

KIF2C Facilitates Tumor Growth and Metastasis in Pancreatic Ductal Adenocarcinoma
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Survival Curve and Correlation Analysis
2.2. Patients and Specimens
2.3. Immunohistochemical (IHC) Staining
2.4. Cell Culture
2.5. Plasmid Construction and Transfection
2.6. Cell Proliferation, Migration, and Invasion Assays
2.7. RNA Isolation and Quantitative RT-PCR
2.8. Western Blot
2.9. Antibodies
2.10. Animals Model
2.11. Embedding and Slicing
2.12. Cell Cycle Detection
2.13. H&E Staining
2.14. Statistical Analysis
2.15. Data Availability Statement
2.16. Ethics
3. Results
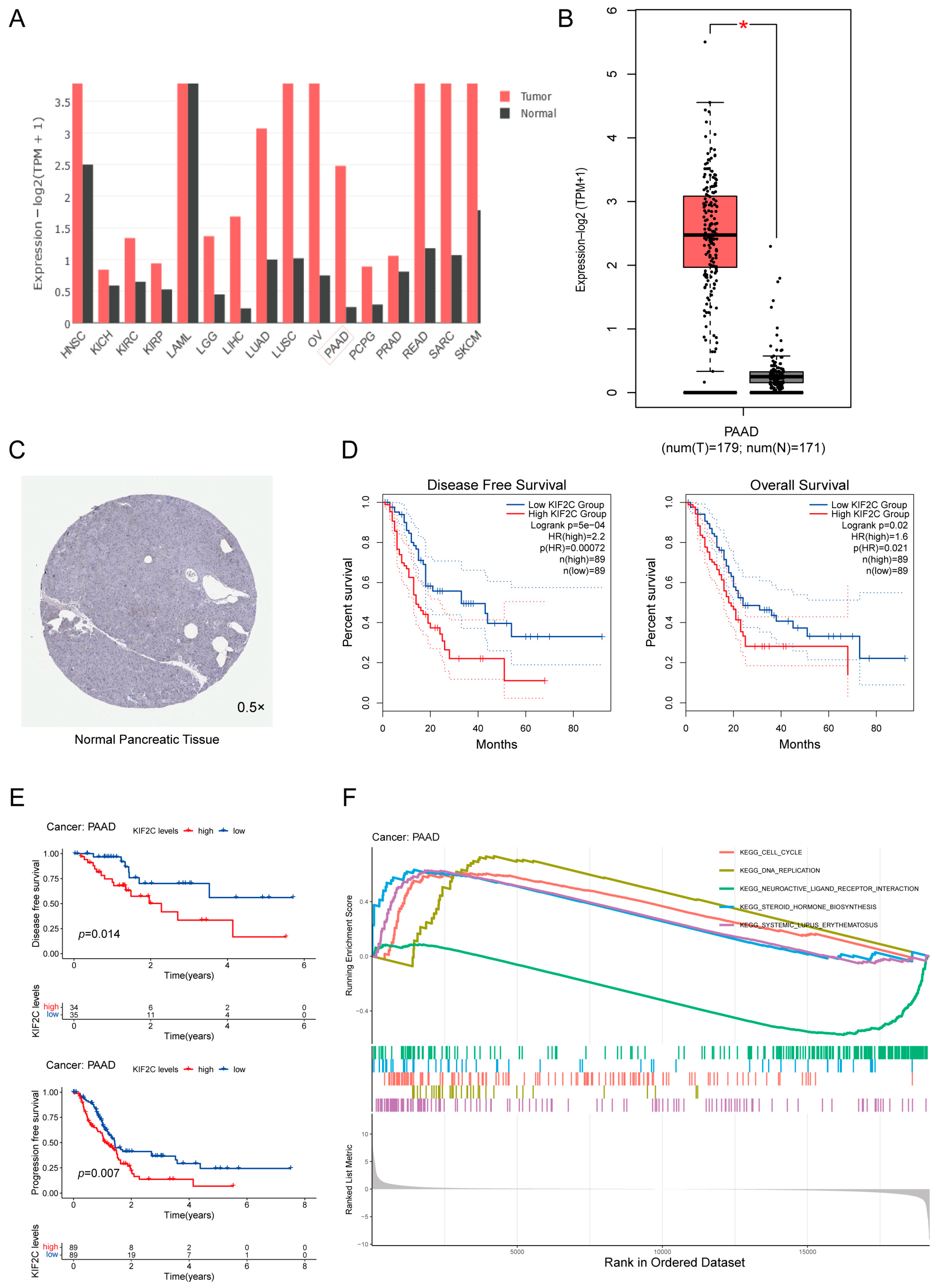
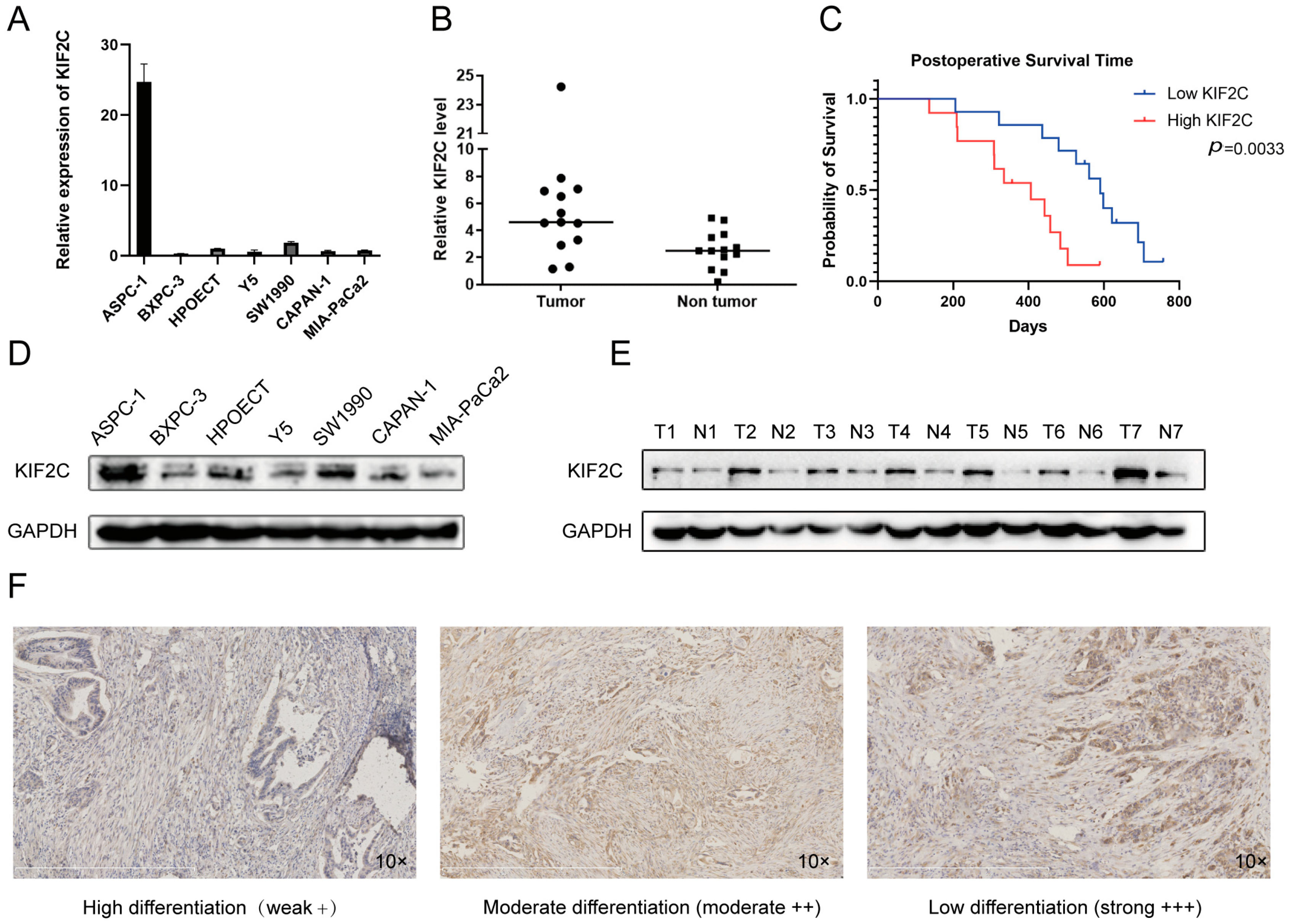
3.1. Upregulation of KIF2C in PDAC
3.2. Abnormal Expression of KIF2C in Clinical Specimens
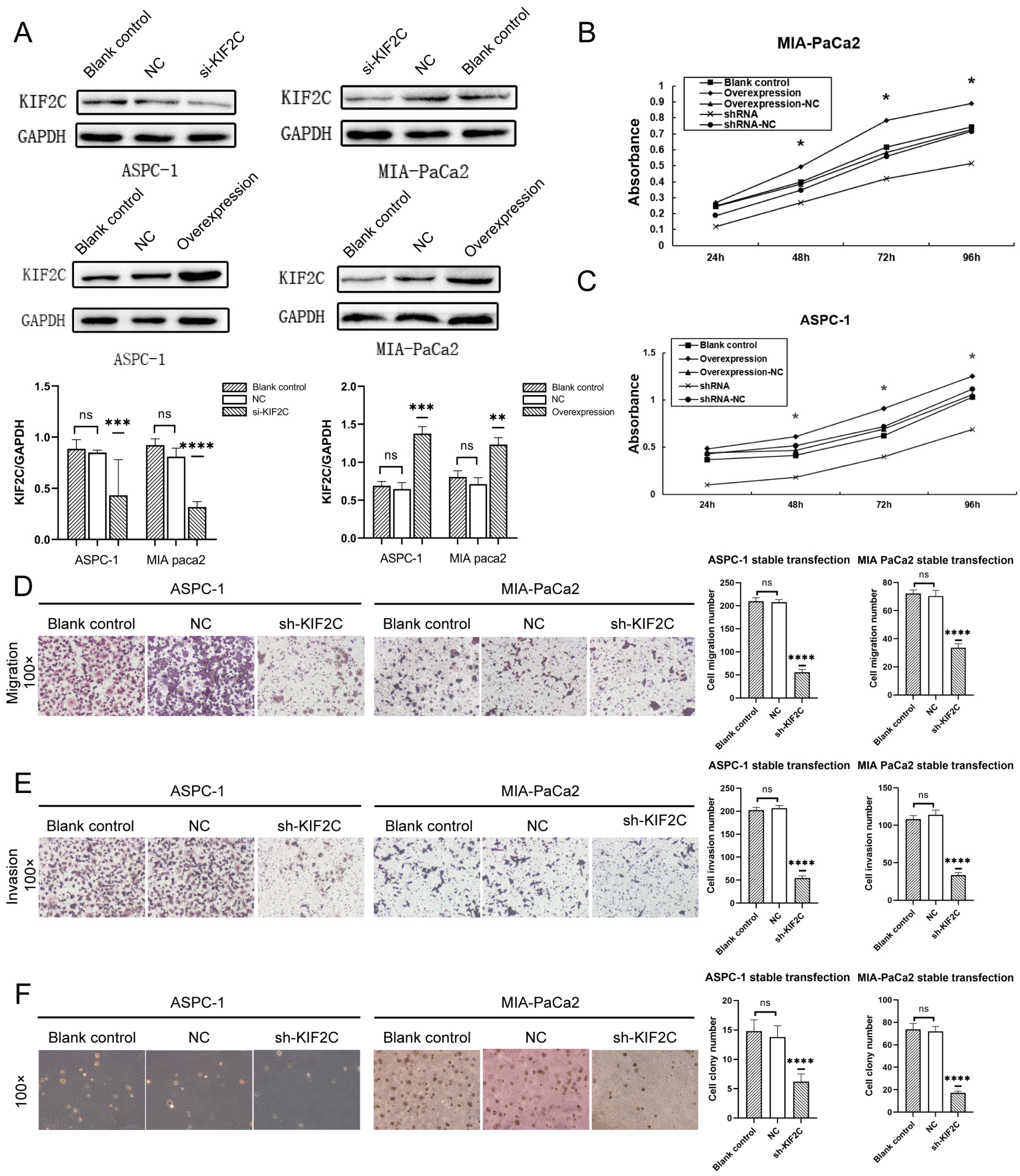
3.3. Knockdown of KIF2C Inhibits PDAC Cell Proliferation, Migration, and Invasion In Vitro
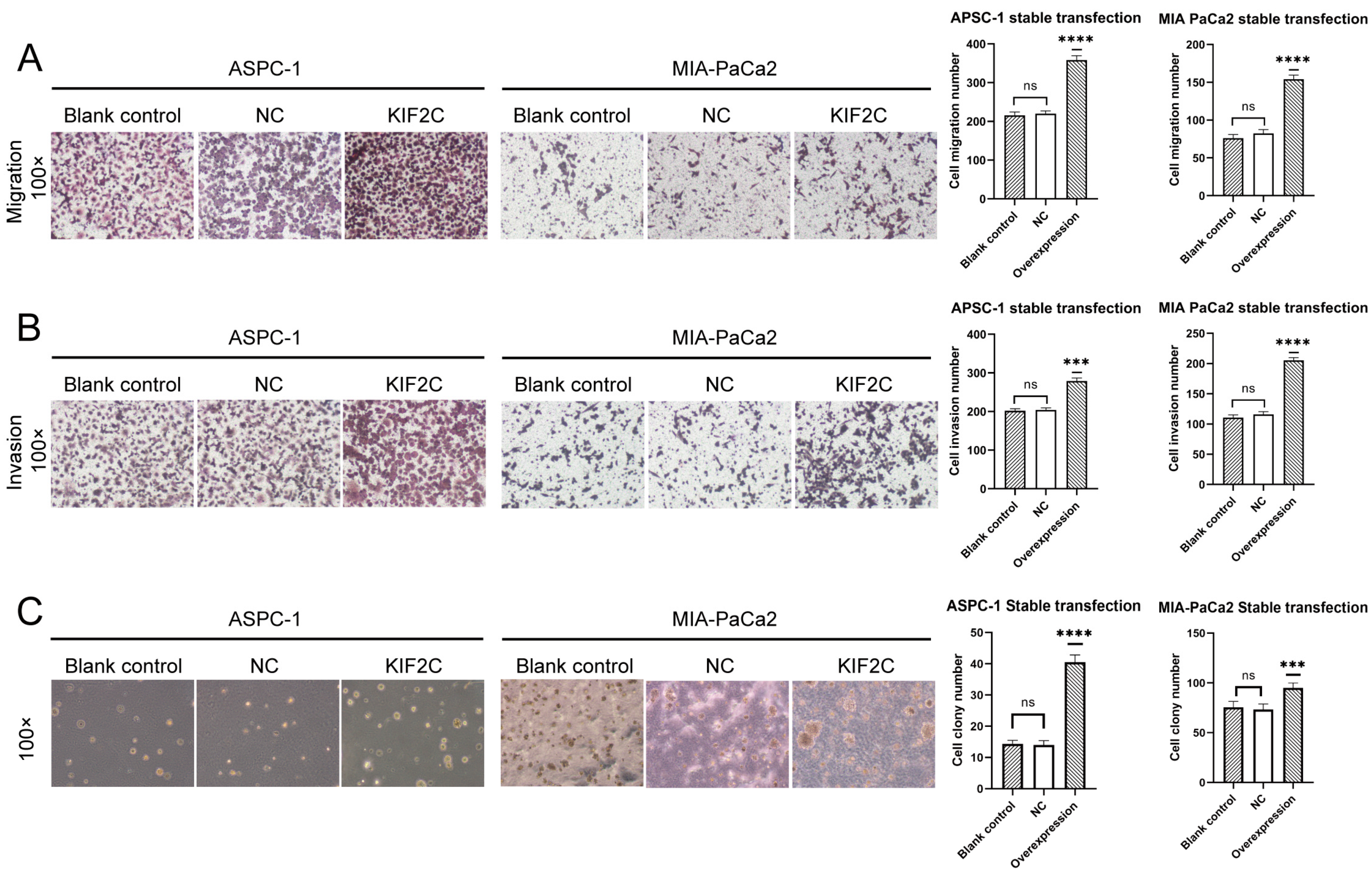
3.4. Overexpression of KIF2C Promotes PDAC Cell Proliferation, Migration, and Invasion In Vitro
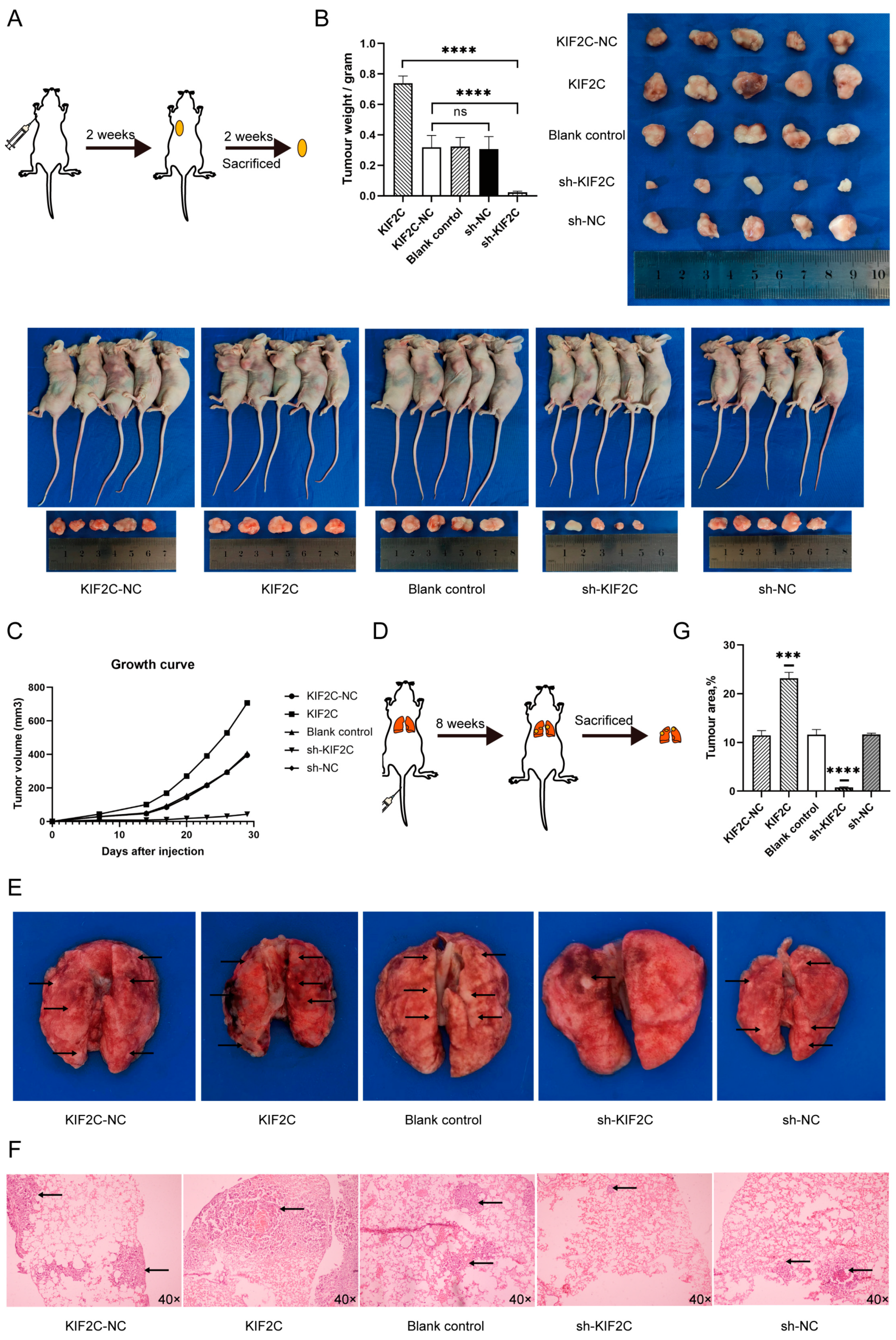
3.5. KIF2C Promotes PDAC Cell Proliferation, Migration, and Invasion In Vivo
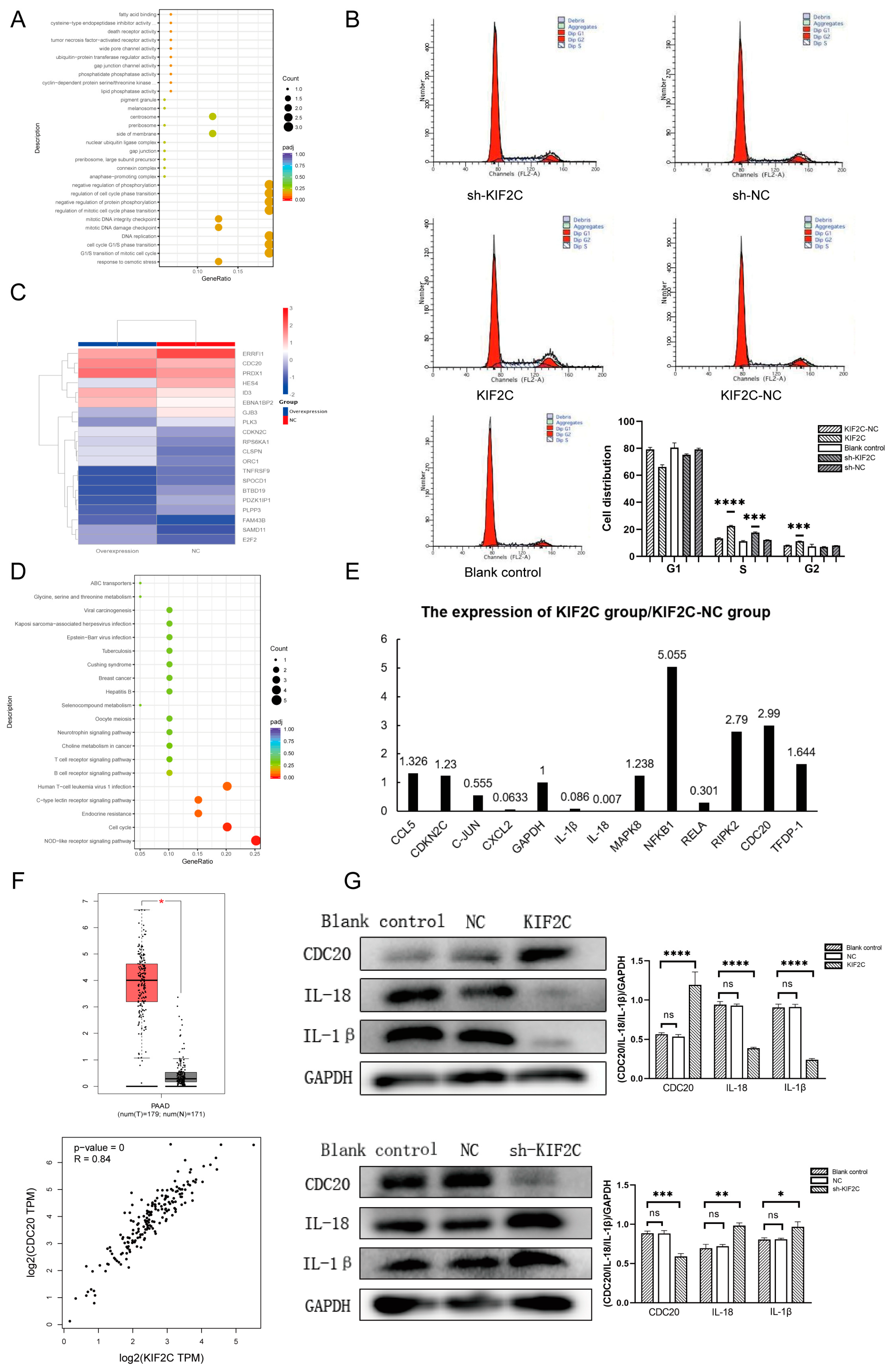
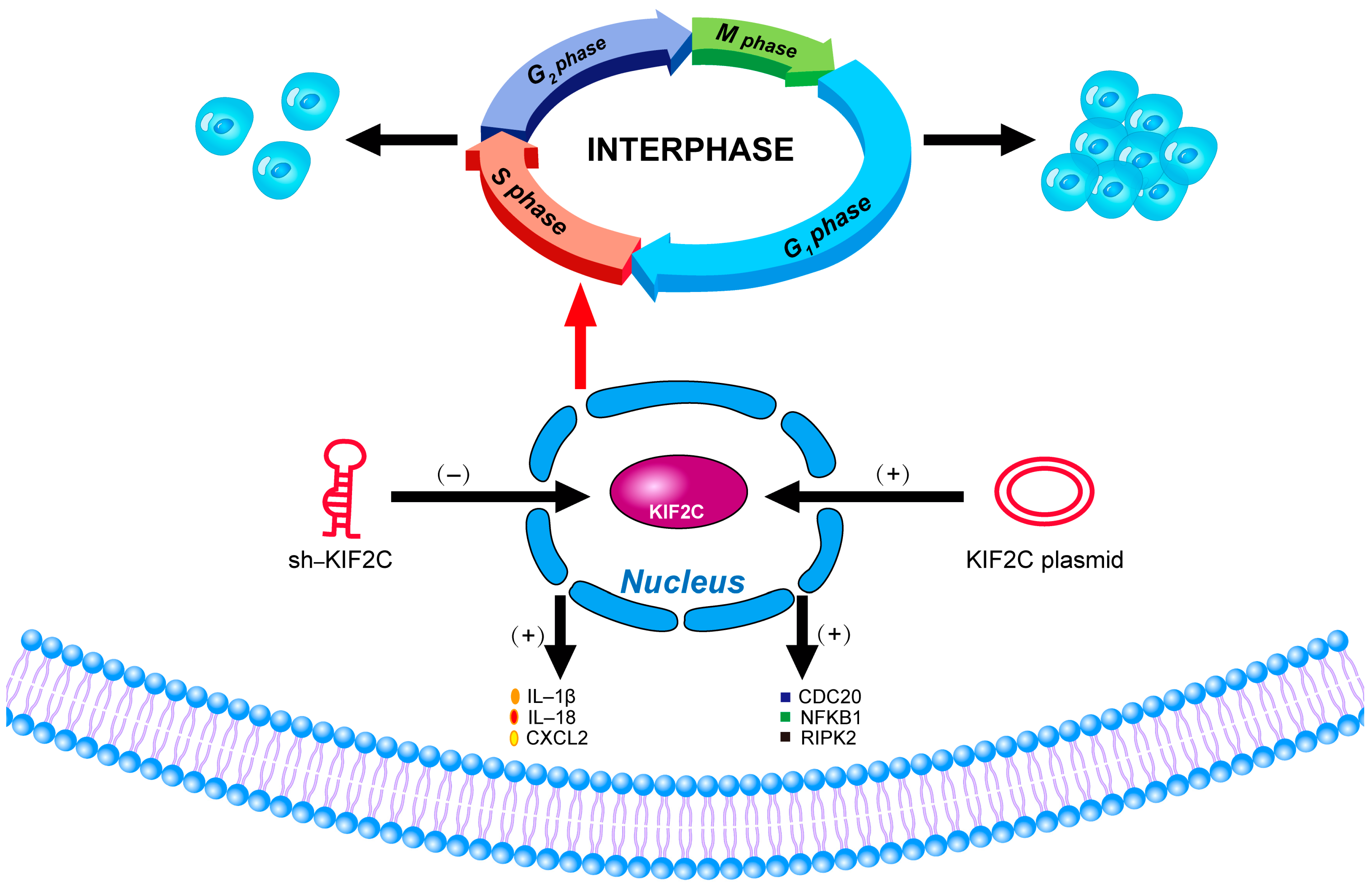
3.6. The Potential Mechanism of KIF2C in PDAC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Abbassi, R.; Algül, H. Palliative chemotherapy in pancreatic cancer-treatment sequences. Transl. Gastroenterol. Hepatol. 2019, 4, 56. [Google Scholar] [CrossRef] [PubMed]
- Kleeff, J.; Korc, M.; Apte, M.; La Vecchia, C.; Johnson, C.D.; Biankin, A.V.; Neale, R.E.; Tempero, M.; Tuveson, D.A.; Hruban, R.H.; et al. Pancreatic cancer. Nat. Rev. Dis. Prim. 2016, 2, 16022. [Google Scholar] [CrossRef]
