
Emerging Role of IGF-1 in Prostate Cancer: A Promising Biomarker and Therapeutic Target


Abstract
:Simple Summary
Abstract
1. Introduction
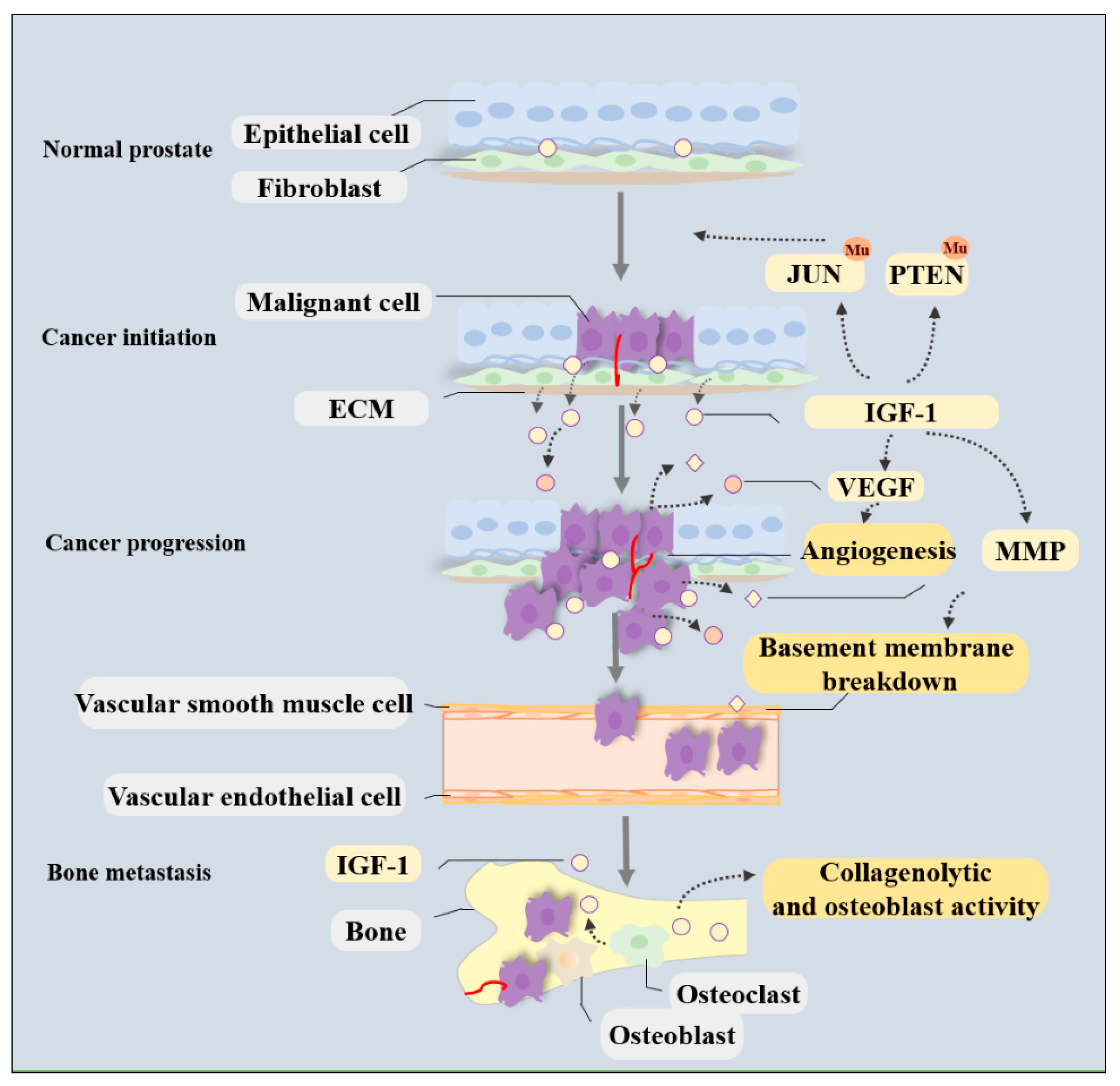
2. Circulating IGF-1 Levels and Local IGF-1R Expression Are Related to PCa Development
2.1. Tumor Initiation: Risk of Tumorigenesis
2.2. Tumor Progression: Cell Migration and Metastatic Spread
2.3. Biological Tumor Recurrence

{kind=link}
{kind=link}
| Study Population | Region | PCa Risk/Stage/Survival Related to IGF-1 or IGF-1R Expression | Main Conclusion | References |
|---|---|---|---|---|
| 152 PCa cases and 152 control subjects (median age = 60) | United States |
|
| [20] (Science,1998) |
| 630 PCa cases and 630 control subjects (median age = 65) | Europea |
|
| [19] (Cancer Epidemiol Biomarkers Prev, 2007) |
| 1709 PCa cases and 1778 control subjects | United States |
|
| [22] (Int J Cancer, 2015) |
| 793 PCa cases underwent radical prostatectomy and 272 men with negative prostate biopsy (mean age = 65) | Korea |
|
| [34] (Cancer Med, 2018) |
| 156 PCa cases (median age = 67) and 271 control subjects (median age = 69) | Austria |
|
| [32] (European Urology, 2005) |
| 72 PCa cases and 50 control subjects (median age = 67) | Unknown |
|
| [23] (Prostate, 2014) |
| 753 PCa cases in various stages (median age = 66) | United Kingdom |
|
| [28] (Carcinogenesis, 2018) |
| 360 patients underwent surgery for PCa or BPH (median age = 68) | Japan |
|
| [29] (Translational Research, 2021) |
| 130 patients with PCa (median age = 63) | United Kingdom |
|
| [13] (Prostate, 2017) |
| 136 patients with PCa (median age = 69) | Japan |
|
| [42] (British Journal of Cancer, 2017) |
| 215 patients of PCa with bone metastasis (median age = 70) or 111 patients of PCa with bone metastasis (median age = 70.6) | United Kingdom |
|
| [3,43] (BMC Cancer, 2013) (J Clin Oncol, 2006) |
2.4. Cancer-Specific Survival
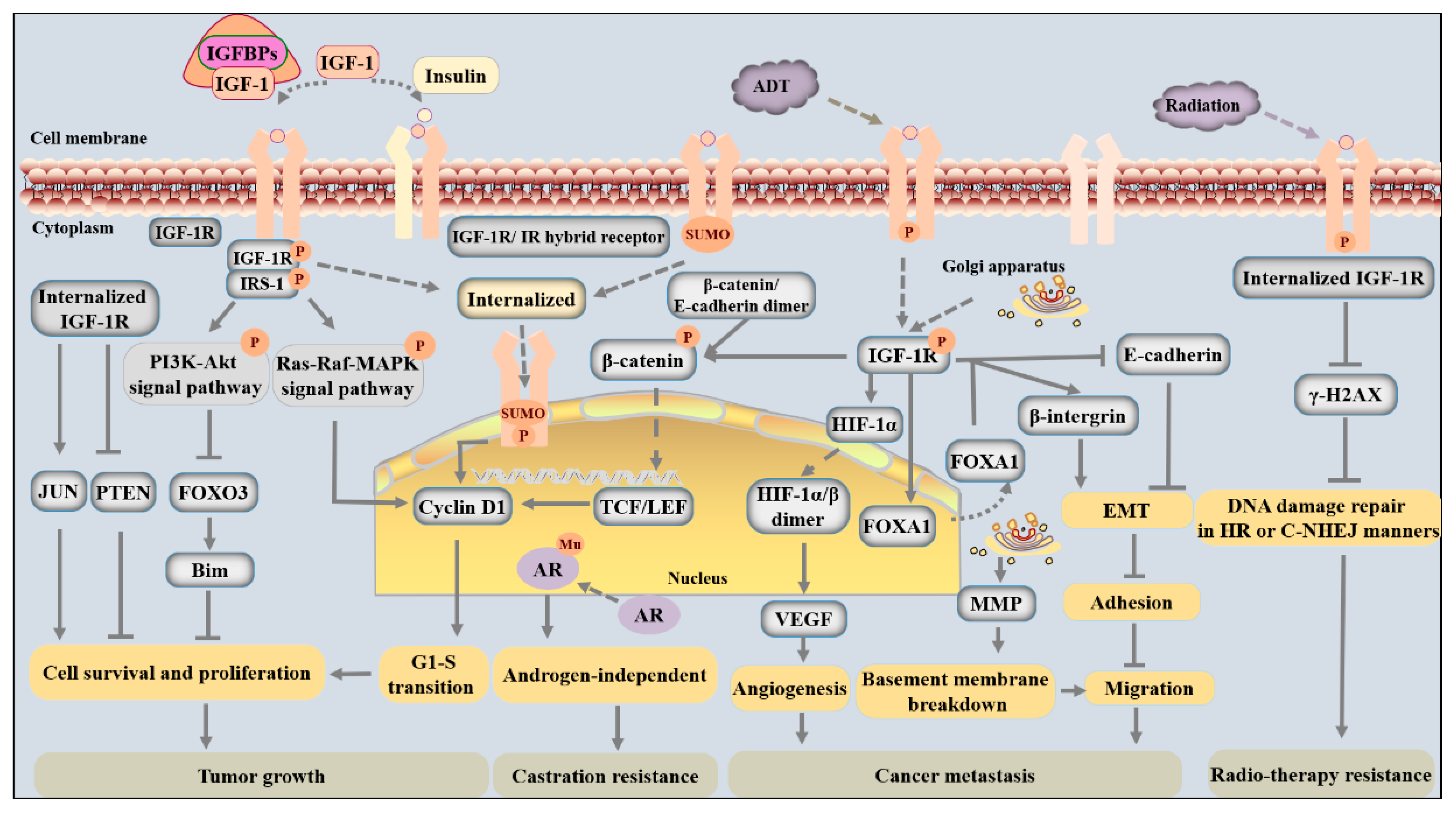
3. The Mechanism by Which IGF-1 Signaling Regulates PCa Progression
3.1. Cancer Cell Survival and Renewal
3.2. Cancer Cell Migration and Metastasis
3.2.1. Cell Adhesion and Motility
3.2.2. Angiogenesis
3.2.3. Bone Metastasis
3.3. Radiosensitivity/Radioresistance
3.4. Castration Sensitivity/Resistance
4. IGF-1 Signaling: A Target for PCa Treatment
4.1. Anti-IGF-1R Monoclonal Antibodies
4.1.1. Cixutumumab (IMC-A12)
4.1.2. Figitumumab
4.1.3. A12
4.1.4. Ganitumab (AMG 479)
4.1.5. Other IGF-1R-Targeting Monoclonal Antibodies
| Agents | Property | Phase/Region | Population | Treatment Schemes | Key Activity Data | Safety and Adverse Events | References |
|---|---|---|---|---|---|---|---|
| Cixutumumab | Recombinant human mAb, IgG1, targets IGF-1R | Phase II (2015/2020/USA) | 210 patients with mHSPC (PSA ≥ 5 ng/mL) | Cixutumumab (10 mg/kg, intravenously, 2 times/28 d, last 28 w) + Bicalutamide (oral, daily) + LHRH agonist |
|
| [90,99] (J Clin Oncol, 2015); (Prostate Cancer and Prostatic Diseases, 2020) |
| Phase I (2019/USA) | 16 patients with mCRPC (PSA ≥ 2 ng/mL) | Cixutumumab (6 mg/kg, 12 w) + Temsirolimus |
|
| [98] (Clinical Genitourinary Cancer, 2020) | ||
| Phase II (2015/USA) | 132 patients with docetaxel-treated mCRPC (PSA ≥ 2 ng/mL) | Cixutumumab (6 mg/kg, intravenously,3 times/21 d) + Mitoxantrone + Prednisone |
|
| [100] (Eur J Cancer, 2015) | ||
| Figitumumab | Full human mAb, IgG2, blocks IGF-1 binds to IGF-1R | Phase II (2012/Canada) | 16 patients with localized PCa (PSA ≥ 10 ng/mL) | Figitumumab (20 mg/kg, intravenously every 3 weeks, 3 cycles) |
|
| [102] (Clin Cancer Res, 2012) |
| Phase II (2014/UK) | 204 patients with mCRPC | Figitumumab (10 or 20 mg/kg, intravenously, 12 m) + Docetaxel vs. Docetaxel alone |
|
| [101] (Clin Cancer Res, 2014) |
4.2. IGF-Neutralizing Antibodies
4.2.1. Xentuzumab (BI 836845)
4.2.2. Dusigitumab (MEDI-573)
4.3. IGF-1R Inhibitory Small-Molecule Agents
4.3.1. Linsitinib (OSI-906)
4.3.2. BMS-754807
4.3.3. Picropodophyllin (AXL1717/PPP)
4.4. Brief Summary
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ADT | Androgen deprivation therapy |
| ALS | Acid-labile subunit |
| ASOs | Antisense oligonucleotides |
| BAD | Bcl-2-associated agonist of cell death |
| BCR | Biochemical recurrence |
| BPH | Benign prostatic hyperplasia |
| C-NHEJ | Classical nonhomologous end joining |
| CRPC | Castration-Resistant Prostate Cancer |
| DSBs | Double-strand breaks |
| ECM | Extracellular matrix |
| EMT | Epithelial-mesenchymal transition |
| FOXA1 | Forkhead box A1 |
| FOXOs | Forkhead box transcription factors |
| GS | Gleason score |
| GSK3α/β | Glycogen synthase kinase-3α/β |
| HIF-1α | Hypoxia-inducible factor-1α |
| HR | Homologous recombination |
| IGF-1 | Insulin-like growth factor-1 |
| IGFBPs | Insulin-like growth factor binding proteins |
| IGF-1R | Insulin-like growth factor-1 receptor |
| INSR | Insulin receptor |
| mAb | Monoclonal antibody |
| mHSPC | Metastatic castration-sensitive prostate cancer |
| MDR | Multidrug resistance |
| MMP | Matrix metalloproteinase |
| OR | Odds ratio |
| OS | Overall Survival |
| PCa | Prostate cancer |
| PFS | Progression-free survival |
| PSA | Prostate-specific antigen |
| PTEN | Gene of phosphate and tensin homolog deleted on chromosome ten |
| RR | Relative risk |
| siRNA | Small-interfering RNA |
| Src | Sarcoma gene |
| TCF/LEF | T-cell factor/lymphoid enhancer factors |
| TRAMP | Transgenic adenocarcinoma of mouse prostate |
| TUBB2B | β-tubulin isoform 2B |
| VEGF | Vascular endothelial growth factor |
References
- Peitzsch, C.; Gorodetska, I.; Klusa, D.; Shi, Q.; Alves, T.C.; Pantel, K.; Dubrovska, A. Metabolic regulation of prostate cancer heterogeneity and plasticity. Semin. Cancer Biol. 2022, 82, 94–119. [Google Scholar] [CrossRef]
