
Golgi-Targeting Anticancer Natural Products
Abstract
:Simple Summary
Abstract

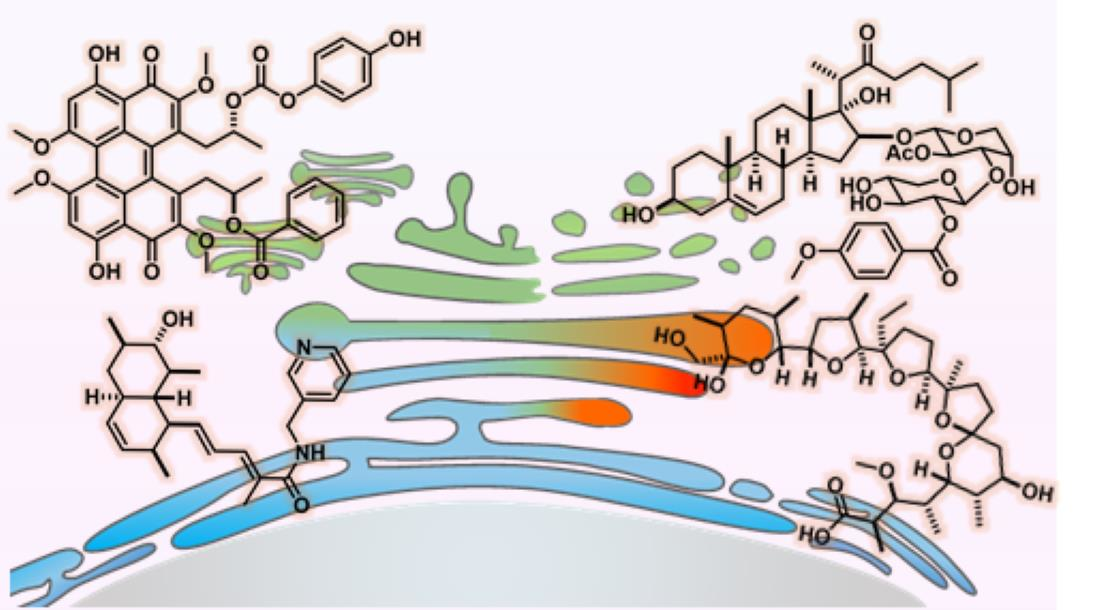{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Golgi Stress Response Pathways
3. Golgi Stress-Inducing Anticancer Natural Products
3.1. Inducers of Golgi Ribbon Disassembly/Fragmentation
3.1.1. Calphostin C
3.1.2. Bafilomycin A1
3.1.3. Tyrphostin AG-1478
3.2. Golgi-Vesiculating Agents
3.2.1. Ilimaquinone
3.2.2. OSW-1
3.2.3. Schweinfurthins
3.3. Inducers of ER–Golgi Fusion
3.3.1. Brefeldin A
3.3.2. AMF-26
3.4. Inducers of Golgi Swelling
3.4.1. Monensin
3.4.2. Nigericin
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilson, C.; Venditti, R.; Rega, L.R.; Colanzi, A.; D’Angelo, G.; De Matteis, M.A. The Golgi apparatus: An organelle with multiple complex functions. Biochem. J. 2011, 433, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schjoldager, K.T.; Narimatsu, Y.; Joshi, H.J.; Clausen, H. Global view of human protein glycosylation pathways and functions. Nat. Rev. Mol. Cell Biol. 2020, 21, 729–749. [Google Scholar] [CrossRef] [PubMed]
- De Graffenried, C.L.; Bertozzi, C.R. The roles of enzyme localization and complex formation in glycan assembly within the Golgi apparatus. Curr. Opin. Cell Biol. 2004, 16, 356–363. [Google Scholar] [CrossRef]
- Mayinger, P. Signaling at the Golgi. Cold Spring Harb. Perspect. Biol. 2011, 3, a005314. [Google Scholar] [CrossRef]
- Chia, J.; Goh, G.; Racine, V.; Ng, S.; Kumar, P.; Bard, F. RNAi screening reveals a large signaling network controlling the Golgi apparatus in human cells. Mol. Syst. Biol. 2012, 8, 629. [Google Scholar] [CrossRef]
- Kulkarni-Gosavi, P.; Makhoul, C.; Gleeson, P.A. Form and function of the Golgi apparatus: Scaffolds, cytoskeleton and signaling. FEBS Lett. 2019, 593, 2289–2305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alborzinia, H.; Ignashkova, T.I.; Dejure, F.R.; Gendarme, M.; Theobald, J.; Wölfl, S.; Lindemann, R.K.; Reiling, J.H. Golgi stress mediates redox imbalance and ferroptosis in human cells. Commun. Biol. 2018, 1, 210. [Google Scholar] [CrossRef] [Green Version]
- Zappa, F.; Failli, M.; De Matteis, M.A. The Golgi complex in disease and therapy. Curr. Opin. Cell Biol. 2018, 50, 102–116. [Google Scholar] [CrossRef]
- Li, J.; Ahat, E.; Wang, Y. Golgi Structure and Function in Health, Stress, and Diseases. Results Probl. Cell Differ. 2019, 67, 441–485. [Google Scholar]
- Bui, S.; Mejia, I.; Díaz, B.; Wang, Y. Adaptation of the Golgi apparatus in cancer cell invasion and metastasis. Front. Cell Dev. Biol. 2021, 9, 806482. [Google Scholar] [CrossRef]
- Spano, D.; Colanzim, A. Golgi Complex: A Signaling Hub in Cancer. Cells 2022, 11, 1990. [Google Scholar] [CrossRef] [PubMed]
- Dinter, A.; Berger, E.G. Golgi-disturbing agents. Histochem. Cell Biol. 1998, 109, 571–590. [Google Scholar] [CrossRef] [PubMed]
- Vanneste, M.; Huang, Q.; Li, M.; Moose, D.; Zhao, L.; Stamnes, M.A.; Schultz, M.; Wu, M.; Henry, M.D. High content screening identifies monensin as an EMT-selective cytotoxic compound. Sci. Rep. 2019, 9, 1200. [Google Scholar] [CrossRef]
- Van Meer, G.; van’t Hof, W. Epithelial sphingolipid sorting is insensitive to reorganization of the Golgi by nocodazole, but is abolished by monensin in MDCK cells and by brefeldin A in Caco-2 cells. J. Cell Sci. 1993, 104, 833–842. [Google Scholar] [CrossRef]
- Machamer, C.E. The Golgi complex in stress and death. Front. Neurosci. 2015, 9, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, K.; Yoshida, H. Golgi stress response and organelle zones. FEBS Lett. 2019, 593, 2330–2340. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, K.; Yoshida, H. Organelle autoregulation-stress responses in the ER, Golgi, mitochondria and lysosome. J. Biochem. 2015, 157, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Mori, K. The unfolded protein response: The dawn of a new field. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2015, 91, 469–480. [Google Scholar] [CrossRef] [Green Version]
