
CAR-Based Immunotherapy of Solid Tumours—A Survey of the Emerging Targets



Abstract
:Simple Summary
Abstract

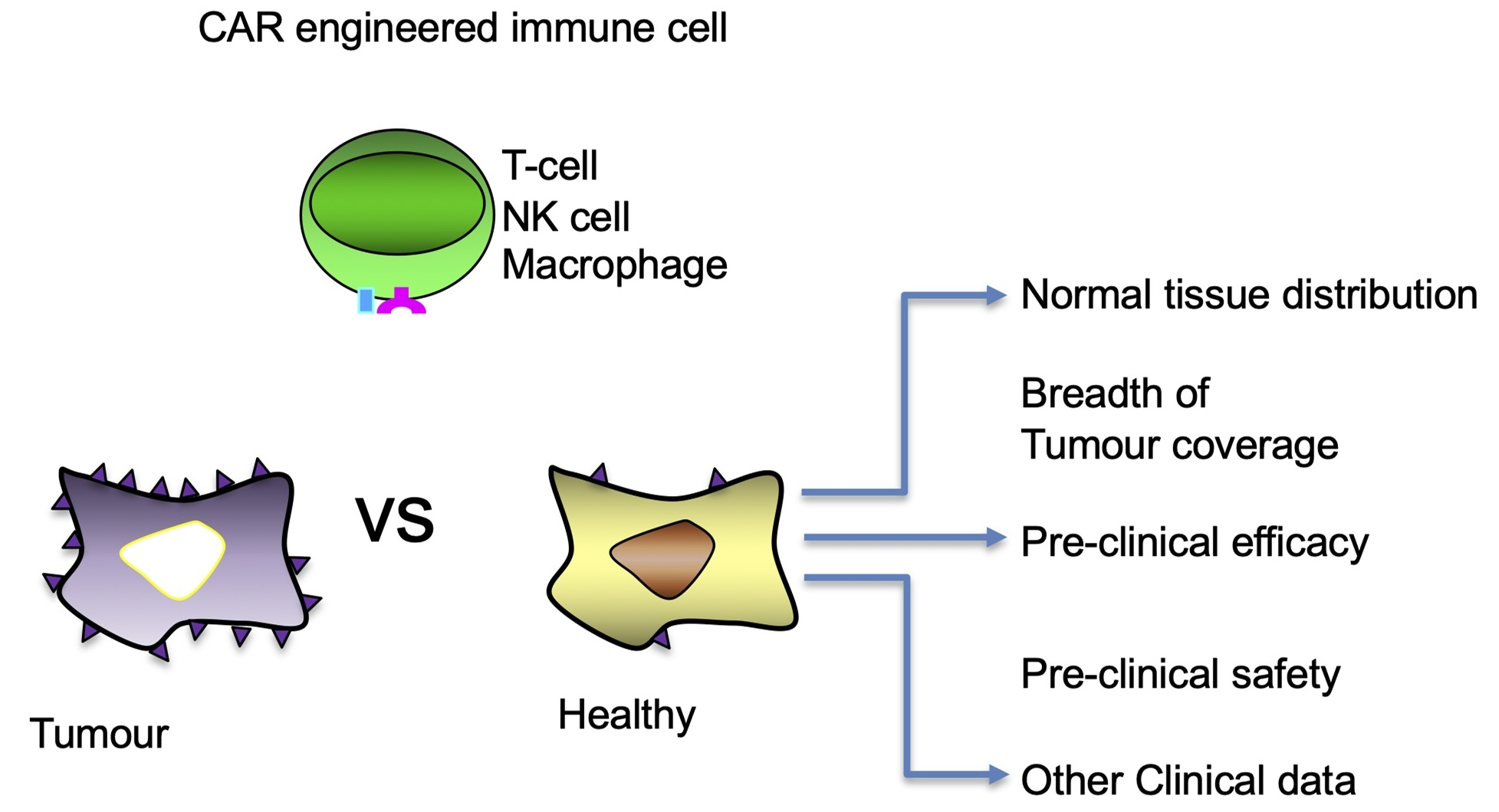
1. Introduction
2. Receptor Tyrosine Kinases
2.1. Fibroblast Growth Factor Receptor 4 (FGFR4)
2.2. Erythropoietin-Producing Hepatocellular Carcinoma (Ephrin) Receptors
2.2.1. Ephrin Type A3 Receptor (EphA3)
2.2.2. Ephrin Type A10 Receptor (EphA10)
2.2.3. Ephrin Type B4 Receptor (EphB4)
2.3. Receptor Tyrosine Kinase-Like Orphan Receptor Family Member (ROR)2
2.4. ALK Receptor Tyrosine Kinase
2.5. AXL Receptor Tyrosine Kinase
2.6. Platelet-Derived Growth Factor Receptor (PDGFR) α
2.7. Protein Tyrosine Kinase 7 (PTK7; Colon Carcinoma Kinase 4; CCK4)
3. Adhesion Molecules
3.1. Intracellular Adhesion Molecule 1 (ICAM-1)
3.2. Cadherins
3.2.1. Cadherin 6
3.2.2. Cadherin 17
3.3. Nectin-4 (Poliovirus Receptor-like 4)
3.4. CD44 v6 Splice Variant (CD44v6)
4. Integrins
4.1. αvβ6 Integrin
4.2. αvβ3 Integrin
5. Vascular Targets
5.1. CLEC14A
5.2. Apelin Receptor
6. Extracellular Matrix Targets
6.1. Fibronectin with Extra Domain B (EDB)
6.2. Fibronectin with Extra Domain A (EDA)
7. Gangliosides
7.1. Ganglioside D3
7.2. Ganglioside M2
8. B7 Family Members
8.1. B7-H4
8.2. B7-H6
9. Prostate Antigens
9.1. Kallikrein-Related Peptidase 2 (KLK2)
9.2. Six-Transmembrane Epithelial Antigen of Prostate (STEAP) Family Members
9.2.1. Six-Transmembrane Epithelial Antigen of Prostate (STEAP) 1
9.2.2. Six-Transmembrane Epithelial Antigen of Prostate (STEAP) 2
10. Notch Ligands
10.1. Delta-like Ligand 3 (DLL3)
10.2. Delta-like Homologue 1 (DLK1)
11. Viral Antigens
11.1. Human Endogenous Retrovirus (HERV) K
11.2. Latent Membrane Protein 1 (LMP1)
12. Glypicans
12.1. Glypican 1
12.2. Glypican 2
13. Placental Antigens
13.1. Placental Alkaline Phosphatase (PLAP)
13.2. Trophoblast Cell Surface Antigen 2 (TROP2)
13.3. Human Leucocyte Antigen G (HLA-G)
13.4. 5T4 (Trophoblast Glycoprotein)
14. Lewis Y
15. MG7
16. Glial Cell Line-Derived Neurotrophic Factor Family Receptor α4 (GFR α4)
17. Leucine-Rich Repeat-Containing G Protein-Coupled Receptor 5 (LGR5)
18. Non-Functioning P2X7 Receptor (nfP2X7)
19. Follicle-Stimulating Hormone Receptor (FSHR)
20. Guanylyl Cyclase 2C (GUCY2C)
21. Thyroid-Stimulating Hormone Receptor (TSHR)
22. C-Type Lectin 4 Family (CLEC4)
23. CD47
24. Transmembrane 4 L6 Family Member 1 (TM4SF1)
25. A Disintegrin and Metalloproteinase 10 (ADAM10)
26. Chlorotoxin Ligands
27. Heat Shock Protein 70 (HSP70)
28. Cripto-1
29. Glucose-Regulated Protein 78 (GRP78)
30. CD147 (Basigin)
31. CD317 (Tetherin)
32. Chondroitin Sulphate Proteoglycan 4 (CSPG4)
33. CD24
34. Müllerian-Inhibiting Substance Type 2 Receptor (MISIIR)
35. SLC3A2 (Solute Carrier Family 3 Member 2)/CD98hc (Heavy Chain)
36. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Halim, L.; Maher, J. CAR T-cell immunotherapy of B-cell malignancy: The story so far. Ther. Adv. Vaccines Immunother. 2020, 8, 2515135520927164. [Google Scholar] [CrossRef] [PubMed]
- Van de Donk, N.; Usmani, S.Z.; Yong, K. CAR T-cell therapy for multiple myeloma: State of the art and prospects. Lancet Haematol. 2021, 8, e446–e461. [Google Scholar] [CrossRef] [PubMed]
- Maher, J.; Davies, D.M. CAR based immunotherapy of solid tumours – a clinically based review of target antigens. Biology 2023, 12, 287. [Google Scholar] [CrossRef]
- Hwang, M.S.; Miller, M.S.; Thirawatananond, P.; Douglass, J.; Wright, K.M.; Hsiue, E.H.; Mog, B.J.; Aytenfisu, T.Y.; Murphy, M.B.; Aitana Azurmendi, P.; et al. Structural engineering of chimeric antigen receptors targeting HLA-restricted neoantigens. Nat. Commun. 2021, 12, 5271. [Google Scholar] [CrossRef] [PubMed]
- Arman, I.; Haus-Cohen, M.; Reiter, Y. The Intracellular Proteome as a Source for Novel Targets in CAR-T and T-Cell Engagers-Based Immunotherapy. Cells 2022, 12, 27. [Google Scholar] [CrossRef] [PubMed]
- Schettini, F.; Barbao, P.; Braso-Maristany, F.; Galvan, P.; Martinez, D.; Pare, L.; De Placido, S.; Prat, A.; Guedan, S. Identification of cell surface targets for CAR-T cell therapies and antibody-drug conjugates in breast cancer. ESMO Open 2021, 6, 100102. [Google Scholar] [CrossRef]
- Alijaj, N.; Moutel, S.; Gouveia, Z.L.; Gray, M.; Roveri, M.; Dzhumashev, D.; Weber, F.; Meier, G.; Luciani, P.; Rossler, J.K.; et al. Novel FGFR4-Targeting Single-Domain Antibodies for Multiple Targeted Therapies against Rhabdomyosarcoma. Cancers 2020, 12, 3313. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, P.M.; Kumar, R.; Li, W.; Hoglund, V.; Wang, L.; Zhang, Y.; Shi, M.; Beak, D.; Cheuk, A.; Jensen, M.C.; et al. FGFR4-Targeted Chimeric Antigen Receptors Combined with Anti-Myeloid Polypharmacy Effectively Treat Orthotopic Rhabdomyosarcoma. Mol. Cancer Ther. 2022, 21, 1608–1621. [Google Scholar] [CrossRef]
- Chan, S.L.; Schuler, M.; Kang, Y.K.; Yen, C.J.; Edeline, J.; Choo, S.P.; Lin, C.C.; Okusaka, T.; Weiss, K.H.; Macarulla, T.; et al. A first-in-human phase 1/2 study of FGF401 and combination of FGF401 with spartalizumab in patients with hepatocellular carcinoma or biomarker-selected solid tumors. J. Exp. Clin. Cancer Res. 2022, 41, 189. [Google Scholar] [CrossRef]
- London, M.; Gallo, E. Critical role of EphA3 in cancer and current state of EphA3 drug therapeutics. Mol. Biol. Rep. 2020, 47, 5523–5533. [Google Scholar] [CrossRef]
- Hafner, C.; Schmitz, G.; Meyer, S.; Bataille, F.; Hau, P.; Langmann, T.; Dietmaier, W.; Landthaler, M.; Vogt, T. Differential gene expression of Eph receptors and ephrins in benign human tissues and cancers. Clin. Chem. 2004, 50, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Ba, T.; Ho, J.; Chen, D.; Cheng, Y.; Wang, L.; Xu, G.; Xu, L.; Zhou, Y.; Wei, Y.; et al. First-in-Human Trial of EphA2-Redirected CAR T-Cells in Patients With Recurrent Glioblastoma: A Preliminary Report of Three Cases at the Starting Dose. Front. Oncol. 2021, 11, 694941. [Google Scholar] [CrossRef] [PubMed]
- Ruff, M.; Sakemura, R.; Cox, M.; Torghabeh, M.H.; Moreno, P.R.; Schick, K.; Sarkaria, J.; Kenderian, S. Exth-32. Development of EphA3 directed chimeric antigen receptor T cell therapy for the treatment of glioblastoma multiforme. Neuro Oncol. 2019, 21, vi88–vi89. [Google Scholar] [CrossRef]
- Swords, R.T.; Greenberg, P.L.; Wei, A.H.; Durrant, S.; Advani, A.S.; Hertzberg, M.S.; Jonas, B.A.; Lewis, I.D.; Rivera, G.; Gratzinger, D.; et al. KB004, a first in class monoclonal antibody targeting the receptor tyrosine kinase EphA3, in patients with advanced hematologic malignancies: Results from a phase 1 study. Leuk. Res. 2016, 50, 123–131. [Google Scholar] [CrossRef]
- Gan, H.; Cher, L.; Inglis, P.-L.; Lwin, Z.; Lau, E.; Wichmann, C.; Ackermann, U.; McDonald, A.; Gunjur, A.; Coombs, N.; et al. Phase I safety and bioimaging trial of ifabotuzumab in patients with glioblastoma. J. Nucl. Med. 2021, 62, 104. [Google Scholar]
- Cha, J.H.; Chan, L.C.; Wang, Y.N.; Chu, Y.Y.; Wang, C.H.; Lee, H.H.; Xia, W.; Shyu, W.C.; Liu, S.P.; Yao, J.; et al. Ephrin receptor A10 monoclonal antibodies and the derived chimeric antigen receptor T cells exert an antitumor response in mouse models of triple-negative breast cancer. J. Biol. Chem. 2022, 298, 101817. [Google Scholar] [CrossRef]