- Fitzmaurice, C.; Abate, D.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdel-Rahman, O.; Abdelalim, A.; Abdoli, A.; Abdollahpour, I.; Abdulle, A.S.M.; et al. Global, Regional, and National Cancer Incidence, Mortality, Years of Life Lost, Years Lived with Disability, and Disability-Adjusted Life-Years for 29 Cancer Groups, 1990 to 2017: A Systematic Analysis for the Global Burden of Disease Study. JAMA Oncol. 2019, 5, 1749–1768. [Google Scholar] [CrossRef] [Green Version]
- Tsuchiya, N.; Wang, L.; Suzuki, H.; Segawa, T.; Fukuda, H.; Narita, S.; Shimbo, M.; Kamoto, T.; Mitsumori, K.; Ichikawa, T.; et al. Impact of IGF-I and CYP19 gene polymorphisms on the survival of patients with metastatic prostate cancer. J. Clin. Oncol. 2006, 24, 1982–1989. [Google Scholar] [CrossRef]
- Rebello, R.J.; Oing, C.; Knudsen, K.E.; Loeb, S.; Johnson, D.C.; Reiter, R.E.; Gillessen, S.; Van der Kwast, T.; Bristow, R.G. Prostate cancer. Nat. Rev. Dis. Prim. 2021, 7, 9. [Google Scholar] [CrossRef]
- Sandhu, S.; Moore, C.M.; Chiong, E.; Beltran, H.; Bristow, R.G.; Williams, S.G. Prostate cancer. Lancet 2021, 398, 1075–1090. [Google Scholar] [CrossRef]
- Wu, Y.; Brodt, P.; Sun, H.; Mejia, W.; Novosyadlyy, R.; Nunez, N.; Chen, X.; Mendoza, A.; Hong, S.H.; Khanna, C.; et al. Insulin-like growth factor-I regulates the liver microenvironment in obese mice and promotes liver metastasis. Cancer Res. 2010, 70, 57–67. [Google Scholar] [CrossRef] [Green Version]
- De Meyts, P.; Whittaker, J. Structural biology of insulin and IGF1 receptors: Implications for drug design. Nat. Rev. Drug Discov. 2002, 1, 769–783. [Google Scholar] [CrossRef]
- Li, L.; Yu, H.; Schumacher, F.; Casey, G.; Witte, J.S. Relation of serum insulin-like growth factor-I (IGF-I) and IGF binding protein-3 to risk of prostate cancer (United States). Cancer Causes Control 2003, 14, 721–726. [Google Scholar] [CrossRef]
- Pollak, M.N.; Schernhammer, E.S.; Hankinson, S.E. Insulin-like growth factors and neoplasia. Nat. Rev. Cancer 2004, 4, 505–518. [Google Scholar] [CrossRef]
- Bailes, J.; Soloviev, M. Insulin-Like Growth Factor-1 (IGF-1) and Its Monitoring in Medical Diagnostic and in Sports. Biomolecules 2021, 11, 217. [Google Scholar] [CrossRef]
- Forbes, B.E.; Blyth, A.J.; Wit, J.M. Disorders of IGFs and IGF-1R signaling pathways. Mol. Cell. Endocrinol. 2020, 518, 111035. [Google Scholar] [CrossRef]
- Rieunier, G.; Wu, X.; Harris, L.E.; Mills, J.V.; Nandakumar, A.; Colling, L.; Seraia, E.; Hatch, S.B.; Ebner, D.V.; Folkes, L.K.; et al. Targeting IGF Perturbs Global Replication through Ribonucleotide Reductase Dysfunction. Cancer Res. 2021, 81, 2128–2141. [Google Scholar] [CrossRef] [PubMed]
- Breen, K.J.; O’Neill, A.; Murphy, L.; Fan, Y.; Boyce, S.; Fitzgerald, N.; Dorris, E.; Brady, L.; Finn, S.P.; Hayes, B.D.; et al. Investigating the role of the IGF axis as a predictor of biochemical recurrence in prostate cancer patients post-surgery. Prostate 2017, 77, 1288–1300. [Google Scholar] [CrossRef] [PubMed]
- Aleksic, T.; Chitnis, M.M.; Perestenko, O.V.; Gao, S.; Thomas, P.H.; Turner, G.D.; Protheroe, A.S.; Howarth, M.; Macaulay, V.M. Type 1 insulin-like growth factor receptor translocates to the nucleus of human tumor cells. Cancer Res. 2010, 70, 6412–6419. [Google Scholar] [CrossRef] [Green Version]
- Goh, L.K.; Sorkin, A. Endocytosis of receptor tyrosine kinases. Cold Spring Harb. Perspect. Biol. 2013, 5, a017459. [Google Scholar] [CrossRef] [Green Version]
- Tentori, L.; Graziani, G. Doping with growth hormone/IGF-1, anabolic steroids or erythropoietin: Is there a cancer risk? Pharmacol. Res. 2007, 55, 359–369. [Google Scholar] [CrossRef]
- Lin, S.L.; Lin, C.Y.; Lee, W.; Teng, C.F.; Shyu, W.C.; Jeng, L.B. Mini Review: Molecular Interpretation of the IGF/IGF-1R Axis in Cancer Treatment and Stem Cells-Based Therapy in Regenerative Medicine. Int. J. Mol. Sci. 2022, 23, 11781. [Google Scholar] [CrossRef]
- Travis, R.C.; Appleby, P.N.; Martin, R.M.; Holly, J.M.P.; Albanes, D.; Black, A.; Bueno-de-Mesquita, H.B.A.; Chan, J.M.; Chen, C.; Chirlaque, M.D.; et al. A Meta-analysis of Individual Participant Data Reveals an Association between Circulating Levels of IGF-I and Prostate Cancer Risk. Cancer Res. 2016, 76, 2288–2300. [Google Scholar] [CrossRef] [Green Version]
- Allen, N.E.; Key, T.J.; Appleby, P.N.; Travis, R.C.; Roddam, A.W.; Rinaldi, S.; Egevad, L.; Rohrmann, S.; Linseisen, J.; Pischon, T.; et al. Serum insulin-like growth factor (IGF)-I and IGF-binding protein-3 concentrations and prostate cancer risk: Results from the European Prospective Investigation into Cancer and Nutrition. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1121–1127. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.M.; Stampfer, M.J.; Giovannucci, E.; Gann, P.H.; Ma, J.; Wilkinson, P.; Hennekens, C.H.; Pollak, M. Plasma insulin-like growth factor-I and prostate cancer risk: A prospective study. Science 1998, 279, 563–566. [Google Scholar] [CrossRef]
- Schmitz, M.; Grignard, G.; Margue, C.; Dippel, W.; Capesius, C.; Mossong, J.; Nathan, M.; Giacchi, S.; Scheiden, R.; Kieffer, N. Complete loss of PTEN expression as a possible early prognostic marker for prostate cancer metastasis. Int. J. Cancer 2007, 120, 1284–1292. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Nimptsch, K.; Shui, I.M.; Platz, E.A.; Wu, K.; Pollak, M.N.; Kenfield, S.A.; Stampfer, M.J.; Giovannucci, E.L. Prediagnostic plasma IGFBP-1, IGF-1 and risk of prostate cancer. Int. J. Cancer 2015, 136, 2418–2426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, J.; Gray, K.P.; Evan, C.; Nakabayashi, M.; Fichorova, R.; Rider, J.; Mucci, L.; Kantoff, P.W.; Sweeney, C.J. Elevated insulin-like growth factor binding protein-1 (IGFBP-1) in men with metastatic prostate cancer starting androgen deprivation therapy (ADT) is associated with shorter time to castration resistance and overall survival. Prostate 2014, 74, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Papatsoris, A.G.; Karamouzis, M.V.; Papavassiliou, A.G. Novel insights into the implication of the IGF-1 network in prostate cancer. Trends Mol. Med. 2005, 11, 52–55. [Google Scholar] [CrossRef]