- Deng, J.; Kang, Y.; Cheng, C.C.; Li, X.; Dai, B.; Katz, M.H.; Men, T.; Kim, M.P.; Koay, E.A.; Huang, H.; et al. DDR1-induced neutrophil extracellular traps drive pancreatic cancer metastasis. JCI Insight 2021, 6, e146133. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.S.; Li, H.; Li, T.J.; Li, S.; Xia, H.Y.; Long, J.; Wu, C.T.; Wang, W.Q.; Zhang, W.H.; Gao, H.L.; et al. Neutrophil Extracellular Traps and Macrophage Extracellular Traps Predict Postoperative Recurrence in Resectable Nonfunctional Pancreatic Neuroendocrine Tumors. Front. Immunol. 2021, 12, 577517. [Google Scholar] [CrossRef]
- Zhang, Y.; Chandra, V.; Riquelme Sanchez, E.; Dutta, P.; Quesada, P.R.; Rakoski, A.; Zoltan, M.; Arora, N.; Baydogan, S.; Horne, W.; et al. Interleukin-17-induced neutrophil extracellular traps mediate resistance to checkpoint blockade in pancreatic cancer. J. Exp. Med. 2020, 217, e20190354. [Google Scholar] [CrossRef]
- Paweletz, N. Walther Flemming: Pioneer of mitosis research. Nat. Rev. Mol. Cell Biol. 2001, 2, 72–75. [Google Scholar] [CrossRef]
- Shelanski, M.L.; Taylor, E.W. Properties of the protein subunit of central-pair and outer-doublet microtubules of sea urchin flagella. J. Cell Biol. 1968, 38, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Gadde, S.; Heald, R. Mechanisms and molecules of the mitotic spindle. Curr. Biol. 2004, 14, R797–R805. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, E.; Ho, T.; Kirschner, M.W. The role of microtubule dynamics in growth cone motility and axonal growth. J. Cell Biol. 1995, 128, 139–155. [Google Scholar] [CrossRef]
- Wordeman, L. Microtubule-depolymerizing kinesins. Curr. Opin. Cell Biol. 2005, 17, 82–88. [Google Scholar] [CrossRef]