- Zhang, X. The Expression Profile and Prognostic Values of EPHA Family Members in Breast Cancer. Front. Oncol. 2021, 11, 619949. [Google Scholar] [CrossRef]
- Kubo, H.; Yagyu, S.; Nakamura, K.; Yamashima, K.; Tomida, A.; Kikuchi, K.; Iehara, T.; Nakazawa, Y.; Hosoi, H. Development of non-viral, ligand-dependent, EPHB4-specific chimeric antigen receptor T cells for treatment of rhabdomyosarcoma. Mol. Ther. Oncolytics 2021, 20, 646–658. [Google Scholar] [CrossRef]
- Yagyu, S.; Mochizuki, H.; Yamashima, K.; Kubo, H.; Saito, S.; Tanaka, M.; Sakamoto, K.; Shimoi, A.; Nakazawa, Y. A lymphodepleted non-human primate model for the assessment of acute on-target and off-tumor toxicity of human chimeric antigen receptor-T cells. Clin. Transl. Immunol. 2021, 10, e1291. [Google Scholar] [CrossRef]
- Jackovich, A.; Gitlitz, B.J.; Tiu-lim, J.W.W.; Duddalwar, V.; King, K.G.; El-Khoueiry, A.B.; Thomas, J.S.; Tsao-Wei, D.; Quinn, D.I.; Gill, P.S.; et al. Phase II trial of soluble EphB4-albumin in combination with PD-1 antibody (pembrolizumab) in relapsed/refractory head neck squamous cell carcinoma. J. Clin. Oncol. 2021, 39, 6016. [Google Scholar] [CrossRef]
- Endo, M.; Kamizaki, K.; Minami, Y. The Ror-Family Receptors in Development, Tissue Regeneration and Age-Related Disease. Front. Cell Dev. Biol. 2022, 10, 891763. [Google Scholar] [CrossRef] [PubMed]
- Rebagay, G.; Yan, S.; Liu, C.; Cheung, N.K. ROR1 and ROR2 in Human Malignancies: Potentials for Targeted Therapy. Front. Oncol. 2012, 2, 34. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Nerreter, T.; Chang, J.; Qi, J.; Li, X.; Karunadharma, P.; Martinez, G.J.; Fallahi, M.; Soden, J.; Freeth, J.; et al. Mining Naive Rabbit Antibody Repertoires by Phage Display for Monoclonal Antibodies of Therapeutic Utility. J. Mol. Biol. 2017, 429, 2954–2973. [Google Scholar] [CrossRef]
- Walker, A.J.; Majzner, R.G.; Zhang, L.; Wanhainen, K.; Long, A.H.; Nguyen, S.M.; Lopomo, P.; Vigny, M.; Fry, T.J.; Orentas, R.J.; et al. Tumor Antigen and Receptor Densities Regulate Efficacy of a Chimeric Antigen Receptor Targeting Anaplastic Lymphoma Kinase. Mol. Ther. 2017, 25, 2189–2201. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, E.; Vitali, A.; Muller, H.; Alonso-Ferrero, M.; Barisa, M.; Gavriil, A.; Piapi, A.; Leboreiro-Babe, C.; Gileadi, T.; Yeung, J.; et al. Targeting of low ALK antigen density neuroblastoma using AND logic-gate engineered CAR-T cells. Cytotherapy 2022, 25, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Liu, M.; Wang, L.; Zhu, K.; Cai, M.; Chen, X.; Feng, Y.; Yang, S.; Fu, S.; Zhi, C.; et al. Remodelling of tumour microenvironment by microwave ablation potentiates immunotherapy of AXL-specific CAR T cells against non-small cell lung cancer. Nat. Commun. 2022, 13, 6203. [Google Scholar] [CrossRef] [PubMed]
- Holtzhausen, A.; Harris, W.; Ubil, E.; Hunter, D.M.; Zhao, J.; Zhang, Y.; Zhang, D.; Liu, Q.; Wang, X.; Graham, D.K.; et al. TAM Family Receptor Kinase Inhibition Reverses MDSC-Mediated Suppression and Augments Anti-PD-1 Therapy in Melanoma. Cancer Immunol. Res. 2019, 7, 1672–1686. [Google Scholar] [CrossRef]
- Myers, K.V.; Amend, S.R.; Pienta, K.J. Targeting Tyro3, Axl and MerTK (TAM receptors): Implications for macrophages in the tumor microenvironment. Mol. Cancer 2019, 18, 94. [Google Scholar] [CrossRef]
- Wei, J.; Sun, H.; Zhang, A.; Wu, X.; Li, Y.; Liu, J.; Duan, Y.; Xiao, F.; Wang, H.; Lv, M.; et al. A novel AXL chimeric antigen receptor endows T cells with anti-tumor effects against triple negative breast cancers. Cell Immunol. 2018, 331, 49–58. [Google Scholar] [CrossRef]
- Cho, J.H.; Okuma, A.; Al-Rubaye, D.; Intisar, E.; Junghans, R.P.; Wong, W.W. Engineering Axl specific CAR and SynNotch receptor for cancer therapy. Sci. Rep. 2018, 8, 3846. [Google Scholar] [CrossRef]
- Gianno, F.; Giovannoni, I.; Cafferata, B.; Diomedi-Camassei, F.; Minasi, S.; Barresi, S.; Buttarelli, F.R.; Alesi, V.; Cardoni, A.; Antonelli, M.; et al. Paediatric-type diffuse high-grade gliomas in the 5th CNS WHO Classification. Pathologica 2022, 114, 422–435. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.K.; Yoon, C.; Yi, B.C.; Tap, W.D.; Simon, M.C.; Yoon, S.S. Platelet-derived growth factor receptor-alpha and -beta promote cancer stem cell phenotypes in sarcomas. Oncogenesis 2018, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- Tap, W.D.; Wagner, A.J.; Schoffski, P.; Martin-Broto, J.; Krarup-Hansen, A.; Ganjoo, K.N.; Yen, C.C.; Abdul Razak, A.R.; Spira, A.; Kawai, A.; et al. Effect of Doxorubicin Plus Olaratumab vs Doxorubicin Plus Placebo on Survival in Patients with Advanced Soft Tissue Sarcomas: The Announce Randomized Clinical Trial. JAMA 2020, 323, 1266–1276. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Wang, J.; Wen, X.; Xu, B.; Que, Y.; Yu, K.; Xu, L.; Zhao, J.; Pan, Q.; Zhou, P.; et al. Chimeric antigen receptor-modified T-cell therapy for platelet-derived growth factor receptor alpha-positive rhabdomyosarcoma. Cancer 2020, 126, 2093–2100. [Google Scholar] [CrossRef]
- Jie, Y.; Liu, G.; Feng, L.; Li, Y.; E, M.; Wu, L.; Li, Y.; Rong, G.; Li, Y.; Wei, H.; et al. PTK7-Targeting CAR T-Cells for the Treatment of Lung Cancer and Other Malignancies. Front. Immunol. 2021, 12, 665970. [Google Scholar] [CrossRef]
- Maitland, M.L.; Sachdev, J.C.; Sharma, M.R.; Moreno, V.; Boni, V.; Kummar, S.; Stringer-Reasor, E.; Lakhani, N.; Moreau, A.R.; Xuan, D.; et al. First-in-Human Study of PF-06647020 (Cofetuzumab Pelidotin), an Antibody-Drug Conjugate Targeting Protein Tyrosine Kinase 7, in Advanced Solid Tumors. Clin. Cancer Res. 2021, 27, 4511–4520. [Google Scholar] [CrossRef]
- Park, S.; Shevlin, E.; Vedvyas, Y.; Zaman, M.; Park, S.; Hsu, Y.S.; Min, I.M.; Jin, M.M. Micromolar affinity CAR T cells to ICAM-1 achieves rapid tumor elimination while avoiding systemic toxicity. Sci. Rep. 2017, 7, 14366. [Google Scholar] [CrossRef]
- Wichert, S.; Juliusson, G.; Johansson, A.; Sonesson, E.; Teige, I.; Wickenberg, A.T.; Frendeus, B.; Korsgren, M.; Hansson, M. A single-arm, open-label, phase 2 clinical trial evaluating disease response following treatment with BI-505, a human anti-intercellular adhesion molecule-1 monoclonal antibody, in patients with smoldering multiple myeloma. PLoS ONE 2017, 12, e0171205. [Google Scholar] [CrossRef]
- Pang, L.; Ren, F.; Xu, X.; Fu, L.; Wang, T.; Guo, Z. Construction and Characterization of Cadherin 6 (CDH6)-Targeting Chimeric Antigen Receptor (CAR) Modified T Cells. J. Environ. Pathol. Toxicol. Oncol. 2022, 41, 55–71. [Google Scholar] [CrossRef]
- Schoffski, P.; Concin, N.; Suarez, C.; Subbiah, V.; Ando, Y.; Ruan, S.; Wagner, J.P.; Mansfield, K.; Zhu, X.; Origuchi, S.; et al. A Phase 1 Study of a CDH6-Targeting Antibody-Drug Conjugate in Patients with Advanced Solid Tumors with Evaluation of Inflammatory and Neurological Adverse Events. Oncol. Res. Treat. 2021, 44, 547–556. [Google Scholar] [CrossRef]
- Feng, Z.; He, X.; Zhang, X.; Wu, Y.; Xing, B.; Knowles, A.; Shan, Q.; Miller, S.; Hojnacki, T.; Ma, J.; et al. Potent suppression of neuroendocrine tumors and gastrointestinal cancers by CDH17CAR T cells without toxicity to normal tissues. Nat. Cancer 2022, 3, 581–594. [Google Scholar] [CrossRef]
