

Beyond Avoiding Hemiplegia after Glioma Surgery: The Need to Map Complex Movement in Awake Patient to Preserve Conation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
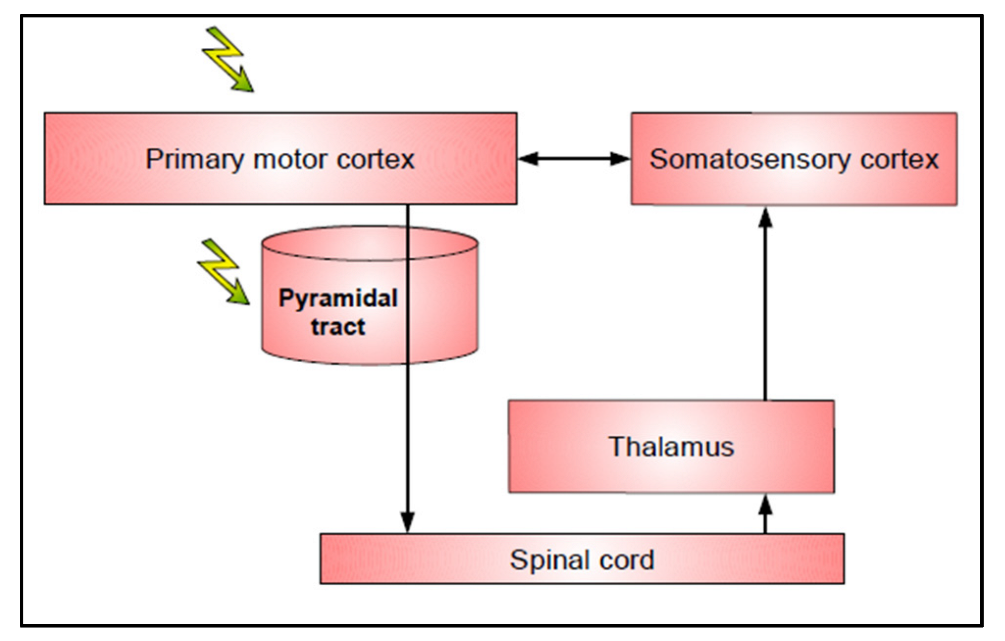
2. First Level: Muscle Contraction and Motor Output
3. Second Level: Movement Coordination and Control
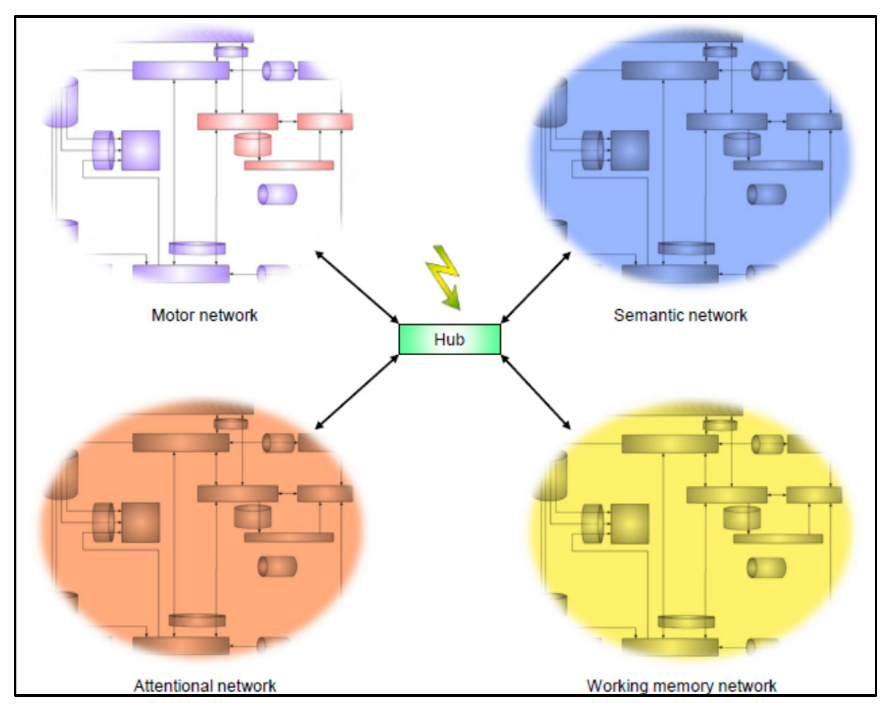
4. The Most Integrated Level: To Action and Conation
5. How to Tailor Resection
6. Conclusions and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Jackson, J.H. On The Anatomical & Physiological Localisation of Movements In The Brain. Lancet 1873, 101, 162–164. [Google Scholar] [CrossRef]
- Dum, R.P.; Strick, P.L. Motor areas in the frontal lobe of the primate. Physiol. Behav. 2002, 77, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Dum, R.P.; Strick, P.L. Spinal Cord Terminations of the Medial Wall Motor Areas in Macaque Monkeys. J. Neurosci. 1996, 16, 6513–6525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dum, R.P.; Strick, P.L. The origin of corticospinal projections from the premotor areas in the frontal lobe. J. Neurosci. 1991, 11, 667–689. [Google Scholar] [CrossRef] [Green Version]
- Graziano, M.S.; Taylor, C.S.; Moore, T.; Cooke, D. The Cortical Control of Movement Revisited. Neuron 2002, 36, 349–362. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Dum, R.; Strick, P. Topographic organization of corticospinal projections from the frontal lobe: Motor areas on the medial surface of the hemisphere. J. Neurosci. 1995, 15, 3284–3306. [Google Scholar] [CrossRef]
- Picard, N.; Strick, P.L. Motor Areas of the Medial Wall: A Review of Their Location and Functional Activation. Cereb. Cortex 1996, 6, 342–353. [Google Scholar] [CrossRef] [Green Version]
- Caminiti, R.; Borra, E.; Comandini, F.V.; Battaglia-Mayer, A.; Averbeck, B.B.; Luppino, G. Computational Architecture of the Parieto-Frontal Network Underlying Cognitive-Motor Control in Monkeys. Eneuro 2017, 4, 1–35. [Google Scholar] [CrossRef]
- De Benedictis, A.; Duffau, H. Brain Hodotopy: From Esoteric Concept to Practical Surgical Applications. Neurosurgery 2011, 68, 1709–1723. [Google Scholar] [CrossRef] [Green Version]
- Ball, T.; Schreiber, A.; Feige, B.; Wagner, M.; Lücking, C.H.; Kristeva-Feige, R. The Role of Higher-Order Motor Areas in Voluntary Movement as Revealed by High-Resolution EEG and fMRI. Neuroimage 1999, 10, 682–694. [Google Scholar] [CrossRef] [Green Version]
- Cunnington, R.; Windischberger, C.; Deecke, L.; Moser, E. The Preparation and Execution of Self-Initiated and Externally-Triggered Movement: A Study of Event-Related fMRI. Neuroimage 2002, 15, 373–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallivan, J.P.; Culham, J.C. Neural coding within human brain areas involved in actions. Curr. Opin. Neurobiol. 2015, 33, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Pool, E.-M.; Rehme, A.K.; Eickhoff, S.B.; Fink, G.R.; Grefkes, C. Functional resting-state connectivity of the human motor network: Differences between right- and left-handers. Neuroimage 2015, 109, 298–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bestmann, S.; Duque, J. Transcranial Magnetic Stimulation: Decomposing the Processes Underlying Action Preparation. Neuroscientist 2015, 22, 392–405. [Google Scholar] [CrossRef] [PubMed]