- Jun, D.Y.; Park, H.S.; Lee, J.Y.; Kim, Y.H. Regulation of the human mitotic centromere-associated kinesin (MCAK) promoter by the transcription factors Sp1 and E2F1. Biochim. Biophys. Acta 2008, 1779, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.T.; Rankin, K.E.; von Dassow, G.; Peris, L.; Wagenbach, M.; Ovechkina, Y.; Andrieux, A.; Job, D.; Wordeman, L. MCAK associates with the tips of polymerizing microtubules. J. Cell Biol. 2005, 169, 391–397. [Google Scholar] [CrossRef]
- Tanenbaum, M.E.; Medema, R.H.; Akhmanova, A. Regulation of localization and activity of the microtubule depolymerase MCAK. Bioarchitecture 2011, 1, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Braun, A.; Dang, K.; Buslig, F.; Baird, M.A.; Davidson, M.W.; Waterman, C.M.; Myers, K.A. Rac1 and Aurora A regulate MCAK to polarize microtubule growth in migrating endothelial cells. J. Cell Biol. 2014, 206, 97–112. [Google Scholar] [CrossRef] [Green Version]
- Cooper, J.R.; Wagenbach, M.; Asbury, C.L.; Wordeman, L. Catalysis of the microtubule on-rate is the major parameter regulating the depolymerase activity of MCAK. Nat. Struct. Mol. Biol. 2010, 17, 77–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hertzer, K.M.; Ems-McClung, S.C.; Kline-Smith, S.L.; Lipkin, T.G.; Gilbert, S.P.; Walczak, C.E. Full-length dimeric MCAK is a more efficient microtubule depolymerase than minimal domain monomeric MCAK. Mol. Biol. Cell 2006, 17, 700–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maney, T.; Wagenbach, M.; Wordeman, L. Molecular dissection of the microtubule depolymerizing activity of mitotic centromere-associated kinesin. J. Biol. Chem. 2001, 276, 34753–34758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talapatra, S.K.; Harker, B.; Welburn, J.P. The C-terminal region of the motor protein MCAK controls its structure and activity through a conformational switch. eLife 2015, 4, e06421. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Shen, T.; Guerois, R.; Zhang, F.; Kuerban, H.; Lv, Y.; Gigant, B.; Knossow, M.; Wang, C. New Insights into the Coupling between Microtubule Depolymerization and ATP Hydrolysis by Kinesin-13 Protein Kif2C. J. Biol. Chem. 2015, 290, 18721–18731. [Google Scholar] [CrossRef] [Green Version]
- Eichenlaub-Ritter, U. Microtubule dynamics and tumor invasion involving MCAK. Cell Cycle 2015, 14, 3353. [Google Scholar] [CrossRef] [Green Version]
- Ritter, A.; Sanhaji, M.; Friemel, A.; Roth, S.; Rolle, U.; Louwen, F.; Yuan, J. Functional analysis of phosphorylation of the mitotic centromere-associated kinesin by Aurora B kinase in human tumor cells. Cell Cycle 2015, 14, 3755–3767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wordeman, L.; Mitchison, T.J. Identification and partial characterization of mitotic centromere-associated kinesin, a kinesin-related protein that associates with centromeres during mitosis. J. Cell Biol. 1995, 128, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Li, T.F.; Zeng, H.J.; Shan, Z.; Ye, R.Y.; Cheang, T.Y.; Zhang, Y.J.; Lu, S.H.; Zhang, Q.; Shao, N.; Lin, Y. Overexpression of kinesin superfamily members as prognostic biomarkers of breast cancer. Cancer Cell Int. 2020, 20, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, S.; Dai, M.; Zhang, C.; Teng, K.; Wang, F.; Li, H.; Sun, W.; Feng, Z.; Kang, T.; Guan, X.; et al. KIF2C: A novel link between Wnt/β-catenin and mTORC1 signaling in the pathogenesis of hepatocellular carcinoma. Protein Cell 2021, 12, 788–809. [Google Scholar] [CrossRef]