- Hellawell, G.O.; Turner, G.D.; Davies, D.R.; Poulsom, R.; Brewster, S.F.; Macaulay, V.M. Expression of the type 1 insulin-like growth factor receptor is up-regulated in primary prostate cancer and commonly persists in metastatic disease. Cancer Res. 2002, 62, 2942–2950. [Google Scholar]
- Nickerson, T.; Chang, F.; Lorimer, D.; Smeekens, S.P.; Sawyers, C.L.; Pollak, M. In vivo progression of LAPC-9 and LNCaP prostate cancer models to androgen independence is associated with increased expression of insulin-like growth factor I (IGF-I) and IGF-I receptor (IGF-IR). Cancer Res. 2001, 61, 6276–6280. [Google Scholar]
- Wu, J.; Yu, E. Insulin-like growth factor receptor-1 (IGF-IR) as a target for prostate cancer therapy. Cancer Metastasis Rev. 2014, 33, 607–617. [Google Scholar] [CrossRef] [Green Version]
- Ahearn, T.U.; Peisch, S.; Pettersson, A.; Ebot, E.M.; Zhou, C.K.; Graff, R.E.; Sinnott, J.A.; Fazli, L.; Judson, G.L.; Bismar, T.A.; et al. Expression of IGF/insulin receptor in prostate cancer tissue and progression to lethal disease. Carcinogenesis 2018, 39, 1431–1437. [Google Scholar] [CrossRef]
- Broggi, G.; Lo Giudice, A.; Di Mauro, M.; Pricoco, E.; Piombino, E.; Ferro, M.; Caltabiano, R.; Morgia, G.; Russo, G.I. Insulin signaling, androgen receptor and PSMA immunohistochemical analysis by semi-automated tissue microarray in prostate cancer with diabetes (DIAMOND study). Transl. Res. J. Lab. Clin. Med. 2021, 238, 25–35. [Google Scholar] [CrossRef]
- Rieger, L.; O’Shea, S.; Godsmark, G.; Stanicka, J.; Kelly, G.; O’Connor, R. IGF-1 receptor activity in the Golgi of migratory cancer cells depends on adhesion-dependent phosphorylation of Tyr(1250) and Tyr(1251). Sci. Signal. 2020, 13, eaba3176. [Google Scholar] [CrossRef]
- Ismail, H.A.; Pollak, M.; Behlouli, H.; Tanguay, S.; Bégin, L.R.; Aprikian, A.G. Serum insulin-like growth factor (IGF)-1 and IGF-binding protein-3 do not correlate with Gleason score or quantity of prostate cancer in biopsy samples. BJU Int. 2003, 92, 699–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marszalek, M.; Wachter, J.; Ponholzer, A.; Leitha, T.; Rauchenwald, M.; Madersbacher, S. Insulin-like growth factor 1, chromogranin A and prostate specific antigen serum levels in prostate cancer patients and controls. Eur. Urol. 2005, 48, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Philippou, A.; Armakolas, A.; Koutsilieris, M. Evidence for the Possible Biological Significance of the igf-1 Gene Alternative Splicing in Prostate Cancer. Front. Endocrinol. 2013, 4, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Kim, J.W.; Kim, J.K.; Lee, S.M.; Song, C.; Jeong, I.G.; Hong, J.H.; Kim, C.S.; Ahn, H. Association between serum levels of insulin-like growth factor-1, bioavailable testosterone, and pathologic Gleason score. Cancer Med. 2018, 7, 4170–4180. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, P.J.; Mohan, S.; Cohen, P.; Foster, B.A.; Greenberg, N.M. The insulin-like growth factor axis and prostate cancer: Lessons from the transgenic adenocarcinoma of mouse prostate (TRAMP) model. Cancer Res. 1999, 59, 2203–2209. [Google Scholar]
- Aleksic, T.; Gray, N.; Wu, X.; Rieunier, G.; Osher, E.; Mills, J.; Verrill, C.; Bryant, R.J.; Han, C.; Hutchinson, K.; et al. Nuclear IGF1R Interacts with Regulatory Regions of Chromatin to Promote RNA Polymerase II Recruitment and Gene Expression Associated with Advanced Tumor Stage. Cancer Res. 2018, 78, 3497–3509. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.A.; Campbell, C.I.; Wood, G.A.; Petrik, J.J.; Moorehead, R.A. Reversibility and recurrence of IGF-IR-induced mammary tumors. Oncogene 2009, 28, 2152–2162. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Mao, W.; Dong, M.; Yang, D.; Li, W.; Chen, Y. Serum Insulin-Like Growth Factor-1 (IGF-1): A Novel Prognostic Factor for Early Recurrence of Hepatocellular Carcinoma (HCC). Clin. Lab. 2017, 63, 261–270. [Google Scholar] [CrossRef]
- Shitara, K.; Ito, S.; Misawa, K.; Ito, Y.; Ito, H.; Hosono, S.; Watanabe, M.; Tajima, K.; Tanaka, H.; Muro, K.; et al. Genetic polymorphism of IGF-I predicts recurrence in patients with gastric cancer who have undergone curative gastrectomy. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2012, 23, 659–664. [Google Scholar] [CrossRef]
- Yu, H.; Nicar, M.R.; Shi, R.; Berkel, H.J.; Nam, R.; Trachtenberg, J.; Diamandis, E.P. Levels of insulin-like growth factor I (IGF-I) and IGF binding proteins 2 and 3 in serial postoperative serum samples and risk of prostate cancer recurrence. Urology 2001, 57, 471–475. [Google Scholar] [CrossRef]
- Chitnis, M.M.; Lodhia, K.A.; Aleksic, T.; Gao, S.; Protheroe, A.S.; Macaulay, V.M. IGF-1R inhibition enhances radiosensitivity and delays double-strand break repair by both non-homologous end-joining and homologous recombination. Oncogene 2014, 33, 5262–5273. [Google Scholar] [CrossRef] [Green Version]
