. Molecular Pathways Involved in the Anti-Cancer Activity of Flavonols: A Focus on Myricetin and Kaempferol. Int. J. Mol. Sci. 2022, 23, 4411. [Google Scholar] [CrossRef]
- Lin, Y.; Shi, R.; Wang, X.; Shen, H.-M. Luteolin, a flavonoid with potential for cancer prevention and therapy. Curr. Cancer Drug Targets 2008, 8, 634–646. [Google Scholar] [CrossRef]
- Ganai, S.A.; Sheikh, F.A.; Baba, Z.A.; Mir, M.A.; Mantoo, M.A.; Yatoo, M.A. Anticancer activity of the plant flavonoid luteolin against preclinical models of various cancers and insights on different signalling mechanisms modulated. Phytother. Res. 2021, 35, 3509–3532. [Google Scholar] [CrossRef]
- Xiao, Y.; Yang, H.; Lu, J.; Li, D.; Xu, C.; Risch, H.A. Serum gamma-glutamyltransferase and the overall survival of metastatic pancreatic cancer. BMC Cancer 2019, 19, 1–7. [Google Scholar] [CrossRef]
- Ribatti, D.; Crivellato, E. Mast cells, angiogenesis, and tumour growth. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2012, 1822, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-X.; Chen, Y.-R.; Liu, S.-S.; Ye, Y.-P.; Jiao, H.-L.; Wang, S.-Y.; Xiao, Z.-Y.; Wei, W.-T.; Qiu, J.-F.; Liang, L. MiR-384 inhibits human colorectal cancer metastasis by targeting KRAS and CDC42. Oncotarget 2016, 7, 84826. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Bai, P.-S.; Nan, K.-J.; Sun, H.; Chen, N.-Z.; Qi, X.-G. Pleiotrophin is a potential colorectal cancer prognostic factor that promotes VEGF expression and induces angiogenesis in colorectal cancer. Int. J. Color. Dis. 2012, 27, 287–298. [Google Scholar] [CrossRef]
- Zhu, M.-L.; Zhang, P.-M.; Jiang, M.; Yu, S.-W.; Wang, L. Myricetin induces apoptosis and autophagy by inhibiting PI3K/Akt/mTOR signalling in human colon cancer cells. BMC Complement. Med. Ther. 2020, 20, 1–9. [Google Scholar] [CrossRef]
- Rauf, A.; Shariati, M.A.; Imran, M.; Bashir, K.; Khan, S.A.; Mitra, S.; Emran, T.B.; Badalova, K.; Uddin, M.; Mubarak, M.S. Comprehensive review on naringenin and naringin polyphenols as a potent anticancer agent. Environ. Sci. Pollut. Res. 2022, 29, 31025–31041. [Google Scholar] [CrossRef] [PubMed]
- Ghanbari-Movahed, M.; Jackson, G.; Farzaei, M.H.; Bishayee, A. A Systematic Review of the Preventive and Therapeutic Effects of Naringin against Human Malignancies. Front. Pharmacol. 2021, 12, 250. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Wang, L.; Yang, Y.; Ni, J.; Zhao, L.; Dong, S.; Guo, M. Simultaneous determination of oleanolic acid, ursolic acid, quercetin and apigenin in Swertia mussotii Franch by capillary zone electrophoresis with running buffer modifier. Biomed. Chromatogr. 2015, 29, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Niaz, K.; Maqbool, F.; Ismail Hassan, F.; Abdollahi, M.; Nagulapalli Venkata, K.C.; Nabavi, S.M.; Bishayee, A. Molecular targets underlying the anticancer effects of quercetin: An update. Nutrients 2016, 8, 529. [Google Scholar] [CrossRef]
- Terzić, J.; Grivennikov, S.; Karin, E.; Karin, M. Inflammation and colon cancer. Gastroenterology 2010, 138, 2101–2114.e2105. [Google Scholar] [CrossRef]
- Long, A.G.; Lundsmith, E.T.; Hamilton, K.E. Inflammation and colorectal cancer. Curr. Color. Cancer Rep. 2017, 13, 341–351. [Google Scholar] [CrossRef]
- Nouri, Z.; Fakhri, S.; Nouri, K.; Wallace, C.E.; Farzaei, M.H.; Bishayee, A. Targeting multiple signaling pathways in cancer: The rutin therapeutic approach. Cancers 2020, 12, 2276. [Google Scholar] [CrossRef] [PubMed]
- Chua, L.S. A review on plant-based rutin extraction methods and its pharmacological activities. J. Ethnopharmacol. 2013, 150, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Negahdari, R.; Bohlouli, S.; Sharifi, S.; Maleki Dizaj, S.; Rahbar Saadat, Y.; Khezri, K.; Jafari, S.; Ahmadian, E.; Gorbani Jahandizi, N.; Raeesi, S. Therapeutic benefits of rutin and its nanoformulations. Phytother. Res. 2021, 35, 1719–1738. [Google Scholar] [CrossRef]
- Pan, M.-H.; Chen, W.-J.; Lin-Shiau, S.-Y.; Ho, C.-T.; Lin, J.-K. Tangeretin induces cell-cycle G1 arrest through inhibiting cyclin-dependent kinases 2 and 4 activities as well as elevating Cdk inhibitors p21 and p27 in human colorectal carcinoma cells. Carcinogenesis 2002, 23, 1677–1684. [Google Scholar] [CrossRef]
- He, L.; Lu, N.; Dai, Q.; Zhao, Y.; Zhao, L.; Wang, H.; Li, Z.; You, Q.; Guo, Q. Wogonin induced G1 cell cycle arrest by regulating Wnt/β-catenin signaling pathway and inactivating CDK8 in human colorectal cancer carcinoma cells. Toxicology 2013, 312, 36–47. [Google Scholar] [CrossRef]
- Tan, H.; Li, X.; Yang, W.-H.; Kang, Y. A flavone, Wogonin from Scutellaria baicalensis inhibits the proliferation of human colorectal cancer cells by inducing of autophagy, apoptosis and G2/M cell cycle arrest via modulating the PI3K/AKT and STAT3 signalling pathways. J. BUON 2019, 24, 1143–1149. [Google Scholar] [PubMed]
- Alam, M.; Ahmed, S.; Elasbali, A.M.; Adnan, M.; Alam, S.; Hassan, M.I.; Pasupuleti, V.R. Therapeutic implications of caffeic acid in cancer and neurological diseases. Front. Oncol. 2022, 12, 860508. [Google Scholar] [CrossRef]
- Banerjee, N.; Kim, H.; Krenek, K.; Talcott, S.T.; Mertens-Talcott, S.U. Mango polyphenolics suppressed tumor growth in breast cancer xenografts in mice: Role of the PI3K/AKT pathway and associated microRNAs. Nutr. Res. 2015, 35, 744–751. [Google Scholar] [CrossRef]
- Boghossian, S.; Hawash, A. Chemoprevention in colorectal cancer-where we stand and what we have learned from twenty year’s experience. Surgeon 2012, 10, 43–52. [Google Scholar] [CrossRef]
- Ko, J.-H.; Sethi, G.; Um, J.-Y.; Shanmugam, M.K.; Arfuso, F.; Kumar, A.P.; Bishayee, A.; Ahn, K.S. The role of resveratrol in cancer therapy. Int. J. Mol. Sci. 2017, 18, 2589. [Google Scholar] [CrossRef]
- Bishayee, A. Cancer Prevention and Treatment with Resveratrol: From Rodent Studies to Clinical TrialsResveratrol and Cancer: In vivo and Clinical Studies. Cancer Prev. Res. 2009, 2, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Arslan, A.K.K.; Uzunhisarcıklı, E.; Yerer, M.B.; Bishayee, A. The golden spice curcumin in cancer: A perspective on finalized clinical trials during the last 10 years. J. Cancer Res. Ther. 2022, 18, 19–26. [Google Scholar]
- Singh, A.P.; Singh, R.; Verma, S.S.; Rai, V.; Kaschula, C.H.; Maiti, P.; Gupta, S.C. Health benefits of resveratrol: Evidence from clinical studies. Med. Res. Rev. 2019, 39, 1851–1891. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.-Y.; Zhai, J.; Huan, X.-K.; Xu, W.-W.; Tian, J.; Farhood, B. A Systematic Review of the Therapeutic Potential of Resveratrol During Colorectal Cancer Chemotherapy. Mini Rev. Med. Chem. 2022. [Google Scholar] [CrossRef]
- Ma, Z.; Wang, N.; He, H.; Tang, X. Pharmaceutical strategies of improving oral systemic bioavailability of curcumin for clinical application. J. Control. Release 2019, 316, 359–380. [Google Scholar] [CrossRef]
- Seufferlein, T.; Ettrich, T.J.; Menzler, S.; Messmann, H.; Kleber, G.; Zipprich, A.; Frank-Gleich, S.; Algül, H.; Metter, K.; Odemar, F. Green tea extract to prevent colorectal adenomas, results of a randomized, placebo-controlled clinical trial. Off. J. Am. Coll. Gastroenterol. ACG 2022, 117, 884–894. [Google Scholar] [CrossRef]