- Karagoz, G.E.; Acosta-Alvear, D.; Walter, P. The unfolded protein response: Detecting and responding to fluctuations in the protein-folding capacity of the endoplasmic reticulum. Cold Spring Harb. Perspect. Biol. 2019, 11, a033886. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, H. ER stress and diseases. FEBS J. 2007, 274, 630–658. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [Green Version]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; International Natural Product Sciences Taskforce; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, L.N.; Burdette, J.E.; Carcache de Blanco, E.; Coss, C.C.; Eustaquio, A.S.; Fuchs, J.R.; Kinghorn, A.D.; MacFarlane, A.; Mize, B.K.; Oberlies, N.H.; et al. Discovery of anticancer agents of diverse natural origin. J. Nat. Prod. 2022, 85, 702–719. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Tsai, H.N.; Zhang, X.; Rosania, G.R. The subcellular distribution of small molecules: From pharmacokinetics to synthetic biology. Mol. Pharm. 2011, 8, 1619–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louzoun-Zada, S.; Jaber, Q.Z.; Fridman, M. Guiding drugs to target-harboring organelles: Stretching drug-delivery to a higher level of resolution. Angew. Chem. Int. Ed. Engl. 2019, 58, 15584–15594. [Google Scholar] [CrossRef] [PubMed]
- Ayala, I.; Colanzi, A. Structural organization and function of the Golgi ribbon during cell division. Front. Cell Dev. Biol. 2022, 10, 925228. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Wang, Y. Cell cycle regulation of Golgi membrane dynamics. Trends Cell Biol. 2013, 23, 296–304. [Google Scholar] [CrossRef] [Green Version]
- Oku, M.; Tanakura, S.; Uemura, A.; Sohda, M.; Misumi, Y.; Taniguchi, M.; Wakabayashi, S.; Yoshida, H. Novel cis-acting element GASE regulates transcriptional induction by the Golgi stress response. Cell Struct. Funct. 2011, 36, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, M.; Nadanaka, S.; Tanakura, S.; Sawaguchi, S.; Midori, S.; Kawai, Y.; Yamaguchi, S.; Shimada, Y.; Nakamura, Y.; Matsumura, Y.; et al. TFE3 is a bHLH-ZIP-type transcription factor that regulates the mammalian Golgi stress response. Cell Struct. Funct. 2015, 40, 13–30. [Google Scholar] [CrossRef] [Green Version]
- Reiling, J.H.; Olive, A.J.; Sanyal, S.; Carette, J.E.; Brummelkamp, T.R.; Ploegh, H.L.; Starnbach, M.N.; Sabatini, D.M. A CREB3-ARF4 signalling pathway mediates the response to Golgi stress and susceptibility to pathogens. Nat. Cell Biol. 2013, 15, 1473–1485. [Google Scholar] [CrossRef] [Green Version]
- Miyata, S.; Mizuno, T.; Koyama, Y.; Katayama, T.; Tohyama, M. The endoplasmic reticulum-resident chaperone heat shock protein 47 protects the Golgi apparatus from the effects of O-glycosylation inhibition. PLoS ONE 2013, 8, e69732. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Komori, R.; Taniguchi, M.; Shimaoka, A.; Midori, S.; Yamamoto, M.; Okuda, C.; Tanaka, R.; Sakamoto, M.; Wakabayashi, S.; et al. PGSE Is a Novel Enhancer Regulating the Proteoglycan Pathway of the Mammalian Golgi Stress Response. Cell Struct. Funct. 2019, 44, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumann, J.; Ignashkova, T.I.; Chirasani, S.R.; Ramirez-Peinado, S.; Alborzinia, H.; Gendarme, M.; Kuhnigk, K.; Kramer, V.; Lindemann, R.K.; Reiling, J.H. Golgi stress-induced transcriptional changes mediated by MAPK signaling and three ETS transcription factors regulate MCL1 splicing. Mol. Biol. Cell 2018, 29, 42–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ignashkova, T.I.; Gendarme, M.; Peschk, K.; Eggenweiler, H.M.; Lindemann, R.K.; Reiling, J.H. Cell survival and protein secretion associated with Golgi integrity in response to Golgi stress-inducing agents. Traffic 2017, 18, 530–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh-Hashi, K.; Hasegawa, T.; Mizutani, Y.; Takahashi, K.; Hirata, Y. Elucidation of brefeldin A-induced ER and Golgi stress responses in Neuro2a cells. Mol. Cell. Biochem. 