- Penfield, W.; Boldrey, E. Somatic Motor and Sensory Representation in The Cerebral Cortex of Man As Studied by Electrical Stimulation. Brain 1937, 60, 389–443. [Google Scholar] [CrossRef]
- Brinkman, C. Supplementary motor area of the monkey’s cerebral cortex: Short- and long-term deficits after unilateral ablation and the effects of subsequent callosal section. J. Neurosci. 1984, 4, 918–929. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-H.; Kim, C.H.; Kim, J.S.; Lee, S.K.; Han, J.H.; Kim, C.-Y.; Chung, C.K. Risk factor analysis of the development of new neurological deficits following supplementary motor area resection. J. Neurosurg. 2013, 119, 7–14. [Google Scholar] [CrossRef]
- Krainik, A.; Lehericy, S.; Duffau, H.; Vlaicu, M.; Poupon, F.; Capelle, L.; Cornu, P.; Clemenceau, S.; Sahel, M.; Valery, C.-A.; et al. Role of the supplementary motor area in motor deficit following medial frontal lobe surgery. Neurology 2001, 57, 871–878. [Google Scholar] [CrossRef] [Green Version]
- Krainik, A.; Lehericy, S.; Duffau, H.; Capelle, L.; Chainay, H.; Cornu, P.; Cohen, L.; Boch, A.L.; Mangin, J.F.; Le Bihan, D.; et al. Postoperative Speech Disorder after Medial Frontal Surgery Role of the Supplementary Motor Area. Neurology 2003, 60, 587–594. [Google Scholar] [CrossRef]
- Russell, S.M.; Kelly, P.J. Incidence and Clinical Evolution of Postoperative Deficits after Volumetric Stereotactic Resection of Glial Neoplasms Involving the Supplementary Motor Area. Neurosurgery 2003, 52, 506–516. [Google Scholar] [CrossRef]
- Rech, F.; Herbet, G.; Moritz-Gasser, S.; Duffau, H. Somatotopic organization of the white matter tracts underpinning motor control in humans: An electrical stimulation study. Brain Struct. Funct. 2015, 221, 3743–3753. [Google Scholar] [CrossRef]
- Rossi, M.; Fornia, L.; Puglisi, G.; Leonetti, A.; Zuccon, G.; Fava, E.; Milani, D.; Casarotti, A.; Riva, M.; Pessina, F.; et al. Assessment of the praxis circuit in glioma surgery to reduce the incidence of postoperative and long-term apraxia: A new intraoperative test. J. Neurosurg. 2018, 130, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Duffau, H.; Ng, S.; Lemaitre, A.-L.; Moritz-Gasser, S.; Herbet, G. Constant Multi-Tasking with Time Constraint to Preserve Across-Network Dynamics throughout Awake Surgery for Low-Grade Glioma: A Necessary Step to Enable Patients Resuming an Active Life. Front. Oncol. 2022, 12, 924762. [Google Scholar] [CrossRef]
- Taillandier, L.; Obara, T.; Duffau, H. What Does Quality of Care Mean in Lower-Grade Glioma Patients: A Precision Molecular-Based Management of the Tumor or an Individualized Medicine Centered on Patient’s Choices? Front. Oncol. 2021, 11, 719014. [Google Scholar] [CrossRef]
- Rossi, M.; Viganò, L.; Puglisi, G.; Nibali, M.C.; Leonetti, A.; Gay, L.; Sciortino, T.; Fornia, L.; Callipo, V.; Lamperti, M.; et al. Targeting Primary Motor Cortex (M1) Functional Components in M1 Gliomas Enhances Safe Resection and Reveals M1 Plasticity Potentials. Cancers 2021, 13, 3808. [Google Scholar] [CrossRef]