- Farsad-Naeimi, A.; Alizadeh, M.; Esfahani, A.; Aminabad, E.D. Effect of fisetin supplementation on inflammatory factors and matrix metalloproteinase enzymes in colorectal cancer patients. Food Funct. 2018, 9, 2025–2031. [Google Scholar] [CrossRef]
- Ganesan, K.; Jayachandran, M.; Xu, B. Diet-derived phytochemicals targeting colon cancer stem cells and microbiota in colorectal cancer. Int. J. Mol. Sci. 2020, 21, 3976. [Google Scholar] [CrossRef]
- Greiner, A.K.; Papineni, R.V.; Umar, S. Chemoprevention in gastrointestinal physiology and disease. Natural products and microbiome. Am. J. Physiol.-Gastrointest. Liver Physiol. 2014, 307, G1–G15. [Google Scholar] [CrossRef]
- Dacrema, M.; Ali, A.; Ullah, H.; Khan, A.; Di Minno, A.; Xiao, J.; Martins, A.M.C.; Daglia, M. Spice-Derived Bioactive Compounds Confer Colorectal Cancer Prevention via Modulation of Gut Microbiota. Cancers 2022, 14, 5682. [Google Scholar] [CrossRef]
- O’keefe, S.J. Diet, microorganisms and their metabolites, and colon cancer. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 691–706. [Google Scholar] [CrossRef] [PubMed]









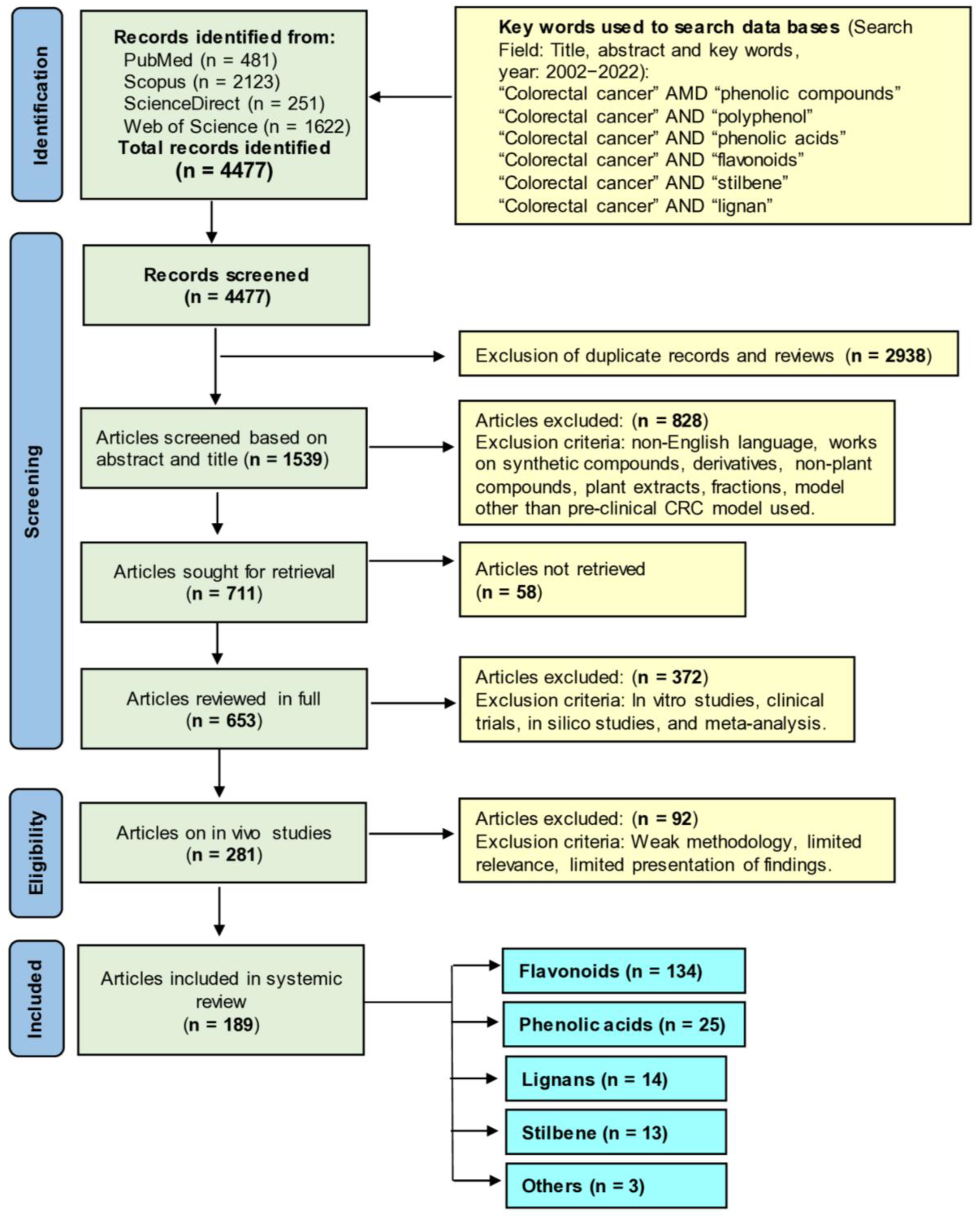{kind=link}
{kind=link}
{kind=link}
{kind=link}
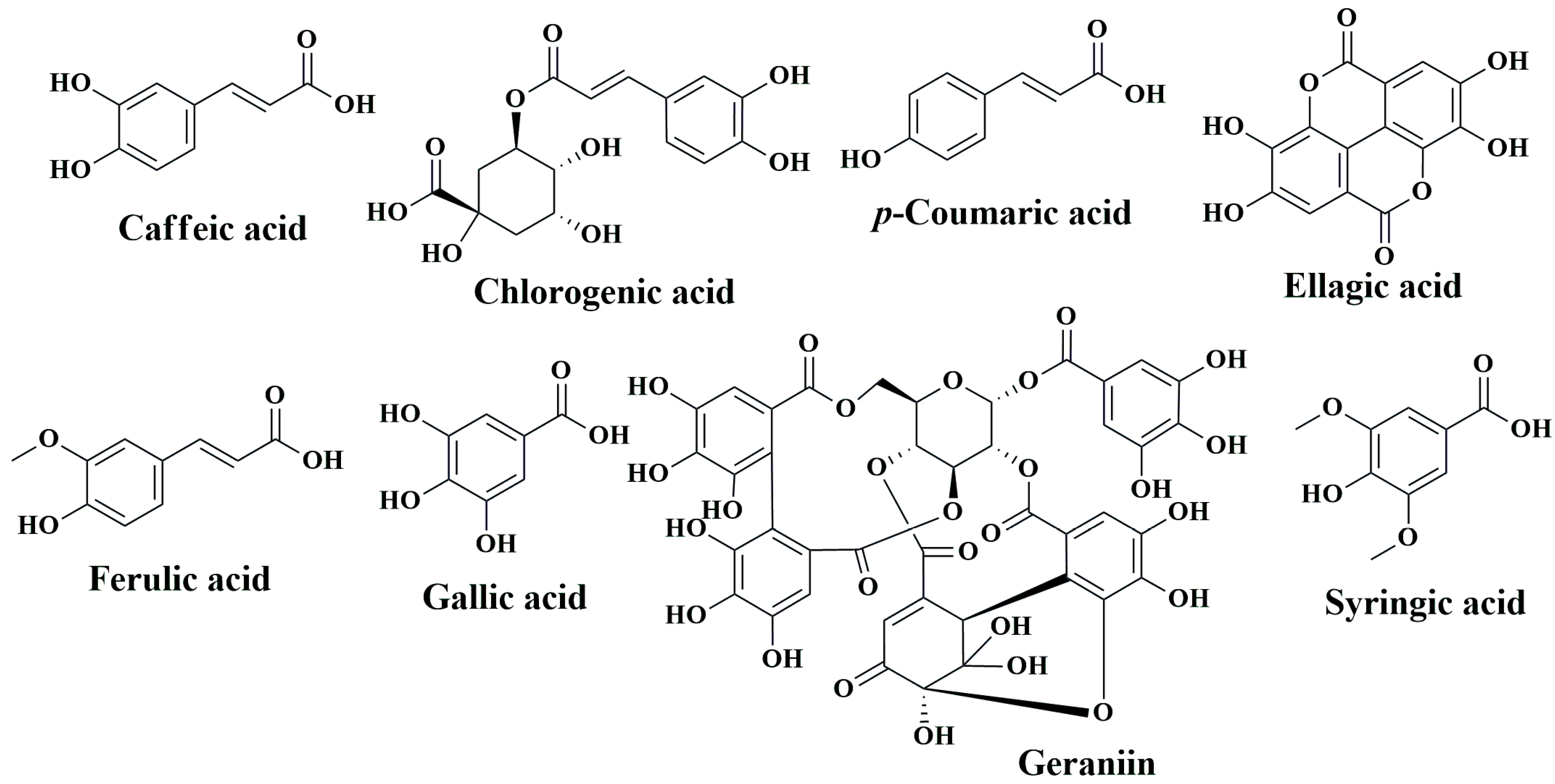{kind=link}
{kind=link}
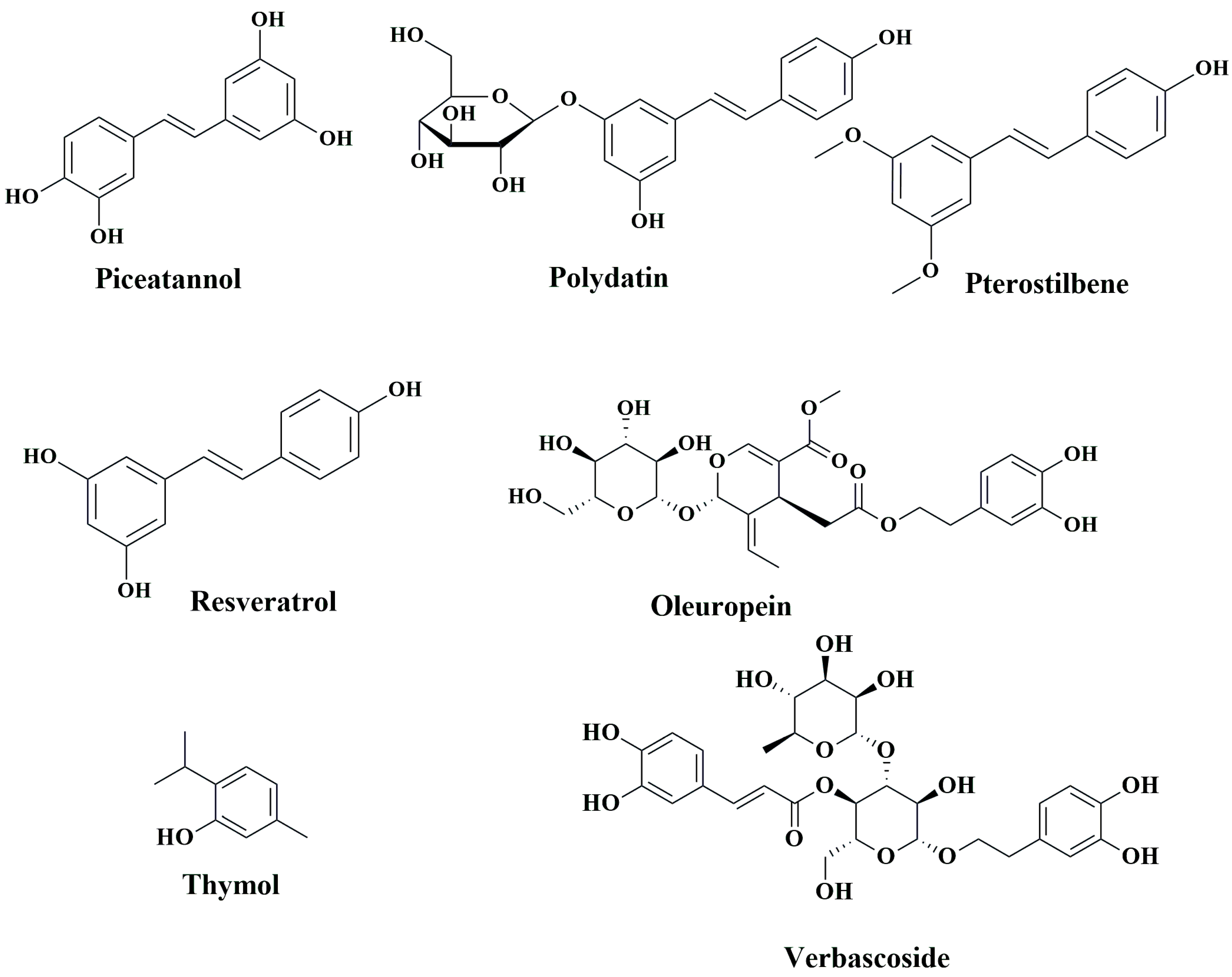{kind=link}
{kind=link}
| Syndrome | Genetic Defects | Clinical Manifestations | References |
|---|---|---|---|
| Hereditary nonpolyposis cancer syndromes | |||
| Lynch syndrome | MLH1, MSH2, MSH6, MSH3, and PMS2 | Increased risk for CRC, (10–47%) depending on gene mutated; asymptomatic unless altered bowel habits, GI bleeding due to tumors/polyps occurs; increased risk for endometrial cancer; extracolonic manifestations are associated as Muir-Torre, Turcot. | [44,46,47] |
| Muir-Torre syndrome (HNPCC + Sebaceous gland malignancies) | MLH1, MSH2, MSH6, and PMS2 | Sebaceous skin tumor/keratoacanthoma and Lynch syndrome features. | [48,49] |
| Turcot syndrome type 1 (HNPCC with primary brain tumors) | MMR, MLH1, and PMS2 | Features of Lynch syndrome + primary brain tumors. | [50,51,52,53] |
| Hereditary polyposis colorectal cancers | |||
| Familial adenomatous polyposis (FAP) syndrome | APC | More than colorectal adenomatous polyps; 100% cancer risk | [50,54] |
| Turcot syndrome type II (FAP with Primary Brain tumors) | APC | FAP syndrome + primary brain tumors, medulloblastoma, glioblastoma, astrocytoma. | [50,51,52,53] |
| Gardner syndrome | APC | FAP syndrome+ extraintestinal manifestations of desmoid tumors; sebaceous cysts; osteomas of mandible, skull, fibromatosis, congenital hypertrophy of retinal pigment epithelium (CHRPE); adrenal adenomas. | [55,56] |
| Adenomatous polyposis syndromes | APC and MUTYH | Increased number of colorectal adenomas (10–100 s), serrated polyposis, mixed polyps; duodenal adenomas are common; 43–33% increased risk of CRC; increased thyroid nodules, adrenal lesions, jawbone cysts. | [50,57,58,59] |
| Juvenile polyposis coli | BMPR1A and SMAD4 | Multiple hamartomatous polyps in the GI tract- mainly colorectum; rectal bleeding due to polyps is a common presenting symptom; anemia due to bleeding is common; extracolonic manifestations hereditary hemorrhagic Telangiectasia (HHT) telangiectasias of buccal mucosa and skin, epistaxis, and anemia, with AV malformations; colorectal cancer risk 38.7% increased. | [60,61,62] |
| Peutz-Jeghers syndrome | STK11 | Mucocutaneous pigmentation; hamartomatous polyps; 39% increased risk for CRC. | [63,64] |
| Cowden syndrome (multiple hamartomasyndrome) | PTEN | Mucocutaneous lesions and macrocephaly; skin manifestations; uterine leiomyomas, ovarian cysts; multiple hamartomas on any organ; increased risk of breast, thyroid, renal, endometrial, and colorectal cancer; 9–16% risk of CRC.; increased risk for malignant melanomas; specific dysplastic gangliocytoma of the cerebellum; Lhermitte-Duclos disease is specific to Cowden disease. | [65,66] |
| Drugs | Study Design | Mechanism | Main Findings | References |
|---|---|---|---|---|
| Aspirin | Meta-analysis | COX-2 inhibition | There was a dose-dependent reduction in the risk of CR by aspirin. An aspirin dose of 75–100 mg/day reduced the risk by 10%, and 325 mg/day reduced the risk by 35% (Meta-analysis of 45 studies [RR = 0.73, 95% confidence interval (CI) 0.69–0.78]) | [103,104,105,106] |
| Non-aspirin NSAIDS | Meta-analysis | COX-2 inhibition | Data from 23 studies suggested using higher doses of non-aspirin NSAIDs in the general population aged 40 years or older reduced CRC risk, specifically for white women, for distal colon cancer. (Pooled ODDs ratio was 0.74 (0.67–0.81), I2 = 75.9%, p < 0.001.) | [107] |
| Sulindac+ DFMO | RCT | Sulindac inhibits COX-2 DFMO- irreversibly inhibits Ornithine decarboxylase (polyamine synthesis) | Significant reduction of recurrent adenomas (12 vs. 41%, risk ratio 0.30), advanced adenomas (0.7 vs. 8.5%, risk ratio 0.09), and multiple adenomas (0.7 vs. 13.2%, risk ratio 0.06) | [108,109] |
| DFMO + Aspirin | RCT | Aspirin inhibits COX-2 DFMO inhibits polyamine synthesis Both combined may have a synergistic action. | After one year of treatment, in the DFMO + aspirin arm vs. placebo, there was a significant reduction in rectal aberrant crypt foci (precursor of rectal carcinoma). (74% vs. 45%, p = 0.020). No statistically significant reduction of colorectal adenomas was observed. | [110] |
| Erlotinib + Sulindac | RCT | Erlotinib is an EGFR inhibitor; sulindac is a COX-2 inhibitor. | In 82 patients of familial adenomatous polyposis, Sulindac + Erlotinib was associated with a 69.4% decrease in those with an intact colorectum compared with placebo (95% CI, 28.8−109.2%; p = 0.009) | [111] |
| Celecoxib | Meta-analysis | Selective COX-2 inhibitor, more specific for inflammation, with fewer GI side effects. Celecoxib has higher cardiovascular mortality | 3 RCTs (involving 4420 patients) and 3 post-trial studies (2159) showed a significant reduction in the incidence of adenoma RR (0.67 [95% CI, 0.62–0.72] compared with placebo). There was an increased risk of cardiovascular mortality with twice dosing 400 mg celecoxib (RR 3.42 [95% CI, 1.56–7.46]). Once-a-day dosing did not show an increased CV risk. (1.01 [95% CI, 0.70–1.46]). | [112] |
| Clopidogrel | Case-control Study | Clopidogrel inhibits platelet aggregation via irreversible inhibition of the P2Y12 receptor | Clopidogrel decreased CRC risk in patients receiving treatment >1 year. (0.65% AOR; 95% CI, 0.55–0.78). Dual antiplatelet therapy (Clopidogrel aspirin) had the same effect as either drug is taken as monotherapy. | [113] |
| Metformin | Meta-analysis | Activates AMPK, inhibits mTOR pathway | Metformin users had a significantly lower incidence. CRC (RR 0.76, CI 0.69–0.84, p < 0.001) compared with non-metformin users. Further analysis on the overall survival of metastatic CRC patients revealed significantly higher survival rates in metformin users (HR 0.77, CI 0.68–0.87, p < 0.001). | [114] |