- Chatterjee, S.; Sinha, S.; Kundu, C.N. Nectin cell adhesion molecule-4 (NECTIN-4): A potential target for cancer therapy. Eur. J. Pharmacol. 2021, 911, 174516. [Google Scholar] [CrossRef]
- Li, F.; Zhao, S.; Wei, C.; Hu, Y.; Xu, T.; Xin, X.; Zhu, T.; Shang, L.; Ke, S.; Zhou, J.; et al. Development of Nectin4/FAP-targeted CAR-T cells secreting IL-7, CCL19, and IL-12 for malignant solid tumors. Front. Immunol. 2022, 13, 958082. [Google Scholar] [CrossRef]
- Powles, T.; Rosenberg, J.E.; Sonpavde, G.P.; Loriot, Y.; Duran, I.; Lee, J.L.; Matsubara, N.; Vulsteke, C.; Castellano, D.; Wu, C.; et al. Enfortumab Vedotin in Previously Treated Advanced Urothelial Carcinoma. N. Engl. J. Med. 2021, 384, 1125–1135. [Google Scholar] [CrossRef]
- Klumper, N.; Ralser, D.J.; Ellinger, J.; Roghmann, F.; Albrecht, J.; Below, E.; Alajati, A.; Sikic, D.; Breyer, J.; Bolenz, C.; et al. Membranous NECTIN-4 expression frequently decreases during metastatic spread of urothelial carcinoma and is associated with enfortumab vedotin resistance. Clin. Cancer Res. 2022. [Google Scholar] [CrossRef]
- Senbanjo, L.T.; Chellaiah, M.A. CD44: A Multifunctional Cell Surface Adhesion Receptor Is a Regulator of Progression and Metastasis of Cancer Cells. Front. Cell Dev. Biol. 2017, 5, 18. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, K.; Hackert, T.; Zoller, M. CD44/CD44v6 a Reliable Companion in Cancer-Initiating Cell Maintenance and Tumor Progression. Front. Cell Dev. Biol. 2018, 6, 97. [Google Scholar] [CrossRef]
- Casucci, M.; Nicolis di Robilant, B.; Falcone, L.; Camisa, B.; Norelli, M.; Genovese, P.; Gentner, B.; Gullotta, F.; Ponzoni, M.; Bernardi, M.; et al. CD44v6-targeted T cells mediate potent antitumor effects against acute myeloid leukemia and multiple myeloma. Blood 2013, 122, 3461–3472. [Google Scholar] [CrossRef] [PubMed]
- Hekele, A.; Dall, P.; Moritz, D.; Wels, W.; Groner, B.; Herrlich, P.; Ponta, H. Growth retardation of tumors by adoptive transfer of cytotoxic T lymphocytes reprogrammed by CD44v6-specific scFv:zeta-chimera. Int. J. Cancer 1996, 68, 232–238. [Google Scholar] [CrossRef]
- Tang, L.; Huang, H.; Tang, Y.; Li, Q.; Wang, J.; Li, D.; Zhong, Z.; Zou, P.; You, Y.; Cao, Y.; et al. CD44v6 chimeric antigen receptor T cell specificity towards AML with FLT3 or DNMT3A mutations. Clin. Transl. Med. 2022, 12, e1043. [Google Scholar] [CrossRef] [PubMed]
- Haist, C.; Schulte, E.; Bartels, N.; Bister, A.; Poschinski, Z.; Ibach, T.C.; Geipel, K.; Wiek, C.; Wagenmann, M.; Monzel, C.; et al. CD44v6-targeted CAR T-cells specifically eliminate CD44 isoform 6 expressing head/neck squamous cell carcinoma cells. Oral Oncol. 2021, 116, 105259. [Google Scholar] [CrossRef] [PubMed]
- Tijink, B.M.; Buter, J.; de Bree, R.; Giaccone, G.; Lang, M.S.; Staab, A.; Leemans, C.R.; van Dongen, G.A. A phase I dose escalation study with anti-CD44v6 bivatuzumab mertansine in patients with incurable squamous cell carcinoma of the head and neck or esophagus. Clin. Cancer Res. 2006, 12, 6064–6072. [Google Scholar] [CrossRef] [PubMed]
- Riechelmann, H.; Sauter, A.; Golze, W.; Hanft, G.; Schroen, C.; Hoermann, K.; Erhardt, T.; Gronau, S. Phase I trial with the CD44v6-targeting immunoconjugate bivatuzumab mertansine in head and neck squamous cell carcinoma. Oral Oncol. 2008, 44, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Whilding, L.M.; Parente-Pereira, A.C.; Zabinski, T.; Davies, D.M.; Petrovic, R.M.G.; Kao, Y.V.; Saxena, S.A.; Romain, A.; Costa-Guerra, J.A.; Violette, S.; et al. Targeting of Aberrant alphavbeta6 Integrin Expression in Solid Tumors Using Chimeric Antigen Receptor-Engineered T Cells. Mol. Ther. 2017, 25, 259–273. [Google Scholar] [CrossRef]
- Pameijer, C.R.; Navanjo, A.; Meechoovet, B.; Wagner, J.R.; Aguilar, B.; Wright, C.L.; Chang, W.C.; Brown, C.E.; Jensen, M.C. Conversion of a tumor-binding peptide identified by phage display to a functional chimeric T cell antigen receptor. Cancer Gene Ther. 2007, 14, 91–97. [Google Scholar] [CrossRef]
- Phanthaphol, N.; Somboonpatarakun, C.; Suwanchiwasiri, K.; Chieochansin, T.; Sujjitjoon, J.; Wongkham, S.; Maher, J.; Junking, M.; Yenchitsomanus, P.T. Chimeric Antigen Receptor T Cells Targeting Integrin alphavbeta6 Expressed on Cholangiocarcinoma Cells. Front. Oncol. 2021, 11, 657868. [Google Scholar] [CrossRef]
- Muliaditan, T.; Halim, L.; Whilding, L.M.; Draper, B.; Achkova, D.Y.; Kausar, F.; Glover, M.; Bechman, N.; Arulappu, A.; Sanchez, J.; et al. Synergistic T cell signaling by 41BB and CD28 is optimally achieved by membrane proximal positioning within parallel chimeric antigen receptors. Cell Rep. Med. 2021, 2, 100457. [Google Scholar] [CrossRef]
- Larcombe-Young, D.; Whilding, L.; Davies, D.M.; Draper, B.; Bechman, N.; Maher, J. Generation of human parallel chimeric antigen receptor (pCAR) T cells to achieve synergistic T cell co-stimulation. STAR Protoc. 2022, 3, 101414. [Google Scholar] [CrossRef]
- Whilding, L.M.; Halim, L.; Draper, B.; Parente-Pereira, A.C.; Zabinski, T.; Davies, D.M.; Maher, J. CAR T-Cells Targeting the Integrin alphavbeta6 and Co-Expressing the Chemokine Receptor CXCR2 Demonstrate Enhanced Homing and Efficacy against Several Solid Malignancies. Cancers 2019, 11, 674. [Google Scholar] [CrossRef]
- Hollebecque, A.; Lopez, J.; Piha-Paul, S.; Dowlati, A.; Patnaik, A.; Galvao, V.; Buckorny, B.; Sehgal, K.; Kingsley, E.; Sanborn, R.; et al. 731 A first-in-human trial of an integrin beta-6 targeted antibody-drug conjugate (ADC), SGN-B6A, in patients with advanced solid tumors: Interim results of a phase 1 study (SGNB6A-001). J. Immunother. Cancer 2022, 10, A763. [Google Scholar] [CrossRef]
- Cobb, D.A.; de Rossi, J.; Liu, L.; An, E.; Lee, D.W. Targeting of the alpha(v) beta(3) integrin complex by CAR-T cells leads to rapid regression of diffuse intrinsic pontine glioma and glioblastoma. J. Immunother. Cancer 2022, 10, e003816. [Google Scholar] [CrossRef] [PubMed]
- Wallstabe, L.; Mades, A.; Frenz, S.; Einsele, H.; Rader, C.; Hudecek, M. CAR T cells targeting alpha(v)beta(3) integrin are effective against advanced cancer in preclinical models. Adv. Cell Gene Ther. 2018, 1, e11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, E.; Gu, J.; Xue, J.; Lin, C.; Liu, C.; Li, M.; Hao, J.; Setrerrahmane, S.; Chi, X.; Qi, W.; et al. Accurate control of dual-receptor-engineered T cell activity through a bifunctional anti-angiogenic peptide. J. Hematol. Oncol. 2018, 11, 44. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.; Whitworth, K.; Jinks, E.; Nagy, Z.; Bicknell, R.; Lee, S.P. An evaluation of the tumour endothelial marker CLEC14A as a therapeutic target in solid tumours. J. Pathol. Clin. Res. 2020, 6, 308–319. [Google Scholar] [CrossRef]
- Zhuang, X.; Maione, F.; Robinson, J.; Bentley, M.; Kaul, B.; Whitworth, K.; Jumbu, N.; Jinks, E.; Bystrom, J.; Gabriele, P.; et al. CAR T cells targeting tumor endothelial marker CLEC14A inhibit tumor growth. JCI Insight 2020, 5, e138808. [Google Scholar] [CrossRef]