- Aleksic, T.; Verrill, C.; Bryant, R.J.; Han, C.; Worrall, A.R.; Brureau, L.; Larré, S.; Higgins, G.S.; Fazal, F.; Sabbagh, A.; et al. IGF-1R associates with adverse outcomes after radical radiotherapy for prostate cancer. Br. J. Cancer 2017, 117, 1600–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchiya, N.; Narita, S.; Inoue, T.; Saito, M.; Numakura, K.; Huang, M.; Hatakeyama, S.; Satoh, S.; Saito, S.; Ohyama, C.; et al. Insulin-like growth factor-1 genotypes and haplotypes influence the survival of prostate cancer patients with bone metastasis at initial diagnosis. BMC Cancer 2013, 13, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Liu, H.; Waraky, A.; Haglund, F.; Agarwal, P.; Jernberg-Wiklund, H.; Warsito, D.; Larsson, O. SUMO-modified insulin-like growth factor 1 receptor (IGF-1R) increases cell cycle progression and cell proliferation. J. Cell. Physiol. 2017, 232, 2722–2730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Packham, S.; Warsito, D.; Lin, Y.; Sadi, S.; Karlsson, R.; Sehat, B.; Larsson, O. Nuclear translocation of IGF-1R via p150(Glued) and an importin-β/RanBP2-dependent pathway in cancer cells. Oncogene 2015, 34, 2227–2238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, T.R.; Zhau, H.E.; Odero-Marah, V.A.; Osunkoya, A.O.; Kimbro, K.S.; Tighiouart, M.; Liu, T.; Simons, J.W.; O’Regan, R.M. Insulin-like growth factor-I-dependent up-regulation of ZEB1 drives epithelial-to-mesenchymal transition in human prostate cancer cells. Cancer Res. 2008, 68, 2479–2488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, T.E.; Epa, V.C.; Garrett, T.P.; Ward, C.W. Structure and function of the type 1 insulin-like growth factor receptor. Cell. Mol. Life Sci. 2000, 57, 1050–1093. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Choi, E.; Yu, H.; Bai, X.C. Structural basis of the activation of type 1 insulin-like growth factor receptor. Nat. Commun. 2019, 10, 4567. [Google Scholar] [CrossRef] [Green Version]
- Sayeed, A.; Fedele, C.; Trerotola, M.; Ganguly, K.K.; Languino, L.R. IGF-IR promotes prostate cancer growth by stabilizing α5β1 integrin protein levels. PLoS ONE 2013, 8, e76513. [Google Scholar] [CrossRef] [Green Version]
- Goel, H.L.; Breen, M.; Zhang, J.; Das, I.; Aznavoorian-Cheshire, S.; Greenberg, N.M.; Elgavish, A.; Languino, L.R. beta1A integrin expression is required for type 1 insulin-like growth factor receptor mitogenic and transforming activities and localization to focal contacts. Cancer Res. 2005, 65, 6692–6700. [Google Scholar] [CrossRef] [Green Version]
- Siech, C.; Rutz, J.; Maxeiner, S.; Grein, T.; Sonnenburg, M.; Tsaur, I.; Chun, F.K.; Blaheta, R.A. Insulin-like Growth Factor-1 Influences Prostate Cancer Cell Growth and Invasion through an Integrin α3, α5, αV, and β1 Dependent Mechanism. Cancers 2022, 14, 363. [Google Scholar] [CrossRef]
- Feng, Z.; Levine, A.J. The regulation of energy metabolism and the IGF-1/mTOR pathways by the p53 protein. Trends Cell Biol. 2010, 20, 427–434. [Google Scholar] [CrossRef] [Green Version]
- Rinner, O.; Mueller, L.N.; Hubálek, M.; Müller, M.; Gstaiger, M.; Aebersold, R. An integrated mass spectrometric and computational framework for the analysis of protein interaction networks. Nat. Biotechnol. 2007, 25, 345–352. [Google Scholar] [CrossRef]
- Wang, S.; Wang, N.; Yu, B.; Cao, M.; Wang, Y.; Guo, Y.; Zhang, Y.; Zhang, P.; Yu, X.; Wang, S.; et al. Circulating IGF-1 promotes prostate adenocarcinoma via FOXO3A/BIM signaling in a double-transgenic mouse model. Oncogene 2019, 38, 6338–6353. [Google Scholar] [CrossRef]
- Habrowska-Górczyńska, D.E.; Kozieł, M.J.; Kowalska, K.; Piastowska-Ciesielska, A.W. FOXO3a and Its Regulators in Prostate Cancer. Int. J. Mol. Sci. 2021, 22, 12530. [Google Scholar] [CrossRef]
- Lee, J.; Ju, J.; Park, S.; Hong, S.J.; Yoon, S. Inhibition of IGF-1 signaling by genistein: Modulation of E-cadherin expression and downregulation of β-catenin signaling in hormone refractory PC-3 prostate cancer cells. Nutr. Cancer 2012, 64, 153–162. [Google Scholar] [CrossRef]
- Feng, Z. p53 regulation of the IGF-1/AKT/mTOR pathways and the endosomal compartment. Cold Spring Harb. Perspect. Biol. 2010, 2, a001057. [Google Scholar] [CrossRef] [Green Version]
- Nordstrand, A.; Bergström, S.H.; Thysell, E.; Bovinder-Ylitalo, E.; Lerner, U.H.; Widmark, A.; Bergh, A.; Wikström, P. Inhibition of the insulin-like growth factor-1 receptor potentiates acute effects of castration in a rat model for prostate cancer growth in bone. Clin. Exp. Metastasis 2017, 34, 261–271. [Google Scholar] [CrossRef] [Green Version]
- Åberg, M.; Edén, D.; Siegbahn, A. Activation of β1 integrins and caveolin-1 by TF/FVIIa promotes IGF-1R signaling and cell survival. Apoptosis Int. J. Program. Cell Death 2020, 25, 519–534. [Google Scholar] [CrossRef]
- Juan-Rivera, M.C.; Martínez-Ferrer, M. Integrin Inhibitors in Prostate Cancer. Cancers 2018, 10, 44. [Google Scholar] [CrossRef] [Green Version]
- Melincovici, C.S.; Boşca, A.B.; Şuşman, S.; Mărginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular endothelial growth factor (VEGF)—Key factor in normal and pathological angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar]
- Rieger, L.; O’Connor, R. Controlled Signaling-Insulin-Like Growth Factor Receptor Endocytosis and Presence at Intracellular Compartments. Front. Endocrinol. 2020, 11, 620013. [Google Scholar] [CrossRef] [PubMed]
- Kiely, P.A.; Leahy, M.; O’Gorman, D.; O’Connor, R. RACK1-mediated integration of adhesion and insulin-like growth factor I (IGF-I) signaling and cell migration are defective in cells expressing an IGF-I receptor mutated at tyrosines 1250 and 1251. J. Biol. Chem. 2005, 280, 7624–7633. [Google Scholar] [CrossRef] [Green Version]
- Brodt, P.; Fallavollita, L.; Khatib, A.M.; Samani, A.A.; Zhang, D. Cooperative regulation of the invasive and metastatic phenotypes by different domains of the type I insulin-like growth factor receptor beta subunit. J. Biol. Chem. 2001, 276, 33608–33615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sroka, I.C.; McDaniel, K.; Nagle, R.B.; Bowden, G.T. Differential localization of MT1-MMP in human prostate cancer tissue: Role of IGF-1R in MT1-MMP expression. Prostate 2008, 68, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, C.; Goswami, S.; Basu, S.; Chakroborty, D. Angiogenesis Inhibition in Prostate Cancer: An Update. Cancers 2020, 12, 2382. [Google Scholar] [CrossRef] [PubMed]