| UCDA | Cohort Study | Has antioxidant action. Prevents NF-κB and AP1 activity. Inhibits c-Myc | Chronic liver disease patients with UCDA have a reduced risk of colorectal cancer. UDCA use was associated with a reduced risk of CRC (hazard ratio, 0.60; 95% confidence interval [CI], 0.39–0.92). | [115] |
| Statin | Meta-analysis | 3-HMGCOA reductase inhibitor decreases cholesterol synthesis. Antioxidant activity; shows pro-apoptotic effects on human CRC lines. Anti-inflammatory properties | 14 studies involving 130,994 patients. In terms of post-diagnosis statin uses, the pooled HR of all-cause mortality was 0.86 (95% CI, 0.76–0.98), and the pooled HR of CSM was 0.79 (95%CI, 0.70–0.89) (Cancer-Specific Mortality). | [116,117] |
| Menopausal hormone therapy (combined estrogen-progestin) | Nationwide Cohort Study (Norway) | Estrogens have been proposed to alter bile acid composition, modulate colonic transit. Decrease production of mitogenic insulin-like growth factor | The current use of postmenopausal hormone therapy was associated with a decreased CRC risk. RR (for combined estrogen-progestin therapy) in oral formulations was 0.86 (95% CI 0.71 to 1.05) | [118] |
| Bisphosphonates | Meta-analysis | Inhibits osteoclastic bone resorption, Anti-apoptotic effect | Meta-analysis of 34 studies and 4,508,261 participants. There was a significant reduction in the risk of CRC. (RR = 0.89, 95% CI: 0.81–0.98) | [119] |
| Phytocompound | Source | Animal Model Studied | Dose and Route of Administration | Mode of Action | Reference |
|---|---|---|---|---|---|
| Flavonoids | |||||
| 2,3,5,4′-tetrahydroxystilbene-2-O-β-D-glucoside | Polygonum multiflorum Thunb | AOM-induced colon carcinogenesis in male F144 rats | Oral administration, 30, 150, 250 mg/kg | Decreased the number ofACF by 47–54%; suppressed tumor growth; downregulated NF-κB in nucleus and cytoplasm; downregulated CEA | [170] |
| 4′-hydroxychalcone | Herb, teas, and spices | APCMin/+ mice | Oral administration, 10 mg/kg | Reduced the incidences and size of adenomas; induced apoptosis; suppressed proliferation of polyps; downregulated Ki-67; downregulated c-Myc, Axin2 and CD44 gene expression | [171] |
| Aciculatin | Chrysopogon aciculatus | HCT116 induced tumor xenograft SCID mice | Intraperitoneal injection, 30 mg/kg | Suppressed tumor growth without losing weight; upregulated the expression of p53 and downregulated the expression of Ki-67; induced apoptosis; arrested cells in sub G1 phase | [172] |
| Apigenin | Parsley, wheat, onions, apples, and tea plants | AOM-induced CF-1 mice and Min mice carrying mutant APC gene | Oral administration of 0.1% dietary apigenin | Reduced ACF formation and ODC activity | [173] |
| Male BALB/c-nu mice | Intraperitoneal injection, 20 mg/kg | Induced apoptosis of CRC cells; upregulated FADD expression and its phosphorylation | [174] | ||
| Male BALB/c-nu mice injected with SW480 cells | Route of administration not reported, 50 mg/kg | Elevated transgelin and downregulation of MMP-9 expression via reducing Akt phosphorylation at Ser473 and Thr308 | [175] | ||
| APCMin/+ mice | Oral gavage, 25 and 50 mg/kg | Reduced the number of polyps; induced of p53 activity | [176] | ||
| Nude BALB/c mice injected with HT-29 cells | Subcutaneous injection, 35 mg/kg | Induced apoptosis; induced autophagy through inhibition mTOR/PI3K/Akt signaling pathway | [177] | ||
| SCID mice | Oral gavage, 25 mg/kg | Suppressed prosurvival regulators Mcl-1, Akt, and ERK | [178] | ||
| NEDD9 knock downed DLD1 cells mediated metastasis model in female athymic nude mice | Intraperitoneal injection, 20 mg/kg | Suppressed invasion, migration, and metastasis by downregulating overexpressed Neural precursor cells expressed NEDD9 | [179] | ||
| Baicalein | Scutellaria baicalensis Georgi | AMO and DSS induced colon tumor in male ICR mice | Oral administration, 1,5 and 10 mg/kg | Restored colon length; reduced tissue inflammation. | [180] |
| SW620 xenograft in BALB/c nude mice | Intraperitoneal injection, 50 mg/kg | Suppressed tumor growth by 55% without losing body weight | [181] | ||
| CT-26 derived tumor in female BALB/c mice | Intraperitoneal injection, 20 and 40 mg/kg | Reduced tumor growth rate; downregulated TLR4 and p-IκBα protein expression; inhibited NF-κB | [182] | ||
| HT-29 cell-induced tumor xenograft in male nude mice | Oral administration, 10 mg/kg | Suppressed tumor growth by 29.33% compared to the control group; induced apoptosis; upregulated p53 and p21 | [183] | ||
| DLD-1 tumor xenograft in BALB/c athymic nude mice | Intragastric administration, 20 mg/kg | Suppression of tumor growth; inhibition of ERK phosphorylation; downregulation of MMP-2 and MMP-9 | [184] | ||
| HCT116 tumor xenograft in NSG immunodeficient mice | Intraperitoneal injection, 50 mg/kg | Suppressed tumorigenesis; inhibited colon cancer growth; induced apoptosis and senescence | [185] | ||
| HCT116 tumor xenograft in athymic BALB/c nude mice | Intraperitoneal injection, 80 mg/kg | Suppressed tumor growth; induced senescence; upregulated DEPP; activated Ras/Raf/MEK/ERK pathway | [186] | ||
| HT-29 tumor xenograft in nude mice | Intraperitoneal injection, 50 and 100 mg/kg | Suppressed tumor growth | [187] | ||
| HCT116 tumor xenograft in athymic BALB/c nude mice | Intraperitoneal injection, 100 and 200 mg/kg | Suppressed tumor growth; induced apoptosis; suppressed cancer stem cells; inhibited EMT and cyclin D1 | [188] | ||
| APCMin/+ mice | Oral administration, 30 mg/kg | Reduced tumor numbers; suppressed IL-1β, IL-2, IL-6, and IL-10 | [189] | ||
| HCT116 tumor xenograft in male BALB/c nude mice | Intraperitoneal injection, 100 mg/kg | Suppressed tumor growth; decreased circMYH9, mir761 and HDGF | [190] | ||
| Boeravinone B | Boerhaavia diffusa | DMH-induced CRC in Swiss albino Wistar rats | Intraperitoneal injection, 20 and 40 mg/kg | Decreased the number of tumor incidences; downregulated LPO; upregulated catalase, SOD and GSH; downregulated TNF-α, IL-1β, IL-6, COX-2, PGE2 and iNOS; upregulated levels of IL-4 and IL-10; down regulated MPO; downregulated the expression of GDI2 mRNA | [191] |
| Chrysin | Passiflora caerulea, Passiflora incarnata, Oroxylum indicum | AOM-induced ACF in male F344 rats | Dietary administration, 0.001% and 0.01% | Reduced mitotic index and increased apoptotic index; reduced the frequency of ACF | [192] |
| Male albino rats injected with DMH + DSS | Oral administration, 125 and 250 mg/kg | Reduced the level of CXCL1, AREG and MMP-9 | [193] | ||
| Curcumin | Curcuma longa | DSS-induced colitis in C57BL/6 mice | Oral consumption as dietary supplement, 0.6% | Reduced tumor incidences; inhibited nuclear translocation of β-catenin; downregulated TNF-α and interferon-γ; downregulated COX-2 and p53 | [194] |
| HCT116 tumor xenograft in female ICR SCID mice | Intragastric administration, 500 mg/kg | Suppressed tumor growth; inhibited proteasome; suppressed proliferation; induced apoptosis | [195] | ||