- Rossi, M.; Puglisi, G.; Nibali, M.C.; Viganò, L.; Sciortino, T.; Gay, L.; Leonetti, A.; Zito, P.; Riva, M.; Bello, L. Asleep or awake motor mapping for resection of perirolandic glioma in the nondominant hemisphere? Development and validation of a multimodal score to tailor the surgical strategy. J. Neurosurg. 2022, 136, 16–29. [Google Scholar] [CrossRef]
- Duffau, H. What Direct Electrostimulation of the Brain Taught Us About the Human Connectome: A Three-Level Model of Neural Disruption. Front. Hum. Neurosci. 2020, 14, 315. [Google Scholar] [CrossRef]
- Duffau, H. Acute functional reorganisation of the human motor cortex during resection of central lesions: A study using intraoperative brain mapping. J. Neurol. Neurosurg. Psychiatry 2001, 70, 506–513. [Google Scholar] [CrossRef] [Green Version]
- Ius, T.; Angelini, E.; de Schotten, M.T.; Mandonnet, E.; Duffau, H. Evidence for potentials and limitations of brain plasticity using an atlas of functional resectability of WHO grade II gliomas: Towards a “minimal common brain”. Neuroimage 2011, 56, 992–1000. [Google Scholar] [CrossRef]
- Duffau, H. A personal consecutive series of surgically treated 51 cases of insular WHO Grade II glioma: Advances and limitations. J. Neurosurg. 2009, 110, 696–708. [Google Scholar] [CrossRef] [Green Version]
- Laplane, D.; Talairach, J.; Meininger, V.; Bancaud, J.; Orgogozo, J. Clinical consequences of corticectomies involving the supplementary motor area in man. J. Neurol. Sci. 1977, 34, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Nachev, P.; Wydell, H.; O’Neill, K.; Husain, M.; Kennard, C. The role of the pre-supplementary motor area in the control of action. NeuroImage 2007, 36, T155–T163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krainik, A.; Duffau, H.; Capelle, L.; Cornu, P.; Boch, A.L.; Mangin, J.F.; Le Bihan, D.; Marsault, C.; Chiras, J.; Lehéricy, S. Role of the healthy hemisphere in recovery after resection of the supplementary motor area. Neurology 2004, 62, 1323–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassal, M.; Charroud, C.; Deverdun, J.; Le Bars, E.; Molino, F.; Bonnetblanc, F.; Boyer, A.; Dutta, A.; Herbet, G.; Moritz-Gasser, S.; et al. Recovery of functional connectivity of the sensorimotor network after surgery for diffuse low-grade gliomas involving the supplementary motor area. J. Neurosurg. 2017, 126, 1181–1190. [Google Scholar] [CrossRef] [Green Version]
- Giampiccolo, D.; Parisi, C.; Meneghelli, P.; Tramontano, V.; Basaldella, F.; Pasetto, M.; Pinna, G.; Cattaneo, L.; Sala, F. Long-term motor deficit in brain tumour surgery with preserved intra-operative motor-evoked potentials. Brain Commun. 2021, 3, fcaa226. [Google Scholar] [CrossRef]
- Yazawa, S.; Ikeda, A.; Kunieda, T.; Mima, T.; Nagamine, T.; Ohara, S.; Terada, K.; Taki, W.; Kimura, J.; Shibasaki, H. Human supplementary motor area is active in preparation for both voluntary muscle relaxation and contraction: Subdural recording of Bereitschaftspotential. Neurosci. Lett. 1998, 244, 145–148. [Google Scholar] [CrossRef]