- Odero-Marah, V.; Hawsawi, O.; Henderson, V.; Sweeney, J. Epithelial-Mesenchymal Transition (EMT) and Prostate Cancer. Adv. Exp. Med. Biol. 2018, 1095, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Mansor, R.; Holly, J.; Barker, R.; Biernacka, K.; Zielinska, H.; Koupparis, A.; Rowe, E.; Oxley, J.; Sewell, A.; Martin, R.M.; et al. IGF-1 and hyperglycaemia-induced FOXA1 and IGFBP-2 affect epithelial to mesenchymal transition in prostate epithelial cells. Oncotarget 2020, 11, 2543–2559. [Google Scholar] [CrossRef]
- Potter, A.S.; Casa, A.J.; Lee, A.V. Forkhead box A1 (FOXA1) is a key mediator of insulin-like growth factor I (IGF-I) activity. J. Cell. Biochem. 2012, 113, 110–121. [Google Scholar] [CrossRef] [Green Version]
- Calvani, M.; Rapisarda, A.; Uranchimeg, B.; Shoemaker, R.H.; Melillo, G. Hypoxic induction of an HIF-1alpha-dependent bFGF autocrine loop drives angiogenesis in human endothelial cells. Blood 2006, 107, 2705–2712. [Google Scholar] [CrossRef]
- Tang, X.; Zhang, Q.; Shi, S.; Yen, Y.; Li, X.; Zhang, Y.; Zhou, K.; Le, A.D. Bisphosphonates suppress insulin-like growth factor 1-induced angiogenesis via the HIF-1alpha/VEGF signaling pathways in human breast cancer cells. Int. J. Cancer 2010, 126, 90–103. [Google Scholar] [CrossRef] [Green Version]
- Acker, T.; Plate, K.H. Role of hypoxia in tumor angiogenesis-molecular and cellular angiogenic crosstalk. Cell Tissue Res. 2003, 314, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Berish, R.B.; Ali, A.N.; Telmer, P.G.; Ronald, J.A.; Leong, H.S. Translational models of prostate cancer bone metastasis. Nat. Rev. Urol. 2018, 15, 403–421. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X. Interactions between cancer cells and bone microenvironment promote bone metastasis in prostate cancer. Cancer Commun. 2019, 39, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, E.T.; Brown, J. Prostate cancer bone metastases promote both osteolytic and osteoblastic activity. J. Cell. Biochem. 2004, 91, 718–729. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.L.; Pienta, K.J.; Kim, W.J.; Cooper, C.R. The effect of bone-associated growth factors and cytokines on the growth of prostate cancer cells derived from soft tissue versus bone metastases in vitro. Int. J. Oncol. 2003, 22, 921–926. [Google Scholar] [CrossRef]
- Crane, J.L.; Cao, X. Function of matrix IGF-1 in coupling bone resorption and formation. J. Mol. Med. 2014, 92, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Graves, D.T. Impact of the host response and osteoblast lineage cells on periodontal disease. Front. Immunol. 2022, 13, 998244. [Google Scholar] [CrossRef]
- Hiraga, T.; Myoui, A.; Hashimoto, N.; Sasaki, A.; Hata, K.; Morita, Y.; Yoshikawa, H.; Rosen, C.J.; Mundy, G.R.; Yoneda, T. Bone-derived IGF mediates crosstalk between bone and breast cancer cells in bony metastases. Cancer Res. 2012, 72, 4238–4249. [Google Scholar] [CrossRef] [Green Version]
- Buijs, J.T.; van der Pluijm, G. Osteotropic cancers: From primary tumor to bone. Cancer Lett. 2009, 273, 177–193. [Google Scholar] [CrossRef]
- Litwin, M.S.; Tan, H.J. The Diagnosis and Treatment of Prostate Cancer: A Review. JAMA 2017, 317, 2532–2542. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.; Chibly, A.M.; Burd, R.; Limesand, K.H. Insulin-Like Growth Factor-1-Mediated DNA Repair in Irradiated Salivary Glands Is Sirtuin-1 Dependent. J. Dent. Res. 2017, 96, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Cosaceanu, D.; Budiu, R.A.; Carapancea, M.; Castro, J.; Lewensohn, R.; Dricu, A. Ionizing radiation activates IGF-1R triggering a cytoprotective signaling by interfering with Ku-DNA binding and by modulating Ku86 expression via a p38 kinase-dependent mechanism. Oncogene 2007, 26, 2423–2434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, D.; Watanabe, H.; Shibuya, H.; Miura, M. Redundancy of radioresistant signaling pathways originating from insulin-like growth factor I receptor. J. Biol. Chem. 2003, 278, 6702–6709. [Google Scholar] [CrossRef] [Green Version]
- Venkatachalam, S.; Mettler, E.; Fottner, C.; Miederer, M.; Kaina, B.; Weber, M.M. The impact of the IGF-1 system of cancer cells on radiation response—An In Vitro study. Clin. Transl. Radiat. Oncol. 2017, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ge, R.; Wang, Z.; Montironi, R.; Jiang, Z.; Cheng, M.; Santoni, M.; Huang, K.; Massari, F.; Lu, X.; Cimadamore, A.; et al. Epigenetic modulations and lineage plasticity in advanced prostate cancer. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2020, 31, 470–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quigley, D.A.; Dang, H.X.; Zhao, S.G.; Lloyd, P.; Aggarwal, R.; Alumkal, J.J.; Foye, A.; Kothari, V.; Perry, M.D.; Bailey, A.M.; et al. Genomic Hallmarks and Structural Variation in Metastatic Prostate Cancer. Cell 2018, 174, 758–769.e759. [Google Scholar] [CrossRef] [Green Version]
- Baures, M.; Puig Lombardi, E.; Di Martino, D.; Zeitouni, W.; Pacreau, E.; Dos Santos, L.; Dariane, C.; Boutillon, F.; Guidotti, J.E.; Goffin, V. Transcriptomic Signature and Growth Factor Regulation of Castration-Tolerant Prostate Luminal Progenitor Cells. Cancers 2022, 14, 3775. [Google Scholar] [CrossRef]
- Wu, J.D.; Haugk, K.; Woodke, L.; Nelson, P.; Coleman, I.; Plymate, S.R. Interaction of IGF signaling and the androgen receptor in prostate cancer progression. J. Cell. Biochem. 2006, 99, 392–401. [Google Scholar] [CrossRef]