| AOM-DSS induced CRC in male C57BL/6 mice | Oral gavage, 500 mg/kg | Reduced CRC tumor number; downregulated IL-1β, IL-6, COX-2 and β-catenin; suppressed Axin2 by inhibiting Wnt/β-catenin pathway | [196] | ||
| AOM-induced colonic preneoplastic lesion in C57BL/KsJ-db/db obese mice | Dietary supplement, 0.2% and 2.0% | Inhibited colonic premalignant lesion | [197] | ||
| HCT116 tumor xenograft in athymic nu/nu nude mice | Oral administration, 1 g/kg | Enhanced the efficacy of radiation therapy; suppressed NF-κB activity and expression | [198] | ||
| Colo205 and LoVo tumor xenografts in athymic nu/nu mice | Tail vein injection, 40 mg/kg | Inhibited tumor growth; suppressed angiogenesis | [199] | ||
| AOM-induced colon carcinogenesis in Il10−/− mice | Oral administration, 1% | Reduced colon tumors | [200] | ||
| AOM/DSS-induced colitis in C5757BL/6 mice | Oral administration, 25 mg/kg | Suppressed colitis-associated colon cancer and reduced tumor number | [201] | ||
| Cyanidin | Blackberries (Rubus fruticosus) | ApcMin/+ mice | Dietary supplementation, 0.03%, 0.1% or 0.3% | Reduced adenoma counts | [202] |
| Daidzein | Soybeans and soy-based products, and nuts | Male albino rats injected with DMH + DSS | Oral administration, 5 and 10 mg/kg | Reduced the level of CXCL1, AREG and MMP-9 | [193] |
| Delphinidin | Berries, pomegranates, eggplant, roselle, and wine | Male BALB/c nude mice xenograft with luciferase-transfected DLD-1 cells | Intraperitoneal injection, 100 μM | Suppressed integrin/FAK nexus; elevated miR-204–3p levels | [203] |
| Diosmetin | Chamomile, parsley, rosemary, rooibos tea, green tea, and other plants fof the mint and citrus family (Lamiaceae) | NCr nu/nu nude mice injected with HCT-116 cells | Oral administration, 50 and 100 mg/kg | Downregulated Bcl-2; upregulated Bax | [204] |
| EGCG | Camellia sinensis L. Ktze | SW837 xenograft in male BALB/c nude mice | Oral administration, 0.01% and 0.1% | Reduced tumor growth; inhibited phosphorylation of VEGFR-2, Akt and ERK | [205] |
| AOM-induced colonic premalignant lesions C57BL/KsJ-db/db mice | Oral administration, 0.01% and 0.1% | Decreased p-IGF-IR, p-GSK-3β, β-catenin, COX-2 and cyclin D1 in colonic mucosa; reduced IGF-I, insulin, triglyceride, cholesterol and leptin in serum | [206] | ||
| AOM-induced colonic carcinogenesis in ICR mice | Oral administration, 0.25% and 0.5% | Inhibited large ACF formation; inhibited iNOS and COX-2 | [207] | ||
| HCT116-SDCSCs tumor xenograft in athymic nude mice | Cells were pretreated, 100 μM | Suppressed tumor formation; downregulated Notch1, Bmi1, Suz12, and Ezh1; upregulated miR-34a, miR-145 and miR-200c | [208] | ||
| DMH-induced colon carcinogenesis in Wister rats | Oral administration, 0.2% | Inhibited ACF and induced apoptosis | [209] | ||
| DMH-induced CRC in male Wistar rats | Oral administration, 50, 100 and 200 mg/kg | Lowered ACF formation; reduced tumor volume | [210] | ||
| Eriodictyol | Eriodictyon californicum | DMH-induced colon carcinogenesis in male albino Wistar rats | Intragastrical administration, 200 mg/kg | Suppressed the number of polyps, ACF and lipid peroxidation levels; upregulated catalase, SOD, GPX, GST, GSH and GR | [211] |
| Euxanthone | Polygala caudata | HT-29 cells induced tumor in BALB/c nude mice | Intraperitoneal injection, 20 and 40 mg/kg | Suppressed tumor growth; induced apoptosis; upregulated Bax; downregulated Bcl-2; induced caspase-3 cleavage; downregulated CIP2A expression and upregulated PP2A | [212] |
| Fisetin | Strawberry, apple, persimmon, grapes, onion, and cucumber | AOM and DSS induced CAC in male BALB/c mice | Oral administration, 20 mg/kg | Suppressed dysplastic lesions; induced apoptosis in colonic tissue; downregulated Bcl-2 and STAT3 | [213] |
| FC1 mice, 3K1 mice, ApcMin/+ males, 3K1ApcMin/+ mice, B6 congenic strain, B6 FC13K1ApcMin/+ mice | Intraperitoneal injection, 1 mg/animal | Upregulated AMPK phosphorylation; suppressed PI3K/Akt/mTOR signaling | [214] | ||
| Male athymic nude mice | Oral administration, 400 and 800 mg/kg | Induced apoptosis, caspase-8 and cyt.; inhibited IGF1R and Akt | [215] | ||
| CT-26 tumor in BALB/c nude mice | Subcutaneous injection, 5 mg/kg | Suppressed oncoprotein securin in p53-independent fashion | [216] | ||
| BALB/c mice | Tail vein injection, 50 mg/kg | Inhibited programmed cell death and angiogenesis | [217] | ||
| HCT116 tumor xenograft in mice NOD/Shi-scid-IL2R gamma (null) (NOG) | Intraperitoneal injection, 30, 60 and 120 mg/kg | Suppressed tumor growth in a dose-dependent manner | [218] | ||
| Flavone | Fruits and vegetables | DMM-induced colon carcinogenesis in C57BL/6J mice | Subcutaneous injection, 15 and 400 mg/kg | Suppressed ACF formation and multiplicity | [219] |
| Formononetin | Astragalus membranaceus | Female BALB/c-nu/nu mice injected with HCT-116 cells | Intraperitoneal injection, 20 mg/kg | Decreased VEGF, MMP-2 and MMP-9 levels | [220] |
| Furowanin A | Millettia pachycarpa Benth | HT-29 tumor xenograft in male athymic BALB/c nude mice | Intraperitoneal injection, 20 and 40 mg/kg | Suppressed tumor growth, induced apoptosis and autophagy; upregulated cleaved caspase-3, LC3BII, Beclin and p27; downregulated Ki-67, pSTAT3, Mcl-1, p62, and cyclin D | [221] |
| Genistein | Genista tinctoria | DMH-induced colon cancer in Wistar rats | Oral administration, 2.5 mg/kg | Regulated tumor microenvironment; upregulated SOD, CAT, GPx, GR, vitamin A, vitamin C, vitamin E and GSH; activated NRF2 and HO-1; reduced expression of CD133, CD44 and β-catenin | [222] |
| AOM-induced colon cancer in Sprague-Dawley rats | Dietary supplementation, 140 mg/kg | Suppressed the expression of cyclin-D1 and c-Myc; decreased expression of Wnt5a, Sfrp1, Sfrp2, and Sfrp5; downregulated Wnt/ β-catenin pathway | [223] | ||
| HCT116 tumor xenograft in athymic BALB/c mice | Oral administration, 75 mg/kg | Didn’t inhibit tumor growth; suppress metastasis; downregulated MMP-2 and EGFR3 | [224] | ||
| Genkwanin | Dried flower buds of Daphne genkwa | APCMin/+ mice | Oral administration, 12.5 and 25 mg/kg | Inducted host defense; reduced proinflammatory cytokine levels | [225] |
| AOM/DSS-induced C57BL/6J mice | Oral administration, 22.5 mg/kg | Suppressed colon cancer growth by triggering tumor cell death; inhibited of pro-inflammatory cytokines | [226] | ||
| Hesperidin | Citrus fruits | AOM-induced Swiss albino mice | Oral administration, 25 mg/kg | Inhibited NF-κB, iNOS and COX-2; reduced cellular oxidative indicators and improved antioxidant status | [227] |
| AOM-induced male Swiss albino mice | Oral administration, 25 mg/kg | Inhibited the constitutively active Aurora-A driven PI3K/Akt/GSK-3 and mTOR; activated autophagy | [228] | ||
| AOM-induced male F344 rats | Oral administration, 1000 ppm | Inhibited ACF formation; reduced colonic mucosal ODC activity and polyamine levels in the blood | [229] | ||
| DMH-induced CRC in albino rats | Oral administration, 25 mg/kg | Elevated the expression of Smad4 and activin A | [230] | ||