- Gerloff, C.; Corwell, B.; Chen, R.; Hallett, M.; Cohen, L.G. Stimulation over the human supplementary motor area interferes with the organization of future elements in complex motor sequences. Brain 1997, 120, 1587–1602. [Google Scholar] [CrossRef] [Green Version]
- Deiber, M.-P.; Honda, M.; Ibañez, V.; Sadato, N.; Hallett, M. Mesial Motor Areas in Self-Initiated Versus Externally Triggered Movements Examined with fMRI: Effect of Movement Type and Rate. J. Neurophysiol. 1999, 81, 3065–3077. [Google Scholar] [CrossRef] [Green Version]
- Hoffstaedter, F.; Grefkes, C.; Zilles, K.; Eickhoff, S.B. The “What” and “When” of Self-Initiated Movements. Cereb. Cortex 2012, 23, 520–530. [Google Scholar] [CrossRef] [Green Version]
- Shima, K.; Tanji, J. Neuronal Activity in the Supplementary and Presupplementary Motor Areas for Temporal Organization of Multiple Movements. J. Neurophysiol. 2000, 84, 2148–2160. [Google Scholar] [CrossRef] [Green Version]
- Shima, K.; Tanji, J. Both Supplementary and Presupplementary Motor Areas Are Crucial for the Temporal Organization of Multiple Movements. J. Neurophysiol. 1998, 80, 3247–3260. [Google Scholar] [CrossRef]
- Tanji, J. Sequential Organization of Multiple Movements: Involvement of Cortical Motor Areas. Annu. Rev. Neurosci. 2001, 24, 631–651. [Google Scholar] [CrossRef] [Green Version]
- Vingerhoets, G.; Nys, J.; Honoré, P.; Vandekerckhove, E.; Vandemaele, P. Human Left Ventral Premotor Cortex Mediates Matching of Hand Posture to Object Use. PLoS ONE 2013, 8, e70480. [Google Scholar] [CrossRef]
- Rojkova, K.; Volle, E.; Urbanski, M.; Humbert, F.; Dell’Acqua, F.; De Schotten, M.T. Atlasing the frontal lobe connections and their variability due to age and education: A spherical deconvolution tractography study. Brain Struct. Funct. 2016, 221, 1751–1766. [Google Scholar] [CrossRef]
- Penfield, W. The supplementary motor area in the cerebral cortex of man. Rchiv Psychiatr. Nervenkrankh. Ver. Mit Z. Für Die Gesamte Neurol. Psychiatr. 1950, 185, 670–674. [Google Scholar] [CrossRef]
- Lüders, H.; Lesser, R.P.; Morris, H.H.; Dinner, D.S. Negative Motor Responses Elicited by Stimulation of the Human Cortex. In Advances in Epileptology; Wolf, P., Dam, M., Janz, D., Eds.; Raven Press: New York, NY, USA, 1987; pp. 229–231. [Google Scholar]
- Filevich, E.; Kühn, S.; Haggard, P. Negative motor phenomena in cortical stimulation: Implications for inhibitory control of human action. Cortex 2012, 48, 1251–1261. [Google Scholar] [CrossRef]
- Lüders, H.O.; Lüders, H.; Dinner, D.S.; Morris, H.H.; Wyllie, E.; Comair, Y.G. Cortical electrical stimulation in humans. The negative motor areas. Adv. Neurol. 1995, 67, 115–129. [Google Scholar]
- Mikuni, N.; Ohara, S.; Ikeda, A.; Hayashi, N.; Nishida, N.; Taki, J.; Enatsu, R.; Matsumoto, R.; Shibasaki, H.; Hashimoto, N. Evidence for a wide distribution of negative motor areas in the perirolandic cortex. Clin. Neurophysiol. 2006, 117, 33–40. [Google Scholar] [CrossRef]