- Yu, E.Y.; Li, H.; Higano, C.S.; Agarwal, N.; Pal, S.K.; Alva, A.; Heath, E.I.; Lam, E.T.; Gupta, S.; Lilly, M.B.; et al. SWOG S0925: A Randomized Phase II Study of Androgen Deprivation Combined with Cixutumumab Versus Androgen Deprivation Alone in Patients with New Metastatic Hormone-Sensitive Prostate Cancer. J. Clin. Oncol. 2015, 33, 1601–1608. [Google Scholar] [CrossRef]
- Fan, W.; Yanase, T.; Morinaga, H.; Okabe, T.; Nomura, M.; Daitoku, H.; Fukamizu, A.; Kato, S.; Takayanagi, R.; Nawata, H. Insulin-like growth factor 1/insulin signaling activates androgen signaling through direct interactions of Foxo1 with androgen receptor. J. Biol. Chem. 2007, 282, 7329–7338. [Google Scholar] [CrossRef] [Green Version]
- Pandini, G.; Mineo, R.; Frasca, F.; Roberts, C.T., Jr.; Marcelli, M.; Vigneri, R.; Belfiore, A. Androgens up-regulate the insulin-like growth factor-I receptor in prostate cancer cells. Cancer Res. 2005, 65, 1849–1857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liotti, A.; La Civita, E.; Cennamo, M.; Crocetto, F.; Ferro, M.; Guadagno, E.; Insabato, L.; Imbimbo, C.; Palmieri, A.; Mirone, V.; et al. Periprostatic adipose tissue promotes prostate cancer resistance to docetaxel by paracrine IGF-1 upregulation of TUBB2B beta-tubulin isoform. Prostate 2021, 81, 407–417. [Google Scholar] [CrossRef]
- Niu, X.B.; Fu, G.B.; Wang, L.; Ge, X.; Liu, W.T.; Wen, Y.Y.; Sun, H.R.; Liu, L.Z.; Wang, Z.J.; Jiang, B.H. Insulin-like growth factor-I induces chemoresistence to docetaxel by inhibiting miR-143 in human prostate cancer. Oncotarget 2017, 8, 107157–107166. [Google Scholar] [CrossRef] [Green Version]
- Heidegger, I.; Massoner, P.; Sampson, N.; Klocker, H. The insulin-like growth factor (IGF) axis as an anticancer target in prostate cancer. Cancer Lett. 2015, 367, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Samani, A.A.; Yakar, S.; LeRoith, D.; Brodt, P. The role of the IGF system in cancer growth and metastasis: Overview and recent insights. Endocr. Rev. 2007, 28, 20–47. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.D.; Odman, A.; Higgins, L.M.; Haugk, K.; Vessella, R.; Ludwig, D.L.; Plymate, S.R. In vivo effects of the human type I insulin-like growth factor receptor antibody A12 on androgen-dependent and androgen-independent xenograft human prostate tumors. Clin. Cancer Res. 2005, 11, 3065–3074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McHugh, D.J.; Chudow, J.; DeNunzio, M.; Slovin, S.F.; Danila, D.C.; Morris, M.J.; Scher, H.I.; Rathkopf, D.E. A Phase I Trial of IGF-1R Inhibitor Cixutumumab and mTOR Inhibitor Temsirolimus in Metastatic Castration-resistant Prostate Cancer. Clin. Genitourin. Cancer 2020, 18, 171–178.e2. [Google Scholar] [CrossRef]
- Wong, R.L.; Duong, M.T.; Tangen, C.M.; Agarwal, N.; Cheng, H.H.; Vogelzang, N.J.; Hussain, M.; Thompson, I.M., Jr.; Quinn, D.I.; Yu, E.Y. Survival outcomes and risk group validation from SWOG S0925: A randomized phase II study of cixutumumab in new metastatic hormone-sensitive prostate cancer. Prostate Cancer Prostatic Dis. 2020, 23, 486–493. [Google Scholar] [CrossRef]
- Hussain, M.; Rathkopf, D.; Liu, G.; Armstrong, A.; Kelly, W.K.; Ferrari, A.; Hainsworth, J.; Joshi, A.; Hozak, R.R.; Yang, L.; et al. A randomised non-comparative phase II trial of cixutumumab (IMC-A12) or ramucirumab (IMC-1121B) plus mitoxantrone and prednisone in men with metastatic docetaxel-pretreated castration-resistant prostate cancer. Eur. J. Cancer 2015, 51, 1714–1724. [Google Scholar] [CrossRef] [Green Version]
- de Bono, J.S.; Piulats, J.M.; Pandha, H.S.; Petrylak, D.P.; Saad, F.; Aparicio, L.M.; Sandhu, S.K.; Fong, P.; Gillessen, S.; Hudes, G.R.; et al. Phase II randomized study of figitumumab plus docetaxel and docetaxel alone with crossover for metastatic castration-resistant prostate cancer. Clin. Cancer Res. 2014, 20, 1925–1934. [Google Scholar] [CrossRef] [Green Version]
- Chi, K.N.; Gleave, M.E.; Fazli, L.; Goldenberg, S.L.; So, A.; Kollmannsberger, C.; Murray, N.; Tinker, A.; Pollak, M. A phase II pharmacodynamic study of preoperative figitumumab in patients with localized prostate cancer. Clin. Cancer Res. 2012, 18, 3407–3413. [Google Scholar] [CrossRef] [Green Version]
- Burtrum, D.; Zhu, Z.; Lu, D.; Anderson, D.M.; Prewett, M.; Pereira, D.S.; Bassi, R.; Abdullah, R.; Hooper, A.T.; Koo, H.; et al. A fully human monoclonal antibody to the insulin-like growth factor I receptor blocks ligand-dependent signaling and inhibits human tumor growth in vivo. Cancer Res. 2003, 63, 8912–8921. [Google Scholar]
- Wu, J.D.; Haugk, K.; Coleman, I.; Woodke, L.; Vessella, R.; Nelson, P.; Montgomery, R.B.; Ludwig, D.L.; Plymate, S.R. Combined in vivo effect of A12, a type 1 insulin-like growth factor receptor antibody, and docetaxel against prostate cancer tumors. Clin. Cancer Res. 2006, 12, 6153–6160. [Google Scholar] [CrossRef] [Green Version]
- Fahrenholtz, C.D.; Beltran, P.J.; Burnstein, K.L. Targeting IGF-IR with ganitumab inhibits tumorigenesis and increases durability of response to androgen-deprivation therapy in VCaP prostate cancer xenografts. Mol. Cancer Ther. 2013, 12, 394–404. [Google Scholar] [CrossRef] [Green Version]
- Galet, C.; Gray, A.; Said, J.W.; Castor, B.; Wan, J.; Beltran, P.J.; Calzone, F.J.; Elashoff, D.; Cohen, P.; Aronson, W.J. Effects of calorie restriction and IGF-1 receptor blockade on the progression of 22Rv1 prostate cancer xenografts. Int. J. Mol. Sci. 2013, 14, 13782–13795. [Google Scholar] [CrossRef] [Green Version]
- Konijeti, R.; Koyama, S.; Gray, A.; Barnard, R.J.; Said, J.W.; Castor, B.; Elashoff, D.; Wan, J.; Beltran, P.J.; Calzone, F.J.; et al. Effect of a low-fat diet combined with IGF-1 receptor blockade on 22Rv1 prostate cancer xenografts. Mol. Cancer Ther. 2012, 11, 1539–1546. [Google Scholar] [CrossRef] [Green Version]