| Hinokiflavone | Selaginella tamariscina, Juniperus phoenicea, and Rhus succedanea | CT-26 tumor in female BALB/c mice | Intraperitoneal injection, 25 and 50 mg/kg | Suppressed tumor growth and proliferation; induced apoptosis; downregulated Ki-67 and MMP-9 | [231] |
| Icariside II | Epimedi Herba | SW620 tumor xenograft in nude BALB/c mice | Intraperitoneal injection, 25 mg/kg | Suppressed tumor growth; induced apoptosis | [232] |
| Icaritin | Epimedium sp. | HT-29 tumor xenograft in male nude mice | Oral gavage, 10 mg/kg | Suppressed tumor growth and volume | [233] |
| Isoangustone A | Glycyrrhiza sp. | SW480 tumor xenograft in male BALB/c nu/nu mice | Intraperitoneal injection, 10 mg/kg | Suppressed tumor growth; induced autophagic cell death; upregulated phosphorylation of AMPK, ACC and LC3B-1 and II levels | [234] |
| Isoliquiritigenin | Glycyrrhiza glabra | AOM/DSS-induced colon carcinogenesis in male BALB/c mice | Intragastrical administration, 3, 15 and 75 mg/kg | Suppressed tumorigenesis; inhibited macrophage polarization; upregulated TNF-α, INF-γ and IL-12; downregulated TGF-β, IL-10 and IL-1 and COX-2 | [235] |
| Glycyrrhiza uralensis Fisher | AOM-treated colon carcinogenesis in 344 rats | Oral administration, 100 ppm dietary supplementation | Suppressed ACF formation; induced apoptosis | [236] | |
| Isorhamnetin | Opuntia ficus-indica | HT-29 RFP xenograft in immunosuppressed mice | Oral administration, dose not reported | Elevated cleaved caspase-9, Hdac11, and Bai1 proteins | [237] |
| FVB/N mice treated with AOM/DSS | Oral administration, dietary supplement, dose not reported | Inhibited nuclear translocation of β-catenin and c-Src stimulation; activated CSK | [238] | ||
| Kaempferol | Apple, tea, broccoli, and grapefruit | DMH-induced colorectal carcinogenesis in male Wistar rats | Oral administration, 200 mg/kg | Restored CAT, SOD, and GPx | [239] |
| DMH-induced colon carcinoma in male Sprague Dawley rats | Oral administration, 200 mg/kg | Reduced multiple plaque lesions and preneoplastic lesions | [240] | ||
| DMH-induced colitis in Sprague-Dawley albino rats | Oral administration, 200 mg/kg | Reduced multiplicity of the ACF; downregulated COX-2 and PCNA | [241] | ||
| Luteolin | Celery, parsley, broccoli, onion leaves, carrots, peppers, cabbages, and tea | DMH-induced carcinogenesis in male Wistar rats | Subcutaneous injection, 0.2 mg/kg | Reduced the number of tumor polyps and colon polyploids; decreased COX-2 level in blood and colonic tissue | [242] |
| AOM-induced CRC in male BALB/c mice | Oral administration, 1.2 mg/kg | Reduced the levels of alkaline phosphatase and lactate dehydrogenase; suppressed iNOS and COX-2 | [243] | ||
| AOM-induced CRC in male BALB/c mice | Oral administration, 1.2 mg/kg | Reduced cytochrome b5, cytochrome P450 and cytochrome b5; enhanced the expression of UDP-GT and GST in colonic tissue; upregulated Nrf2 | [244] | ||
| CT-26 mediated lung metastasis | Oral administration, 10 and 50 mg/kg | Suppressed lung nodules and nodule volume; inhibited MMP-9 expression | [245] | ||
| AOM-induced colon carcinogenesis in BALB/c mice | Oral administration, 1.2 mg/kg | Inhibited MMP-2 and MMP-9; downregulated γ-glutamyl transferase, 5′ nucleotidase, cathepsin D, and carcinoembryonic antigen | [246] | ||
| HT-29 tumor xenograft in BALB/c nude mice | Intragastric administration, 100 mg/kg | Suppressed CRC metastasis; upregulated miR-384; downregulated pleiotrophin expression | [247] | ||
| HT-29 tumor xenograft in BALB/c nude mice | Intraperitoneal injection, 50 mg/kg | Inhibited tumor growth; induced apoptosis | [248] | ||
| Lysionotin | Lysionotus pauciflorus Maxim | HCT116 tumor xenograft in athymic nude mice | Intraperitoneal injection, 20 mg/kg | Suppressed tumor growth; induced ferroptosis | [249] |
| Magnolin | Magnolia biondii | HCT116 tumor xenograft in female BALB/c athymic nude mice | Intraperitoneal injection, 20 mg/kg | Suppressed tumor growth; downregulated LIF, STAT3 and Mcl-1 | [250] |
| Morin | Old fustic (Chlorophora tinctoria) and osage orange (Maclura pomifera) | Male athymic nude mice injected with HCT-116 cells | Intraperitoneal injection, 30 and 60 mg/kg | Inactivated NF-κB signaling | [251] |
| Male albino Wistar rats injected with DMH | Intraperitoneal injection, 30 and 60 mg/kg | Modulated tumor metabolism via β-cateinin/c-myc signaling, glycolysis and glutaminolysis pathways | [252] | ||
| Pirc rats (F344/NTac-Apc am1137) | Dietary supplementation, 50 mg/kg | Restored the sensitivity to apoptosis by inhibiting LMW-PTP | [253] | ||
| Male albino Wistar rats injected with DMH | Intragastric administration, 50 mg/kg | Reduced ACF formation; suppressed fecal and mucosal biotransformation enzymes | [254] | ||
| Male albino Wistar rats injected with DMH | Intragastric administration, 50 mg/kg | Inhibited NF-κB and inflammatory mediators; suppressed proapoptotic pathway | [255] | ||
| DMH-induced colon carcinogenesis in a male Wistar rats | Oral administration, 50 mg/kg | Reduced lipid hydroperoxides and CD; increased superoxide SOD, CAT, GST, GPx, GR; decreased GSH | [256] | ||
| Myricetin | Tea, barriers, fruits, vegetables | DMH-induced rat colon carcinogenesis | Dietary supplementation, 50, 100 and 200 mg/kg myricetin | Restored CAT, GPx and GSH | [257] |
| APCMin/+ C57BL/6J mice | Oral gavage, 100 mg/kg | Promoted apoptosis in adenomatous polyps; lowered IL-6 and PGE2; downregulated p38 MAPK/Akt/mTOR signaling pathway | [258] | ||
| AOM/DSS-induced in BALB/c mice | Oral gavage, 40 and 100 mg/kg | Inhibited the development of colorectal tumors and colorectal polyps; decreased the levels of TNF-, IL-1, IL-6, NF-κB, p-NF-κB, COX-2, PCNA, and cyclin D1 | [259] | ||
| AOM/DSS-induced colitis in C57BL/6 mice | Oral administration, 100 mg/kg | Decreased CSF/M-CSF, IL-6, and TNF-α in colonic mucosa; inhibited NF-κB/IL-6/STAT3 pathway | [260] | ||
| Naringenin | Oranges, lemons, and grapefruit | AOM-induced colon carcinogenesis in rats | Dietary supplement, 0.02% | Reduce the number of HMACF by 51% and the proliferative index by 32% | [261] |
| DSS-induced murine colitis model | Oral administration, 50 mg/kg | Decreased iNOS, ICAM-1, MCP-1, COX-2, TNF-α, and IL-6 transcript levels | [262] | ||
| HT-29 tumor xenograft in athymic NIH Swiss nude mice | Oral administration, 40 mg/kg | Suppressed tumor growth; inhibited COX-1 | [263] | ||
| Naringin | Oranges, lemons, and grapefruit | DMH-induced female Wistar rats | Oral gavage, 10, 100, 200 mg/kg | Reduced cell proliferation and tissue iron levels; upregulated antioxidant mineral levels | [264] |
| AOM/DSS-induce Male C57BL/6 mice | Oral gavage, 50 and 100 mg/kg | Suppressed ER stress-induced autophagy in colorectal mucosal cells | [265] | ||
| AOM-induced ACF in Sprague Dawley rats | Oral administration, 200 mg/kg | Reduced total number of ACF; suppressed proliferation; induced apoptosis; downregulated COX-2 and iNOS | [266] | ||
| Nobiletin | Peel of various Citrus fruits | AOM-DSS-induced colon carcinogenesis in male CD-1 mice | Oral, dietary supplement, 100 ppm | Reduced tumor incidences and multiplicity | [267] |