- Chen, J.; Li, Z.; Zhao, Q.; Chen, L. Roles of apelin/APJ system in cancer: Biomarker, predictor, and emerging therapeutic target. J. Cell Physiol. 2022, 237, 3734–3751. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, F.; Zhong, J.; Zhu, T.; Zheng, Y.; Zhao, T.; Xie, Q.; Ma, F.; Li, R.; Tang, Q.; et al. Using apelin-based synthetic Notch receptors to detect angiogenesis and treat solid tumors. Nat. Commun. 2020, 11, 2163. [Google Scholar] [CrossRef]
- White, E.S.; Muro, A.F. Fibronectin splice variants: Understanding their multiple roles in health and disease using engineered mouse models. IUBMB Life 2011, 63, 538–546. [Google Scholar] [CrossRef]
- Ventura, E.; Weller, M.; Macnair, W.; Eschbach, K.; Beisel, C.; Cordazzo, C.; Claassen, M.; Zardi, L.; Burghardt, I. TGF-beta induces oncofetal fibronectin that, in turn, modulates TGF-beta superfamily signaling in endothelial cells. J. Cell Sci. 2018, 131, jcs209619. [Google Scholar] [CrossRef]
- Lieverse, R.I.Y.; Marcus, D.; van der Wiel, A.M.A.; Van Limbergen, E.J.; Theys, J.; Yaromina, A.; Lambin, P.; Dubois, L.J. Human fibronectin extra domain B as a biomarker for targeted therapy in cancer. Mol. Oncol. 2020, 14, 1555–1568. [Google Scholar] [CrossRef]
- Hooper, A.T.; Marquette, K.; Chang, C.B.; Golas, J.; Jain, S.; Lam, M.H.; Guffroy, M.; Leal, M.; Falahatpisheh, H.; Mathur, D.; et al. Anti-Extra Domain B Splice Variant of Fibronectin Antibody-Drug Conjugate Eliminates Tumors with Enhanced Efficacy When Combined with Checkpoint Blockade. Mol. Cancer Ther. 2022, 21, 1462–1472. [Google Scholar] [CrossRef]
- Mhawech, P.; Dulguerov, P.; Assaly, M.; Ares, C.; Allal, A.S. EB-D fibronectin expression in squamous cell carcinoma of the head and neck. Oral Oncol. 2005, 41, 82–88. [Google Scholar] [CrossRef]
- Li, F.; Hooper, A.T.; Golas, J.; Chang, C.B.; Neubert, H.; King, L. Evaluation of EDB Fibronectin in Plasma, Patient-Derived Xenograft Formalin-Fixed Paraffin-Embedded and Fresh Frozen Tumor Tissues Using Immunoaffinity LC-MS/MS. J. Proteome Res. 2022, 21, 2331–2340. [Google Scholar] [CrossRef] [PubMed]
- Petrini, I.; Sollini, M.; Bartoli, F.; Barachini, S.; Montali, M.; Pardini, E.; Burzi, I.S.; Erba, P.A. ED-B-Containing Isoform of Fibronectin in Tumor Microenvironment of Thymomas: A Target for a Theragnostic Approach. Cancers 2022, 14, 2592. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.; Wickman, E.; Shaw, T.I.; Anido, A.A.; Langfitt, D.; Zhang, J.; Porter, S.N.; Pruett-Miller, S.M.; Tillman, H.; Krenciute, G.; et al. Antitumor Effects of CAR T Cells Redirected to the EDB Splice Variant of Fibronectin. Cancer Immunol. Res. 2021, 9, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, C.; Yang, Z.; Yin, H. CAR-T-Cell Therapy for Solid Tumors Positive for Fibronectin Extra Domain B. Cells 2022, 11, 2863. [Google Scholar] [CrossRef] [PubMed]
- Mock, J.; Stringhini, M.; Villa, A.; Weller, M.; Weiss, T.; Neri, D. An engineered 4-1BBL fusion protein with “activity on demand”. Proc. Natl. Acad. Sci. USA 2020, 117, 31780–31788. [Google Scholar] [CrossRef] [PubMed]
- Kumra, H.; Reinhardt, D.P. Fibronectin-targeted drug delivery in cancer. Adv. Drug Deliv. Rev. 2016, 97, 101–110. [Google Scholar] [CrossRef]
- Martin-Otal, C.; Lasarte-Cia, A.; Serrano, D.; Casares, N.; Conde, E.; Navarro, F.; Sanchez-Moreno, I.; Gorraiz, M.; Sarrion, P.; Calvo, A.; et al. Targeting the extra domain A of fibronectin for cancer therapy with CAR-T cells. J. Immunother. Cancer 2022, 10, e004479. [Google Scholar] [CrossRef]
- Lo, A.S.; Ma, Q.; Liu, D.L.; Junghans, R.P. Anti-GD3 chimeric sFv-CD28/T-cell receptor zeta designer T cells for treatment of metastatic melanoma and other neuroectodermal tumors. Clin. Cancer Res. 2010, 16, 2769–2780. [Google Scholar] [CrossRef]
- Nakakuma, H.; Horikawa, K.; Kawaguchi, T.; Hidaka, M.; Nagakura, S.; Hirai, S.; Kageshita, T.; Ono, T.; Kagimoto, T.; Iwamori, M.; et al. Common phenotypic expression of gangliosides GM3 and GD3 in normal human tissues and neoplastic skin lesions. Jpn. J. Clin. Oncol. 1992, 22, 308–312. [Google Scholar] [PubMed]
- Kasprowicz, A.; Sophie, G.D.; Lagadec, C.; Delannoy, P. Role of GD3 Synthase ST8Sia I in Cancers. Cancers 2022, 14, 1299. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.; Sumughan, S.; Dellacecca, E.R.; Shivde, R.S.; Lancki, N.; Mukhatayev, Z.; Vaca, C.C.; Han, F.; Barse, L.; Henning, S.W.; et al. Benign tumors in TSC are amenable to treatment by GD3 CAR T cells in mice. JCI Insight 2021, 6, e152014. [Google Scholar] [CrossRef] [PubMed]
- Adachi, K.; Kano, Y.; Nagai, T.; Okuyama, N.; Sakoda, Y.; Tamada, K. IL-7 and CCL19 expression in CAR-T cells improves immune cell infiltration and CAR-T cell survival in the tumor. Nat. Biotechnol. 2018, 36, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Sachdev, J.C.; Bauer, T.M.; Chawla, S.P.; Pant, S.; Patnaik, A.; Wainberg, Z.A.; Inamdar, S.P.; Marina, N.; Sun, S.; Schmidt, M.; et al. Phase 1a/1b study of first-in-class B7-H4 antibody, FPA150, as monotherapy in patients with advanced solid tumors. J. Clin. Oncol. 2019, 37, 2529. [Google Scholar] [CrossRef]
- Smith, J.B.; Lanitis, E.; Dangaj, D.; Buza, E.; Poussin, M.; Stashwick, C.; Scholler, N.; Powell, D.J., Jr. Tumor Regression and Delayed Onset Toxicity Following B7-H4 CAR T Cell Therapy. Mol. Ther. 2016, 24, 1987–1999. [Google Scholar] [CrossRef]
- Pinheiro, P.F.; Justino, G.C.; Marques, M.M. NKp30—A prospective target for new cancer immunotherapy strategies. Br. J. Pharmacol. 2020, 177, 4563–4580. [Google Scholar] [CrossRef]
- Zhang, T.; Wu, M.R.; Sentman, C.L. An NKp30-Based Chimeric Antigen Receptor Promotes T Cell Effector Functions and Antitumor Efficacy In Vivo. J. Immunol. 2012, 189, 2290–2299. [Google Scholar] [CrossRef]
- Falchook, G.; Spigel, D.; Patel, M.; Bashir, B.; Ulahannan, S.; Duffy, C.; Maier, D.; Azuma, H. 480 A first-in-human phase I dose-escalation trial of the B7-H6/CD3 T-cell engager BI 765049 ± ezabenlimab (BI 754091) in patients with advanced solid tumors expressing B7-H6. J. Immunother. Cancer 2021, 9, A510. [Google Scholar] [CrossRef]
- Drake, C.; Garcia, A.; Chang, C.-W.; Yang, B.-H.; Ibitokou, S.; Pride, C.; Markov, S.; Liao, A.; Pribadi, M.; Pan, Y.; et al. 326 Off-the-shelf iPSC-derived CAR-T cells targeting KLK2 demonstrate prolonged tumor control and survival in xenograft models of prostate cancer. J. Immunother. Cancer 2022, 10, A343. [Google Scholar] [CrossRef]
- Xu, M.; Evans, L.; Bizzaro, C.L.; Quaglia, F.; Verrillo, C.E.; Li, L.; Stieglmaier, J.; Schiewer, M.J.; Languino, L.R.; Kelly, W.K. STEAP1-4 (Six-Transmembrane Epithelial Antigen of the Prostate 1-4) and Their Clinical Implications for Prostate Cancer. Cancers 2022, 14, 4034. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Lorvik, K.B.; Jin, Y.; Beck, C.; Sike, A.; Persiconi, I.; Kvaloy, E.; Saatcioglu, F.; Dunn, C.; Kyte, J.A. Development of STEAP1 targeting chimeric antigen receptor for adoptive cell therapy against cancer. Mol. Ther. Oncolytics 2022, 26, 189–206. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Bergsland, E.; Aggarwal, R.; Aparicio, A.; Beltran, H.; Crabtree, J.S.; Hann, C.L.; Ibrahim, T.; Byers, L.A.; Sasano, H.; et al. DLL3 as an Emerging Target for the Treatment of Neuroendocrine Neoplasms. Oncologist 2022, 27, 940–951. [Google Scholar] [CrossRef]