- Gnjatic, S.; Cao, Y.; Reichelt, U.; Yekebas, E.F.; Nölker, C.; Marx, A.H.; Erbersdobler, A.; Nishikawa, H.; Hildebrandt, Y.; Bartels, K.; et al. NY-CO-58/KIF2C is overexpressed in a variety of solid tumors and induces frequent T cell responses in patients with colorectal cancer. Int. J. Cancer 2010, 127, 381–393. [Google Scholar] [CrossRef]
- Honnappa, S.; Gouveia, S.M.; Weisbrich, A.; Damberger, F.F.; Bhavesh, N.S.; Jawhari, H.; Grigoriev, I.; van Rijssel, F.J.; Buey, R.M.; Lawera, A.; et al. An EB1-binding motif acts as a microtubule tip localization signal. Cell 2009, 138, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Maney, T.; Hunter, A.W.; Wagenbach, M.; Wordeman, L. Mitotic centromere-associated kinesin is important for anaphase chromosome segregation. J. Cell Biol. 1998, 142, 787–801. [Google Scholar] [CrossRef] [Green Version]
- Walczak, C.E.; Mitchison, T.J.; Desai, A. XKCM1: A Xenopus kinesin-related protein that regulates microtubule dynamics during mitotic spindle assembly. Cell 1996, 84, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Ems-McClung, S.C.; Walczak, C.E. Kinesin-13s in mitosis: Key players in the spatial and temporal organization of spindle microtubules. Semin. Cell Dev. Biol. 2010, 21, 276–282. [Google Scholar] [CrossRef] [Green Version]
- Holmfeldt, P.; Stenmark, S.; Gullberg, M. Differential functional interplay of TOGp/XMAP215 and the KinI kinesin MCAK during interphase and mitosis. EMBO J. 2004, 23, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.C.; Kim, N.H. Spindle assembly checkpoint and its regulators in meiosis. Hum. Reprod. Update 2012, 18, 60–72. [Google Scholar] [CrossRef] [Green Version]