- Beltran, P.J.; Mitchell, P.; Chung, Y.A.; Cajulis, E.; Lu, J.; Belmontes, B.; Ho, J.; Tsai, M.M.; Zhu, M.; Vonderfecht, S.; et al. AMG 479, a fully human anti-insulin-like growth factor receptor type I monoclonal antibody, inhibits the growth and survival of pancreatic carcinoma cells. Mol. Cancer Ther. 2009, 8, 1095–1105. [Google Scholar] [CrossRef] [Green Version]
- Tolcher, A.W.; Sarantopoulos, J.; Patnaik, A.; Papadopoulos, K.; Lin, C.C.; Rodon, J.; Murphy, B.; Roth, B.; McCaffery, I.; Gorski, K.S.; et al. Phase I, pharmacokinetic, and pharmacodynamic study of AMG 479, a fully human monoclonal antibody to insulin-like growth factor receptor 1. J. Clin. Oncol. 2009, 27, 5800–5807. [Google Scholar] [CrossRef]
- Rosen, L.S.; Puzanov, I.; Friberg, G.; Chan, E.; Hwang, Y.C.; Deng, H.; Gilbert, J.; Mahalingam, D.; McCaffery, I.; Michael, S.A.; et al. Safety and pharmacokinetics of ganitumab (AMG 479) combined with sorafenib, panitumumab, erlotinib, or gemcitabine in patients with advanced solid tumors. Clin. Cancer Res. 2012, 18, 3414–3427. [Google Scholar] [CrossRef] [Green Version]
- von Mehren, M.; Britten, C.D.; Pieslor, P.; Saville, W.; Vassos, A.; Harris, S.; Galluppi, G.R.; Darif, M.; Wainberg, Z.A.; Cohen, R.B.; et al. A phase 1, open-label, dose-escalation study of BIIB022 (anti-IGF-1R monoclonal antibody) in subjects with relapsed or refractory solid tumors. Investig. New Drugs 2014, 32, 518–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atzori, F.; Tabernero, J.; Cervantes, A.; Prudkin, L.; Andreu, J.; Rodríguez-Braun, E.; Domingo, A.; Guijarro, J.; Gamez, C.; Rodon, J.; et al. A phase I pharmacokinetic and pharmacodynamic study of dalotuzumab (MK-0646), an anti-insulin-like growth factor-1 receptor monoclonal antibody, in patients with advanced solid tumors. Clin. Cancer Res. 2011, 17, 6304–6312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Lipari, P.; Wang, X.; Hailey, J.; Liang, L.; Ramos, R.; Liu, M.; Pachter, J.A.; Bishop, W.R.; Wang, Y. A fully human insulin-like growth factor-I receptor antibody SCH 717454 (Robatumumab) has antitumor activity as a single agent and in combination with cytotoxics in pediatric tumor xenografts. Mol. Cancer Ther. 2010, 9, 410–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frappaz, D.; Federico, S.M.; Pearson, A.D.; Gore, L.; Macy, M.E.; DuBois, S.G.; Aerts, I.; Iannone, R.; Geschwindt, R.; Van Schanke, A.; et al. Phase 1 study of dalotuzumab monotherapy and ridaforolimus-dalotuzumab combination therapy in paediatric patients with advanced solid tumours. Eur. J. Cancer 2016, 62, 9–17. [Google Scholar] [CrossRef]
- Anderson, P.M.; Bielack, S.S.; Gorlick, R.G.; Skubitz, K.; Daw, N.C.; Herzog, C.E.; Monge, O.R.; Lassaletta, A.; Boldrini, E.; Pápai, Z.; et al. A phase II study of clinical activity of SCH 717454 (robatumumab) in patients with relapsed osteosarcoma and Ewing sarcoma. Pediatr. Blood Cancer 2016, 63, 1761–1770. [Google Scholar] [CrossRef] [Green Version]
- de Bono, J.; Lin, C.C.; Chen, L.T.; Corral, J.; Michalarea, V.; Rihawi, K.; Ong, M.; Lee, J.H.; Hsu, C.H.; Yang, J.C.; et al. Two first-in-human studies of xentuzumab, a humanised insulin-like growth factor (IGF)-neutralising antibody, in patients with advanced solid tumours. Br. J. Cancer 2020, 122, 1324–1332. [Google Scholar] [CrossRef] [Green Version]
- Parra-Guillen, Z.P.; Schmid, U.; Janda, A.; Freiwald, M.; Troconiz, I.F. Model-Informed Dose Selection for Xentuzumab, a Dual Insulin-Like Growth Factor-I/II-Neutralizing Antibody. Clin. Pharmacol. Ther. 2020, 107, 597–606. [Google Scholar] [CrossRef]
- Weyer-Czernilofsky, U.; Hofmann, M.H.; Friedbichler, K.; Baumgartinger, R.; Adam, P.J.; Solca, F.; Kraut, N.; Nguyen, H.M.; Corey, E.; Liu, G.; et al. Antitumor Activity of the IGF-1/IGF-2-Neutralizing Antibody Xentuzumab (BI 836845) in Combination with Enzalutamide in Prostate Cancer Models. Mol. Cancer Ther. 2020, 19, 1059–1069. [Google Scholar] [CrossRef] [Green Version]
- Doi, T.; Kuboki, Y.; Naito, Y.; Ishida, M.; Tanaka, T.; Takeuchi, Y. A phase 1 trial of xentuzumab, an IGF-neutralizing antibody, in Japanese patients with advanced solid tumors. Cancer Sci. 2022, 113, 1010–1017. [Google Scholar] [CrossRef]
- Schmid, P.; Sablin, M.P.; Bergh, J.; Im, S.A.; Lu, Y.S.; Martínez, N.; Neven, P.; Lee, K.S.; Morales, S.; Pérez-Fidalgo, J.A.; et al. A phase Ib/II study of xentuzumab, an IGF-neutralising antibody, combined with exemestane and everolimus in hormone receptor-positive, HER2-negative locally advanced/metastatic breast cancer. Breast Cancer Res. 2021, 23, 8. [Google Scholar] [CrossRef]
- Saad, F. Evidence for the efficacy of enzalutamide in postchemotherapy metastatic castrate-resistant prostate cancer. Ther. Adv. Urol. 2013, 5, 201–210. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Chesebrough, J.W.; Cartlidge, S.A.; Ricketts, S.A.; Incognito, L.; Veldman-Jones, M.; Blakey, D.C.; Tabrizi, M.; Jallal, B.; Trail, P.A.; et al. Dual IGF-I/II-neutralizing antibody MEDI-573 potently inhibits IGF signaling and tumor growth. Cancer Res. 2011, 71, 1029–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haluska, P.; Menefee, M.; Plimack, E.R.; Rosenberg, J.; Northfelt, D.; LaVallee, T.; Shi, L.; Yu, X.Q.; Burke, P.; Huang, J.; et al. Phase I dose-escalation study of MEDI-573, a bispecific, antiligand monoclonal antibody against IGFI and IGFII, in patients with advanced solid tumors. Clin. Cancer Res. 2014, 20, 4747–4757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bid, H.K.; London, C.A.; Gao, J.; Zhong, H.; Hollingsworth, R.E.; Fernandez, S.; Mo, X.; Houghton, P.J. Dual targeting of the type 1 insulin-like growth factor receptor and its ligands as an effective antiangiogenic strategy. Clin. Cancer Res. 2013, 19, 2984–2994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuentes-Baile, M.; Ventero, M.P.; Encinar, J.A.; García-Morales, P.; Poveda-Deltell, M.; Pérez-Valenciano, E.; Barberá, V.M.; Gallego-Plazas, J.; Rodríguez-Lescure, Á.; Martín-Nieto, J.; et al. Differential Effects of IGF-1R Small Molecule Tyrosine Kinase Inhibitors BMS-754807 and OSI-906 on Human Cancer Cell Lines. Cancers 2020, 12, 3717. [Google Scholar] [CrossRef]