- Blackhall, F.; Jao, K.; Greillier, L.; Cho, B.C.; Penkov, K.; Reguart, N.; Majem, M.; Nackaerts, K.; Syrigos, K.; Hansen, K.; et al. Efficacy and Safety of Rovalpituzumab Tesirine Compared With Topotecan as Second-Line Therapy in DLL3-High SCLC: Results From the Phase 3 TAHOE Study. J. Thorac. Oncol. 2021, 16, 1547–1558. [Google Scholar] [CrossRef]
- Johnson, M.L.; Dy, G.K.; Mamdani, H.; Dowlati, A.; Schoenfeld, A.J.; Pacheco, J.M.; Sanborn, R.E.; Menon, S.P.; Santiago, L.; Yaron, Y.; et al. Interim results of an ongoing phase 1/2a study of HPN328, a tri-specific, half-life extended, DLL3-targeting, T-cell engager, in patients with small cell lung cancer and other neuroendocrine cancers. J. Clin. Oncol. 2022, 40, 8566. [Google Scholar] [CrossRef]
- Owonikoko, T.K.; Champiat, S.; Johnson, M.L.; Govindan, R.; Izumi, H.; Lai, W.V.V.; Borghaei, H.; Boyer, M.J.; Boosman, R.J.; Hummel, H.-D.; et al. Updated results from a phase 1 study of AMG 757, a half-life extended bispecific T-cell engager (BiTE) immuno-oncology therapy against delta-like ligand 3 (DLL3), in small cell lung cancer (SCLC). J. Clin. Oncol. 2021, 39, 8510. [Google Scholar] [CrossRef]
- Pittaway, J.F.H.; Lipsos, C.; Mariniello, K.; Guasti, L. The role of delta-like non-canonical Notch ligand 1 (DLK1) in cancer. Endocr. Relat. Cancer 2021, 28, R271–R287. [Google Scholar] [CrossRef]
- Zhai, Y.; He, K.; Huang, L.; Shang, X.; Wang, G.; Yuan, G.; Han, Z.G. DLK1-directed chimeric antigen receptor T-cell therapy for hepatocellular carcinoma. Liver Int. 2022, 42, 2524–2537. [Google Scholar] [CrossRef]
- Baldwin, E.T.; Gotte, M.; Tchesnokov, E.P.; Arnold, E.; Hagel, M.; Nichols, C.; Dossang, P.; Lamers, M.; Wan, P.; Steinbacher, S.; et al. Human endogenous retrovirus-K (HERV-K) reverse transcriptase (RT) structure and biochemistry reveals remarkable similarities to HIV-1 RT and opportunities for HERV-K-specific inhibition. Proc. Natl. Acad. Sci. USA 2022, 119, e2200260119. [Google Scholar] [CrossRef]
- Dervan, E.; Bhattacharyya, D.D.; McAuliffe, J.D.; Khan, F.H.; Glynn, S.A. Ancient Adversary—HERV-K (HML-2) in Cancer. Front. Oncol. 2021, 11, 658489. [Google Scholar] [CrossRef]
- Burn, A.; Roy, F.; Freeman, M.; Coffin, J.M. Widespread expression of the ancient HERV-K (HML-2) provirus group in normal human tissues. PLoS Biol. 2022, 20, e3001826. [Google Scholar] [CrossRef]
- Krishnamurthy, J.; Rabinovich, B.A.; Mi, T.; Switzer, K.C.; Olivares, S.; Maiti, S.N.; Plummer, J.B.; Singh, H.; Kumaresan, P.R.; Huls, H.M.; et al. Genetic Engineering of T Cells to Target HERV-K, an Ancient Retrovirus on Melanoma. Clin. Cancer Res. 2015, 21, 3241–3251. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Krishnamurthy, J.; Wei, Y.; Li, M.; Hunt, K.; Johanning, G.L.; Cooper, L.J.; Wang-Johanning, F. Chimeric antigen receptor T cells targeting HERV-K inhibit breast cancer and its metastasis through downregulation of Ras. Oncoimmunology 2015, 4, e1047582. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Zhou, Y.; Li, W.; Tang, Q.; Chen, R.; Zhu, J.; Feng, Z. T cells expressing a LMP1-specific chimeric antigen receptor mediate antitumor effects against LMP1-positive nasopharyngeal carcinoma cells in vitro and in vivo. J. Biomed. Res. 2014, 28, 468–475. [Google Scholar] [CrossRef]
- He, H.Z.; Xing, Y.Y.; Zhang, Y.; Xu, Y.X.; Tian, Z.; Xing, H.Y.; Tang, K.J.; Rao, Q.; Wang, J.X.; Wang, M. Functional investigation of chimeric antigen receptor T cells targeting LMP1 antigen. Zhonghua Xue Ye Xue Za Zhi 2022, 43, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Harada, S.; Ando, M.; Ando, J.; Ishii, M.; Yamaguchi, T.; Yamazaki, S.; Toyota, T.; Ohara, K.; Ohtaka, M.; Nakanishi, M.; et al. Dual-antigen targeted iPSC-derived chimeric antigen receptor-T cell therapy for refractory lymphoma. Mol. Ther. 2022, 30, 534–549. [Google Scholar] [CrossRef]
- Tang, X.; Tang, Q.; Mao, Y.; Huang, X.; Jia, L.; Zhu, J.; Feng, Z. CD137 Co-Stimulation Improves The Antitumor Effect Of LMP1-Specific Chimeric Antigen Receptor T Cells In Vitro And In Vivo. Onco Targets Ther. 2019, 12, 9341–9350. [Google Scholar] [CrossRef]
- Wang, S.; Qiu, Y.; Bai, B. The Expression, Regulation, and Biomarker Potential of Glypican-1 in Cancer. Front. Oncol. 2019, 9, 614. [Google Scholar] [CrossRef]
- Kato, D.; Yaguchi, T.; Iwata, T.; Katoh, Y.; Morii, K.; Tsubota, K.; Takise, Y.; Tamiya, M.; Kamada, H.; Akiba, H.; et al. GPC1 specific CAR-T cells eradicate established solid tumor without adverse effects and synergize with anti-PD-1 Ab. Elife 2020, 9, e49392. [Google Scholar] [CrossRef]
- Bosse, K.R.; Raman, P.; Zhu, Z.; Lane, M.; Martinez, D.; Heitzeneder, S.; Rathi, K.S.; Kendsersky, N.M.; Randall, M.; Donovan, L.; et al. Identification of GPC2 as an Oncoprotein and Candidate Immunotherapeutic Target in High-Risk Neuroblastoma. Cancer Cell 2017, 32, 295–309.e212. [Google Scholar] [CrossRef]
- Heitzeneder, S.; Bosse, K.R.; Zhu, Z.; Zhelev, D.; Majzner, R.G.; Radosevich, M.T.; Dhingra, S.; Sotillo, E.; Buongervino, S.; Pascual-Pasto, G.; et al. GPC2-CAR T cells tuned for low antigen density mediate potent activity against neuroblastoma without toxicity. Cancer Cell 2022, 40, 53–69.e9. [Google Scholar] [CrossRef]
- Yekehfallah, V.; Pahlavanneshan, S.; Sayadmanesh, A.; Momtahan, Z.; Ma, B.; Basiri, M. Generation and Functional Characterization of PLAP CAR-T Cells against Cervical Cancer Cells. Biomolecules 2022, 12, 1296. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Berahovich, R.; Zhou, H.; Liu, X.; Li, F.; Xu, S.; Wei, Y.; Ouaret, D.; Bodmer, W.; Wu, L.; et al. PLAP -CAR T cells mediate high specific cytotoxicity against colon cancer cells. Front. Biosci. 2020, 25, 1765–1786. [Google Scholar] [CrossRef]
- Chen, H.; Wei, F.; Yin, M.; Zhao, Q.; Liu, Z.; Yu, B.; Huang, Z. CD27 enhances the killing effect of CAR T cells targeting trophoblast cell surface antigen 2 in the treatment of solid tumors. Cancer Immunol. Immunother. 2021, 70, 2059–2071. [Google Scholar] [CrossRef] [PubMed]
- Heist, R.S.; Guarino, M.J.; Masters, G.; Purcell, W.T.; Starodub, A.N.; Horn, L.; Scheff, R.J.; Bardia, A.; Messersmith, W.A.; Berlin, J.; et al. Therapy of Advanced Non-Small-Cell Lung Cancer with an SN-38-Anti-Trop-2 Drug Conjugate, Sacituzumab Govitecan. J. Clin. Oncol. 2017, 35, 2790–2797. [Google Scholar] [CrossRef] [PubMed]
- Wahby, S.; Fashoyin-Aje, L.; Osgood, C.L.; Cheng, J.; Fiero, M.H.; Zhang, L.; Tang, S.; Hamed, S.S.; Song, P.; Charlab, R.; et al. FDA Approval Summary: Accelerated Approval of Sacituzumab Govitecan-hziy for Third-line Treatment of Metastatic Triple-negative Breast Cancer. Clin. Cancer Res. 2021, 27, 1850–1854. [Google Scholar] [CrossRef]
- Matusz-Fisher, A.; Tan, A.R. Sacituzumab govitecan and other antibody-drug conjugates targeting trophoblast cell-surface antigen 2 (Trop-2) in breast cancer. Ann. Transl. Med. 2022, 10, 1031. [Google Scholar] [CrossRef] [PubMed]
- Meric-Bernstam, F.; Spira, A.I.; Lisberg, A.E.; Sands, J.; Yamamoto, N.; Johnson, M.L.; Yoh, K.; Garon, E.B.; Heist, R.S.; Petrich, A.; et al. TROPION-PanTumor01: Dose analysis of the TROP2-directed antibody-drug conjugate (ADC) datopotamab deruxtecan (Dato-DXd, DS-1062) for the treatment (Tx) of advanced or metastatic non-small cell lung cancer (NSCLC). J. Clin. Oncol. 2021, 39, 9058. [Google Scholar] [CrossRef]