| Orientin | Ocimum sanctum | DMH-induced CRC in male Wister rats | Intraperitoneal injection, 10 mg/kg | Reduced NF-κB, TNF-α and IL-6; downregulated Ki-67 and PCNA; suppressed iNOS and COX-2 | [268] |
| DMH-induced CRC in male Wister rats | Intraperitoneal injection, 10 mg/kg | Suppressed ACF and crypt multiplicity; elevated the level of antioxidants; downregulated phase I enzymes and upregulated phase II enzymes | [269] | ||
| Oroxylin A | Scutellaria baicalensis | AOM-DSS induced CRC in C57BL/6 mice | Dietary supplementation, 50, 100 and 200 mg/kg | Suppressed tumor formation and colitis associated CRC; induced apoptosis; downregulated IL-6, IL-1β, p-STAT3, cyclin D, and Bcl-2; upregulated Bax | [270] |
| HCT116 tumor xenograft in male athymic BALB/c nude mice and AOM-DSS induced colon carcinogenesis in male C57BL/6 mice | Oral administration, 150 and 300 mg/kg | Suppressed carcinogenesis and primary colon cancer progression; reduced triglyceride; downregulated HIF1α, Srebp1, FASN, ADRP and FABP7; upregulated CPT1 | [271] | ||
| Pectolinarigenin | Cirsium chanroenicum | Murine CT26 CRC cells were introduced into BALB/C mice | Intraperitoneal injection, 25 and 50 mg/kg | Induced apoptotic death of cancer cells; suppression STAT3 | [272] |
| Peonidin | Sweet potato (Ipomoea batatas) | AOM-induced CF-1 mice | Dietary supplementation, 10 to 30% | Blocked cell cycle at the G1 phase; activated caspase-3 | [273] |
| Petunidin | Lycium ruthenicum | Nude mice | Intraperitoneal injection, 25 and 50 mg/kg | Induced ferroptosis via inhibiting SLC7A11 | [274] |
| Phloretin | Manchurian apricot | COLO 205 cells derived tumor in BALB/c nude mice | Route of administration not reported, 25 mg/kg | Inhibited tumor growth; upregulated p53, p21 and E-cadherin | [275] |
| Polyphenon E | AOM-induced colon carcinogenesis in F344 rats | Oral administration, 0.24% | Induced apoptosis; decreased eicosanoid, prostaglandin E2, and interleukin B4 in plasma; decreased nuclear β-catenin and increased expression of RXRα,β and γ in adenocarcinomas | [276] | |
| Procyanidin | Cider apple (Malus domestica) | AOM-induced Wistar rats | Oral administration, 0.01% | Suppressed protein kinase; down-regulated of polyamine production; stimulated caspase-3 | [277] |
| Male C57/BL6 mice transfected with CT26 cells | Oral gavage, 30 mg/kg | Reduced cellular oxidative stress through modulation of Nrf2/ARE signaling | [278] | ||
| Quercetin | Apples, nuts, cauliflower, cabbage, onions, grapes, berries, broccoli, citrus fruits, cherries, green tea, and coffee | AOM-induced colon tumor in C57BL/6J male mice | Dietary supplementation, 0.5% | Induced apoptosis; upregulated CB1-R; downregulated STAT3 and p-STAT3; downregulated Bax/Bcl-2 ratio | [279] |
| Subcutaneous DLD-1 human colon tumor fragment implant in male athymic nu/nu mice | Intraperitoneal injection, 30 mg/kg | Enhanced radiosensitivity by inhibiting ATM-mediated signaling pathway | [280] | ||
| AOM-induced CRC in male weanling Sprague-Dawley rats | Dietary supplement, 4.5 g/kg | Reduced the number of crypts; inhibited proliferation; induced apoptosis; suppressed COX-1, COX-2 and iNOS | [281] | ||
| AOM/DSS induced colon carcinogenesis in C57BL/6J mice | Dietary supplementation, 30 mg/kg | Reduced number and size of colon tumors; suppressed inflammation; downregulated LOP, NO, SOD, G6PD, and GSH | [282] | ||
| CT-26 lung tumor metastasis in BALB/c mice | Intraperitoneal injection, 50 mg/kg | Suppressed lung metastasis; induced apoptosis | [283] | ||
| HT-29 tumor xenograft in BALB/c nude mice | Subcutaneous injection, 10 mg/kg | Enhanced radiosensitivity; inhibited Notch-1 signaling | [284] | ||
| Rutin | Buckwheat, Mez, Labisia pumila, Sophora japonica L., Schum, Canna indica L., and Ruta graveolens L. | SW480 cell-induced tumor xenograft | Intraperitoneal injection, 20 mg/kg | Suppressed tumor growth; decreased angiogenesis and VEGF levels | [285] |
| Scutellarin | Scutellaria barbata | AOM/DSS-induced male C57BL/6 mice | Intraperitoneal injection, 25, 50 and 100 mg/kg | Inhibited Wnt/β-catenin signal transduction | [286] |
| RKO cells were subcutaneously implanted into female nude mice | Intraperitoneal injection, 50, 150 and 300 mg/kg | Suppressed tumor growth and metastasis | [287] | ||
| AOM/DSS-induced mice | Intraperitoneal injection, 25, 50 and 100 mg/kg | Suppressed the Hedgehog signaling cascade | [288] | ||
| Silibinin | Silybum marianum | LoVo cell deposition on eight days old fertilized chicken egg | Route of administration not reported, 9.64 μg/mL | Decreased in VDI; upregulated Flt-1 gene | [289] |
| AOM-induced CRC in male Wistar rats | Intragastric intonation, 300 mg/kg | Suppressed preneoplastic lesion formation; activated apoptosis; registered sub G0/G1 cell cycle arrest; reduced MMP-7, IL-1β and TNF-α | [290] | ||
| Tangeretin | Peel of citrus fruits | HT-29 induced tumor xenograft in BALB/c nude mice | Route of administration not reported, 5 mg/kg | Suppressed tumor growth | [291] |
| Taxifolin | Olive oil, grapes, citrus fruits, and onion | HCT116 tumor xenograft in athymic male nude mice | Intraperitoneal injection, 15 and 25 mg/kg | Suppressed tumor growth; induced apoptosis; inhibited cyclin D; degraded β-catenin; inhibited of Akt phosphorylation | [292] |
| Tricin | Rice bran, oats, barley, and wheat | Colon26-Luc colon tumor and lung metastasis model in BALB/c mice | Oral gavage, 19 and 37.5 mg/kg | Suppressed tumor growth; reduced metastasis incidence | [293] |
| AOM-DSS induced CRC in male Crj: CD-1 mice | Dietary supplement, 50 and 250 ppm | Restored colonic length; reduced number of incidences and multiplicity of adenomas and adenocarcinomas; downregulated PCNA and TNF-α | [294] | ||
| Troxerutin | Tea and coffee | DMH-induced colon carcinogenesis in male albino Wistar rats | Oral administration, 12.5, 25 and 50 mg/kg | Lowered ACF formation and crypt multiplicity; reduced cytochrome P450, cytochrome b5, cytochrome P4502E1, NADPH-cytochrome P450 reductase, and NADH-cytochrome b5 reductase and upregulates phase GST, DTD and UDPGT | [295] |
| Vitexin | Passionflower, bamboo leaves, pearl, and millet | HCT116 tumor xenograft in nude BALB/c mice | Oral administration, 25, 50 and 100 mg/kg | Suppressed tumor growth; increased phosphorylation of JNK; upregulated LC3 II and ApoL1 | [296] |
| HCT116DR tumor xenograft in female athymic BALB/c nude mice | Oral administration, 25 and 50 mg/kg | Suppressed tumor growth; induced apoptosis; downregulated HSP90, HSP70, HSP27, Atg7, Beclin-1, LC3 II and Bcl-2; upregulated Bax and PARP1; cleaved caspase-3 and caspase-9 | [297] | ||
| Wogonin | Scutellaria baicalensis, Scutellaria radix | AOM/DSS-induced colitis related colon cancer in C57BL/6 mice | Gastric intubation, 60 mg/kg | Decreased cell proliferation; lowered the expression and secretion of IL-6 and IL-1β and expression of NF-κB; increased Nrf2 nuclear translocation | [298] |