- Borggraefe, I.; Catarino, C.B.; Rémi, J.; Vollmar, C.; Peraud, A.; Winkler, P.A.; Noachtar, S. Lateralization of cortical negative motor areas. Clin. Neurophysiol. 2016, 127, 3314–3321. [Google Scholar] [CrossRef]
- Duffau, H. Does post-lesional subcortical plasticity exist in the human brain? Neurosci. Res. 2009, 65, 131–135. [Google Scholar] [CrossRef]
- Rech, F.; Herbet, G.; Gaudeau, Y.; Mézières, S.; Moureau, J.-M.; Moritz-Gasser, S.; Duffau, H. A probabilistic map of negative motor areas of the upper limb and face: A brain stimulation study. Brain 2019, 142, 952–965. [Google Scholar] [CrossRef] [PubMed]
- Trevisi, G.; Eickhoff, S.B.; Chowdhury, F.; Jha, A.; Rodionov, R.; Nowell, M.; Miserocchi, A.; McEvoy, A.W.; Nachev, P.; Diehl, B. Probabilistic electrical stimulation mapping of human medial frontal cortex. Cortex 2018, 109, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Viganò, L.; Howells, H.; Fornia, L.; Rossi, M.; Nibali, M.C.; Puglisi, G.; Leonetti, A.; Simone, L.; Bello, L.; Cerri, G. Negative motor responses to direct electrical stimulation: Behavioral assessment hides different effects on muscles. Cortex 2021, 137, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Schucht, P.; Moritz-Gasser, S.; Herbet, G.; Raabe, A.; Duffau, H. Subcortical electrostimulation to identify network subserving motor control. Hum. Brain Mapp. 2012, 34, 3023–3030. [Google Scholar] [CrossRef] [PubMed]
- Rech, F.; Duffau, H.; Pinelli, C.; Masson, A.; Roublot, P.; Billy-Jacques, A.; Brissart, H.; Civit, T. Intraoperative identification of the negative motor network during awake surgery to prevent deficit following brain resection in premotor regions. Neurochirurgie 2017, 63, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Duffau, H. New Philosophy, Clinical Pearls, and Methods for Intraoperative Cognition Mapping and Monitoring “à la carte” in Brain Tumor Patients. Neurosurgery 2021, 88, 919–930. [Google Scholar] [CrossRef]
- Rech, F.; Herbet, G.; Moritz-Gasser, S.; Duffau, H. Disruption of bimanual movement by unilateral subcortical electrostimulation. Hum. Brain Mapp. 2013, 35, 3439–3445. [Google Scholar] [CrossRef]
- Schubotz, R.I.; Anwander, A.; Knösche, T.R.; von Cramon, D.Y.; Tittgemeyer, M. Anatomical and functional parcellation of the human lateral premotor cortex. Neuroimage 2010, 50, 396–408. [Google Scholar] [CrossRef] [Green Version]
- Graziano, M.S. Ethological Action Maps: A Paradigm Shift for the Motor Cortex. Trends Cogn. Sci. 2016, 20, 121–132. [Google Scholar] [CrossRef]
- Catani, M. A little man of some importance. Brain 2017, 140, 3055–3061. [Google Scholar] [CrossRef] [Green Version]
- Nachev, P.; Kennard, C.; Husain, M. Functional role of the supplementary and pre-supplementary motor areas. Nat. Rev. Neurosci. 2008, 9, 856–869. [Google Scholar] [CrossRef]
- Geyer, S.; Matelli, M.; Luppino, G.; Zilles, K. Functional neuroanatomy of the primate isocortical motor system. Anat. Embryol. 2000, 202, 443–474. [Google Scholar] [CrossRef]
- Dum, R.P.; Strick, P.L. Frontal Lobe Inputs to the Digit Representations of the Motor Areas on the Lateral Surface of the Hemisphere. J. Neurosci. 2005, 25, 1375–1386. [Google Scholar] [CrossRef] [Green Version]