- Zhao, W.; Jia, L.; Zhang, M.; Huang, X.; Qian, P.; Tang, Q.; Zhu, J.; Feng, Z. The killing effect of novel bi-specific Trop2/PD-L1 CAR-T cell targeted gastric cancer. Am. J. Cancer Res. 2019, 9, 1846–1856. [Google Scholar]
- Zhu, H.; Fang, X.; Tuhin, I.J.; Tan, J.; Ye, J.; Jia, Y.; Xu, N.; Kang, L.; Li, M.; Lou, X.; et al. CAR T cells equipped with a fully human scFv targeting Trop2 can be used to treat pancreatic cancer. J. Cancer Res. Clin. Oncol. 2022, 148, 2261–2274. [Google Scholar] [CrossRef]
- Anna, F.; Bole-Richard, E.; LeMaoult, J.; Escande, M.; Lecomte, M.; Certoux, J.M.; Souque, P.; Garnache, F.; Adotevi, O.; Langlade-Demoyen, P.; et al. First immunotherapeutic CAR-T cells against the immune checkpoint protein HLA-G. J. Immunother. Cancer 2021, 9, e001998. [Google Scholar] [CrossRef]
- Martin-Villa, J.M.; Vaquero-Yuste, C.; Molina-Alejandre, M.; Juarez, I.; Suarez-Trujillo, F.; Lopez-Nares, A.; Palacio-Gruber, J.; Barrera-Gutierrez, L.; Fernandez-Cruz, E.; Rodriguez-Sainz, C.; et al. HLA-G: Too Much or Too Little? Role in Cancer and Autoimmune Disease. Front. Immunol. 2022, 13, 796054. [Google Scholar] [CrossRef]
- Obermajer, N.; Zwolak, A.; Van De Ven, K.; Versmissen, S.; Brajic, A.; Petley, T.; Weinstock, D.; Aligo, J.; Yi, F.; Jarantow, S.; et al. Abstract ND07: JNJ-78306358: A first-in-class bispecific T cell redirecting HLA-G antibody. Cancer Res. 2022, 82, ND07. [Google Scholar] [CrossRef]
- Harrop, R.; O’Neill, E.; Stern, P.L. Cancer stem cell mobilization and therapeutic targeting of the 5T4 oncofetal antigen. Ther. Adv. Vaccines Immunother. 2019, 7, 2515135518821623. [Google Scholar] [CrossRef] [PubMed]
- Owens, G.L.; Sheard, V.E.; Kalaitsidou, M.; Blount, D.; Lad, Y.; Cheadle, E.J.; Edmondson, R.J.; Kooner, G.; Gilham, D.E.; Harrop, R. Preclinical Assessment of CAR T-Cell Therapy Targeting the Tumor Antigen 5T4 in Ovarian Cancer. J. Immunother. 2018, 41, 130–140. [Google Scholar] [CrossRef]
- Guo, C.; Dong, E.; Lai, Q.; Zhou, S.; Zhang, G.; Wu, M.; Yue, X.; Tao, Y.; Peng, Y.; Ali, J.; et al. Effective antitumor activity of 5T4-specific CAR-T cells against ovarian cancer cells in vitro and xenotransplanted tumors in vivo. MedComm 2020, 1, 338–350. [Google Scholar] [CrossRef]
- Shapiro, G.I.; Vaishampayan, U.N.; LoRusso, P.; Barton, J.; Hua, S.; Reich, S.D.; Shazer, R.; Taylor, C.T.; Xuan, D.; Borghaei, H. First-in-human trial of an anti-5T4 antibody-monomethylauristatin conjugate, PF-06263507, in patients with advanced solid tumors. Investig. New Drugs 2017, 35, 315–323. [Google Scholar] [CrossRef]
- Kobayashi, K.; Sakamoto, J.; Kito, T.; Yamamura, Y.; Koshikawa, T.; Fujita, M.; Watanabe, T.; Nakazato, H. Lewis blood group-related antigen expression in normal gastric epithelium, intestinal metaplasia, gastric adenoma, and gastric carcinoma. Am. J. Gastroenterol. 1993, 88, 919–924. [Google Scholar] [PubMed]
- Ritchie, D.S.; Neeson, P.J.; Khot, A.; Peinert, S.; Tai, T.; Tainton, K.; Chen, K.; Shin, M.; Wall, D.M.; Honemann, D.; et al. Persistence and efficacy of second generation CAR T cell against the LeY antigen in acute myeloid leukemia. Mol. Ther. 2013, 21, 2122–2129. [Google Scholar] [CrossRef]
- Testa, L.; Mano, M.; Arai, R.J.; Bonadio, R.C.; Serrano, S.V.; Zorzetto, M.M.C.; Crocamo, S.; Smaletz, O.; Freitas-Junior, R.; Hoff, P.M. Phase II trial of humanized anti-Lewis Y monoclonal antibody for advanced hormone receptor-positive breast cancer that progressed following endocrine therapy. Clinics 2021, 76, e3146. [Google Scholar] [CrossRef]
- Yuan, J. Abstract 3766: MG7-car, a first-in-class T-cell therapy for gastric cancer. Cancer Res. 2017, 77, 3766. [Google Scholar] [CrossRef]
- Yuan, J. Abstract CT056: The first-in-human clinical trial of MG7-CART for metastasis colon cancer. Cancer Res. 2018, 78, CT506. [Google Scholar] [CrossRef]
- Bhoj, V.G.; Li, L.; Parvathaneni, K.; Zhang, Z.; Kacir, S.; Arhontoulis, D.; Zhou, K.; McGettigan-Croce, B.; Nunez-Cruz, S.; Gulendran, G.; et al. Adoptive T cell immunotherapy for medullary thyroid carcinoma targeting GDNF family receptor alpha 4. Mol. Ther. Oncolytics 2021, 20, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Lin, W.; Wen, L.; Li, G. Lgr5 in cancer biology: Functional identification of Lgr5 in cancer progression and potential opportunities for novel therapy. Stem Cell Res. Ther. 2019, 10, 219. [Google Scholar] [CrossRef]
- Wang, W.; Bandara, V.; Lokman, N.; Napoli, S.; Gundsambuu, B.; Oehler, M.; Barry, S.C.; Ricciardelli, C. LGR5 CAR-T cells: A novel potential treatment against high grade serous ovarian cancer. Cancer Res. 2022, 82, 5183. [Google Scholar] [CrossRef]
- Chen, H.-C.; Mueller, N.; Stott, K.; Rivers, E.; Kapeni, C.; Sauer, C.M.; Beke, F.; Walsh, S.; Ashman, N.; O’Brien, L.; et al. LGR5 targeting molecules as therapeutic agents for multiple cancer types. bioRxiv 2022, preprint. [Google Scholar] [CrossRef]
- Pegoraro, A.; De Marchi, E.; Adinolfi, E. P2X7 Variants in Oncogenesis. Cells 2021, 10, 189. [Google Scholar] [CrossRef]
- Lara, R.; Adinolfi, E.; Harwood, C.A.; Philpott, M.; Barden, J.A.; Di Virgilio, F.; McNulty, S. P2X7 in Cancer: From Molecular Mechanisms to Therapeutics. Front. Pharmacol. 2020, 11, 793. [Google Scholar] [CrossRef]
- Gilbert, S.M.; Oliphant, C.J.; Hassan, S.; Peille, A.L.; Bronsert, P.; Falzoni, S.; Di Virgilio, F.; McNulty, S.; Lara, R. ATP in the tumour microenvironment drives expression of nfP2X(7), a key mediator of cancer cell survival. Oncogene 2019, 38, 194–208. [Google Scholar] [CrossRef] [PubMed]
- Perales-Puchalt, A.; Svoronos, N.; Rutkowski, M.R.; Allegrezza, M.J.; Tesone, A.J.; Payne, K.K.; Wickramasinghe, J.; Nguyen, J.M.; O’Brien, S.W.; Gumireddy, K.; et al. Follicle-Stimulating Hormone Receptor Is Expressed by Most Ovarian Cancer Subtypes and Is a Safe and Effective Immunotherapeutic Target. Clin. Cancer Res. 2017, 23, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Urbanska, K.; Stashwick, C.; Poussin, M.; Powell, D.J., Jr. Follicle-Stimulating Hormone Receptor as a Target in the Redirected T-cell Therapy for Cancer. Cancer Immunol. Res. 2015, 3, 1130–1137. [Google Scholar] [CrossRef] [PubMed]
- Magee, M.S.; Kraft, C.L.; Abraham, T.S.; Baybutt, T.R.; Marszalowicz, G.P.; Li, P.; Waldman, S.A.; Snook, A.E. GUCY2C-directed CAR-T cells oppose colorectal cancer metastases without autoimmunity. Oncoimmunology 2016, 5, e1227897. [Google Scholar] [CrossRef]
- Magee, M.S.; Abraham, T.S.; Baybutt, T.R.; Flickinger, J.C., Jr.; Ridge, N.A.; Marszalowicz, G.P.; Prajapati, P.; Hersperger, A.R.; Waldman, S.A.; Snook, A.E. Human GUCY2C-Targeted Chimeric Antigen Receptor (CAR)-Expressing T Cells Eliminate Colorectal Cancer Metastases. Cancer Immunol. Res. 2018, 6, 509–516. [Google Scholar] [CrossRef]
- Kim, R.D.; Cleary, J.M.; Leal, A.D.; Parikh, A.R.; Ryan, D.P.; Wang, S.; Bahamon, B.; Gupta, N.; Moss, A.; Pye, J.; et al. A phase 1 study of TAK-164, an anti-guanylyl cyclase C (GCC) antibody-drug conjugate (ADC), in patients (pts) with advanced gastrointestinal (GI) cancers expressing GCC. J. Clin. Oncol. 2021, 39, 3050. [Google Scholar] [CrossRef]