- Kline-Smith, S.L.; Khodjakov, A.; Hergert, P.; Walczak, C.E. Depletion of centromeric MCAK leads to chromosome congression and segregation defects due to improper kinetochore attachments. Mol. Biol. Cell 2004, 15, 1146–1159. [Google Scholar] [CrossRef] [Green Version]
- Wordeman, L.; Wagenbach, M.; von Dassow, G. MCAK facilitates chromosome movement by promoting kinetochore microtubule turnover. J. Cell Biol. 2007, 179, 869–879. [Google Scholar] [CrossRef]
- Illingworth, C.; Pirmadjid, N.; Serhal, P.; Howe, K.; Fitzharris, G. MCAK regulates chromosome alignment but is not necessary for preventing aneuploidy in mouse oocyte meiosis I. Development 2010, 137, 2133–2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohi, R.; Burbank, K.; Liu, Q.; Mitchison, T.J. Nonredundant functions of Kinesin-13s during meiotic spindle assembly. Curr. Biol. 2007, 17, 953–959. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Paydar, M.; Wang, F.; Li, Y.; Wang, L.; Barrette, B.; Bessho, T.; Kwok, B.H.; Peng, A. Kinesin Kif2C in regulation of DNA double strand break dynamics and repair. eLife 2020, 9, e53402. [Google Scholar] [CrossRef] [PubMed]
- Negrini, S.; Gorgoulis, V.G.; Halazonetis, T.D. Genomic instability—An evolving hallmark of cancer. Nat. Rev. Mol. Cell Biol. 2010, 11, 220–228. [Google Scholar] [CrossRef]
- O’Connor, M.J. Targeting the DNA Damage Response in Cancer. Mol. Cell 2015, 60, 547–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roos, W.P.; Thomas, A.D.; Kaina, B. DNA damage and the balance between survival and death in cancer biology. Nat. Rev. Cancer 2016, 16, 20–33. [Google Scholar] [CrossRef]