2021, 476, 3869–3877. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, E.; Nakano, H.; Morimoto, M.; Tamaoki, T. Calphostin C (UCN-1028C), a novel microbial compound, is a highly potent and specific inhibitor of protein kinase C. Biochem. Biophys. Res. Commun. 1989, 159, 548–553. [Google Scholar] [CrossRef]
- Kobayashi, E.; Ando, K.; Nakano, H.; Iida, T.; Ohno, H.; Morimoto, M.; Tamaoki, T. Calphostins (UCN-1028), novel and specific inhibitors of protein kinase C. I. Fermentation, isolation, physico-chemical properties and biological activities. J. Antibiot. 1989, 42, 1470–1474. [Google Scholar] [CrossRef] [Green Version]
- Bruns, R.F.; Miller, F.D.; Merriman, R.L.; Howbert, J.J.; Heath, W.F.; Kobayashi, E.; Takahashi, I.; Tamaoki, T.; Nakano, H. Inhibition of protein kinase C by calphostin C is light-dependent. Biochem. Biophys. Res. Commun. 1991, 176, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Robitaille, K.; Daviau, A.; Lachance, G.; Couture, J.P.; Blouin, R. Calphostin C-induced apoptosis is mediated by a tissue transglutaminase-dependent mechanism involving the DLK/JNK signaling pathway. Cell Death Differ. 2008, 15, 1522–1531. [Google Scholar] [CrossRef] [Green Version]
- Kaul, A.; Maltese, W.A. Killing of cancer cells by the photoactivatable protein kinase C inhibitor, calphostin C, involves induction of endoplasmic reticulum stress. Neoplasia 2009, 11, 823–834. [Google Scholar] [CrossRef] [Green Version]
- Nishizuka, Y. Studies and perspectives of protein kinase C. Science 1986, 233, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Mackay, H.J.; Twelves, C.J. Targeting the protein kinase C family: Are we there yet? Nat. Rev. Cancer 2007, 7, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Merlic, C.A.; Aldrich, C.C.; Albaneze-Walker, J.; Saghatelian, A. Carbene complexes in the synthesis of complex natural products: Total synthesis of the Calphostins. J. Am. Chem. Soc. 2000, 122, 3224–3225. [Google Scholar] [CrossRef] [Green Version]
- Mulrooey, C.A.; O’Brien, E.M.; Morgan, B.J.; Kozlowski, M.C. Perylenequinones: Isolation, synthesis, and biological activity. Eur. J. Org. Chem. 2012, 21, 3887–3904. [Google Scholar] [CrossRef] [Green Version]
- Nakano, H.; Omura, S. Chemical biology of natural indolocarbazole products: 30 years since the discovery of staurosporine. J. Antibiot. 2009, 62, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Alonso, M.; Muñiz, M.; Hall, C.; Velasco, A.; Hidalgo, J. Calphostin C induces selective disassembly of the Golgi complex by a protein kinase C-independent mechanism. Eur. J. Cell Biol. 1998, 76, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Morgado-Díaz, J.A.; Montesano, G.; De Souza Fernandes, S.; Redondo, P.A.; Fernandes de Souza, W.; Albuquerque-Xavier, A.C.; Leve, F.; Tanaka, M.N.; Martins de Araujo, W.; Oliveira, S.S.; et al. Golgi complex disassembly caused by light-activated calphostin C involves MAPK and PKA. Tissue Cell 2007, 39, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Werner, G.; Hagenmaier, H.; Drautz, H.; Baumgartner, A.; Zähner, H. Metabolic products of microorganisms. 224. Bafilomycins, a new group of macrolide antibiotics. Production, isolation, chemical structure and biological activity. J. Antibiot. 1984, 37, 110–117. [Google Scholar] [CrossRef]
- Bowman, E.J.; Siebers, A.; Altendorf, K. Bafilomycins: A class of inhibitors of membrane ATPases from microorganisms, animal cells, and plant cells. Proc. Natl. Acad. Sci. USA 1988, 85, 7972–7976. [Google Scholar] [CrossRef] [Green Version]