- Catani, M.; Dell’Acqua, F.; Vergani, F.; Malik, F.; Hodge, H.; Roy, P.; Valabregue, R.; de Schotten, M.T. Short frontal lobe connections of the human brain. Cortex 2012, 48, 273–291. [Google Scholar] [CrossRef]
- Zhang, R.; Geng, X.; Lee, T.M.C. Large-scale functional neural network correlates of response inhibition: An fMRI meta-analysis. Anat. Embryol. 2017, 222, 3973–3990. [Google Scholar] [CrossRef] [Green Version]
- Aron, A.R.; Herz, D.M.; Brown, P.; Forstmann, B.U.; Zaghloul, K. Frontosubthalamic Circuits for Control of Action and Cognition. J. Neurosci. 2016, 36, 11489–11495. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, M.; de Champfleur, N.M.; Deverdun, J.; Moritz-Gasser, S.; Herbet, G.; Duffau, H. Role of fronto-striatal tract and frontal aslant tract in movement and speech: An axonal mapping study. Anat. Embryol. 2014, 220, 3399–3412. [Google Scholar] [CrossRef]
- Vergani, F.; Lacerda, L.M.; Martino, J.; Attems, J.; Morris, C.; Mitchell, P.; de Schotten, M.T.; Dell’Acqua, F. White matter connections of the supplementary motor area in humans. J. Neurol. Neurosurg. Psychiatry 2014, 85, 1377–1385. [Google Scholar] [CrossRef]
- Fontaine, D.; Capelle, L.; Duffau, H. Somatotopy of the Supplementary Motor Area: Evidence from Correlation of the Extent of Surgical Resection with the Clinical Patterns of Deficit. Neurosurgery 2002, 50, 297–305. [Google Scholar]
- Mitz, A.R.; Wise, S.P. The somatotopic organization of the supplementary motor area: Intracortical microstimulation mapping. J. Neurosci. 1987, 7, 1010–1021. [Google Scholar] [CrossRef] [Green Version]
- Tanné-Gariépy, J.; Rouiller, E.M.; Boussaoud, D. Parietal inputs to dorsal versus ventral premotor areas in the macaque monkey: Evidence for largely segregated visuomotor pathways. Exp. Brain Res. 2002, 145, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.B.; Ferraina, S.; Bianchi, L.; Caminiti, R. Cortical Networks for Visual Reaching: Physiological and Anatomical Organization of Frontal and Parietal Lobe Arm Regions. Cereb. Cortex 1996, 6, 102–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesaran, B.; Nelson, M.J.; Andersen, R.A. Dorsal Premotor Neurons Encode the Relative Position of the Hand, Eye, and Goal during Reach Planning. Neuron 2006, 51, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Hoshi, E.; Tanji, J. Distinctions between dorsal and ventral premotor areas: Anatomical connectivity and functional properties. Curr. Opin. Neurobiol. 2007, 17, 234–242. [Google Scholar] [CrossRef]
- Borra, E.; Luppino, G. Functional anatomy of the macaque temporo-parieto-frontal connectivity. Cortex 2017, 97, 306–326. [Google Scholar] [CrossRef] [PubMed]
- Davare, M.; Kraskov, A.; Rothwell, J.C.; Lemon, R.N. Interactions between areas of the cortical grasping network. Curr. Opin. Neurobiol. 2011, 21, 565–570. [Google Scholar] [CrossRef] [Green Version]
- Rizzolatti, G.; Luppino, G. The Cortical Motor System. Neuron 2001, 31, 889–902. [Google Scholar] [CrossRef] [Green Version]