- Lord, C.J.; Ashworth, A. The DNA damage response and cancer therapy. Nature 2012, 481, 287–294. [Google Scholar] [CrossRef]
- An, L.; Zhang, J.; Feng, D.; Zhao, Y.; Ouyang, W.; Shi, R.; Zhou, X.; Yu, Z.; Wei, S.; Min, J.; et al. KIF2C Is a Novel Prognostic Biomarker and Correlated with Immune Infiltration in Endometrial Cancer. Stem Cells Int. 2021, 2021, 1434856. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wu, Z.; Yang, L.; Jeong, J.H.; Zhu, Y.; Lu, J.; Wang, B.; Wang, N.; Wang, Y.; Shen, K.; et al. Characterization of Kinesin Family Member 2C as a Proto-Oncogene in Cervical Cancer. Front. Pharmacol. 2022, 12, 785981. [Google Scholar] [CrossRef]
- Mo, S.; Fang, D.; Zhao, S.; Thai Hoa, P.T.; Zhou, C.; Liang, T.; He, Y.; Yu, T.; Chen, Y.; Qin, W.; et al. Down regulated oncogene KIF2C inhibits growth, invasion, and metastasis of hepatocellular carcinoma through the Ras/MAPK signaling pathway and epithelial-to-mesenchymal transition. Ann. Transl. Med. 2022, 10, 151. [Google Scholar] [CrossRef]
- Cheng, S.; Castillo, V.; Sliva, D. CDC20 associated with cancer metastasis and novel mushroom-derived CDC20 inhibitors with antimetastatic activity. Int. J. Oncol. 2019, 54, 2250–2256. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xue, Y.B.; Li, H.; Qiu, D.; Wang, Z.W.; Tan, S.S. Inhibition of Cell Survival by Curcumin Is Associated with Downregulation of Cell Division Cycle 20 (Cdc20) in Pancreatic Cancer Cells. Nutrients 2017, 9, 109. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.Z.; Ma, Y.; Ji, B.; Liu, Y.; Hwu, P.; Abbruzzese, J.L.; Logsdon, C.; Wang, H. Increased CDC20 expression is associated with pancreatic ductal adenocarcinoma differentiation and progression. J. Hematol. Oncol. 2012, 5, 15. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Variables | n | KIF2C Expression | p Value | |