- Manabe, T.; Yoshimori, T.; Henomatsu, N.; Tashiro, Y. Inhibitors of vacuolar-type H+-ATPase suppresses proliferation of cultured cells. J. Cell. Physiol. 1993, 157, 445–452. [Google Scholar] [CrossRef]
- Cotter, K.; Stransky, L.; McGuire, C.; Forgac, M. Recent insights into the structure, regulation, and function of the V-ATPases. Trends Biochem. Sci. 2015, 40, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Wang, J.; Hassan, A.; Lee, C.H.; Xie, X.S.; Li, X. Molecular basis of V-ATPase inhibition by bafilomycin A1. Nat. Commun. 2021, 12, 1782. [Google Scholar] [CrossRef] [PubMed]
- Gagliardi, S.; Gatti, P.A.; Belfiore, P.; Zocchetti, A.; Clarke, G.D.; Farina, C. Synthesis and structure-activity relationships of bafilomycin A1 derivatives as inhibitors of vacuolar H+-ATPase. J. Med. Chem. 1998, 41, 1883–1893. [Google Scholar] [CrossRef] [PubMed]
- Shibata, H.; Tsuchikawa, H.; Hayashi, T.; Matsumori, N.; Murata, M.; Usui, T. Modification of bafilomycin structure to efficiently synthesize solid-state NMR probes that selectively bind to vacuolar-type ATPase. Chem. Asian J. 2015, 10, 915–924. [Google Scholar] [CrossRef]
- Mauvezin, C.; Neufeld, T.P. Bafilomycin A1 disrupts autophagic flux by inhibiting both V-ATPase-dependent acidification and Ca-P60A/SERCA-dependent autophagosome-lysosome fusion. Autophagy 2015, 11, 1437–1438. [Google Scholar] [CrossRef] [Green Version]
- Palokangas, H.; Ying, M.; Väänänen, K.; Saraste, J. Retrograde transport from the pre-Golgi intermediate compartment and the Golgi complex is affected by the vacuolar H+-ATPase inhibitor bafilomycin A1. J. Mol. Cell Biol. 1998, 9, 3561–3578. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Jiang, K.; Liu, P.; Zhang, X.; Dong, X.; Gao, J.; Liu, Q.; Barr, M.P.; Zhang, Q.; Hou, X.; et al. Bafilomycin A1 induces caspase-independent cell death in hepatocellular carcinoma cells via targeting of autophagy and MAPK pathways. Sci. Rep. 2016, 6, 37052. [Google Scholar] [CrossRef]
- Axelsson, M.A.; Karlsson, N.G.; Steel, D.M.; Ouwendijk, J.; Nilsson, T.; Hansson, G.C. Neutralization of pH in the Golgi apparatus causes redistribution of glycosyltransferases and changes in the O-glycosylation of mucins. Glycobiology 2001, 11, 633–644. [Google Scholar] [CrossRef] [Green Version]
- Satoh, M.; Hirayoshi, K.; Yokota, S.; Hosokawa, N.; Nagata, K. Intracellular interaction of collagen-specific stress protein HSP47 with newly synthesized procollagen. J. Cell Biol. 1996, 133, 469–483. [Google Scholar] [CrossRef]
- Levitzki, A.; Gazit, A. Tyrosine kinase inhibition: An approach to drug development. Science 1995, 267, 1782–1788. [Google Scholar] [CrossRef]
- Cohen, P.; Cross, D.; Jänne, P.A. Kinase drug discovery 20 years after Imatinib: Progress and future directions. Nat. Rev. Drug Discov. 2021, 20, 551–569. [Google Scholar] [CrossRef] [PubMed]
- Levitzki, A. Protein kinase inhibitors as a therapeutic modality. Acc. Chem. Res. 2003, 36, 462–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, A.G.; Doherty, M.M.; Walker, F.; Weinstock, J.; Nerrie, M.; Vitali, A.; Murphy, R.; Johns, T.G.; Scott, A.M.; Levitzki, A.; et al. Preclinical analysis of the analinoquinazoline AG1478, a specific small molecule inhibitor of EGF receptor tyrosine kinase. Biochem. Pharmacol. 2006, 71, 1422–1434. [Google Scholar] [CrossRef] [PubMed]
- Caja, L.; Sancho, P.; Bertran, E.; Ortiz, C.; Campbell, J.S.; Fausto, N.; Fabregat, I. The tyrphostin AG1478 inhibits proliferation and induces death of liver tumor cells through EGF receptor-dependent and independent mechanisms. Biochem. Pharmacol. 2011, 82, 1583–1592. [Google Scholar] [CrossRef]