- Li, H.; Zhou, X.; Wang, G.; Hua, D.; Li, S.; Xu, T.; Dong, M.; Cui, X.; Yang, X.; Wu, Y.; et al. CAR-T Cells Targeting TSHR Demonstrate Safety and Potent Preclinical Activity against Differentiated Thyroid Cancer. J. Clin. Endocrinol. Metab. 2022, 107, 1110–1126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wei, H.; Fan, L.; Fang, M.; He, X.; Lu, B.; Pang, Z. CLEC4s as Potential Therapeutic Targets in Hepatocellular Carcinoma Microenvironment. Front. Cell Dev. Biol. 2021, 9, 681372. [Google Scholar] [CrossRef] [PubMed]
- Willingham, S.B.; Volkmer, J.P.; Gentles, A.J.; Sahoo, D.; Dalerba, P.; Mitra, S.S.; Wang, J.; Contreras-Trujillo, H.; Martin, R.; Cohen, J.D.; et al. The CD47-signal regulatory protein alpha (SIRPa) interaction is a therapeutic target for human solid tumors. Proc. Natl. Acad. Sci. USA 2012, 109, 6662–6667. [Google Scholar] [CrossRef]
- La, H.T.; Tran, D.B.T.; Tran, H.M.; Nguyen, L.T. Third-Generation Anti-CD47-Specific CAR-T Cells Effectively Kill Cancer Cells and Reduce the Genes Expression in Lung Cancer Cell Metastasis. J. Immunol. Res. 2021, 2021, 5575260. [Google Scholar] [CrossRef]
- Shu, R.; Evtimov, V.J.; Hammett, M.V.; Nguyen, N.N.; Zhuang, J.; Hudson, P.J.; Howard, M.C.; Pupovac, A.; Trounson, A.O.; Boyd, R.L. Engineered CAR-T cells targeting TAG-72 and CD47 in ovarian cancer. Mol. Ther. Oncolytics 2021, 20, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Golubovskaya, V.; Berahovich, R.; Zhou, H.; Xu, S.; Harto, H.; Li, L.; Chao, C.C.; Mao, M.M.; Wu, L. CD47-CAR-T Cells Effectively Kill Target Cancer Cells and Block Pancreatic Tumor Growth. Cancers 2017, 9, 139. [Google Scholar] [CrossRef]
- Lakhani, N.; Orloff, M.; Fu, S.; Liu, Y.; Wang, Y.; Zhou, H.; Lin, K.; Liu, F.; Yan, S.; Patnaik, A. 295 First-in-human Phase I trial of IBI188, an anti-CD47 targeting monoclonal antibody, in patients with advanced solid tumors and lymphomas. J. Immunother. Cancer 2020, 8, A180. [Google Scholar] [CrossRef]
- Daver, N.G.; Vyas, P.; Kambhampati, S.; Malki, M.M.A.; Larson, R.A.; Asch, A.S.; Mannis, G.N.; Chai-Ho, W.; Tanaka, T.N.; Bradley, T.J.; et al. Tolerability and efficacy of the first-in-class anti-CD47 antibody magrolimab combined with azacitidine in frontline TP53m AML patients: Phase 1b results. J. Clin. Oncol. 2022, 40, 7020. [Google Scholar] [CrossRef]
- Fu, F.; Yang, X.; Zheng, M.; Zhao, Q.; Zhang, K.; Li, Z.; Zhang, H.; Zhang, S. Role of Transmembrane 4 L Six Family 1 in the Development and Progression of Cancer. Front. Mol. Biosci. 2020, 7, 202. [Google Scholar] [CrossRef]
- Rad, E.R.; Foeng, J.; McPeak, D.; Tyllis, T.; Abbott, C.; Bandara, V.; Napoli, S.; Gundsambuu, B.; Barry, S.; Sadlon, T.; et al. Abstract 5505: ADAM10-targeting CAR-T cells inhibit colon cancer cell growth in vivo. Cancer Res. 2022, 82, 5505. [Google Scholar] [CrossRef]
- Wang, D.; Starr, R.; Chang, W.C.; Aguilar, B.; Alizadeh, D.; Wright, S.L.; Yang, X.; Brito, A.; Sarkissian, A.; Ostberg, J.R.; et al. Chlorotoxin-directed CAR T cells for specific and effective targeting of glioblastoma. Sci. Transl. Med. 2020, 12, eaaw2672. [Google Scholar] [CrossRef] [PubMed]
- Pfister, K.; Radons, J.; Busch, R.; Tidball, J.G.; Pfeifer, M.; Freitag, L.; Feldmann, H.J.; Milani, V.; Issels, R.; Multhoff, G. Patient survival by Hsp70 membrane phenotype: Association with different routes of metastasis. Cancer 2007, 110, 926–935. [Google Scholar] [CrossRef]
- Bashiri Dezfouli, A.; Yazdi, M.; Benmebarek, M.R.; Schwab, M.; Michaelides, S.; Micciche, A.; Geerts, D.; Stangl, S.; Klapproth, S.; Wagner, E.; et al. CAR T Cells Targeting Membrane-Bound Hsp70 on Tumor Cells Mimic Hsp70-Primed NK Cells. Front. Immunol. 2022, 13, 883694. [Google Scholar] [CrossRef]
- Ishii, H.; Afify, S.M.; Hassan, G.; Salomon, D.S.; Seno, M. Cripto-1 as a Potential Target of Cancer Stem Cells for Immunotherapy. Cancers 2021, 13, 2491. [Google Scholar] [CrossRef]
- Bianco, C.; Salomon, D.S. Targeting the embryonic gene Cripto-1 in cancer and beyond. Expert Opin. Ther. Pat. 2010, 20, 1739–1749. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, N.S.; Goncalves, D.R.; Balbinot, B.; Visioli, F. Is GRP78 (Glucose-regulated protein 78) a prognostic biomarker in differents types of cancer? A systematic review and meta-analysis. Pathol. Res. Pract. 2023, 242, 154301. [Google Scholar] [CrossRef]
- Hebbar, N.; Epperly, R.; Vaidya, A.; Thanekar, U.; Moore, S.E.; Umeda, M.; Ma, J.; Patil, S.L.; Langfitt, D.; Huang, S.; et al. CAR T cells redirected to cell surface GRP78 display robust anti-acute myeloid leukemia activity and do not target hematopoietic progenitor cells. Nat. Commun. 2022, 13, 587. [Google Scholar] [CrossRef]
- Yu, W.; Zhang, H.; Yuan, Y.; Tang, J.; Chen, X.; Liu, T.; Zhao, X. Chimeric Antigen Receptor T Cells Targeting Cell Surface GRP78 to Eradicate Acute Myeloid Leukemia. Front. Cell Dev. Biol. 2022, 10, 928140. [Google Scholar] [CrossRef]
- Rasche, L.; Duell, J.; Castro, I.C.; Dubljevic, V.; Chatterjee, M.; Knop, S.; Hensel, F.; Rosenwald, A.; Einsele, H.; Topp, M.S.; et al. GRP78-directed immunotherapy in relapsed or refractory multiple myeloma—Results from a phase 1 trial with the monoclonal immunoglobulin M antibody PAT-SM6. Haematologica 2015, 100, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Landras, A.; Reger de Moura, C.; Jouenne, F.; Lebbe, C.; Menashi, S.; Mourah, S. CD147 Is a Promising Target of Tumor Progression and a Prognostic Biomarker. Cancers 2019, 11, 803. [Google Scholar] [CrossRef]
- Bian, H.; Zheng, Z.H.; Wei, D.; Wen, A.; Zhang, Z.; Lian, J.Q.; Kang, W.Z.; Hao, C.Q.; Wang, J.; Xie, R.H.; et al. Safety and efficacy of meplazumab in healthy volunteers and COVID-19 patients: A randomized phase 1 and an exploratory phase 2 trial. Signal Transduct. Target. Ther. 2021, 6, 194. [Google Scholar] [CrossRef]
- Zhang, R.Y.; Wei, D.; Liu, Z.K.; Yong, Y.L.; Wei, W.; Zhang, Z.Y.; Lv, J.J.; Zhang, Z.; Chen, Z.N.; Bian, H. Doxycycline Inducible Chimeric Antigen Receptor T Cells Targeting CD147 for Hepatocellular Carcinoma Therapy. Front. Cell Dev. Biol. 2019, 7, 233. [Google Scholar] [CrossRef] [PubMed]
- Tseng, H.C.; Xiong, W.; Badeti, S.; Yang, Y.; Ma, M.; Liu, T.; Ramos, C.A.; Dotti, G.; Fritzky, L.; Jiang, J.G.; et al. Efficacy of anti-CD147 chimeric antigen receptors targeting hepatocellular carcinoma. Nat. Commun. 2020, 11, 4810. [Google Scholar] [CrossRef]
- Chen, X.H.; Chen, R.; Shi, M.Y.; Tian, R.F.; Zhang, H.; Xin, Z.Q.; Chen, Z.N.; Wang, K. Chimeric antigen receptor T cells targeting CD147 for non-small cell lung cancer therapy. Transl. Oncol. 2022, 16, 101309. [Google Scholar] [CrossRef]
- Li, X.; Zhang, G.; Chen, Q.; Lin, Y.; Li, J.; Ruan, Q.; Chen, Y.; Yu, G.; Wan, X. CD317 Promotes the survival of cancer cells through apoptosis-inducing factor. J. Exp. Clin. Cancer Res. 2016, 35, 117. [Google Scholar] [CrossRef] [PubMed]
- Roth, P.; Haensch, L.; Myburgh, R.; Silginer, M.; Weiss, T.; Gramatzki, D.; Vasella, F.; Manz, M.; Weller, M.; Peipp, M. Identification of CD317 as a novel target for chimeric antigen receptor (CAR)-based immunotherapy for glioblastoma. Brain Tumor Res. Treat. 2022, 10, S7-1. Available online: https://btrt.org/pdf/10.14791/btrt.2022.10.F-1272 (accessed on 28 January 2023).