| AOM-DSS-induced CRC animal model in C57BL/6 mice | Route of administration not reported, 50 and 100 mg/kg | Reduced tumor multiplicity; reverted colon length to normal | [299] | ||
| SW480 induced tumor xenograft in BALB/c nude mice | Intraperitoneal injection, 2 mM | Downregulated of YAP-1 and IRF3; upregulated p-YAP | [300] | ||
| Xanthohumol | Humulus lupulus | AOM-induced colorectal carcinogenesis in male Sprague-Dawley rats | Oral gavage, 5 mg/kg | Suppressed tumor growth; induced apoptosis; suppressed COX-2 and iNOS | [301] |
| Zapotin | Tropical fruit zapote blanco (Casimiroa edulis) | AOM/DSS-induced female CF-1 mice | Intragastric administration, 5 and 10 mg/kg | Induced cell cycle arrest and apoptosis | [302] |
| Phenolic acids | |||||
| Caffeic acid | Coffee, wine tea | CT-26 lung metastasis in BALB/c mice | Oral administration, 0.1 and 0.5 g/kg | Inhibited lung metastasis; suppressed MEK1, TOPK, and TAP-induced activation of AP1, NF-κB and ERK signaling; inhibited TAP, EGF and H-Ras induced neoplastic transformation | [303] |
| HCT116 tumor xenograft in NSG mice | Intraperitoneal injection, 10 mg/kg | Inhibited CSC growth and self-renewal by inhibition of PI3K/Akt signaling | [304] | ||
| HCT116 tumor xenograft in BALB/c AnN Foxn-1 nude mice | Oral administration, 50 nmol/kg | Inhibited PI3K/Akt/mTOR pathway; suppressed MMP-9, cyclin D1, Cdk4, cyclin E, PCNA, FASN c-Myc, and N-cadherin expression; upregulated p21 | [305] | ||
| HT-29 tumor xenograft in BALB/c nude mice | Intragastric administration, CAPE (10 mg/kg); CAPE-pNO2 5, (10 and 20 mg/kg) | Inhibited tumor growth and VEGF expression; upregulated p53, p27, p21, cyt. c, and cleaved caspase-3; downregulated procaspase-3, Cdk2, and c-Myc; | [306] | ||
| HCT116 tumor xenograft in nude mice | Oral administration, 0.2 and 2 mg/kg | Suppressed tumor growth; displayed cell cycle arrest in S phase and autophagic cell death | [307] | ||
| Chlorogenic acid | Apple, betel, coffee beans, kiwi, grapes, eggplant, pear, plum, potato, and tea | MAM acetate-induced carcinogenesis hamsters | Oral administration, 0.025% dietary supplement | Reduced colon tumor incidences; registered antioxidative effect; inhibited the activity of microsomal enzyme | [308] |
| AOM-induced ACF in colon of male F344 rats | Oral administration, 0.025% dietary supplement | Reduced ACF formation and growth | [309] | ||
| Ellagic acid | AOM-induced colon tumors in rats | Oral administration, 250, 2500 and 5000 ppm | Inhibited the incidence of adenocarcinomas in the small intestine | [310] | |
| DMH-induced colon cancer in rats | Oral administration, 60 mg/kg | Lowered the frequency of ACF and lipid peroxidation; increased the activity of CAT, SOD, GPx, GR and GST; restored the levels of vitamin C, vitamin E and GSH | [311] | ||
| DMH-induced colon cancer in Wistar albino rats | Oral administration, 60 mg/kg | Inhibited NF-κB, iNOS, COX-2, TNF-α and IL-6; restored the levels 5′-ND, γ-GT, CEA, AFP and LDH | [312] | ||
| DMH-induced colon cancer in rats | Oral administration, 60 mg/kg | Inhibited PI3K-p58 activation; downregulated Akt and Bcl-2; upregulated Bax | [313] | ||
| DMH-induced colorectal cancer in rats | Oral administration, 60 mg/kg | Inhibited ACF formation; increased the activity of CAT, SOD, GPx, and GR; inhibited ODC expression through inhibition of c-MYC | [314] | ||
| DMH-induced colon cancer in male Laca mice | Oral administration, 10 mg/kg | Restored colon membrane alterations | [315] | ||
| Ferulic acid | Rice, wheat, pineapple, grains, and peanuts | AOM-induced colon cancer in male Fischer 344 rats | Oral administration, 250 ppm and 500 ppm | Reduced number and size of adenomas; increased the activity of GST and QR | [316] |
| AOM-induced colon carcinogenesis in F344 rats | Dietary supplement of 3-(4′-geranyloxy-3-methoxyphenyl)-2propenoate (geranylated derivative of ferulic acid) 0.1% and 0.2% | Decreased the number of ACF | [317] | ||
| Gallic acid | Barriers and pomegranates | DMH-induced colon cancer in male Wister rats | Oral administration, 50 mg/kg | Reduced lipid peroxidation, LOOH, CD, SOD, CAT, GSH, GR and GPx; reduced ascorbic acid and tocopherol levels | [318] |
| SW480 induced tumor xenograft in NOD SCID gamma NSG mice | Intraperitoneal injection, 200 mg/kg | Exerted antitumor activity mediated by interaction with G-quadruplexes | [319] | ||
| DSS-induced acute colitis in C57BL/6 mice | Oral administration, 5 and 25 mg/kg | Suppressed acute colitis; inhibited phosphorylation of STAT3 | [320] | ||
| HCT116 and HT-29 tumor xenografts in BALB/c nude mice | Intraperitoneal injection, 80 mg/kg | Suppressed p-SRC, p-EGFR, p-Akt and p-STAT3 | [321] | ||
| Ulcerative colitis in rats | Oral administration, 10 mg/kg | Suppressed colon cancer; induced ferroptosis | [322] | ||
| DMH-induced colon cancer in male albino Wister rats | Oral administration, 50 mg/kg | Elevated the activity of cytochrome P450, cytochrome b5, GST, DT-diaphorase and γ-GT | [323] | ||
| Geraniin | Phyllanthus amarus | SW480 tumor xenograft in nude mice | Oral administration, 10, 20 and 40 mg/kg | Suppressed tumor growth; induced apoptosis; inhibited phosphorylation of PI3K and Akt | [324] |
| p-Coumaric acid | Mushrooms, apples, pears, barriers, oranges, and beans | DMH-induced colon carcinogenesis in male albino Wistar rats | Intragastric intubation, 100 mg/kg | Reduced ACF, DACF, MDF and BCAC | [325] |
| Syringic acid | Olives, dates, pumpkins, grapes, and palms | DSS-induced mice | Oral administration, 25 mg/kg | Decreased the level of iNOS, COX-2, TNF-α, IL-1β and IL-6; reduced activation and accumulation of p-STAT-3 by decreasing expression of p65-NF-κB | [326] |
| DMH-induced colorectal cancer in male rats | Oral administration, 50 mg/kg | Reduced tumor incidences, tumor volume and average number of tumors | [327] | ||
| Lignans | |||||
| Arctigenin | Arctium lappa, Forsythia suspensa. | CT-26 cells derived lung metastasis model in BALB/c mice | Oral gavage, 50 mg/kg | Reduced the number of lung nodules; induced apoptosis in lung tissue; inhibited EMT in lung tissue; induced cleavage of caspase-3, caspase-9, and PARP; downregulated Bcl-2 and Bcl-xL; upregulated Bax | [328] |
| Daurinol | Haplophyllum dauricum | HCT116 tumor xenograft in athymic BALB/c (Slc/nu) nude mice | Oral administration, 5 and 10 mg/kg | Suppressed tumor growth; upregulated p-Chk1(Ser345)/Chk1 | [329] |
| Dehydrodiisoeugenol | Myristica fragrans Houtt | HCT116, zsw480, and patient-derived xenograft in female NOD/SCID mice | Intraperitoneal injection, 40 mg/kg | Suppressed tumor growth by inducing ER stress; upregulated BiP, PERK, and IRE1α | [330] |
| Gomisin A | Schisandra chinesis | CT-26 induced lung metastasis in female BALB/c mice | Intraperitoneal injection, 50 mg/kg | Suppressed lung metastasis; reduced the number of lung nodules; increased phosphorylation of AMPK and p38 in lung tissue | [331] |
| Honokiol | Magnolia grandiflora | SW620 tumor xenograft in female athymic BALB/c nude mice nu/nu | Intragastric administration, 50 mg/kg | Inhibited proliferation of CRC; upregulated TGF-β1 and p53 by upregulating BMP7 | [332] |