- Pan, H.; Yu, J.; Zhang, L.; Carpenter, A.; Zhu, H.; Li, L.; Ma, D.; Yuan, J. A novel small molecule regulator of guanine nucleotide exchange activity of the ADP-ribosylation factor and Golgi membrane trafficking. J. Biol. Chem. 2008, 283, 31087–31096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugawara, T.; Nakatsu, D.; Kii, H.; Maiya, N.; Adachi, A.; Yamamoto, A.; Kano, F.; Murata, M. PKCδ and ε regulate the morphological integrity of the ER-Golgi intermediate compartment (ERGIC) but not the anterograde and retrograde transports via the Golgi apparatus. Biochim. Biophys. Acta 2012, 1823, 861–875. [Google Scholar] [CrossRef] [Green Version]
- Boncompain, G.; Gareil, N.; Tessier, S.; Lescure, A.; Jones, T.R.; Kepp, O.; Kroemer, G.; Del Nery, E.; Perez, F. BML-265 and Tyrphostin AG1478 disperse the Golgi apparatus and abolish protein transport in human cells. Front. Cell Dev. Biol. 2019, 7, 232. [Google Scholar] [CrossRef] [Green Version]
- López, M.D.; Quiñoá, E.; Riguera, R. Dactyltronic acids from the sponge Dactylospongia elegans. J. Nat. Prod. 1994, 57, 992–996. [Google Scholar] [CrossRef]
- Takizawa, P.A.; Yucel, J.K.; Veit, B.; Faulkner, D.J.; Deerinck, T.; Soto, G.; Ellisman, M.; Malhotra, V. Complete vesiculation of Golgi membranes and inhibition of protein transport by a novel sea sponge metabolite, ilimaquinone. Cell 1993, 73, 1079–1090. [Google Scholar] [CrossRef]
- Sonoda, H.; Okada, T.; Jahangeer, S.; Nakamura, S. Requirement of phospholipase D for ilimaquinone-induced Golgi membrane fragmentation. J. Biol. Chem. 2007, 282, 34085–34092. [Google Scholar] [CrossRef] [Green Version]
- De Figueiredo, P.; Polizotto, R.S.; Drecktrah, D.; Brown, W.J. Membrane tubule-mediated reassembly and maintenance of the Golgi complex is disrupted by phospholipase A2 antagonists. Mol. Biol. Cell 1999, 10, 1763–1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radeke, H.S.; Digits, C.A.; Casaubon, R.L.; Snapper, M.L. Interactions of (-)-ilimaquinone with methylation enzymes: Implications for vesicular-mediated secretion. Chem. Biol. 1999, 6, 639–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, P.H.; Chueh, S.C.; Kung, F.L.; Pan, S.L.; Shen, Y.C.; Guh, J.H. Ilimaquinone, a marine sponge metabolite, displays anticancer activity via GADD153-mediated pathway. Eur. J. Pharmacol. 2007, 556, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Surti, M.; Patel, M.; Redhwan, A.; Al-Keridis, L.A.; Adnan, M.; Alshammari, N.; Reddy, M.N. Ilimaquinone (Marine Sponge Metabolite) induces apoptosis in HCT-116 human colorectal carcinoma cells via mitochondrial-mediated apoptosis pathway. Mar. Drugs 2022, 20, 582. [Google Scholar] [CrossRef]
- Mimaki, Y.; Kuroda, M.; Kameyama, A.; Sashida, Y.; Hirano, T.; Oka, K.; Maekawa, R.; Wada, T.; Sugita, K.; Buetler, J.A. Cholestane glycosides with potent cytostatic activities on various tumor cells from Ornithogalum saundersiae bulbs. Bioorg. Med. Chem. Lett. 1997, 7, 633–636. [Google Scholar] [CrossRef]
- Mimaki, Y. Structures and biological activities of plant glycosides: Cholestane glycosides from Ornithogalum saundersiae, O. thyrsoides and Galtonia candicans, and their cytotoxic and antitumor activities. Nat. Prod. Commun. 2006, 1, 247–253. [Google Scholar] [CrossRef]
- Tang, Y.; Li, N.; Duan, J.; Tao, W. Structure, bioactivity, and chemical synthesis of OSW-1 and other steroidal glycosides in the Genus Ornithogalum. Chem. Rev. 2013, 113, 5480–5514. [Google Scholar] [CrossRef]
- Zhou, Y.; Garcia-Prieto, C.; Carney, D.A.; Xu, R.H.; Pelicano, H.; Kang, Y.; Yu, W.; Lou, C.; Kondo, S.; Liu, J.; et al. OSW-1: A natural compound with potent anticancer activity and a novel mechanism of action. J. Natl. Cancer Inst. 2005, 97, 1781–1785. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, R.; Sakurai, K. Development of chemical probes for functional analysis of anticancer saponin OSW-1. Chem. Rec. 2019, 19, 2362–2369. [Google Scholar] [CrossRef]
- Zhan, Z.; Liu, Z.; Lai, J.; Zhang, C.; Chen, Y.; Huang, H. Anticancer effects and mechanisms of OSW-1 isolated from Ornithogalum saundersiae: A review. Front. Oncol. 2021, 11, 747718. [Google Scholar] [CrossRef]