- Beard, R.E.; Zheng, Z.; Lagisetty, K.H.; Burns, W.R.; Tran, E.; Hewitt, S.M.; Abate-Daga, D.; Rosati, S.F.; Fine, H.A.; Ferrone, S.; et al. Multiple chimeric antigen receptors successfully target chondroitin sulfate proteoglycan 4 in several different cancer histologies and cancer stem cells. J. Immunother. Cancer 2014, 2, 25. [Google Scholar] [CrossRef] [PubMed]
- Leuci, V.; Donini, C.; Grignani, G.; Rotolo, R.; Mesiano, G.; Fiorino, E.; Gammaitoni, L.; D’Ambrosio, L.; Merlini, A.; Landoni, E.; et al. CSPG4-Specific CAR.CIK Lymphocytes as a Novel Therapy for the Treatment of Multiple Soft-Tissue Sarcoma Histotypes. Clin. Cancer Res. 2020, 26, 6321–6334. [Google Scholar] [CrossRef] [PubMed]
- Harrer, D.C.; Dorrie, J.; Schaft, N. CSPG4 as Target for CAR-T-Cell Therapy of Various Tumor Entities-Merits and Challenges. Int. J. Mol. Sci. 2019, 20, 5942. [Google Scholar] [CrossRef]
- Geldres, C.; Savoldo, B.; Hoyos, V.; Caruana, I.; Zhang, M.; Yvon, E.; Del Vecchio, M.; Creighton, C.J.; Ittmann, M.; Ferrone, S.; et al. T lymphocytes redirected against the chondroitin sulfate proteoglycan-4 control the growth of multiple solid tumors both in vitro and in vivo. Clin. Cancer Res. 2014, 20, 962–971. [Google Scholar] [CrossRef]
- Panagiotou, E.; Syrigos, N.K.; Charpidou, A.; Kotteas, E.; Vathiotis, I.A. CD24: A Novel Target for Cancer Immunotherapy. J. Pers. Med. 2022, 12, 1235. [Google Scholar] [CrossRef] [PubMed]
- Maliar, A.; Servais, C.; Waks, T.; Chmielewski, M.; Lavy, R.; Altevogt, P.; Abken, H.; Eshhar, Z. Redirected T cells that target pancreatic adenocarcinoma antigens eliminate tumors and metastases in mice. Gastroenterology 2012, 143, 1375–1384.e5. [Google Scholar] [CrossRef] [PubMed]
- Klapdor, R.; Wang, S.; Morgan, M.; Dork, T.; Hacker, U.; Hillemanns, P.; Buning, H.; Schambach, A. Characterization of a Novel Third-Generation Anti-CD24-CAR against Ovarian Cancer. Int. J. Mol. Sci. 2019, 20, 660. [Google Scholar] [CrossRef]
- Rodriguez-Garcia, A.; Sharma, P.; Poussin, M.; Boesteanu, A.C.; Minutolo, N.G.; Gitto, S.B.; Omran, D.K.; Robinson, M.K.; Adams, G.P.; Simpkins, F.; et al. CAR T Cells Targeting MISIIR for the Treatment of Ovarian Cancer and Other Gynecologic Malignancies. Mol. Ther. 2020, 28, 548–560. [Google Scholar] [CrossRef]
- Pellizzari, G.; Martinez, O.; Crescioli, S.; Page, R.; Di Meo, A.; Mele, S.; Chiaruttini, G.; Hoinka, J.; Batruch, I.; Prassas, I.; et al. Immunotherapy using IgE or CAR T cells for cancers expressing the tumor antigen SLC3A2. J. Immunother. Cancer 2021, 9, e002140. [Google Scholar] [CrossRef]
- Cano-Crespo, S.; Chillaron, J.; Junza, A.; Fernandez-Miranda, G.; Garcia, J.; Polte, C.; de la Ballina, L.R.; Ignatova, Z.; Yanes, Ó.; Zorzano, A.; et al. CD98hc (SLC3A2) sustains amino acid and nucleotide availability for cell cycle progression. Sci. Rep. 2019, 9, 14065. [Google Scholar] [CrossRef]
- Yu, J.X.; Upadhaya, S.; Tatake, R.; Barkalow, F.; Hubbard-Lucey, V.M. Cancer cell therapies: The clinical trial landscape. Nat. Rev. Drug Discov. 2020, 19, 583–584. [Google Scholar] [CrossRef] [PubMed]
- Glover, M.; Avraamides, S.; Maher, J. How Can We Engineer CAR T Cells to Overcome Resistance? Biologics 2021, 15, 175–198. [Google Scholar] [CrossRef] [PubMed]
- Qi, C.; Gong, J.; Li, J.; Liu, D.; Qin, Y.; Ge, S.; Zhang, M.; Peng, Z.; Zhou, J.; Cao, Y.; et al. Claudin18.2-specific CAR T cells in gastrointestinal cancers: Phase 1 trial interim results. Nat. Med. 2022, 28, 1189–1198. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Target Class | Target |
|---|---|
| Glycosylphosphatidyl-inositol-anchored cell surface protein | Mesothelin |
| Carcinoembryonic antigen | |
| Receptor tyrosine kinase | HER2 |
| Epidermal growth factor receptor (EGFR) | |
| EGFR variant III | |
| Receptor tyrosine kinase-like orphan receptor family member (ROR) 1 | |
| c-Met | |
| Vascular endothelial growth factor receptor (VEGFR) 2 | |
| Erythropoietin-producing human hepatocellular carcinoma type A receptor 2 (EphA2) | |
| Mucins | Mucin (MUC) 1 |
| MUC 16 | |
| Tumour-associated glycoprotein (TAG) 72 | |
| Claudins | Claudin 6 |
| Claudin 18.2 | |
| Folate receptors | Folate receptor α |
| Interleukin (IL) receptor | IL-13 receptor α2 |
| Gangliosides | GD2 |
| B7 family members | B7-H3 |
| PD-L1 | |
| Glypicans | Glypican 3 |
| Prostate antigens | Prostate-specific membrane antigen |
| Prostate stem cell antigen | |
| Adhesion molecules | Epithelial cell adhesion molecule (EpCAM) |
| Neuronal L1 cell adhesion molecule (L1CAM) | |
| Miscellaneous | NKG2D ligands |
| CD70 | |
| Carboxy anhydrase IX | |
| Fibroblast activation protein | |
| CD133 | |
| Roundabout guidance receptor (ROBO) 1 |
| Target | Pre-Clinical In Vitro | Pre-Clinical In Vivo | Ongoing Clinical Trial |
|---|---|---|---|
| FGFR4 | X | X | |
| EphA3 | X | ||
| EphA10 | X | X | |
| EphB4 | X | X | |
| ROR2 | X | ||
| ALK | X | X | |
| AXL | X | X | X |
| PDGFR | X | X | |
| PTK7 | X | X | |
| ICAM-1 | X | X | |
| Cadherin 6 | X | ||
| Cadherin 17 | X | X | |
| Nectin-4 | X | X | |
| CD44v6 | X | X | X |
| αvβ6 integrin | X | X | |
| αvβ3 integrin | X | X | |
| CLEC14A | X | X | |
| Apelin receptor | X | X | |
| EDB | X | X | |
| EDA | X | X | |
| GD3 | X | X | |
| GM2 | X | X | X |
| B7-H4 | X | X | |
| B7-H6 | X | X | |
| KLK2 | X | X | |
| STEAP1 | X | X | |
| STEAP2 | Not reported | Not reported | |
| DLL3 | Not reported | Not reported | X |
| DLK1 | X | X | |
| HERV K | X | X | |
| LMP1 | X | X | |
| Glypican 1 | X | X | |
| Glypican 2 | X | X | |
| TROP2 | X | X | |
| HLA-G | X | X | X |
| 5T4 | X | X | X |
| Lewis Y | X | X | X |
| MG7 | Not reported | Not reported | X |
| GFR α4 | X | X | X |
| LGR5 | X | X | In preparation |
| nfP2X7 | Not reported | Not reported | |
| FSHR | X | X | X |
| GUCY2C | X | X | X |
| TSHR | X | X | |
| CLEC4 | Not reported | Not reported | |
| CD47 | X | X | |
| TM4SF1 | Not reported | Not reported | X |
| ADAM10 | X | X | |
| Chlorotoxin ligands | X | X | X |
| HSP70 | X | ||
| Cripto-1 | Not reported | Not reported | |
| GRP78 | X | X | |
| CD147 | X | X | X (non-Hodgkin’s lymphoma) |
| CD317 | X | X | |
| CSPG4 | X | X | |
| CD24 | X | X | |
| MISIIR | X | X | |
| SLC3A2 | X | X |
| Disease | Sponsor | Notes | Identifier |
|---|---|---|---|
| AXL + solid tumours | 2nd Affiliated Hospital Guangzhou Medical University | One of multiple targets | NCT03198052 |
| AXL + sarcomas | Shanghai PerHum Therapeutics | CXCR5 armoured CAR T-cells | NCT05128786 |
| Disease | Sponsor | Notes | Identifier |
|---|---|---|---|
| CD44v6 + Solid tumours | Shenzhen Geno-Immune Medical Institute | NCT04427449 | |
| CD44v6 + Breast cancer | Shenzhen Geno-Immune Medical Institute | One of multiple targets | NCT04430595 |
| Disease | Sponsor | Notes | Identifier |
|---|---|---|---|
| Solid tumours | Shanghai East Hospital | Allogeneic NK host cells | NCT05137275 |
| Solid tumours | Wuxi People’s Hospital | NK host cells | NCT05194709 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maher, J.; Davies, D.M. CAR-Based Immunotherapy of Solid Tumours—A Survey of the Emerging Targets. Cancers 2023, 15, 1171. https://doi.org/10.3390/cancers15041171
Maher J, Davies DM. CAR-Based Immunotherapy of Solid Tumours—A Survey of the Emerging Targets. Cancers. 2023; 15(4):1171. https://doi.org/10.3390/cancers15041171
Chicago/Turabian StyleMaher, John, and David M. Davies. 2023. "CAR-Based Immunotherapy of Solid Tumours—A Survey of the Emerging Targets" Cancers 15, no. 4: 1171. https://doi.org/10.3390/cancers15041171
APA StyleMaher, J., & Davies, D. M. (2023). CAR-Based Immunotherapy of Solid Tumours—A Survey of the Emerging Targets. Cancers, 15(4), 1171. https://doi.org/10.3390/cancers15041171