|---|---|---|---|---|
| Low/Moderate | High | |||
| All cases | 70 | 42 | 28 | |
| Age (years) | ||||
| <60 | 32 | 18 | 14 | |
| ≥60 | 38 | 16 | 22 | 0.643 |
| Gender | ||||
| Male | 35 | 23 | 12 | |
| Female | 35 | 16 | 20 | 0.111 |
| TNM stage | ||||
| IA | 27 | 17 | 10 | |
| IB | 17 | 8 | 9 | |
| IIA | 0 | 0 | 0 | |
| B | 11 | 4 | 7 | |
| III | 0 | 0 | 0 | |
| IV | 14 | 4 | 10 | 0.028 (<0.05) |
| Differentiated degree | ||||
| Low/moderate | 46 | 17 | 29 | |
| High | 24 | 17 | 7 | 0.022 (<0.05) |
| Drinking history | ||||
| Yes | 21 | 12 | 9 | |
| No | 49 | 22 | 27 | 0.412 |
| Obstructive jaundice | ||||
| Yes | 39 | 17 | 22 | |
| No | 31 | 17 | 14 | 0.420 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Zhao, F.; Wu, Q.; Wang, Z.; Ren, H.; Zhang, Q.; Wang, Z.; Xu, J. KIF2C Facilitates Tumor Growth and Metastasis in Pancreatic Ductal Adenocarcinoma. Cancers 2023, 15, 1502. https://doi.org/10.3390/cancers15051502
Huang X, Zhao F, Wu Q, Wang Z, Ren H, Zhang Q, Wang Z, Xu J. KIF2C Facilitates Tumor Growth and Metastasis in Pancreatic Ductal Adenocarcinoma. Cancers. 2023; 15(5):1502. https://doi.org/10.3390/cancers15051502
Chicago/Turabian StyleHuang, Xing, Feng Zhao, Quan Wu, Zitong Wang, Haiyue Ren, Qiqi Zhang, Zhe Wang, and Jin Xu. 2023. "KIF2C Facilitates Tumor Growth and Metastasis in Pancreatic Ductal Adenocarcinoma" Cancers 15, no. 5: 1502. https://doi.org/10.3390/cancers15051502
APA StyleHuang, X., Zhao, F., Wu, Q., Wang, Z., Ren, H., Zhang, Q., Wang, Z., & Xu, J. (2023). KIF2C Facilitates Tumor Growth and Metastasis in Pancreatic Ductal Adenocarcinoma. Cancers, 15(5), 1502. https://doi.org/10.3390/cancers15051502