- Burgett, A.W.; Poulsen, T.B.; Wangkanont, K.; Anderson, D.R.; Kikuchi, C.; Shimada, K.; Okubo, S.; Fortner, K.C.; Mimaki, Y.; Kuroda, M.; et al. Natural products reveal cancer cell dependence on oxysterol-binding proteins. Nat. Chem. Biol. 2011, 7, 639–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bensen, R.C.; Gunay, G.; Finneran, M.C.; Jhingan, I.; Acar, H.; Burgett, A.W.G. Small molecule targeting of oxysterol-binding protein (OSBP)-related protein 4 and OSBP inhibits ovarian cancer cell proliferation in monolayer and spheroid cell models. ACS Pharmacol. Transl. Sci. 2021, 4, 744–756. [Google Scholar] [CrossRef] [PubMed]
- Pietrangelo, A.; Ridgway, N.D. Bridging the molecular and biological functions of the oxysterol-binding protein family. Cell. Mol. Life Sci. 2018, 75, 3079–3098. [Google Scholar] [CrossRef] [PubMed]
- Olkkonen, V.M. The emerging roles of OSBP-related proteins in cancer: Impacts through phosphoinositide metabolism and protein-protein interactions. Biochem. Pharmacol. 2022, 196, 114455. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Uchida, Y.; Yachi, R.; Kudlyk, T.; Lupashin, V.; Inoue, T.; Taguchi, T.; Arai, H. Oxysterol-binding protein (OSBP) is required for the perinuclear localization of intra-Golgi v-SNAREs. Mol. Biol. Cell 2013, 24, 3534–3544. [Google Scholar] [CrossRef] [PubMed]
- Goto, A.; Charman, M.; Ridgway, N.D. Oxysterol-binding protein activation at endoplasmic reticulum-Golgi contact sites reorganizes phosphatidylinositol 4-phosphate pools. J. Biol. Chem. 2016, 291, 1336–1347. [Google Scholar] [CrossRef] [Green Version]
- Nakatsu, F.; Kawasaki, A. Functions of oxysterol-binding proteins at membrane contact sites and their control by phosphoinositide metabolism. Front. Cell Dev. Biol. 2021, 9, 664788. [Google Scholar] [CrossRef]
- Yamada, R.; Takeshita, T.; Hiraizumi, M.; Shinohe, D.; Ohta, Y.; Sakurai, K. Fluorescent analog of OSW-1 and its cellular localization. Bioorg. Med. Chem. Lett. 2014, 24, 1839–1842. [Google Scholar] [CrossRef]
- Pietrangelo, A.; Ridgway, N.D. Golgi-localization of oxysterol binding protein-related protein 4L (ORP4L) is regulated by ligand binding. J. Cell Biol. 2018, 131, jcs215335. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M.; Sasaki, K.; Fukutani, Y.; Yoshida, H.; Ohsawa, I.; Yohda, M.; Sakurai, K. Anticancer saponin OSW-1 is a novel class of selective Golgi stress inducer. Bioorg. Med. Chem. Lett. 2019, 29, 1732–1736. [Google Scholar] [CrossRef]
- Beutler, J.A.; Shoemaker, R.H.; Johnson, T.; Boyd, M.R. Cytotoxic geranyl stilbenes from Macaranga schweinfurthii. J. Nat. Prod. 1998, 61, 1509–5012. [Google Scholar] [CrossRef] [PubMed]
- Beutler, J.; Johnson, G.; Cragg, G.; Wiemer, D.; Neighbors, J.; Salnikova, M.; Hollingshead, M.; Scudiero, D.; McCloud, T. The Schweinfurthins: Issues in development of a plant-based anticancer lead. In Medicinal and Aromatic Plants: Agricultural, Commercial, Ecological, Legal, Pharmacological and Social Aspects, 1st ed.; Bogers, R.J., Craker, L.E., Lange, D., Eds.; Springer: Dordrecht, The Netherlands, 2006; Volume 17, pp. 301–309. [Google Scholar]
- Koubek, E.J.; Weissenrieder, J.S.; Neighbors, J.D.; Hohl, R.J. Schweinfurthins: Lipid modulators with promising anticancer activity. Lipids 2018, 53, 767–784. [Google Scholar] [CrossRef] [PubMed]
- Neighbors, J.D.; Beutler, J.A.; Wiemer, D.F. Synthesis of nonracemic 3-deoxyschweinfurthin B. J. Org. Chem. 2005, 70, 925–931. [Google Scholar] [CrossRef]
- Kuder, C.H.; Weivoda, M.M.; Zhang, Y.; Zhu, J.; Neighbors, J.D.; Wiemer, D.F.; Hohl, R.J. 3-Deoxyschweinfurthin B lowers cholesterol levels by decreasing synthesis and increasing export in cultured cancer cell lines. Lipids 2015, 50, 1195–1207. [Google Scholar] [CrossRef] [Green Version]