- Mandonnet, E.; Sarubbo, S.; Petit, L. The Nomenclature of Human White Matter Association Pathways: Proposal for a Systematic Taxonomic Anatomical Classification. Front. Neuroanat. 2018, 12, 94. [Google Scholar] [CrossRef] [Green Version]
- Almairac, F.; Herbet, G.; Moritz-Gasser, S.; Duffau, H. Parietal network underlying movement control: Disturbances during subcortical electrostimulation. Neurosurg. Rev. 2014, 37, 513–516, discussion 516–517. [Google Scholar] [CrossRef]
- Fornia, L.; Rossi, M.; Rabuffetti, M.; Bellacicca, A.; Viganò, L.; Simone, L.; Howells, H.; Puglisi, G.; Leonetti, A.; Callipo, V.; et al. Motor impairment evoked by direct electrical stimulation of human parietal cortex during object manipulation. Neuroimage 2022, 248, 118839. [Google Scholar] [CrossRef]
- Haggard, P. Sense of agency in the human brain. Nat. Rev. Neurosci. 2017, 18, 196–207. [Google Scholar] [CrossRef]
- Fornia, L.; Puglisi, G.; Leonetti, A.; Bello, L.; Berti, A.; Cerri, G.; Garbarini, F. Direct electrical stimulation of the premotor cortex shuts down awareness of voluntary actions. Nat. Commun. 2020, 11, 705. [Google Scholar] [CrossRef] [Green Version]
- Khalighinejad, N.; Garrett, N.; Priestley, L.; Lockwood, P.; Rushworth, M.F.S. A habenula-insular circuit encodes the willingness to act. Nat. Commun. 2021, 12, 6329. [Google Scholar] [CrossRef]
- Sumner, P.; Nachev, P.; Morris, P.; Peters, A.M.; Jackson, S.R.; Kennard, C.; Husain, M. Human Medial Frontal Cortex Mediates Unconscious Inhibition of Voluntary Action. Neuron 2007, 54, 697–711. [Google Scholar] [CrossRef] [Green Version]
- Swann, N.C.; Cai, W.; Conner, C.R.; Pieters, T.A.; Claffey, M.P.; George, J.S.; Aron, A.R.; Tandon, N. Roles for the pre-supplementary motor area and the right inferior frontal gyrus in stopping action: Electrophysiological responses and functional and structural connectivity. NeuroImage 2012, 59, 2860–2870. [Google Scholar] [CrossRef] [Green Version]
- Wolpe, N.; Hezemans, F.H.; Rae, C.L.; Zhang, J.; Rowe, J.B. The pre-supplementary motor area achieves inhibitory control by modulating response thresholds. Cortex 2022, 152, 98–108. [Google Scholar] [CrossRef]
- Thura, D.; Cisek, P. Microstimulation of dorsal premotor and primary motor cortex delays the volitional commitment to an action choice. J. Neurophysiol. 2020, 123, 927–935. [Google Scholar] [CrossRef]
- Cos, I.; Pezzulo, G.; Cisek, P. Changes of Mind after Movement Onset Depend on the State of the Motor System. Eneuro 2021, 8, 1–12. [Google Scholar] [CrossRef]
- Herbet, G.; Duffau, H. Revisiting the Functional Anatomy of the Human Brain: Toward a Meta-Networking Theory of Cerebral Functions. Physiol. Rev. 2020, 100, 1181–1228. [Google Scholar] [CrossRef]
- Pascual, J.S.G.; Duffau, H. The need to consider return to work as a main outcome in patients undergoing surgery for diffuse low-grade glioma: A systematic review. Acta Neurochir. 2022, 164, 2789–2809. [Google Scholar] [CrossRef]