| Justicidin A | Justicia procumbens | HT-29 tumor xenograft in NOD-SCID mice | Oral administration, 6.2 mg/kg | Suppressed tumor growth; induced autophagy in tumor tissue; induced apoptosis | [333] |
| Magnolol | Magnolia officinalis | CT-26 and HT-29 tumor in BALB/c and Cg-Foxn1nu/CrlNarl nude mice respectively | Route of administration not reported, 50 and 100 mg/kg | Inhibited tumor growth; induced apoptosis; upregulation of Fas, Fas-L, cleaved caspase-3, cleaved caspase-9 and cleaved caspase-8; downregulated NF-κB, PKCδ, ERK, Akt, C-FLIP, and MCL-1; inhibition of PKCδ-NF-κB signaling | [334] |
| HCT116 tumor xenograft in female BLB/c nude mice | Intraperitoneal injection, 5 mg/kg | Suppressed tumor growth without showing any toxicity | [335] | ||
| Schisandrin B | Schisandra chinensis, Schisandra propinqua, and Schisandra rubriflora | AOM-DSS-induced CRC in C57BL/6 mice | Oral administration, 3.7–30 mg/kg | Suppressed SIRT1 | [336] |
| Secoisolariciresinol | Fitzroya cupressoides and Crossosoma bigelovii | HCT116 tumor xenograft in male BALB/c nude mice | Route of administration not reported, 50, 100 and 200 mg/kg | Inhibited tumor growth; induced pyroptosis; downregulated Ki-67; upregulated N-GSDMD | [337] |
| DSS-induced colitis in mice | Dietary supplementation, 200 mg/kg | Suppressed tumor growth; reduced IL-1β, IL-18, TNF-α and NLRP1 | [338] | ||
| Sesaminol | Sesamum indicum | Ethanol-induced CRC in male C57BL/6NCr mice | Oral administration, 2.5 mg/mice | Reduced colonic lesions; downregulated iNOS and CYP2E1; lowered TNF-α, IL-6, MCP-1 and NF-κB levels; increased cell adhesion by upregulation of ZO-1, occludin and cladulin-1 | [339] |
| Tracheloside | Carthamus tinctorious L. (safflower) | CT-26 lung metastasis in BALB/c mice | Oral administration, 25 and 50 mg/kg | Suppressed lung metastasis; induced apoptosis; upregulated E-cadherin RNA; downregulated N-cadherin, vimentin, snail and twist RNA | [340] |
| Vitexin | Vitex negudo | HCT116 tumor xenograft in female nu/nu mice | Intraperitoneal injection, 40 mg/kg | Inhibited tumor growth and lowered tumor volume; upregulated PUMA and p53; induced PUMA-mediated apoptosis | [341] |
| Stilbenes | |||||
| Piceatannol | Red and white grapes | AOM/DSS-induced colon tumor in C57BL/6J mice | Oral administration, 5 and 12.5 mg/kg | Decreased tumor number and size; decreased Ki-67- and COX-2-positive cell number; downregulated MCP1 and PD1 | [342] |
| Polydatin | Picea sitchensis | Caco-2 tumor xenograft in C57BL/6 mice | Subcutaneously into the tumor, 150 mg/kg | Suppressed tumor growth; upregulated miR-382; downregulated PD-L1 | [343] |
| Pterostilbene | Blueberries and cranberries | AOM-induced colonic ACF preneoplastic lesions and adenomas in male ICR mice | Oral administration, 50 or 250 ppm | Reduced ACF and adenoma formation; induced apoptosis; downregulated iNOS and COX-2; inhibited Wnt/β-catenin signaling through suppressing phosphorylation of GSK3β; inhibited VEGF, cyclin D, MMPs, Ras, PI3K/Akt and EGFR | [344] |
| AOM-induced colon tumors in F344 rats | Oral administration, 40 ppm | Reduced the proliferation of non-metastatic adenomas; downregulated IL-1β, IL-4, TNF-α, PCNA, β-catenin and cyclin D and p-NF-κB/p65 | [345] | ||
| AOM-induced colon tumor in male BALB/c mice | Oral administration, 50 or 250 ppm | Reduced NF-κB through inhibition of PKC-β phosphorylation; downregulated iNOS, COX-2 and aldose reductase; upregulated HO-1, GR and Nrf2 signaling | [346] | ||
| CL187 transplantation model in BALB/c nude mice | Intraperitoneal injection, 25, 50, 100 and 200 mg/kg | Inhibited Top1-mediated DNA damage repair pathway | [347] | ||
| AOM-induced colonic ACF preneoplastic lesions in F344 rats | Oral administration, 40 ppm | Inhibited ACF formation; blocked cell proliferation and iNOS | [348] | ||
| Resveratrol | Grapes, blueberries, raspberries, mulberries, and peanuts | LoVo cell-mediated metastasis model in mice | Intragastric administration, 50, 100 and 150 mg/kg | Inhibited metastasis; decreased tumor size; suppressed TGF-β1/Smad pathway; downregulated Snail and vimentin; upregulated E-cadherin expression | [349] |
| APCCKO/Krasmut mice | Dietary supplementation, 150 ppm and 300 ppm | Suppressed tumor formation; reduced tumor size; downregulated Kras expression | [350] | ||
| DSS-induced colon carcinogenesis in rats | Oral administration, 60 mg/kg | Reduced ACF and MDF | [351] | ||
| HCT116 tumor xenograft in ICR SCID mice | Oral administration, 150 mg/kg | Suppressed tumor growth; induced apoptosis; inhibited NF-κB | [352] | ||
| COLO250-luc tumor xenograft in athymic mice | Injection in tumor, 6 μg/implant | Suppressed tumor growth | [353] | ||
| HT-29 tumor xenograft in BALB/c nu/nu mice | Intragastric administration, 480, 960 and 1920 mg/kg | Suppressed VEGF-mediated angiogenesis | [354] | ||
| Miscellaneous compounds | |||||
| Oleuropein | Olives (Olea europaea) | AOM-induced CRC in female A/J mice | Dietary supplementation, 125 mg/kg | Suppressed preneoplastic lesions; lowered tumor incidences; prevented DNA damage | [355] |
| Thymol | Thymus vulgaris L. | HCT116 tumor xenograft and lung metastasis model in BALB/c nude mice | Intraperitoneal injection, 75 and 150 mg/kg | Induced apoptosis; upregulated E-cadherin; downregulated N-cadherin; suppressed lung metastasis by inhibiting Wnt/β-catenin pathway | [356] |
| Verbascoside | Genus, Cistanche | HCT116 tumor xenograft in BALB/c nude mice | Tail vein injection, 20, 40, and 80 mg/kg | Upregulated HIPK2, p53 and Bax; downregulation Bcl-2 | [357] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De, S.; Paul, S.; Manna, A.; Majumder, C.; Pal, K.; Casarcia, N.; Mondal, A.; Banerjee, S.; Nelson, V.K.; Ghosh, S.; et al. Phenolic Phytochemicals for Prevention and Treatment of Colorectal Cancer: A Critical Evaluation of In Vivo Studies. Cancers 2023, 15, 993. https://doi.org/10.3390/cancers15030993
De S, Paul S, Manna A, Majumder C, Pal K, Casarcia N, Mondal A, Banerjee S, Nelson VK, Ghosh S, et al. Phenolic Phytochemicals for Prevention and Treatment of Colorectal Cancer: A Critical Evaluation of In Vivo Studies. Cancers. 2023; 15(3):993. https://doi.org/10.3390/cancers15030993
Chicago/Turabian StyleDe, Samhita, Sourav Paul, Anirban Manna, Chirantan Majumder, Koustav Pal, Nicolette Casarcia, Arijit Mondal, Sabyasachi Banerjee, Vinod Kumar Nelson, Suvranil Ghosh, and et al. 2023. "Phenolic Phytochemicals for Prevention and Treatment of Colorectal Cancer: A Critical Evaluation of In Vivo Studies" Cancers 15, no. 3: 993. https://doi.org/10.3390/cancers15030993
APA StyleDe, S., Paul, S., Manna, A., Majumder, C., Pal, K., Casarcia, N., Mondal, A., Banerjee, S., Nelson, V. K., Ghosh, S., Hazra, J., Bhattacharjee, A., Mandal, S. C., Pal, M., & Bishayee, A. (2023). Phenolic Phytochemicals for Prevention and Treatment of Colorectal Cancer: A Critical Evaluation of In Vivo Studies. Cancers, 15(3), 993. https://doi.org/10.3390/cancers15030993