- Kuder, C.H.; Sheehy, R.M.; Neighbors, J.D.; Wiemer, D.F.; Hohl, R.J. Functional evaluation of a fluorescent schweinfurthin: Mechanism of cytotoxicity and intracellular quantification. Mol. Pharmacol. 2012, 82, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Péresse, T.; Kovacs, D.; Subra, M.; Bigay, J.; Tsai, M.C.; Polidori, J.; Gautier, R.; Desrat, S.; Fleuriot, L.; Debayle, D.; et al. Molecular and cellular dissection of the oxysterol-binding protein cycle through a fluorescent inhibitor. J. Biol. Chem. 2020, 295, 4277–4288. [Google Scholar] [CrossRef]
- Bao, X.; Zheng, W.; Hata Sugi, N.; Agarwala, K.L.; Xu, Q.; Wang, Z.; Tendyke, K.; Lee, W.; Parent, L.; Li, W.; et al. Small molecule schweinfurthins selectively inhibit cancer cell proliferation and mTOR/AKT signaling by interfering with trans-Golgi-network trafficking. Cancer Biol. Ther. 2015, 16, 589–601. [Google Scholar] [CrossRef] [Green Version]
- Paek, S.M. Recent synthesis and discovery of brefeldin A analogs. Mar. Drugs 2018, 16, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Bennink, J.R.; Kang, H.C.; Haugland, R.P.; Yewdell, J.W. Fluorescent conjugates of brefeldin A selectively stain the endoplasmic reticulum and Golgi complex of living cells. J. Histochem. Cytochem. 1995, 43, 907–915. [Google Scholar] [CrossRef] [Green Version]
- Lippincott-Schwartz, J.; Yuan, L.C.; Bonifacino, J.S.; Klausner, R.D. Rapid redistribution of Golgi proteins into the ER in cells treated with brefeldin A: Evidence for membrane cycling from Golgi to ER. Cell 1989, 56, 801–813. [Google Scholar] [CrossRef]
- Nylander, S.; Kalies, I. Brefeldin A, but not monensin, completely blocks CD69 expression on mouse lymphocytes: Efficacy of inhibitors of protein secretion in protocols for intracellular cytokine staining by flow cytometry. J. Immunol. Methods 1999, 224, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Reaves, B.; Banting, G. Perturbation of the morphology of the trans-Golgi network following Brefeldin A treatment: Redistribution of a TGN-specific integral membrane protein, TGN38. J. Cell Biol. 1992, 116, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Gao, W.; Wang, K.; Huang, Z.; Zhang, L.; Zhang, Z.; Zhou, J.; Nice, E.C.; Huang, C. Brefeldin A inhibits colorectal cancer growth by triggering Bip/Akt-regulated autophagy. FASEB J. 2019, 33, 5520–5534. [Google Scholar] [CrossRef]
- Zhu, J.W.; Nagasawa, H.; Nagura, F.; Mohamad, S.B.; Uto, Y.; Ohkura, K.; Hori, H. Elucidation of strict structural requirements of brefeldin A as an inducer of differentiation and apoptosis. Bioorg. Med. Chem. 2000, 8, 455–463. [Google Scholar] [CrossRef]
- Ohashi, Y.; Iijima, H.; Yamaotsu, N.; Yamazaki, K.; Sato, S.; Okamura, M.; Sugimoto, K.; Dan, S.; Hirono, S.; Yamori, T. AMF-26, a novel inhibitor of the Golgi system, targeting ADP-ribosylation factor 1 (Arf1) with potential for cancer therapy. J. Biol. Chem. 2012, 287, 3885–3897. [Google Scholar] [CrossRef] [Green Version]
- Ohashi, Y.; Okamura, M.; Hirosawa, A.; Tamaki, N.; Akatsuka, A.; Wu, K.M.; Choi, H.W.; Yoshimatsu, K.; Shiina, I.; Yamori, T.; et al. M-COPA, a Golgi disruptor, inhibits cell surface expression of MET protein and exhibits antitumor activity against MET-addicted gastric cancers. Cancer Res. 2016, 76, 3895–3903. [Google Scholar] [CrossRef] [Green Version]
- Hara, Y.; Obata, Y.; Horikawa, K.; Tasaki, Y.; Suzuki, K.; Murata, T.; Shiina, I.; Abe, R. M-COPA suppresses endolysosomal Kit-Akt oncogenic signalling through inhibiting the secretory pathway in neoplastic mast cells. PLoS ONE 2017, 12, e0175514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obata, Y.; Horikawa, K.; Shiina, I.; Takahashi, T.; Murata, T.; Tasaki, Y.; Suzuki, K.; Yonekura, K.; Esumi, H.; Nishida, T.; et al. Oncogenic kit signalling on the Golgi is suppressed by blocking secretory trafficking with M-COPA in gastrointestinal stromal tumours. Cancer Lett. 2018, 415, 1–10. [Google Scholar] [CrossRef]