- Pezzulo, G.; Cisek, P. Navigating the Affordance Landscape: Feedback Control as a Process Model of Behavior and Cognition. Trends Cogn. Sci. 2016, 20, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Herbet, G.; Duffau, H. Contribution of the medial eye field network to the voluntary deployment of visuospatial attention. Nat. Commun. 2022, 13, 328. [Google Scholar] [CrossRef] [PubMed]
- Herbet, G.; Lemaitre, A.-L.; Moritz-Gasser, S.; Cochereau, J.; Duffau, H. The antero-dorsal precuneal cortex supports specific aspects of bodily awareness. Brain 2019, 142, 2207–2214. [Google Scholar] [CrossRef] [PubMed]
- de Schotten, M.T.; Urbanski, M.; Duffau, H.; Volle, E.; Lévy, R.; Dubois, B.; Bartolomeo, P. Direct Evidence for a Parietal-Frontal Pathway Subserving Spatial Awareness in Humans. Science 2005, 309, 2226–2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battaglia-Mayer, A.; Babicola, L.; Satta, E. Parieto-frontal gradients and domains underlying eye and hand operations in the action space. Neuroscience 2016, 334, 76–92. [Google Scholar] [CrossRef]
- Duffau, H.; Mandonnet, E. The “onco-functional balance” in surgery for diffuse low-grade glioma: Integrating the extent of resection with quality of life. Acta Neurochir. 2013, 155, 951–957. [Google Scholar] [CrossRef]
- Duffau, H. Awake Mapping with Transopercular Approach in Right Insular–Centered Low-Grade Gliomas Improves Neurological Outcomes and Return to Work. Neurosurgery 2022, 91, 182–190. [Google Scholar] [CrossRef]
- Obara, T.; Blonski, M.; Brzenczek, C.; Mézières, S.; Gaudeau, Y.; Pouget, C.; Gauchotte, G.; Verger, A.; Vogin, G.; Moureaux, J.-M.; et al. Adult Diffuse Low-Grade Gliomas: 35-Year Experience at the Nancy France Neurooncology Unit. Front. Oncol. 2020, 10, 574679. [Google Scholar] [CrossRef]
- Hamdan, N.; Duffau, H. Extending the multistage surgical strategy for recurrent initially low-grade gliomas: Functional and oncological outcomes in 31 consecutive patients who underwent a third resection under awake mapping. J. Neurosurg. 2022, 136, 1035–1044. [Google Scholar] [CrossRef]
- Mandonnet, E.; Duffau, H. An attempt to conceptualize the individual onco-functional balance: Why a standardized treatment is an illusion for diffuse low-grade glioma patients. Crit. Rev. Oncol. 2018, 122, 83–91. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rech, F.; Duffau, H. Beyond Avoiding Hemiplegia after Glioma Surgery: The Need to Map Complex Movement in Awake Patient to Preserve Conation. Cancers 2023, 15, 1528. https://doi.org/10.3390/cancers15051528
Rech F, Duffau H. Beyond Avoiding Hemiplegia after Glioma Surgery: The Need to Map Complex Movement in Awake Patient to Preserve Conation. Cancers. 2023; 15(5):1528. https://doi.org/10.3390/cancers15051528
Chicago/Turabian StyleRech, Fabien, and Hugues Duffau. 2023. "Beyond Avoiding Hemiplegia after Glioma Surgery: The Need to Map Complex Movement in Awake Patient to Preserve Conation" Cancers 15, no. 5: 1528. https://doi.org/10.3390/cancers15051528
APA StyleRech, F., & Duffau, H. (2023). Beyond Avoiding Hemiplegia after Glioma Surgery: The Need to Map Complex Movement in Awake Patient to Preserve Conation. Cancers, 15(5), 1528. https://doi.org/10.3390/cancers15051528