- Fassnacht, M.; Berruti, A.; Baudin, E.; Demeure, M.J.; Gilbert, J.; Haak, H.; Kroiss, M.; Quinn, D.I.; Hesseltine, E.; Ronchi, C.L.; et al. Linsitinib (OSI-906) versus placebo for patients with locally advanced or metastatic adrenocortical carcinoma: A double-blind, randomised, phase 3 study. Lancet Oncol. 2015, 16, 426–435. [Google Scholar] [CrossRef] [Green Version]
- McKinley, E.T.; Bugaj, J.E.; Zhao, P.; Guleryuz, S.; Mantis, C.; Gokhale, P.C.; Wild, R.; Manning, H.C. 18FDG-PET predicts pharmacodynamic response to OSI-906, a dual IGF-1R/IR inhibitor, in preclinical mouse models of lung cancer. Clin. Cancer Res. 2011, 17, 3332–3340. [Google Scholar] [CrossRef] [Green Version]
- Barata, P.; Cooney, M.; Tyler, A.; Wright, J.; Dreicer, R.; Garcia, J.A. A phase 2 study of OSI-906 (linsitinib, an insulin-like growth factor receptor-1 inhibitor) in patients with asymptomatic or mildly symptomatic (non-opioid requiring) metastatic castrate resistant prostate cancer (CRPC). Investig. New Drugs 2018, 36, 451–457. [Google Scholar] [CrossRef]
- Ciuleanu, T.E.; Ahmed, S.; Kim, J.H.; Mezger, J.; Park, K.; Thomas, M.; Chen, J.; Poondru, S.; VanTornout, J.M.; Whitcomb, D.; et al. Randomised Phase 2 study of maintenance linsitinib (OSI-906) in combination with erlotinib compared with placebo plus erlotinib after platinum-based chemotherapy in patients with advanced non-small cell lung cancer. Br. J. Cancer 2017, 117, 757–766. [Google Scholar] [CrossRef] [Green Version]
- von Mehren, M.; George, S.; Heinrich, M.C.; Schuetze, S.M.; Yap, J.T.; Yu, J.Q.; Abbott, A.; Litwin, S.; Crowley, J.; Belinsky, M.; et al. Linsitinib (OSI-906) for the Treatment of Adult and Pediatric Wild-Type Gastrointestinal Stromal Tumors, a SARC Phase II Study. Clin. Cancer Res. 2020, 26, 1837–1845. [Google Scholar] [CrossRef]
- Wang, Y.J.; Zhang, Y.K.; Kathawala, R.J.; Chen, Z.S. Repositioning of Tyrosine Kinase Inhibitors as Antagonists of ATP-Binding Cassette Transporters in Anticancer Drug Resistance. Cancers 2014, 6, 1925–1952. [Google Scholar] [CrossRef]
- Zhang, H.; Kathawala, R.J.; Wang, Y.J.; Zhang, Y.K.; Patel, A.; Shukla, S.; Robey, R.W.; Talele, T.T.; Ashby, C.R., Jr.; Ambudkar, S.V.; et al. Linsitinib (OSI-906) antagonizes ATP-binding cassette subfamily G member 2 and subfamily C member 10-mediated drug resistance. Int. J. Biochem. Cell Biol. 2014, 51, 111–119. [Google Scholar] [CrossRef]
- Carboni, J.M.; Wittman, M.; Yang, Z.; Lee, F.; Greer, A.; Hurlburt, W.; Hillerman, S.; Cao, C.; Cantor, G.H.; Dell-John, J.; et al. BMS-754807, a small molecule inhibitor of insulin-like growth factor-1R/IR. Mol. Cancer Ther. 2009, 8, 3341–3349. [Google Scholar] [CrossRef] [Green Version]
- Dayyani, F.; Parikh, N.U.; Varkaris, A.S.; Song, J.H.; Moorthy, S.; Chatterji, T.; Maity, S.N.; Wolfe, A.R.; Carboni, J.M.; Gottardis, M.M.; et al. Combined Inhibition of IGF-1R/IR and Src family kinases enhances antitumor effects in prostate cancer by decreasing activated survival pathways. PLoS ONE 2012, 7, e51189. [Google Scholar] [CrossRef] [Green Version]
- Girnita, A.; All-Ericsson, C.; Economou, M.A.; Aström, K.; Axelson, M.; Seregard, S.; Larsson, O.; Girnita, L. The insulin-like growth factor-I receptor inhibitor picropodophyllin causes tumor regression and attenuates mechanisms involved in invasion of uveal melanoma cells. Clin. Cancer Res. 2006, 12, 1383–1391. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Chen, X.; Wang, G.; Lei, D.; Chen, X.; Lin, K.; Li, M.; Lin, H.; Li, D.; Zheng, Q. Picropodophyllin Inhibits the Proliferation of Human Prostate Cancer DU145 and LNCaP Cells via ROS Production and PI3K/AKT Pathway Inhibition. Biol. Pharm. Bull. 2022, 45, 1027–1035. [Google Scholar] [CrossRef]
- Pollak, M. The insulin and insulin-like growth factor receptor family in neoplasia: An update. Nat. Rev. Cancer 2012, 12, 159–169. [Google Scholar] [CrossRef]
- Goel, H.L.; Chang, C.; Pursell, B.; Leav, I.; Lyle, S.; Xi, H.S.; Hsieh, C.C.; Adisetiyo, H.; Roy-Burman, P.; Coleman, I.M.; et al. VEGF/neuropilin-2 regulation of Bmi-1 and consequent repression of IGF-IR define a novel mechanism of aggressive prostate cancer. Cancer Discov. 2012, 2, 906–921. [Google Scholar] [CrossRef] [Green Version]
- Muders, M.H.; Zhang, H.; Wang, E.; Tindall, D.J.; Datta, K. Vascular endothelial growth factor-C protects prostate cancer cells from oxidative stress by the activation of mammalian target of rapamycin complex-2 and AKT-1. Cancer Res. 2009, 69, 6042–6048. [Google Scholar] [CrossRef] [Green Version]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, G.; Zhu, M.; Zhang, M.; Pan, F. Emerging Role of IGF-1 in Prostate Cancer: A Promising Biomarker and Therapeutic Target. Cancers 2023, 15, 1287. https://doi.org/10.3390/cancers15041287
Liu G, Zhu M, Zhang M, Pan F. Emerging Role of IGF-1 in Prostate Cancer: A Promising Biomarker and Therapeutic Target. Cancers. 2023; 15(4):1287. https://doi.org/10.3390/cancers15041287
Chicago/Turabian StyleLiu, Guoqiang, Minggang Zhu, Mingrui Zhang, and Feng Pan. 2023. "Emerging Role of IGF-1 in Prostate Cancer: A Promising Biomarker and Therapeutic Target" Cancers 15, no. 4: 1287. https://doi.org/10.3390/cancers15041287
APA StyleLiu, G., Zhu, M., Zhang, M., & Pan, F. (2023). Emerging Role of IGF-1 in Prostate Cancer: A Promising Biomarker and Therapeutic Target. Cancers, 15(4), 1287. https://doi.org/10.3390/cancers15041287