- Watari, K.; Nakamura, M.; Fukunaga, Y.; Furuno, A.; Shibata, T.; Kawahara, A.; Hosoi, F.; Kuwano, T.; Kuwano, M.; Ono, M. The antitumor effect of a novel angiogenesis inhibitor (an octahydronaphthalene derivative) targeting both VEGF receptor and NF-κB pathway. Int. J. Cancer 2012, 131, 310–321. [Google Scholar] [CrossRef]
- Ohashi, Y.; Okamura, M.; Katayama, R.; Fang, S.; Tsutsui, S.; Akatsuka, A.; Shan, M.; Choi, H.W.; Fujita, N.; Yoshimatsu, K.; et al. Targeting the Golgi apparatus to overcome acquired resistance of non-small cell lung cancer cells to EGFR tyrosine kinase inhibitors. Oncotarget 2017, 9, 1641–1655. [Google Scholar] [CrossRef] [Green Version]
- Haney, M.E., Jr.; Hoehn, M.M. Monensin, a new biologically active compound. I. Discovery and isolation. Antimicrob. Agents Chemother. 1967, 7, 349–352. [Google Scholar] [PubMed]
- Aowicki, D.; Huczyński, A. Structure and antimicrobial properties of monensin A and its derivatives: Summary of the achievements. Biomed. Res. Int. 2013, 2013, 742149. [Google Scholar]
- Yang, K.; Wang, X.; Zhang, H.; Wang, Z.; Nan, G.; Li, Y.; Zhang, F.; Mohammed, M.K.; Haydon, R.C.; Luu, H.H.; et al. The evolving roles of canonical WNT signaling in stem cells and tumorigenesis: Implications in targeted cancer therapies. Lab. Investig. 2016, 96, 116–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wu, X.; Zhang, Z.; Ma, C.; Wu, T.; Tang, S.; Zeng, Z.; Huang, S.; Gong, C.; Yuan, C.; et al. Monensin inhibits cell proliferation and tumor growth of chemo-resistant pancreatic cancer cells by targeting the EGFR signaling pathway. Sci. Rep. 2018, 8, 17914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Kim, K.Y.; Yu, S.N.; Park, S.G.; Yu, H.S.; Seo, Y.K.; Ahn, S.C. Monensin induces PC-3 prostate cancer cell apoptosis via ROS production and Ca2+ homeostasis disruption. Anticancer Res. 2016, 36, 5835–5843. [Google Scholar] [CrossRef] [Green Version]
- Rebelo, R.; Polónia, B.; Santos, L.L.; Vasconcelos, M.H.; Xavier, C.P.R. Drug repurposing opportunities in pancreatic ductal adenocarcinoma. Pharmaceuticals 2021, 14, 280. [Google Scholar] [CrossRef]
- Harned, R.L.; Hidy, P.H.; Corum, C.J.; Jones, K.L. Nigericin a new crystalline antibiotic from an unidentified Streptomyces. Antibiot. Chemother. 1951, 1, 594–596. [Google Scholar]
- Podinovskaia, M.; Prescianotto-Baschong, C.; Buser, D.P.; Spang, A.A. Novel live-cell imaging assay reveals regulation of endosome maturation. eLife 2021, 10, e70982. [Google Scholar] [CrossRef]
- Liu, F.; Li, W.; Hua, S.; Han, Y.; Xu, Z.; Wan, D.; Wang, Y.; Chen, W.; Kuang, Y.; Shi, J.; et al. Nigericin exerts anticancer effects on human colorectal cancer cells by inhibiting Wnt/β-catenin signaling pathway. Mol. Cancer Ther. 2018, 17, 952–965. [Google Scholar] [CrossRef] [Green Version]
- Perregaux, D.; Gabel, C.A. Interleukin-1 beta maturation and release in response to ATP and nigericin. Evidence that potassium depletion mediated by these agents is a necessary and common feature of their activity. J. Biol. Chem. 1994, 269, 15195–15203. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khine, M.N.; Sakurai, K. Golgi-Targeting Anticancer Natural Products. Cancers 2023, 15, 2086. https://doi.org/10.3390/cancers15072086
Khine MN, Sakurai K. Golgi-Targeting Anticancer Natural Products. Cancers. 2023; 15(7):2086. https://doi.org/10.3390/cancers15072086
Chicago/Turabian StyleKhine, Myat Nyein, and Kaori Sakurai. 2023. "Golgi-Targeting Anticancer Natural Products" Cancers 15, no. 7: 2086. https://doi.org/10.3390/cancers15072086
APA StyleKhine, M. N., & Sakurai, K. (2023). Golgi-Targeting Anticancer Natural Products. Cancers, 15(7), 2086. https://doi.org/10.3390/cancers15072086